
Synthesis of Theaflavins and Their Functions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
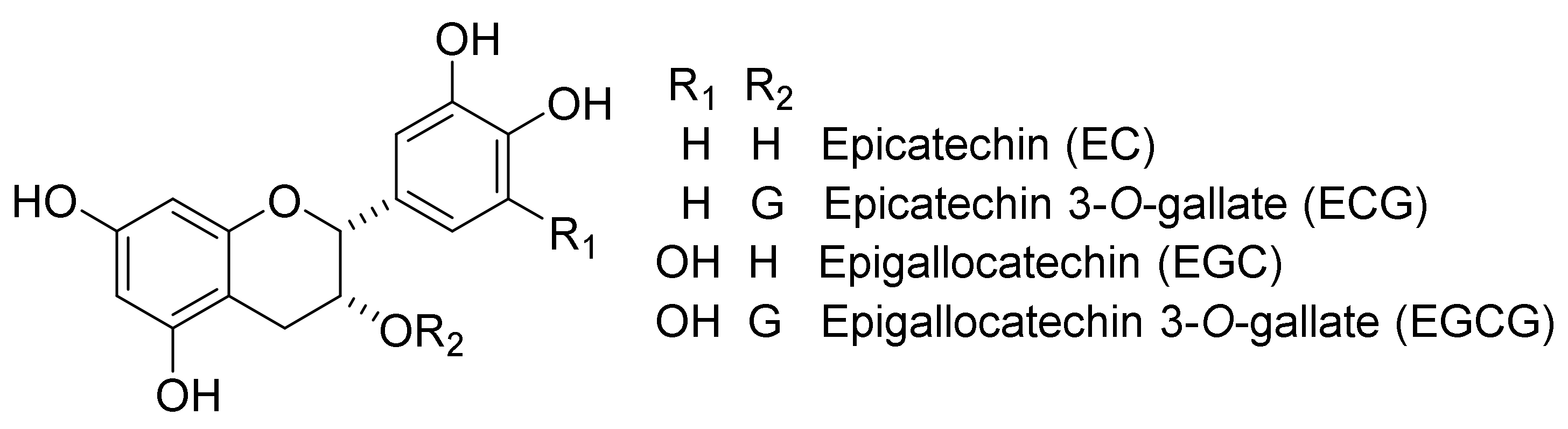
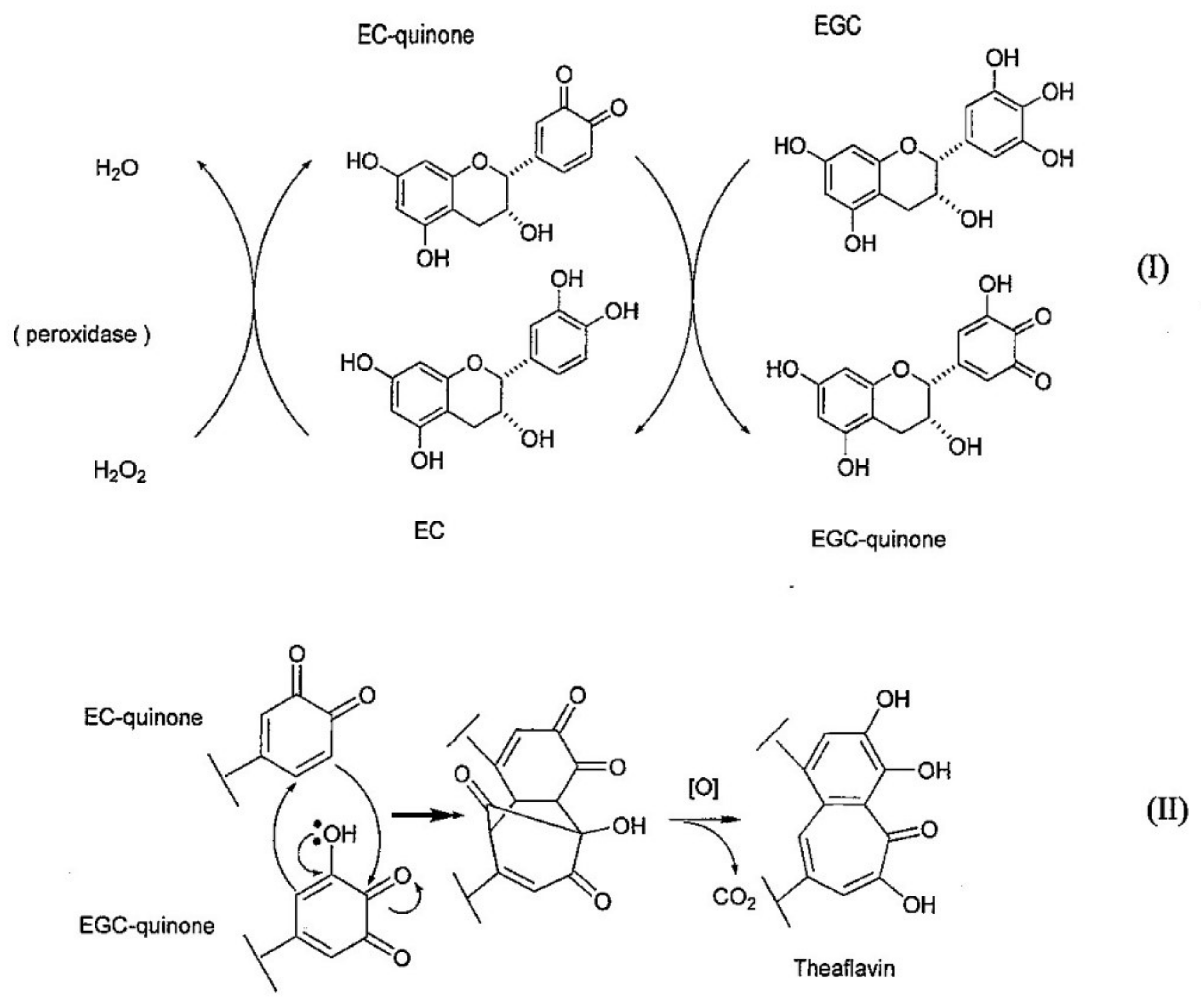
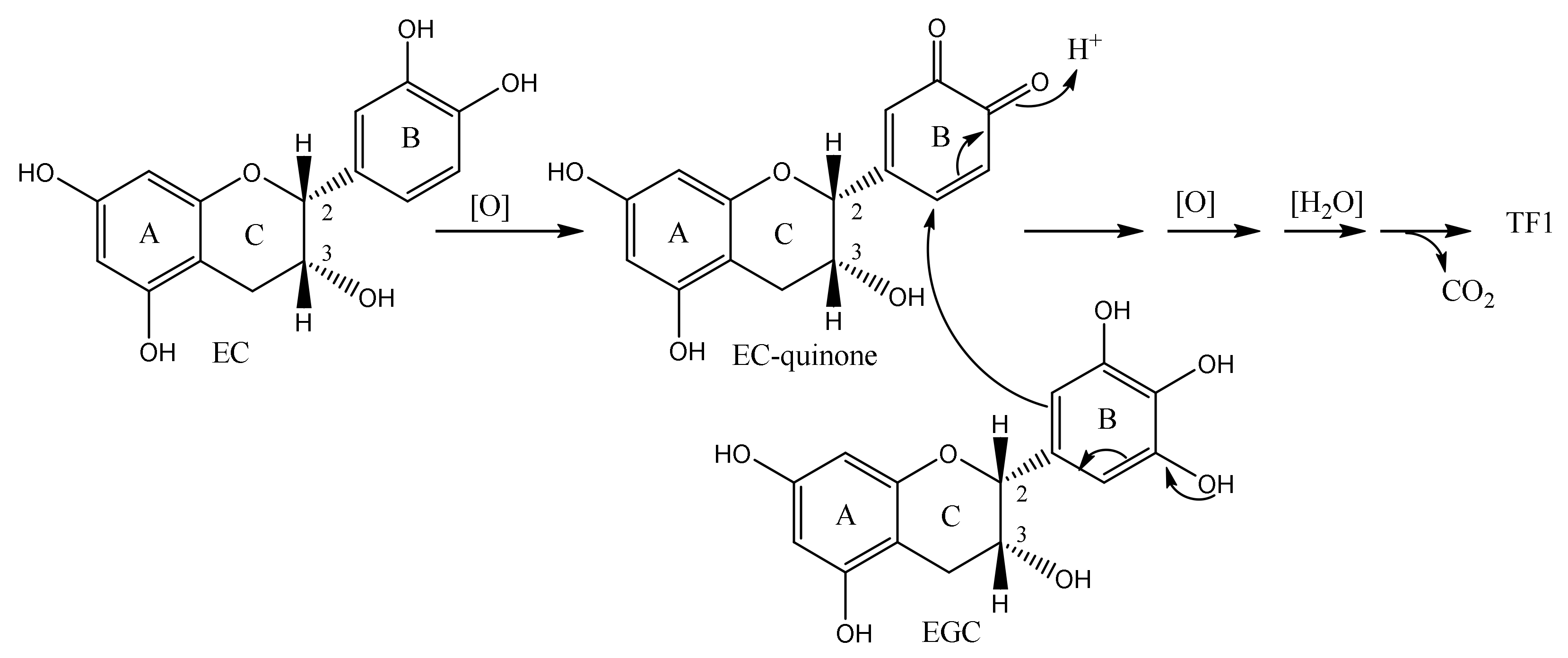
2. Biosynthetic Pathway of TFs
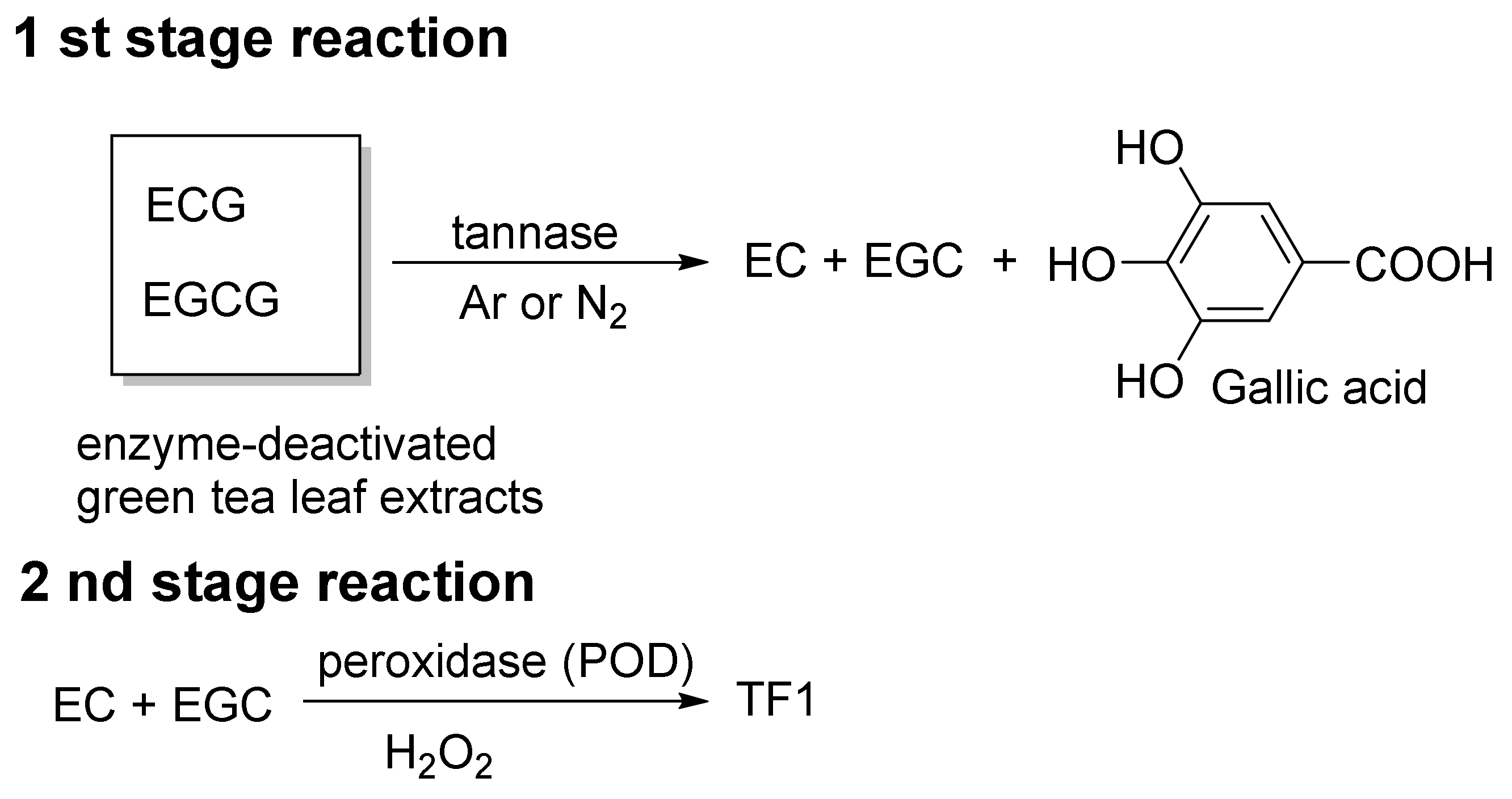
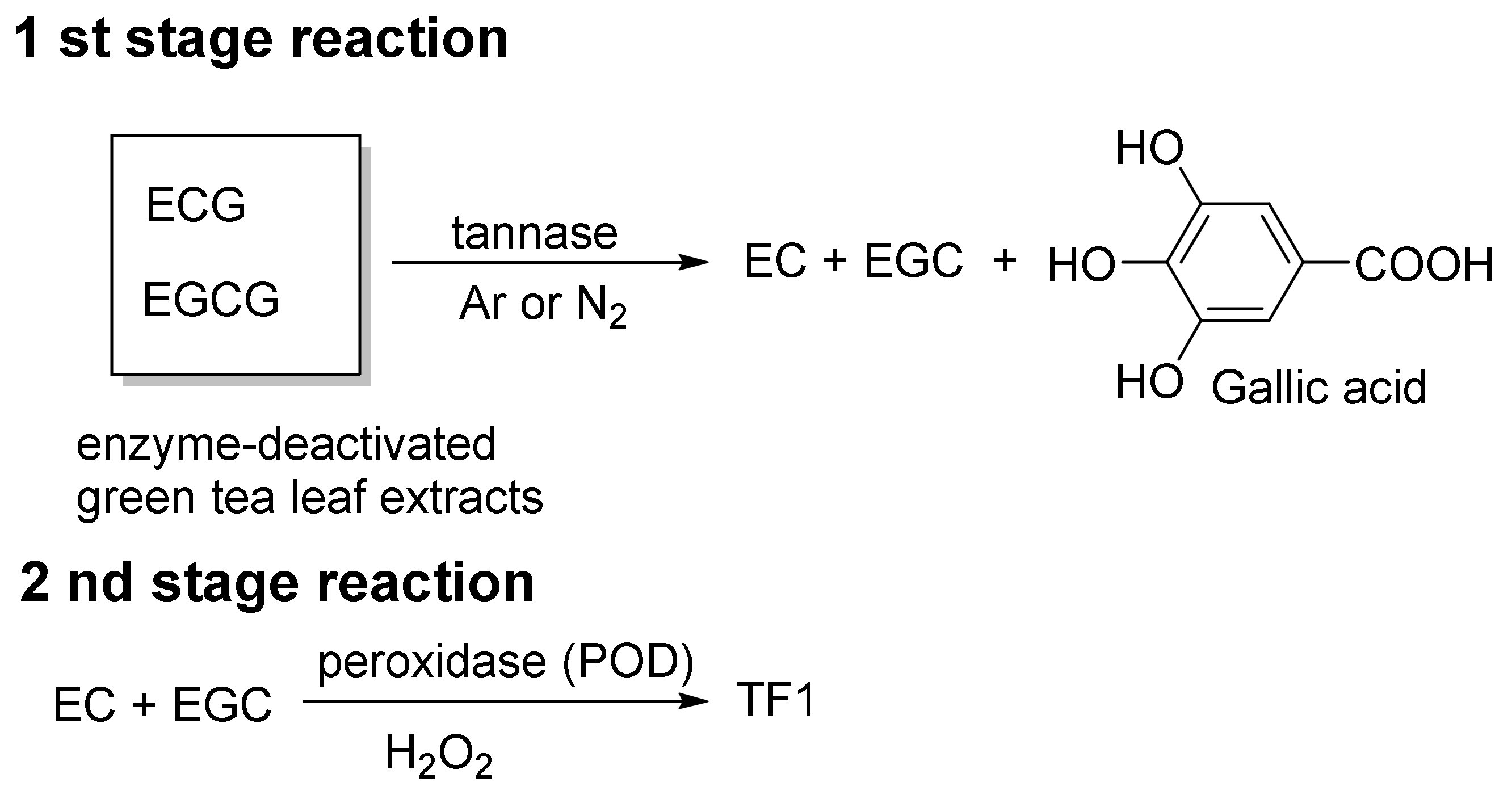
3. Methods for Obtaining TFs and Relevant Obstacles
4. Conventional Synthetic Methods of TFs
- ①
- EC + EGC → TF1
- ②
- EC + EGCG → TF2a
- ③
- ECG + EGC → TF2b
- ④
- ECG + EGCG → TF3
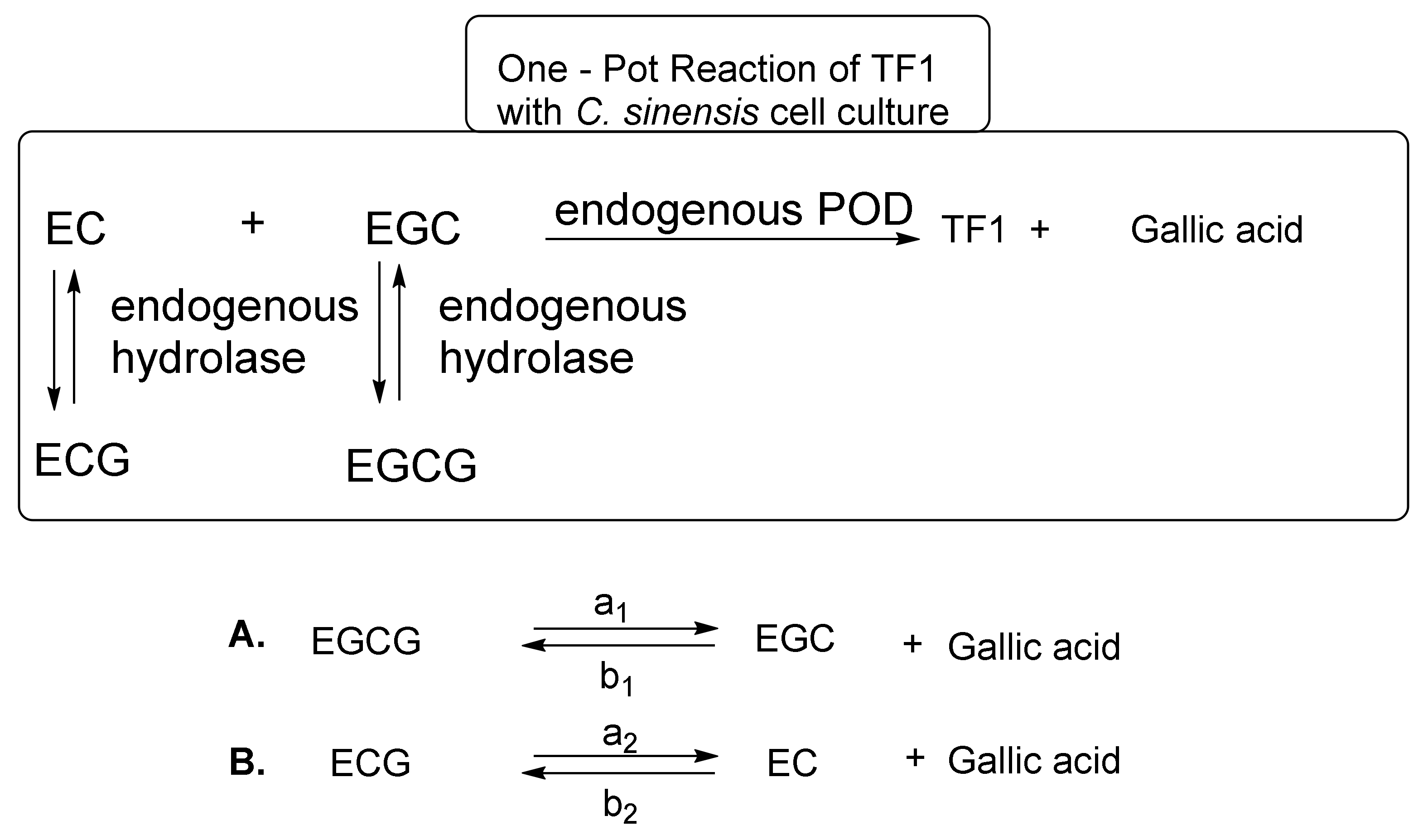
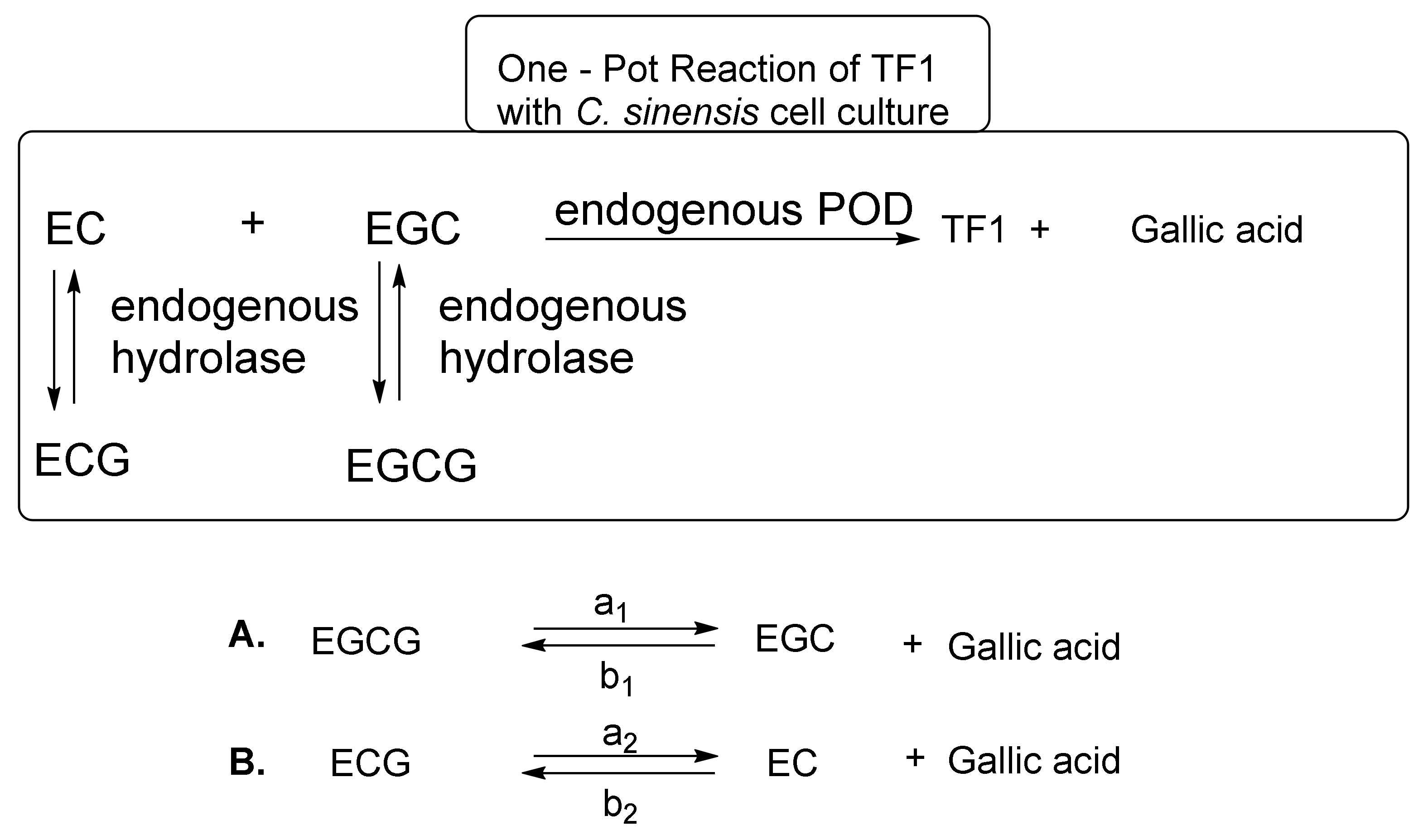
5. Biosynthetic Methods for the Mass Production of Theaflavins
6. Manufacturing Methods of Foods Containing TFs
7. Manufacturing Methods for Grain Flour Processed Foods Containing TFs
8. Health Benefits of TFs
8.1. Antimetabolic Syndrome
8.2. Anti-Periodontitis
8.3. Anti-Norovirus
8.4. Anti-Osteoporosis
9. Conclusions
Acknowledgments
Conflicts of Interest
References
- Tanaka, T.; Mine, C.; Inoue, K.; Matsuda, M.; Kouno, I. Synthesis of theaflavin from epicatechin and epigallocatechin by plant homogenates and role of epicatechin quinone in the synthesis and degradation of theaflavin. J. Agric. Food Chem. 2002, 50, 2142–2148. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.; Yang, C.S.; Ho, C.-T. Peroxidase-mediated oxidation of catechins. Phytochem. Rev. 2004, 3, 229–241. [Google Scholar] [CrossRef]
- Drynan, J.W.; Clifford, M.N.; Obuchowicz, J.; Kuhnert, N. The chemistry of low molecular weight black tea polyphenols. Nat. Prod. Rep. 2010, 27, 417–462. [Google Scholar] [CrossRef] [PubMed]
- He, H.-F. Research progress on theaflavins: Efficacy, formation, and preparation. Food Nutr. Res. 2017, 61, 1344521. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Rankin, G.O.; Tu, Y.; Chen, Y.C. Inhibitory Effects of the Four Main Theaflavin Derivatives Found in Black Tea on Ovarian Cancer Cells. Anticancer Res. 2016, 36, 643–651. [Google Scholar] [PubMed]
- Stensvold, I.; Tverdal, A.; Solvoll, K.; Foss, O.P. Tea consumption relationship to cholesterol, blood pressure, and coronary and total mortality. Prev. Med. 1992, 21, 546–553. [Google Scholar] [CrossRef]
- Green, M.S.; Harari, G. Association of serum lipoproteins and health- related habits with coffee and tea consumption in free-living subjects examined in the Israeli CORDIS study. Prev. Med. 1992, 21, 532–545. [Google Scholar] [CrossRef]
- Iwata, K.; Inayama, T.; Miwa, S. Effects of Chinese Green Tea, Oolong Tea and Black Tea on Plasma and Liver Lipid Metabolism in Fructose-induced Hyperlipidemia Rats. Jpn. J. Nutr. Diet. 1988, 46, 289–298. [Google Scholar] [CrossRef]
- Satoh, T.; Igarashi, M.; Yamada, S.; Takahashi, N.; Watanabe, K. Inhibitory effect of black tea and its combination with acarbose on small intestinal α-glucosidase activity. J. Ethnopharmacol. 2015, 161, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Ichitani, M.; Suzuki, Y.; Unno, T.; Sugawara, T.; Yamahira, T.; Kato, M.; Takihara, T.; Sagesaka, Y.; Kakuda, T.; et al. Black-tea polyphenols suppress postprandial hypertriacylglycerolemia by suppressing lymphatic transport of dietary fat in rats. J. Agric. Food Chem. 2009, 57, 7131–7136. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Pan, S.; Miao, A.; Ling, C.; Pang, S.; Tang, J.; Chen, D.; Zhao, C. Active extracts of black tea (Camellia Sinensis) induce apoptosis of PC-3 prostate cancer cells via mitochondrial dysfunction. Oncol. Rep. 2013, 30, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Peluso, I.; Serafini, M. Antioxidants from black and green tea: From dietary modulation of oxidative stress to pharmacological mechanisms. Br. J. Pharmacol. 2017, 174, 1195–1208. [Google Scholar] [CrossRef] [PubMed]
- Sur, S.; Panda, C.K. Molecular aspects of cancer chemopreventive and therapeutic efficacies of tea and tea polyphenols. Nutrition 2017, 43, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Loke, W.M.; Proudfoot, J.M.; Hodgson, J.M.; McKinley, A.J.; Hime, N.; Magat, M.; Stocker, R.; Croft, K.D. Specific dietary polyphenols attenuate atherosclerosis in apolipoprotein E-knockout mice by alleviating inflammation and endothelial dysfunction. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Jin, F.; Wang, Y.; Li, F.; Wang, L.; Wang, Q.; Ren, Z.; Wang, Y. In vitro and in vivo anti-inflammatory effects of theaflavin-3,3′-digallate on lipopolysaccharide-induced inflammation. Eur. J. Pharmacol. 2017, 794, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Ohba, M.; Oka, T.; Ando, T.; Arahata, S.; Ikegaya, A.; Takagi, H.; Ogo, N.; Zhu, C.; Owada, K.; Kawamori, F.; et al. Antiviral effect of theaflavins against caliciviruses. J. Antibiot. (Tokyo) 2017, 70, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.F.; Bai, L.P.; Huang, W.B.; Li, X.Z.; Zhao, S.S.; Zhong, N.S.; Jiang, Z.H. Comparison of in vitro antiviral activity of tea polyphenols against influenza A and B viruses and structure-activity relationship analysis. Fitoterapia 2014, 93, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Zu, M.; Yang, F.; Zhou, W.; Liu, A.; Du, G.; Zheng, L. In vitro anti-influenza virus and anti-inflammatory activities of theaflavin derivatives. Antivir. Res. 2012, 94, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, Y.; Hosokawa, I.; Ozaki, K.; Nakanishi, T.; Nakae, H.; Matsuo, T. Tea polyphenols inhibit IL-6 production in tumor necrosis factor superfamily 14-stimulated human gingival fibroblasts. Mol. Nutr. Food Res. 2010, 54, S2. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Qi, X.; Huang, S.; Chen, S.; Wu, Y.; Zhao, L. Theaflavins inhibit pathogenic properties of P. gingivalis and MMPs production in P. gingivalis-stimulated human gingival fibroblasts. Arch. Oral. Biol. 2015, 60, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Oka, Y.; Iwai, S.; Amano, H.; Irie, Y.; Yatomi, K.; Ryu, K.; Yamada, S.; Inagaki, K.; Oguchi, K. Tea polyphenols inhibit rat osteoclast formation and differentiation. J. Pharmacol. Sci. 2012, 118, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, K.; Iwamoto, Y.; Kobayashi, Y.; Katsuoka, F.; Kawaguchi, S.; Tsujita, T.; Nakamura, T.; Kato, S.; Yamamoto, M.; Takayanagi, H.; et al. DNA methyltransferase 3a regulates osteoclast differentiation by coupling to an S-adenosylmethionine-producing metabolic pathway. Nat. Med. 2015, 21, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Tanaka, T.; Tamura, S.; Toshima, A.; Tamaya, K.; Miyata, Y.; Tanaka, K.; Matsumoto, K. Alpha-Glucosidase inhibitory profile of catechins and theaflavins. J. Agric. Food Chem. 2007, 55, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Miyata, Y.; Tamaru, S.; Tanaka, T.; Tamaya, K.; Matsui, T.; Nagata, Y.; Tanaka, K. Theflavins and theasinensin A derived from fermented tea have antihyperglycemic and hypotriacylglycerolemic effects in KK-A(y) mice and Sprague-Dawley rats. J. Agric. Food Chem. 2013, 61, 9366–9372. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, M.; Takemoto, H.; Sakurada, H. Synthesis of theaflavins with Camellia sinensis cell culture and inhibition of increase in blood sugar values in high-fat diet mice subjected to sucrose or glucose loading. Tetrahedron Lett. 2014, 55, 5038–5040. [Google Scholar] [CrossRef]
- Takemoto, M.; Takemoto, H.; Kouno, H.; Soeda, T.; Moriya, T.A. Simple, enzymatic biotransformation method using fresh green tea leaves efficiently generates theaflavin-containing fermentation water that has potent physiological functions in mice and humans. Biol. Pharm. Bull. 2017, 40, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Kudo, N.; Arai, Y.; Suhara, Y.; Ishii, T.; Nakayama, T.; Osakabe, N. A single oral Administration of Theaflavins Increases Energy Expenditure and the Expression of Metabolic Genes. PLoS ONE 2015, 10, e0137809. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, M.; Takemoto, H.; Saijo, R. Theaflavin synthesized in a selective, domino-type, one-pot enzymatic biotransformation method with Camellia sinensis cell culture inhibits weight gain and fat accumulation to high-fat diet-induced obese mice. Biol. Pharm. Bull. 2016, 39, 1347–1352. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, M. Process for Selective Production of Theaflavin. Japanese Patent No 5260516, 2 May 2013. [Google Scholar]
- Takemoto, M. Manufacturing Method for Theaflavins Using Raw Tea Leaves. Japanese Patent No 4817206, 9 September 2011. [Google Scholar]
- Takemoto, M. Method of Producing Fermented Tea Drink Rich in Theaflavins. UK Patent GB2471250, 2013.1.30. Japanese Patent No 5472092, 14 February 2014. [Google Scholar]
- Takemoto, M. Method of Producing Fermented Tea Drink. Japanese Patent No 5472093, 14 February 2014. [Google Scholar]
- Takemoto, M. Fermented Tea Drink Containg Methylated Catechin. Japanese Patent No 5419296, 29 November 2013. [Google Scholar]
- Takemoto, M. Cereal Flour Food and Method for Producing Flour Food. Japanese Patent No 5435585, 20 December 2014. [Google Scholar]
- Takemoto, M. Prevention and Amelioration Agent of Lifestyle-Related Disease. Japanese Patent No 5614674, 19 September 2014. [Google Scholar]
- William, G.C.; David, P.A.; Richard, S.; Ambalavanar, T. Improvements in or Relating to Producting Theaflavin. Japan Patent No 4364954, 28 August 2009. [Google Scholar]
- Sinija, V.R.; Mishra, H.N.; Bal, S. Process technology for production of soluble tea powder. J. Food Eng. 2007, 82, 276–283. [Google Scholar] [CrossRef]
- Takino, Y.; Imagawa, H.; Horikawa, H.; Tanaka, A. Studies on the mechanism of the oxidation of tea leaf catechins. Agric. Biol. Chem. 1964, 28, 64–71. [Google Scholar]
- Yanase, E.; Sawaki, K.; Nakatsuka, S. The isolation of a bicyclo[3.2.1] intermediate during formation of benzotropolones, a common nucleus found in black tea pigments. Theaflavins. Synlett 2005, 17, 2661–2663. [Google Scholar]
- Takino, Y.; Imagawa, H. Crystalline reddish orange pigment of manufactured black tea. Agric. Biol. Chem. 1964, 28, 255–256. [Google Scholar]
- Collier, P.D.; Bryce, T.; Mallows, R.; Thomas, P.E. The theaflavins of black tea. Tetrahedron 1973, 29, 125–142. [Google Scholar] [CrossRef]
- Robertson, A. Effects of catechin concentration of the formation of black tea polyphenols during in vitro oxidation. Phytochemistry 1983, 22, 897–903. [Google Scholar] [CrossRef]
- Robertson, A. Effects of physical and chemical conditions on the in vitro oxidation of tea leaf catechins. Phytochemistry 1983, 22, 889–896. [Google Scholar] [CrossRef]
- Margaret, A.D.; Charles, J.F.; David, J.M.; Derwent, S. Fermentation of tea in aqueous suspension. Influence of tea peroxidase. J. Sci. Food Agric. 1981, 32, 920–932. [Google Scholar]
- Subramanian, N.; Venkatesh, P.; Ganguli, S.; Sinkar, V.P. Role of polyphenol oxidase and peroxidase in the generarion of black tea theaflavins. J. Agric. Food Chem. 1999, 47, 2571–2578. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.M.; Tian, S.Y.; Meng, X.F.; Stark, R.E.; Rosen, R.T.; Yang, C.S.; Ho, C.T. Theadibenzotropolone A, a new type pigment from enzymatic oxidation of (−)-epicatechin and (−)-epigallocatechin gallate and characterized from black tea using LC/MS/MS. Tetrahedron Lett. 2002, 43, 7129. [Google Scholar] [CrossRef]
- Sang, S.; Lambert, J.D.; Tian, S.Y.; Hong, J.; Hou, Z.; Ryu, J.H.; Stark, R.E.; Rosen, R.T.; Huang, M.T.; Yang, C.S.; et al. Enzymatic synthesis of tea theaflavin derivatives and their anti-inflammatory and cytotoxia activities. Bioorg. Med. Chem. 2004, 12, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, M. Method for Synthesizing Theaflavin. Japnese Patent No 4696302, 11 March 2011. [Google Scholar]
- Takemoto, M.; Aoshima, Y.; Stoynov, N.; Kutney, J.P. Establishment of Camellia sinensis cell culture with high peroxidase activity and oxidative coupling reaction of dibenzylbutanolides. Tetrahedron Lett. 2002, 43, 6915–6917. [Google Scholar] [CrossRef]
- Kawabe, Y.; Aihara, Y.; Hirose, Y.; Sakurada, A.; Yoshida, A.; Inai, M.; Asakawa, T.; Hamashima, Y.; Kan, T. Synthesis of theaflavins via biomimetic oxidative coupling reactions. Synlett 2013, 24, 479–482. [Google Scholar]
- Asakawa, T.; Kawabe, Y.; Yoshida, A.; Aihara, Y.; Manabe, T.; Hirose, Y.; Sakurada, A.; Inai, M.; Hamashima, Y.; Furuta, T.; et al. Syntheses of methylated catechin and theaflavins using 2-nitrobenzenesulfonyl group to protect and deactive phenol. J. Antibiot. 2016, 69, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Oowatashi, R.; Saito, Y.; Tanaka, T. Nonenzymatic biomimetic synthesis of black tea pigment theaflavins. Synlett 2017, 28, 2505–2508. [Google Scholar] [CrossRef]
- Maeda-Yamamoto, M.; Sano, M.; Matsuda, N.; Miyase, T.; Kawamoto, K.; Suzuki, N.; Yoshimura, M.; Tachibana, H.; Hakamata, K. The change of epigallocatechin-3-O-(3-O-methyl) gallate content in tea of different varieties, tea seasons of crop and processing method. Nippon Shokuhin Kagaku Kogaku Kaishi 2001, 48, 64–68. [Google Scholar] [CrossRef]
- Muramatsu, K.; Fukuyo, M.; Hara, Y. Effect of green tea catechins on plasma cholesterol level in cholesterol-fed rats. J. Nutr. Sci. Vitaminol. (Tokyo) 1986, 32, 613–622. [Google Scholar]
- Ikeda, I.; Imasato, Y.; Sasaki, E.; Nakayama, M.; Nagao, H.; Takeo, T.; Yayabe, F.; Sugano, M. Tea catechins decrease micellar solubility and intestinal absorption of cholesterol in rats. Biochim. Biophys. Acta 1992, 1127, 141–146. [Google Scholar] [CrossRef]
- Hartley, L.; Flowers, N.; Holmes, J.; Clarke, A.; Stranges, S.; Hooper, L.; Rees, K. Green and black tea for the primary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2013, 6, CD009934. [Google Scholar] [CrossRef] [PubMed]
- Santesso, N.; Manheimer, E. A summary of a cochrane review: Green and black tea for the primary prevention of cardiovascular disease. Glob. Adv. Health Med. 2014, 3, 66–67. [Google Scholar] [CrossRef] [PubMed]
- Vernarelli, J.A.; Lambert, J.D. Tea consumption is inversely associated with weight status and other markers for metabolic syndrome in US adults. Eur. J. Nutr. 2013, 52, 1039–1048. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, K.; Usui, Y.; Kan, T.; Ishii, T.; Nakayama, T. Structural specificity of electric potentials in the coulometric-array analysis of catechins and theaflavins. J. Clin. Biochem. Nutr. 2014, 55, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, H. Pharmacology of R-glucosidase inhibition. Eur. J. Clin. Investig. 1994, 24, 3–10. [Google Scholar]
- Hauri, H.P.; Wacker, H.; Rickli, E.E.; Meier, B.B.; Quaroni, A.; Semenza, G. Biosynthesis of sucrase-isomaltase. J. Biol. Chem. 1982, 257, 4522–4528. [Google Scholar] [PubMed]
- Odegaard, A.O.; Pereira, M.A.; Koh, W.P.; Arakawa, K.; Lee, H.P.; Yu, M.C. Coffee, tea, and incident type 2 diabetes: The Singapore Chinese Health Study. Am. J. Clin. Nutr. 2008, 88, 979–985. [Google Scholar] [CrossRef] [PubMed]
- Slots, J. Periodontology: Past, present, perspectives. Periodontol. 2000 2013, 62, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Fentoglu, O.; Koroglu, B.K.; Hicyilmaz, H.; Sert, T.; Ozdem, M.; Sutcu, R.; Tamer, M.N.; Orhan, H.; Ay, Z.Y.; Ozturk, T.M.; et al. Pro-inflammatory cytokine levels in association between periodontal disease and hyperlipidaemia. J. Clin. Periodontol. 2011, 38, 8–16. [Google Scholar]
- Noh, M.K.; Jung, M.; Kim, S.H.; Lee, S.R.; Park, K.H.; Kim, D.H.; Kim, H.H.; Park, Y.G. Assessment of IL-6, IL-8 and TNF-α levels in the gingival tissue of patients with periodontitis. Exp. Ther. Med. 2013, 6, 847–851. [Google Scholar] [CrossRef] [PubMed]
- Maeno, M.; Tanaka, H.; Zhang, F.; Kitami, S.; Nakai, K.; Kawato, T. Direct and indirect effects of IL-17 A on RANKL-induced osteoclastogenesis. J. Hard. Tissue Biol. 2013, 22, 287–292. [Google Scholar] [CrossRef]
- Zhou, J.; Windsor, L.J. Porphyromonas gingivalis affects host collagen degradation by affecting expression, activation, and inhibition of matrix metalloproteinases. J. Periodontal Res. 2006, 41, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, K.; Imamura, T.; Tomikawa, M.; Tancharoen, S.; Tatsuyama, S.; Maruyama, I. DX-9065a inhibits proinflammatory events induced by gingipains and factor XA. J. Periodontal Res. 2006, 41, 148–156. [Google Scholar] [PubMed]
- Zhao, L.; La, V.D.; Grenier, D. Antibacterial, antiadherence, antiprotease, and anti-inflammatory properties of various tea extracts: potential benefits for periodontal diseases. J. Med. Food 2013, 16, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Robilotti, E.; Deresinski, S.; Pinsky, B.A. Norovirus. Clin. Microbiol. Rev. 2015, 28, 134–164. [Google Scholar] [CrossRef] [PubMed]
- Oka, T.; Wang, Q.; Katayama, K.; Saif, L.J. Comprehensive review of human sapoviruses. Clin. Microbiol. Rev. 2015, 28, 32–53. [Google Scholar] [CrossRef] [PubMed]
- Cummings, S.R.; Kelsey, J.L.; Nevitt, M.C.; O’Dowd, K.J. Epidemiology of osteoporosis and osteoporotic fractures. Epidemiol. Rev. 1985, 7, 178–208. [Google Scholar] [CrossRef] [PubMed]
- Tezuka, K.; Nemoto, K.; Tezuka, Y.; Sato, T.; Ikeda, Y.; Kobori, M.; Kawashima, H.; Eguchi, H.; Hakeda, Y.; Kumegawa, M. Identification of matrix metalloproteinase 9 in rabbit osteoclasts. J. Biol. Chem. 1994, 269, 15006–15009. [Google Scholar] [PubMed]
- Logar, D.B.; Komadina, R.; Prezelj, J.; Ostanek, B.; Trost, Z.; Marc, J. Expression of bone resorption genes in osteoarthritis and in osteoporosis. J. Bone Miner. Metab. 2007, 25, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Delaissé, J.M.; Engsig, M.T.; Everts, V.; Ovejero, M.C.; Ferreras, M.; Lund, L.; Vu, T.H.; Werb, Z.; Winding, B.; Lochter, A.; et al. Proteinases in bone resorption: obvious and less obvious roles. Clin. Chim. Acta 2000, 291, 223–234. [Google Scholar] [CrossRef]
- Hegarty, V.M.; May, H.M.; Khaw, K.T. Tea drinking and bone mineral density in older women. Am. J. Clin. Nutr. 2000, 71, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Yasui, T.; Hirose, J.; Aburatani, H.; Tanaka, S. Epigenetic regulation of osteoclast differentiation. Ann. N. Y. Acad. Sci. 2011, 1240, 7–13. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takemoto, M.; Takemoto, H. Synthesis of Theaflavins and Their Functions. Molecules 2018, 23, 918. https://doi.org/10.3390/molecules23040918
Takemoto M, Takemoto H. Synthesis of Theaflavins and Their Functions. Molecules. 2018; 23(4):918. https://doi.org/10.3390/molecules23040918
Chicago/Turabian StyleTakemoto, Masumi, and Hiroaki Takemoto. 2018. "Synthesis of Theaflavins and Their Functions" Molecules 23, no. 4: 918. https://doi.org/10.3390/molecules23040918
APA StyleTakemoto, M., & Takemoto, H. (2018). Synthesis of Theaflavins and Their Functions. Molecules, 23(4), 918. https://doi.org/10.3390/molecules23040918