
The Effects of Plant-Derived Oleanolic Acid on Selected Parameters of Glucose Homeostasis in a Diet-Induced Pre-Diabetic Rat Model


Abstract
1. Introduction
2. Results
2.1. Caloric Intake
2.2. Body Weights
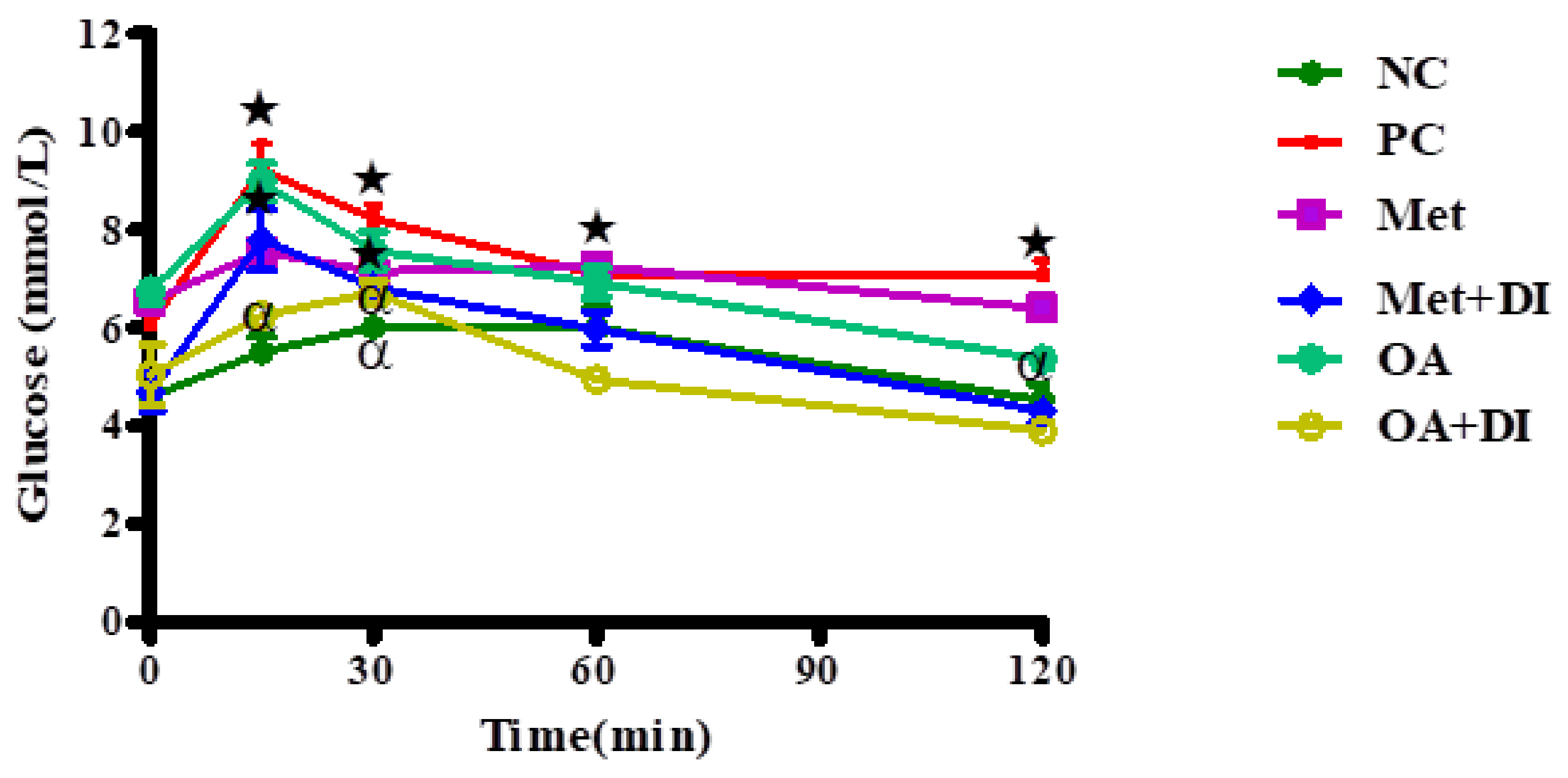
2.3. Oral Glucose Tolerance (OGTT)
2.4. HOMAR2-IR Index
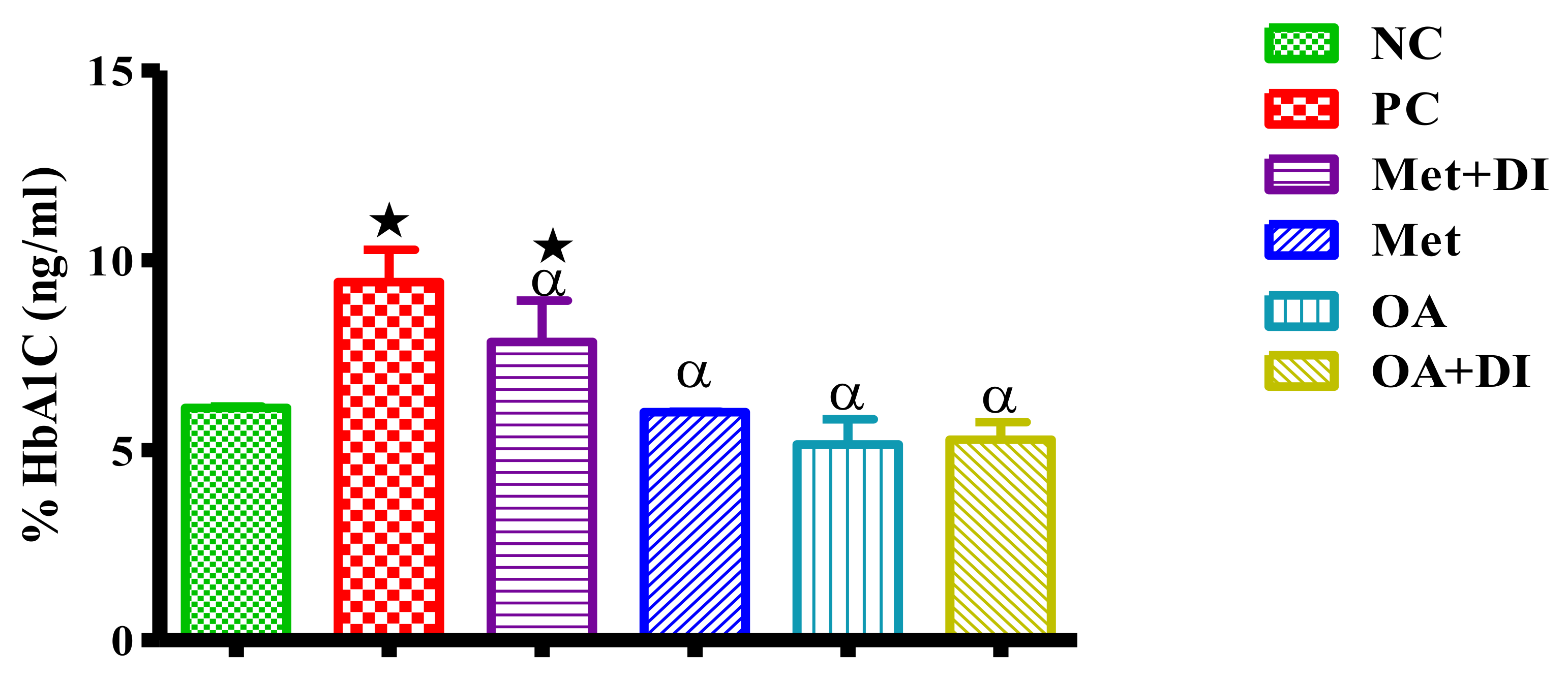
2.5. Glycated Haemoglobin Concentration (HbA1c)
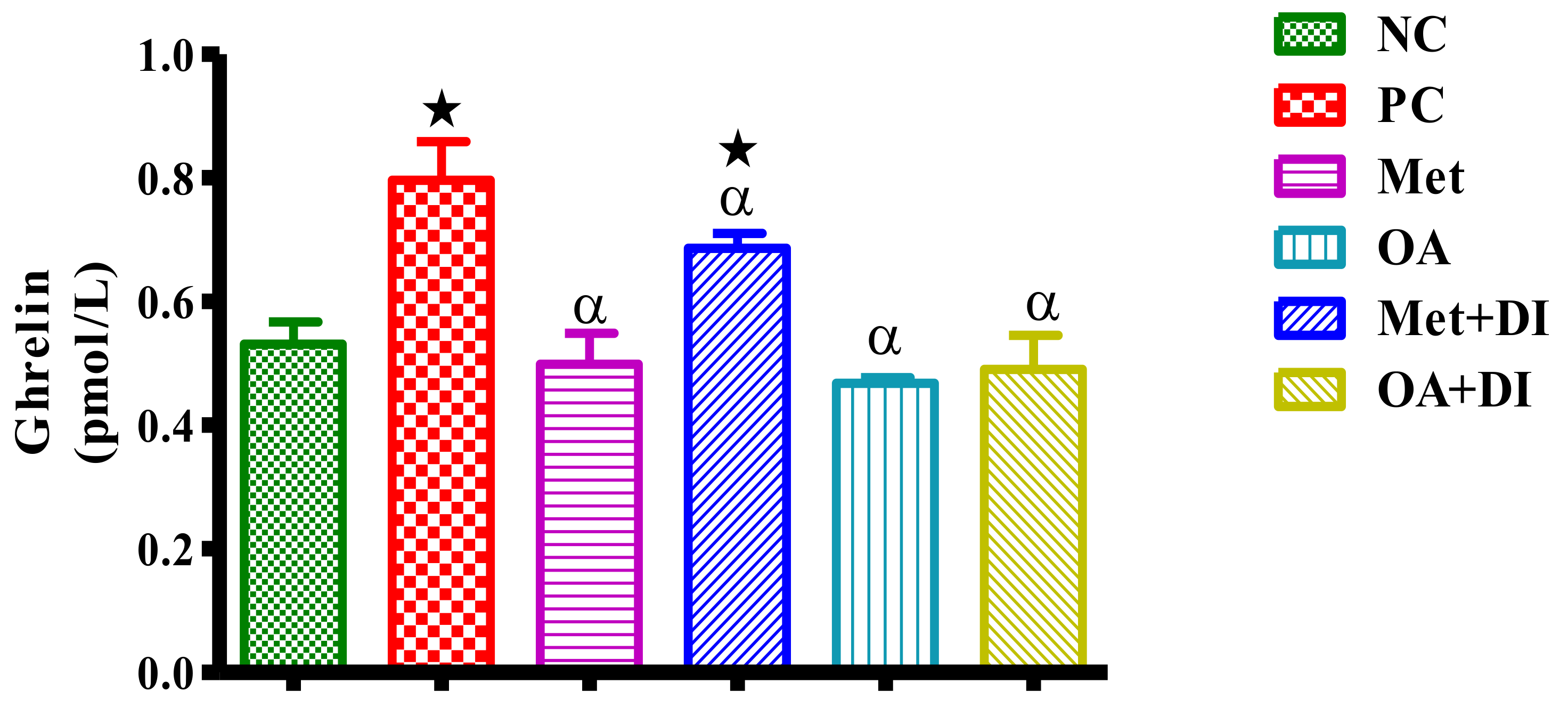
2.6. Ghrelin Concentration
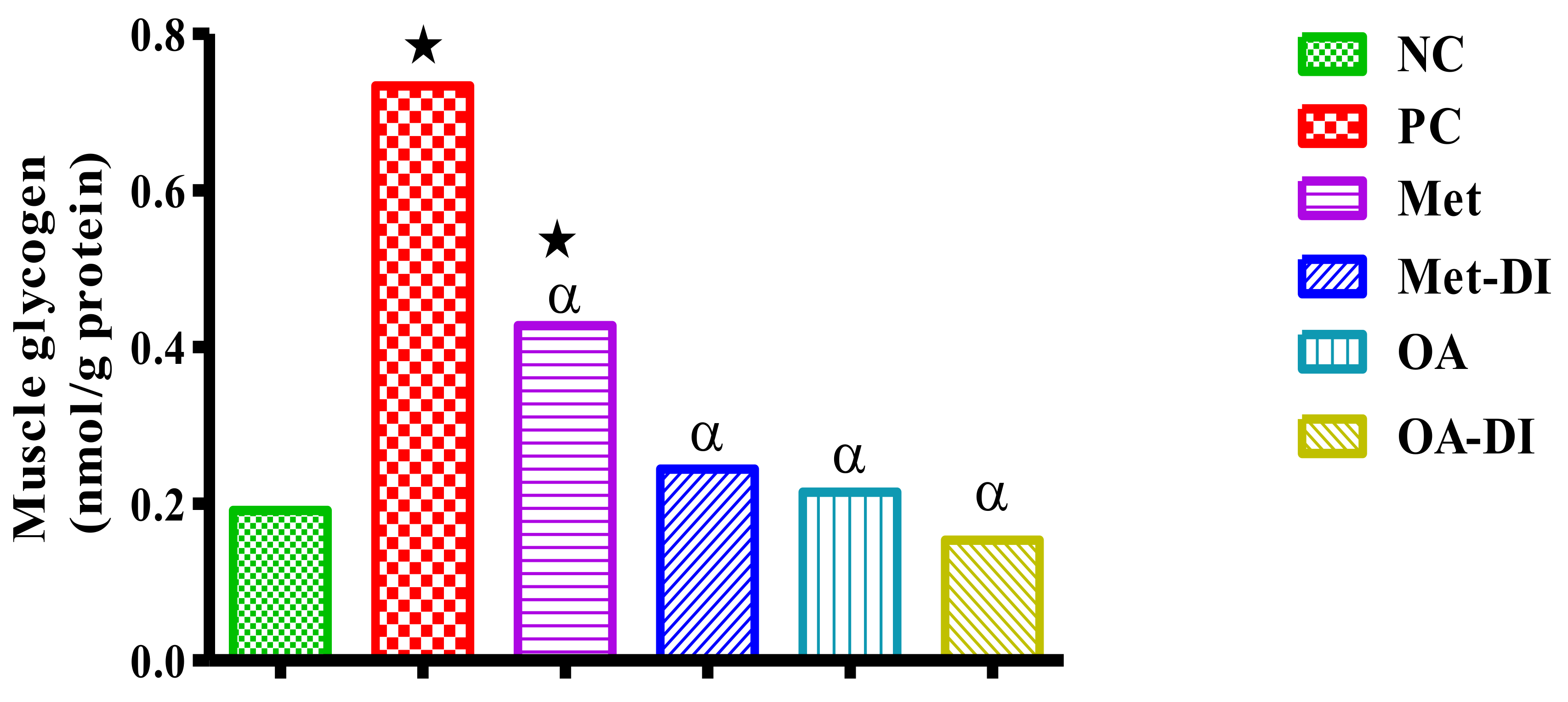
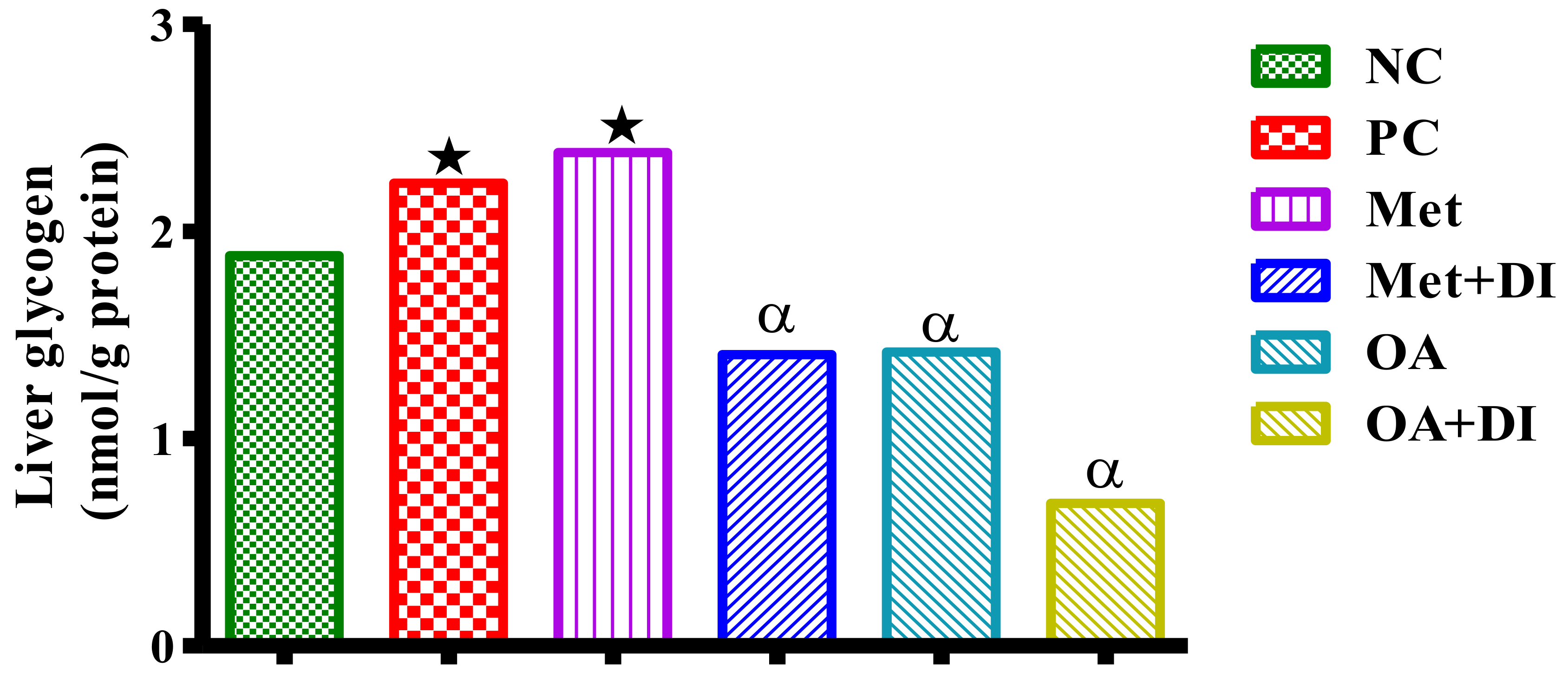
2.7. Skeletal Muscle and Liver Glycogen Concentration
3. Discussion
4. Materials and Methods
4.1. Drugs and Chemicals
4.2. Extraction Method and Administration
4.3. Animals
4.3.1. Induction of Pre-Diabetes
4.3.2. Experimental Design
4.3.3. Treatment of Pre-Diabetic Animals
2.3.4. Blood Collection and Tissue Harvesting
4.4. Biochemical Analysis
4.5. Glycogen Assay
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
Author contributions
Conflicts of Interest
References
- Wang, C.; Zhang, Y.; Zhang, L.; Hou, X.; Lu, H.; Shen, Y.; Chen, R.; Fang, P.; Yu, H.; Li, M. Prevalence of type 2 diabetes among high-risk adults in shanghai from 2002 to 2012. PLoS ONE 2014, 9, e102926. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.-H.; Lee, J.-H.; Kim, J.-W.; Cho, J.H.; Choi, Y.-H.; Ko, S.-H.; Zimmet, P.; Son, H.-Y. Epidemic obesity and type 2 diabetes in asia. Lancet 2006, 368, 1681–1688. [Google Scholar] [CrossRef]
- Panchal, S.; Poudyal, H.; Iyer, A.; Nazer, R.; Alam, A.; Diwan, V.; Kauter, K.; Sernia, C.; Campbell, F.; Ward, P.; et al. High-carbohydrate, high-fat diet–induced metabolic syndrome and cardiovascular remodeling in rats. J. Cardiovasc. Pharm. 2011, 57, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Tuomilehto, J.; Lindström, J.; Eriksson, J.G.; Valle, T.T.; Hämäläinen, H.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S.; Laakso, M.; Louheranta, A.; Rastas, M.; et al. Prevention of type 2 diabetes mellitus by changes in lifestyle among subjects with impaired glucose tolerance. N. Engl. J. Med. 2001, 344, 1343–1350. [Google Scholar] [CrossRef] [PubMed]
- Rebnord, E.W.; Pedersen, E.R.; Strand, E.; Svingen, G.F.T.; Meyer, K.; Schartum-Hansen, H.; Løland, K.H.; Seifert, R.; Ueland, P.M.; Nilsen, D.W.T.; et al. Glycated hemoglobin and long-term prognosis in patients with suspected stable angina pectoris without diabetes mellitus: A prospective cohort study. Atherosclerosis 2015, 240, 115–120. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, A.M.; Johnson, J.D.; Clee, S.M.; Kieffer, T.J. Suppressing hyperinsulinemia prevents obesity but causes rapid onset of diabetes in leptin-deficient lepob/ob mice. Mol. Metab. 2016, 5, 1103–1112. [Google Scholar] [CrossRef] [PubMed]
- Walsh, J.M.; Byrne, J.; Mahony, R.M.; Foley, M.E.; McAuliffe, F.M. Leptin, fetal growth and insulin resistance in non-diabetic pregnancies. Early Hum. Dev. 2014, 90, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Man, R.E.K.; Charumathi, S.; Gan, A.T.L.; Fenwick, E.K.; Tey, C.S.; Chua, J.; Wong, T.-Y.; Cheng, C.-Y.; Lamoureux, E.L. Cumulative incidence and risk factors of prediabetes and type 2 diabetes in a singaporean malay cohort. Diabetes Res. Clin. Pract. 2017, 127, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Mosa, Z.M.; El Badry, Y.A.; Fattah, H.S.; Mohamed, E.G. Comparative study between the effects of some dietary sources and metformin drug on weight reduction in obese rats. Ann. Agric. Sci. 2015, 60, 381–388. [Google Scholar] [CrossRef]
- Watson, C.S. Prediabetes: Screening, diagnosis, and intervention. J. Nurse Pract. 2017, 13, 216–221.e1. [Google Scholar] [CrossRef]
- Chaudhari, P.; Vallarino, C.; Law, E.H.; Seifeldin, R. Evaluation of patients with type 2 diabetes mellitus receiving treatment during the pre-diabetes period: Is early treatment associated with improved outcomes? Diabetes Res. Clin. Pract. 2016, 122, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Courcoulas, A.P.; Belle, S.H.; Neiberg, R.H.; Pierson, S.K.; Eagleton, J.K.; Kalarchian, M.A.; DeLany, J.P.; Lang, W.; Jakicic, J.M. Three-year outcomes of bariatric surgery vs lifestyle intervention for type 2 diabetes mellitus treatment: A randomized clinical trial. JAMA Surg. 2015, 150, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Inzucchi, S.E.; Bergenstal, R.M.; Buse, J.B.; Diamant, M.; Ferrannini, E.; Nauck, M.; Peters, A.L.; Tsapas, A.; Wender, R.; Matthews, D.R. Management of hyperglycaemia in type 2 diabetes, 2015: A patient-centred approach. Update to a position statement of the american diabetes association and the european association for the study of diabetes. Diabetologia 2015, 58, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Katashima, C.K.; Silva, V.R.; Gomes, T.L.; Pichard, C.; Pimentel, G.D. Ursolic acid and mechanisms of actions on adipose and muscle tissue: A systematic review. Obes. Rev. 2017, 18, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Przybylska-Gornowicz, B.; Zakłos-Szyda, M.; Dąbrowski, M.; Michalczuk, L.; Koziołkiewicz, M.; Babuchowski, A.; Zielonka, Ł.; Lewczuk, B.; Gajęcki, M.T. The influence of a natural triterpene preparation on the gastrointestinal tract of gilts with streptozocin-induced diabetes and on cell metabolic activity. J. Funct. Foods 2017, 33, 11–20. [Google Scholar] [CrossRef]
- Khathi, A.; Serumula, M.R.; Myburg, R.B.; Van Heerden, F.R.; Musabayane, C.T. Effects of syzygium aromaticum-derived triterpenes on postprandial blood glucose in streptozotocin-induced diabetic rats following carbohydrate challenge. PLoS ONE 2013, 8, e81632. [Google Scholar] [CrossRef] [PubMed]
- Mkhwanazi, B.N.; Serumula, M.R.; Myburg, R.B.; Van Heerden, F.R.; Musabayane, C.T. Antioxidant effects of maslinic acid in livers, hearts and kidneys of streptozotocin-induced diabetic rats: Effects on kidney function. Ren. Fail. 2014, 36, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Mukundwa, A.; Langa, S.O.; Mukaratirwa, S.; Masola, B. In Vivo effects of diabetes, insulin and oleanolic acid on enzymes of glycogen metabolism in the skin of streptozotocin-induced diabetic male sprague-dawley rats. Biochem. Biophys. Res. Commun. 2016, 471, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Luvuno, M.; Kathi, A.; Mabandla, M.V. Voluntary Ingestion of a High-Fat High-Carbohydrate Diet: A Model for Prediabetes. Master’s Dissertation, University of KwaZulu-Natal, KwaZulu-Natal, South Africa, 2017. [Google Scholar]
- Chen, P.; Zeng, H.; Wang, Y.; Fan, X.; Xu, C.; Deng, R.; Zhou, X.; Bi, H.; Huang, M. Low dose of oleanolic acid protects against lithocholic acid-induced cholestasis in mice: Potential involvement of nuclear factor-e2-related factor 2-mediated upregulation of multidrug resistance-associated proteins. Drug Metab. Dispos. 2014, 42, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Luvuno, M.; Mbongwa, H.P.; Khathi, A. The effects of syzygium aromaticum-derived triterpenes on gastrointestinal ghrelin expression in streptozotocin-induced diabetic rats. Afr. J. Tradit. Complement. Altern. Med. 2016, 13, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Alamri, B.N.; Shin, K.; Chappe, V.; Anini, Y. The role of ghrelin in the regulation of glucose homeostasis. Horm. Mol. Biol. Clin. Investig. 2016, 26, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Theander-Carrillo, C.; Wiedmer, P.; Cettour-Rose, P.; Nogueiras, R.; Perez-Tilve, D.; Pfluger, P.; Castaneda, T.R.; Muzzin, P.; Schürmann, A.; Szanto, I. Ghrelin action in the brain controls adipocyte metabolism. J. Clin. Investig. 2006, 116, 1983. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.H.; Ha, Y.J.; Shim, E.K.; Choi, S.Y.; Jin, J.L.; Yun-Choi, H.S.; Lee, J.R. Insulin-mimetic and insulin-sensitizing activities of a pentacyclic triterpenoid insulin receptor activator. Biochem. J. 2007, 403, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Choi, M.-S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed]
- Magkos, F.; Fraterrigo, G.; Yoshino, J.; Luecking, C.; Kirbach, K.; Kelly, S.C.; de las Fuentes, L.; He, S.; Okunade, A.L.; Patterson, B.W. Effects of moderate and subsequent progressive weight loss on metabolic function and adipose tissue biology in humans with obesity. Cell Metab. 2016, 23, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577. [Google Scholar] [CrossRef] [PubMed]
- Merovci, A.; Solis-Herrera, C.; Daniele, G.; Eldor, R.; Fiorentino, T.V.; Tripathy, D.; Xiong, J.; Perez, Z.; Norton, L.; Abdul-Ghani, M.A. Dapagliflozin improves muscle insulin sensitivity but enhances endogenous glucose production. J. Clin. Investig. 2014, 124, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Noakes, T.D.; Windt, J. Evidence that supports the prescription of low-carbohydrate high-fat diets: A narrative review. Br. J. Sports Med. 2017, 51, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, D.K.; Asati, V.; Bharti, S.K. Chalcones and their therapeutic targets for the management of diabetes: Structural and pharmacological perspectives. Eur. J. Med. Chem. 2015, 92, 839–865. [Google Scholar] [CrossRef] [PubMed]
- Evert, A.B.; Boucher, J.L.; Cypress, M.; Dunbar, S.A.; Franz, M.J.; Mayer-Davis, E.J.; Neumiller, J.J.; Nwankwo, R.; Verdi, C.L.; Urbanski, P. Nutrition therapy recommendations for the management of adults with diabetes. Diabetes Care 2014, 37, S120–S143. [Google Scholar] [CrossRef] [PubMed]
- Colhoun, H.M.; Betteridge, D.J.; Durrington, P.N.; Hitman, G.A.; Neil, H.A.W.; Livingstone, S.J.; Thomason, M.J.; Mackness, M.I.; Charlton-Menys, V.; Fuller, J.H. Primary prevention of cardiovascular disease with atorvastatin in type 2 diabetes in the collaborative atorvastatin diabetes study (cards): Multicentre randomised placebo-controlled trial. Lancet 2004, 364, 685–696. [Google Scholar] [CrossRef]
- Donnan, P.T.; MacDonald, T.M.; Morris, A.D. Adherence to prescribed oral hypoglycaemic medication in a population of patients with type 2 diabetes: A retrospective cohort study. Diabet. Med. 2002, 19, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, M.J.; Wierts, R.; Hoeks, J.; Gemmink, A.; Brans, B.; Mottaghy, F.M.; Schrauwen, P.; van Marken Lichtenbelt, W.D. Glucose uptake in human brown adipose tissue is impaired upon fasting-induced insulin resistance. Diabetologia 2015, 58, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Ngubane, P.S.; Masola, B.; Musabayane, C.T. The effects of syzygium aromaticum-derived oleanolic acid on glycogenic enzymes in streptozotocin-induced diabetic rats. Ren. Fail. 2011, 33, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Shi, G.; Zhang, X.; Gong, S.; Tan, S.; Chen, B.; Che, H.; Li, T. Mesoscale modelling study of the interactions between aerosols and pbl meteorology during a haze episode in china jing–jin–ji and its near surrounding region—Part 2: Aerosols’ radiative feedback effects. Atmos. Chem. Phys. 2015, 15, 3277–3287. [Google Scholar] [CrossRef]
- Wallace, T.M.; Levy, J.C.; Matthews, D.R. Use and abuse of homa modeling. Diabetes Care 2004, 27, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Monje, A.; Catena, A.; Borgnakke, W.S. Association between diabetes mellitus/hyperglycemia and peri-implant diseases: Systematic review and meta-analysis. J. Clin. Periodontol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Richter, B.; Hemmingsen, B.; Metzendorf, M.I.; Takwoingi, Y. Intermediate hyperglycaemia as a predictor for the development of type 2 diabetes: Prognostic factor exemplar review. Cochrane Libr. 2017. [Google Scholar] [CrossRef]
- Myers, R.W.; Guan, H.-P.; Ehrhart, J.; Petrov, A.; Prahalada, S.; Tozzo, E.; Yang, X.; Kurtz, M.M.; Trujillo, M.; Trotter, D.G. Systemic pan-ampk activator mk-8722 improves glucose homeostasis but induces cardiac hypertrophy. Science 2017, 357, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Perumal, V.; Narayanan, N.; Rangarajan, J.; Palanisamy, E.; Kalifa, M.; Maheshwari, U.; Perincheri, P.S.S.; Chinnathambi, S. A study to correlate hba1c levels and left ventricular diastolic dysfunction in newly diagnosed type ii diabetes mellitus. J. Evol. Med. Dent. Sci. 2016, 5, 3412–3417. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Jena, G. Protective role of sodium butyrate, a hdac inhibitor on beta-cell proliferation, function and glucose homeostasis through modulation of p38/erk mapk and apoptotic pathways: Study in juvenile diabetic rat. Chem.-Biol. Interact. 2014, 213, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Musabayane, C.; Mahlalela, N.; Shode, F.; Ojewole, J. Effects of Syzygium cordatum (hochst.) [myrtaccae] leaf extract on plasma glucose and hepatic glycogen in streptozotocin-induce diabetic rats. J. Ethnopharmacol. 2005, 97, 485–490. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Caloric Intake (kcal/g) | ||||
|---|---|---|---|---|
| Experimental Groups | Week 0 | Week 4 | Week 8 | Week 12 |
| NC | 109.18 ± 1.9 (100%) | 125.042 ± 2.34 ↑ (14.52%) | 165.04 ± 1.61 ↑ (51.16%) | 178.40 ± 0.87 ↑ (63.34%) |
| PC | 121.47 ± 1.01 * (100%) | 120.90 ± 0.64 * ↓ (0.47%) | 206.58 ± 0.84 * ↑ (70.07%) | 230.01 ± 0.85 * ↑ (89.36%) |
| Met | 118.09 ± 0.51 * (100%) | 100.54 ± 0.98 * α↓ (14.86%) | 99.51 ± 1.52 * α↓ (15.73%) | 151.66 ± 0.69 * α↑ (28.43%) |
| Met + DI | 115.02 ± 0.67 * (100%) | 102.69 ± 1.17 * α↓ (10.72%) | 120.51 ± 0.75 * α↑ (4.77%) | 144.72 ± 1.64 * α↑ (25.82%) |
| OA | 130.35 ± 0.03 * (100%) | 103.94 ± 2.02 * α↓ (20.26%) | 156.09 ± 1.63 * α↑ (19.75%) | 194.26 ± 1.85 * α↑ (49.03%) |
| OA + DI | 117.58 ± 0.51 * (100%) | 118.04 ± 0.85 *α ↑ (0.40%) | 147.59 ± 2.74 *α ↑ (25.52%) | 168.82 ± 2.22α ↑ (43.58%) |
| HOMAR2-IR Index | |||
|---|---|---|---|
| Experimental Group | Plasma Glucose (mmol/L) | Plasma Insulin (mU/L) | HOMAR2-IR Values |
| NC | 4.60 ± 0.09 | 61.96 ± 0.90 | 12.670 ± 0.61 |
| PC | 5.87 ± 0.32 * | 491.64 ± 3.45 * | 128.26 ± 2.98 * |
| Met | 6.55 ± 0.81 *α | 169.16 ± 3.12 *α | 15.93 ± 1.02 *α |
| Met + DI | 4.52 ± 0.90α | 142.94 ± *α | 49.25 ± 1.15 *α |
| OA | 6.77 ± 1.59 *α | 200.58 ± 2.85 *α | 60.35 ± 2.05 *α |
| OA + DI | 4.47 ± 0.12α | 68.59 ± 2.01α | 13.63 ± 0.95α |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gamede, M.; Mabuza, L.; Ngubane, P.; Khathi, A. The Effects of Plant-Derived Oleanolic Acid on Selected Parameters of Glucose Homeostasis in a Diet-Induced Pre-Diabetic Rat Model. Molecules 2018, 23, 794. https://doi.org/10.3390/molecules23040794
Gamede M, Mabuza L, Ngubane P, Khathi A. The Effects of Plant-Derived Oleanolic Acid on Selected Parameters of Glucose Homeostasis in a Diet-Induced Pre-Diabetic Rat Model. Molecules. 2018; 23(4):794. https://doi.org/10.3390/molecules23040794
Chicago/Turabian StyleGamede, Mlindeli, Lindokuhle Mabuza, Phikelelani Ngubane, and Andile Khathi. 2018. "The Effects of Plant-Derived Oleanolic Acid on Selected Parameters of Glucose Homeostasis in a Diet-Induced Pre-Diabetic Rat Model" Molecules 23, no. 4: 794. https://doi.org/10.3390/molecules23040794
APA StyleGamede, M., Mabuza, L., Ngubane, P., & Khathi, A. (2018). The Effects of Plant-Derived Oleanolic Acid on Selected Parameters of Glucose Homeostasis in a Diet-Induced Pre-Diabetic Rat Model. Molecules, 23(4), 794. https://doi.org/10.3390/molecules23040794