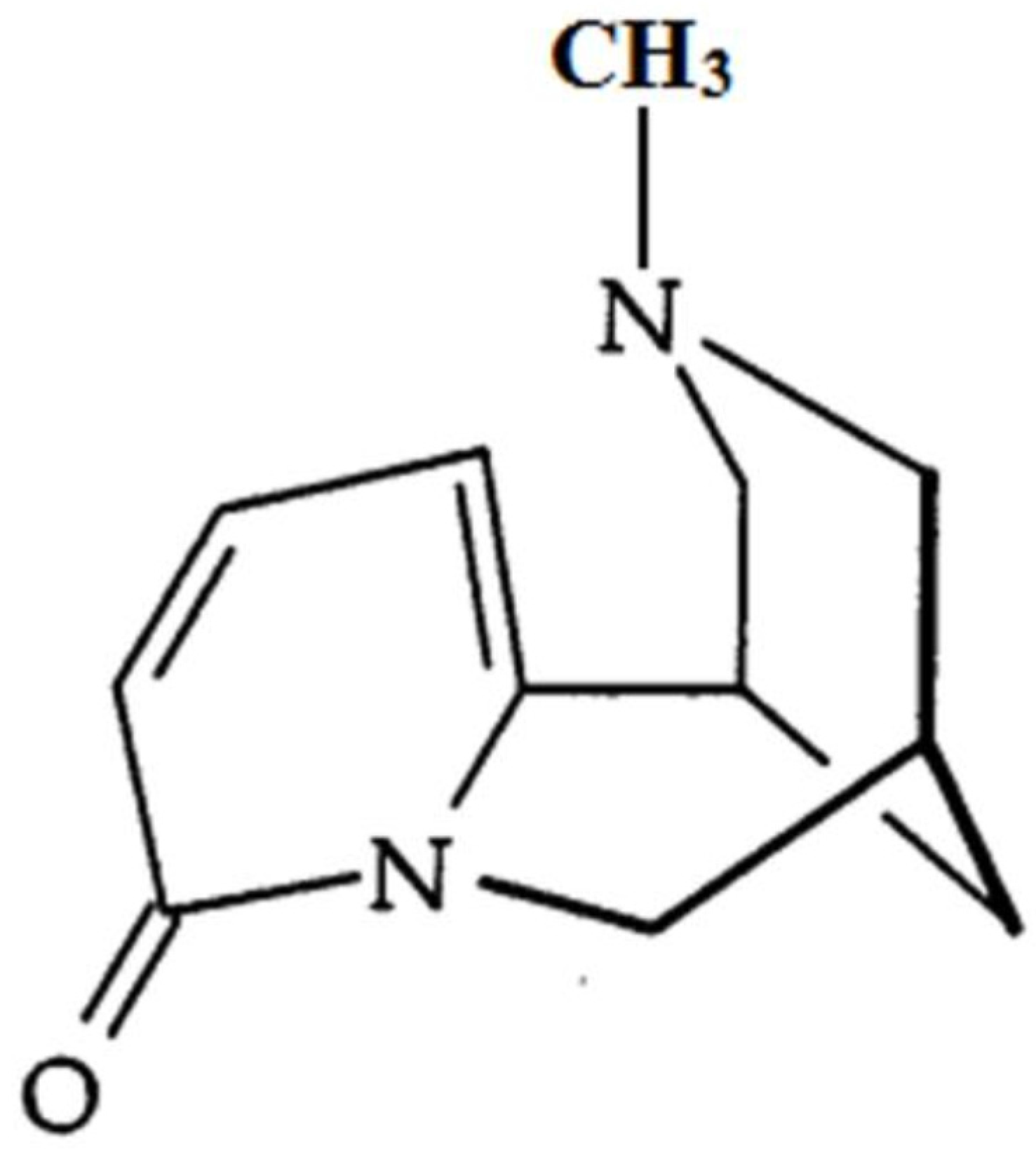
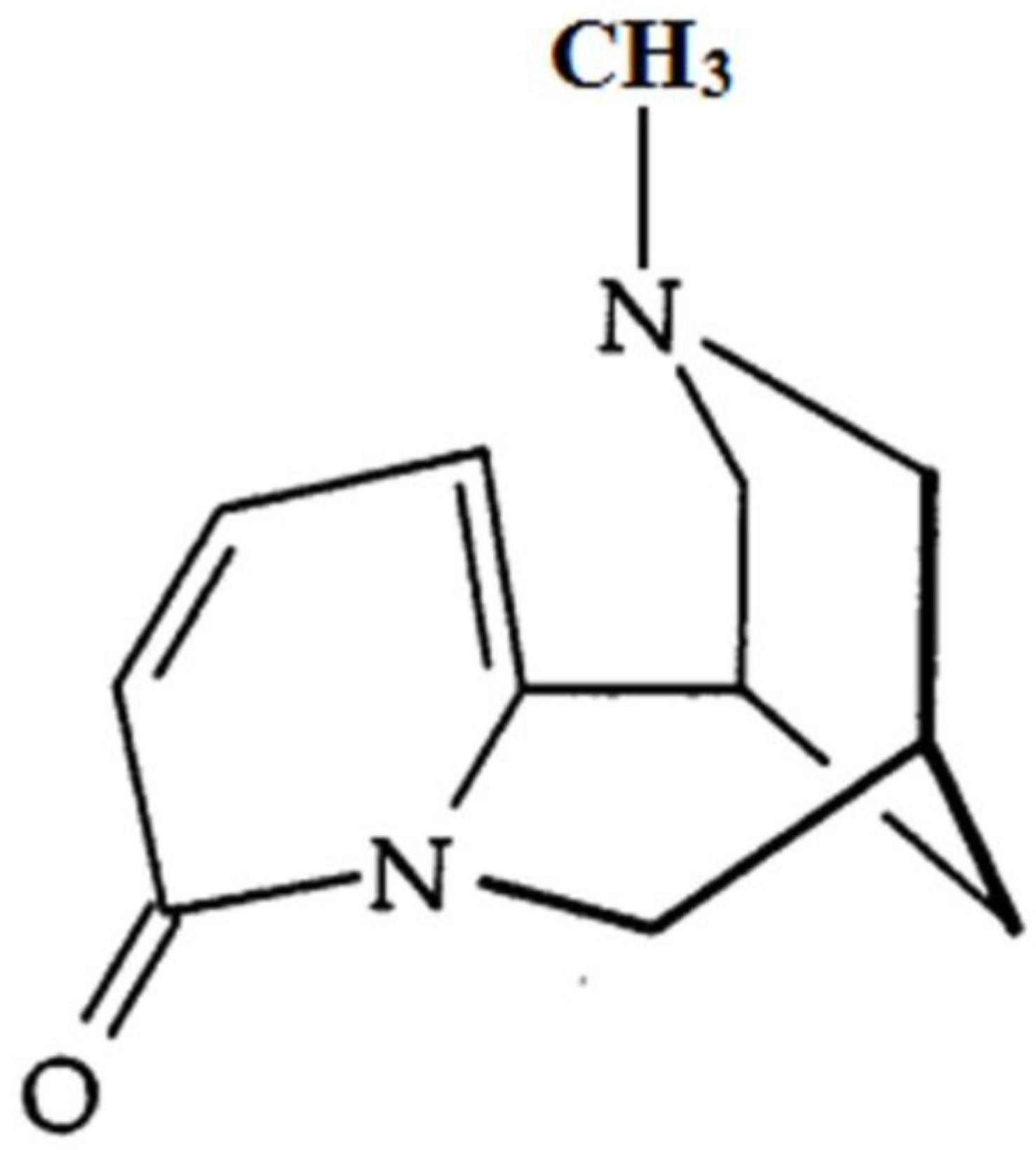
N-Methylcytisine Ameliorates Dextran-Sulfate-Sodium-Induced Colitis in Mice by Inhibiting the Inflammatory Response

Abstract
:1. Introduction
2. Results
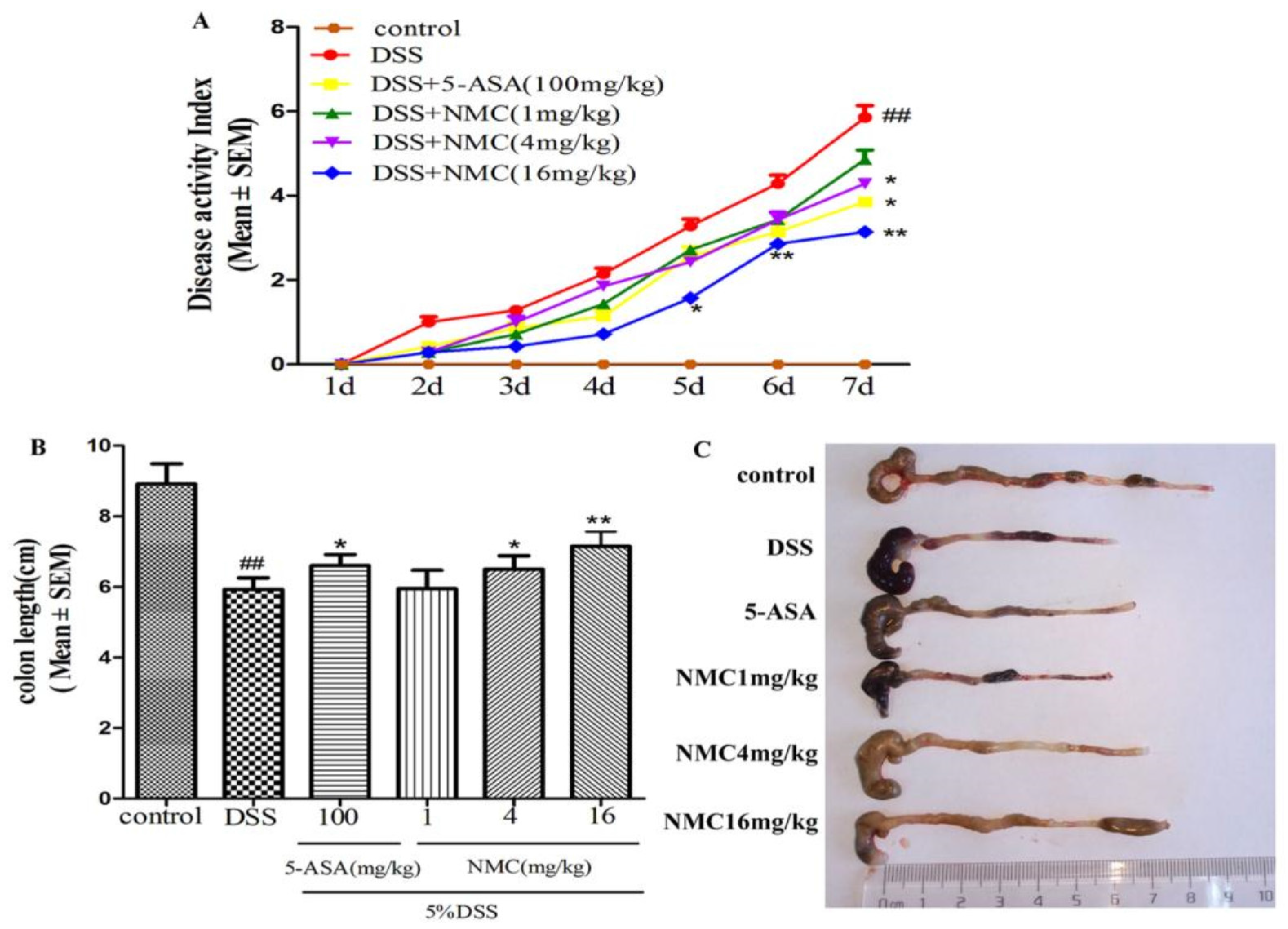
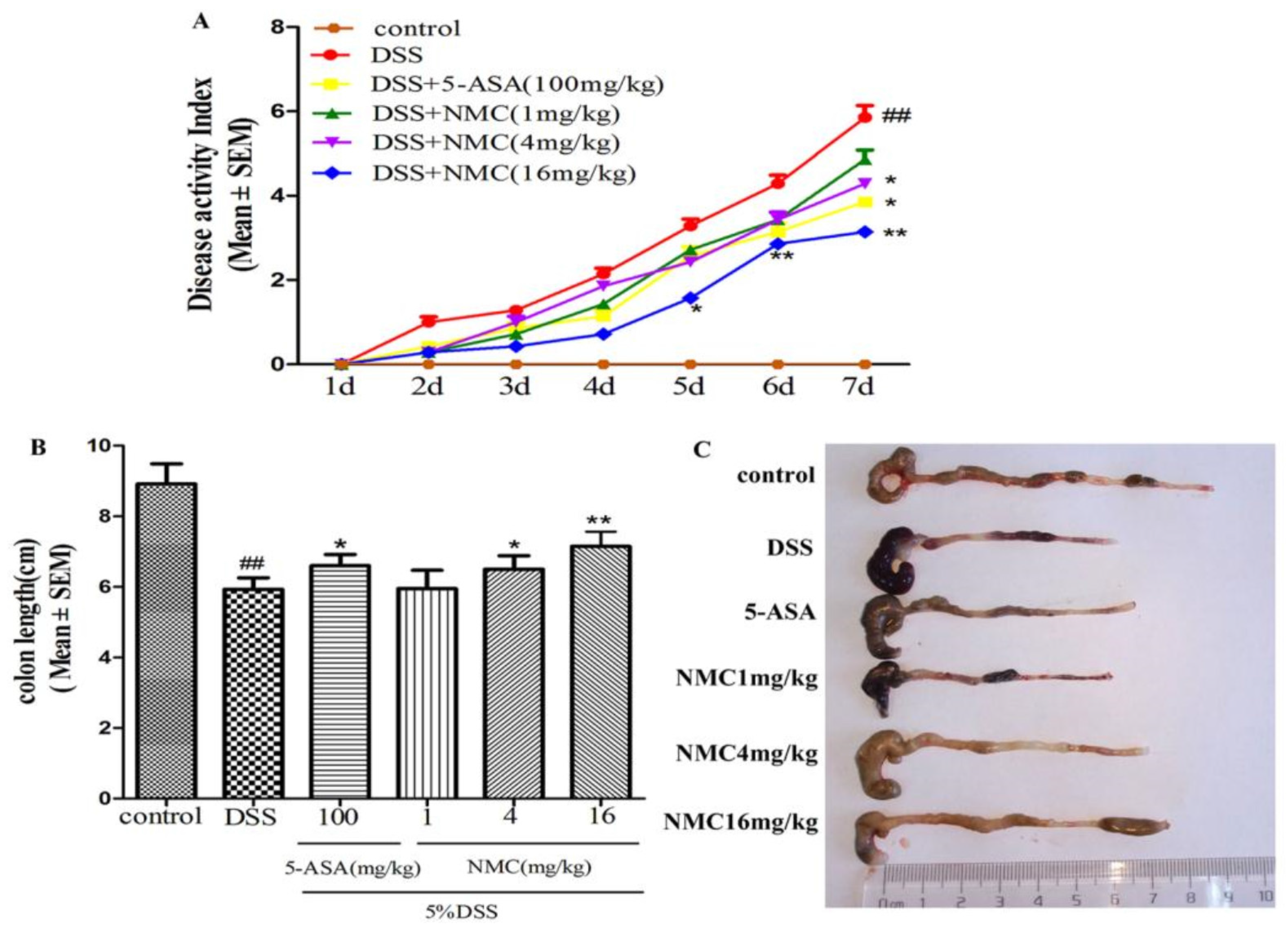
2.1. NMC Ameliorated the Clinical Symptoms of DSS-Induced Colitis in Mice
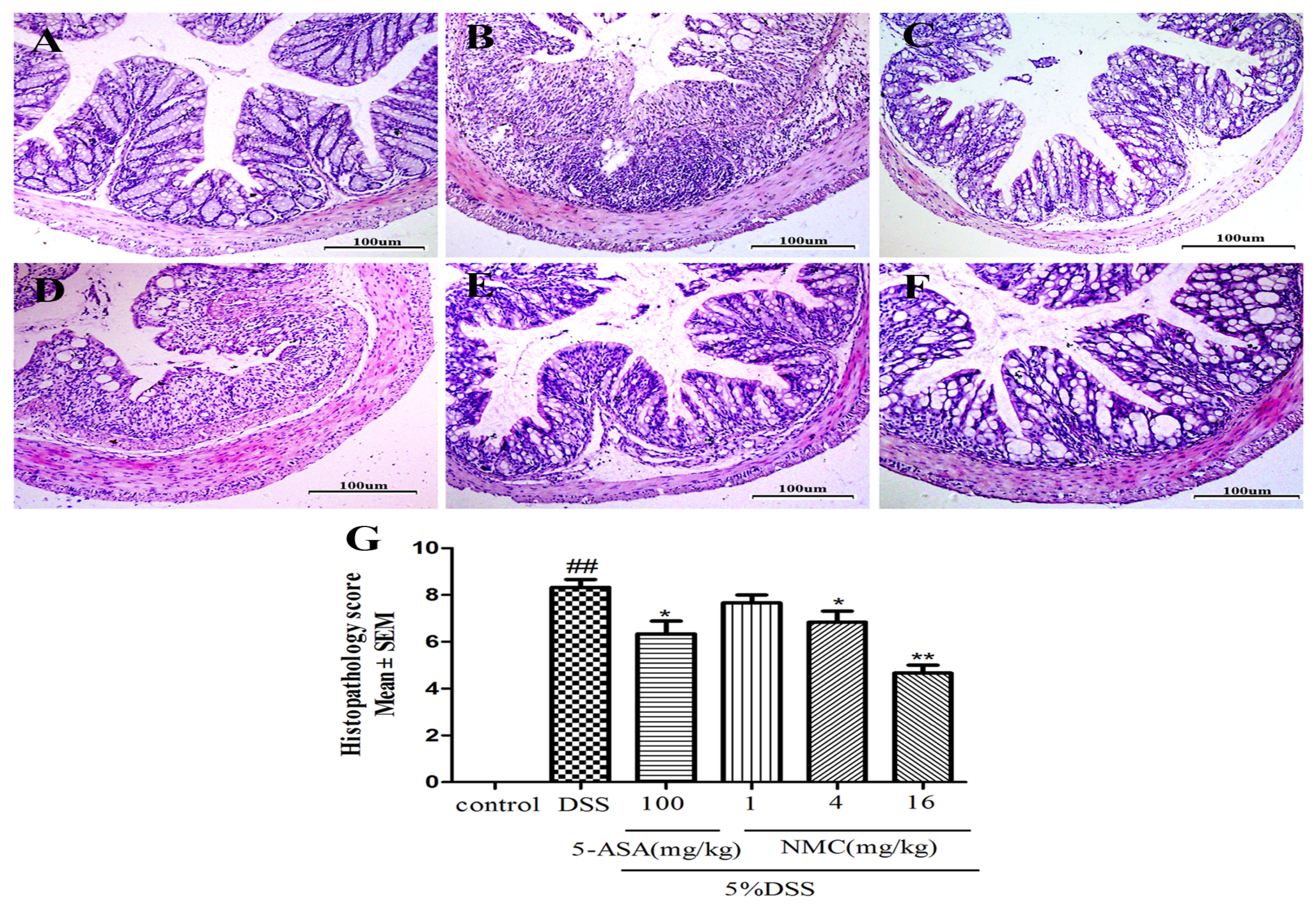
2.2. NMC Attenuated the DSS-Induced Colonic Histopathological Changes
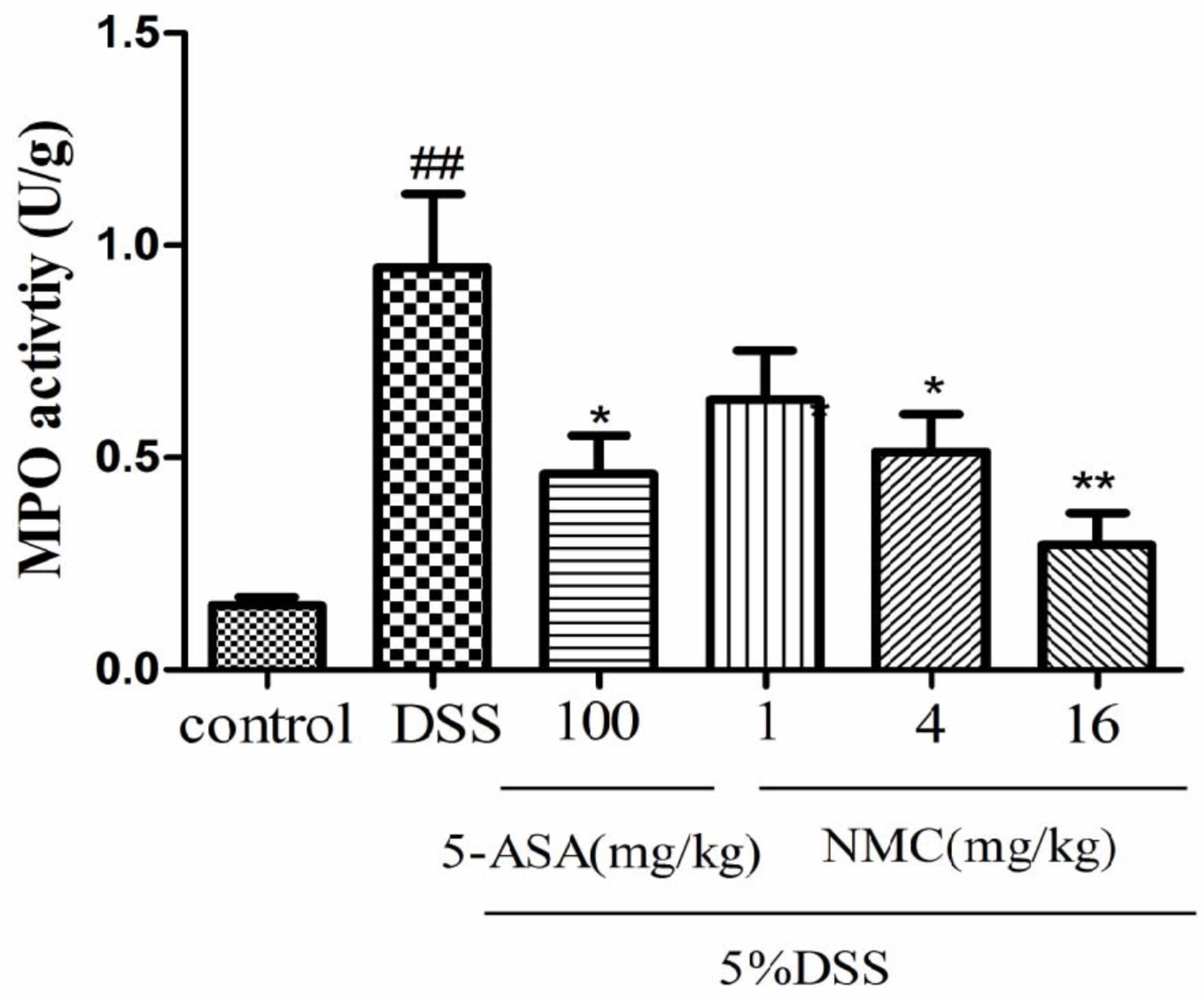
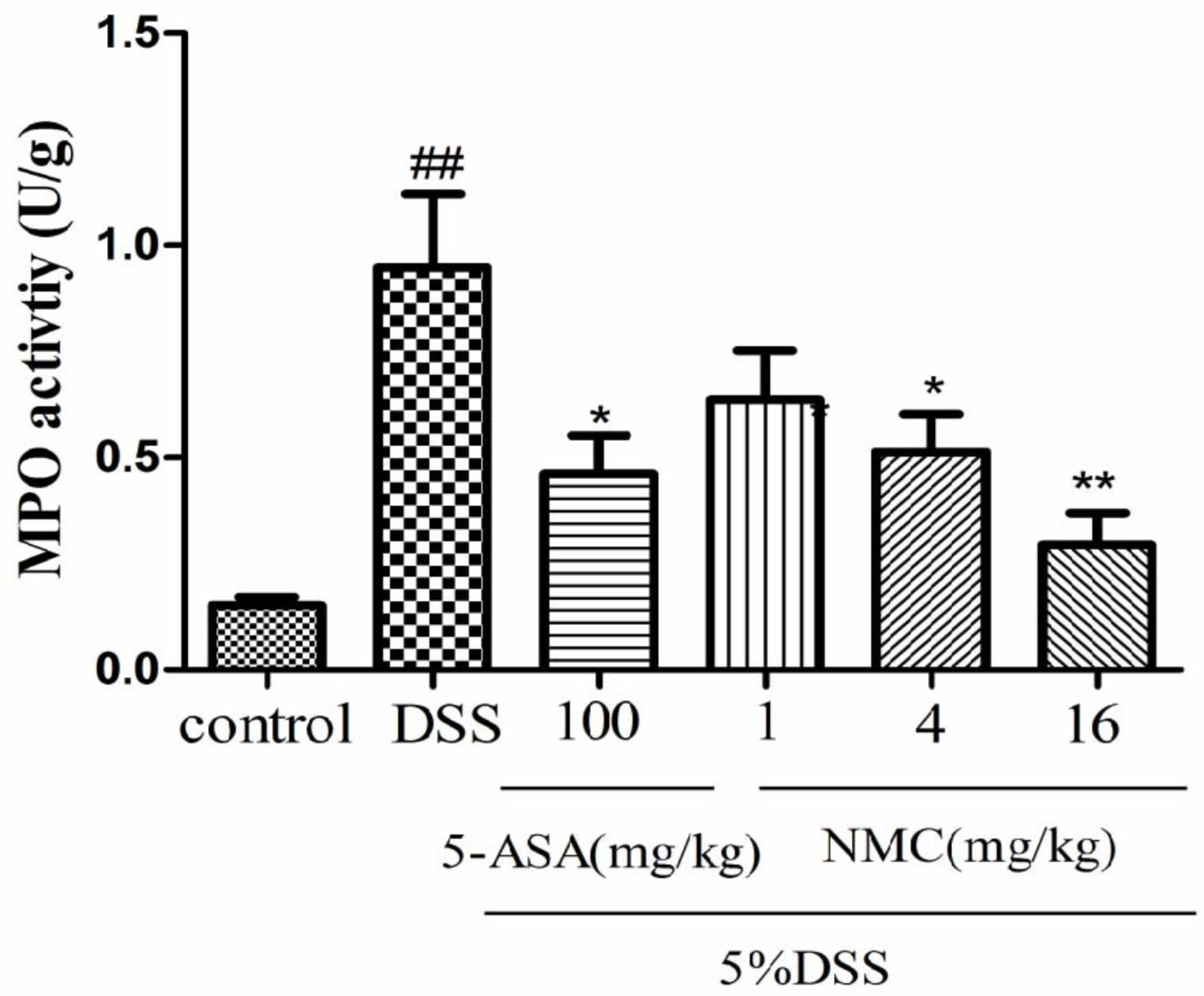
2.3. NMC Decreased the Activity of Myeloperoxidase (MPO) in the Colon
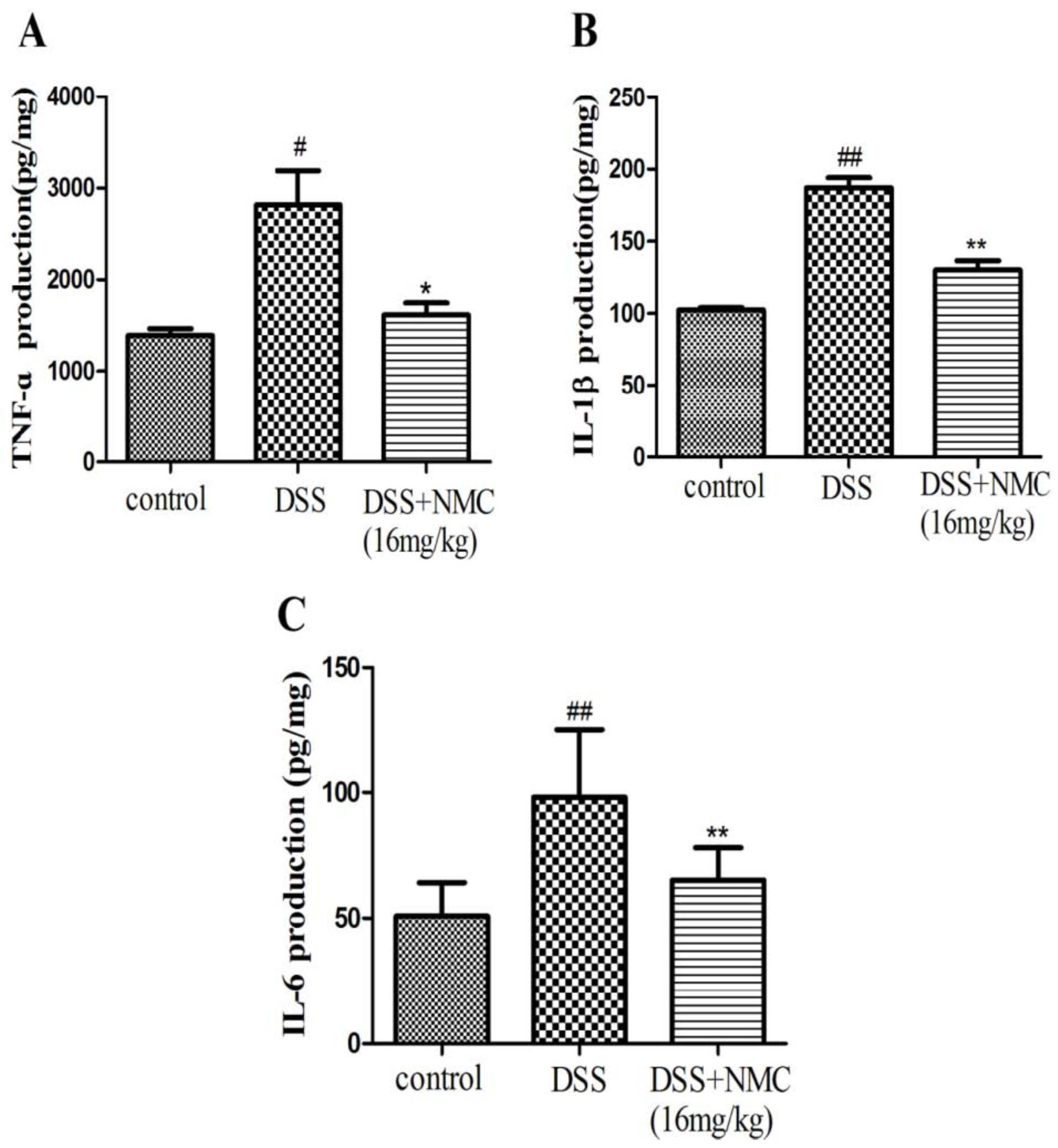
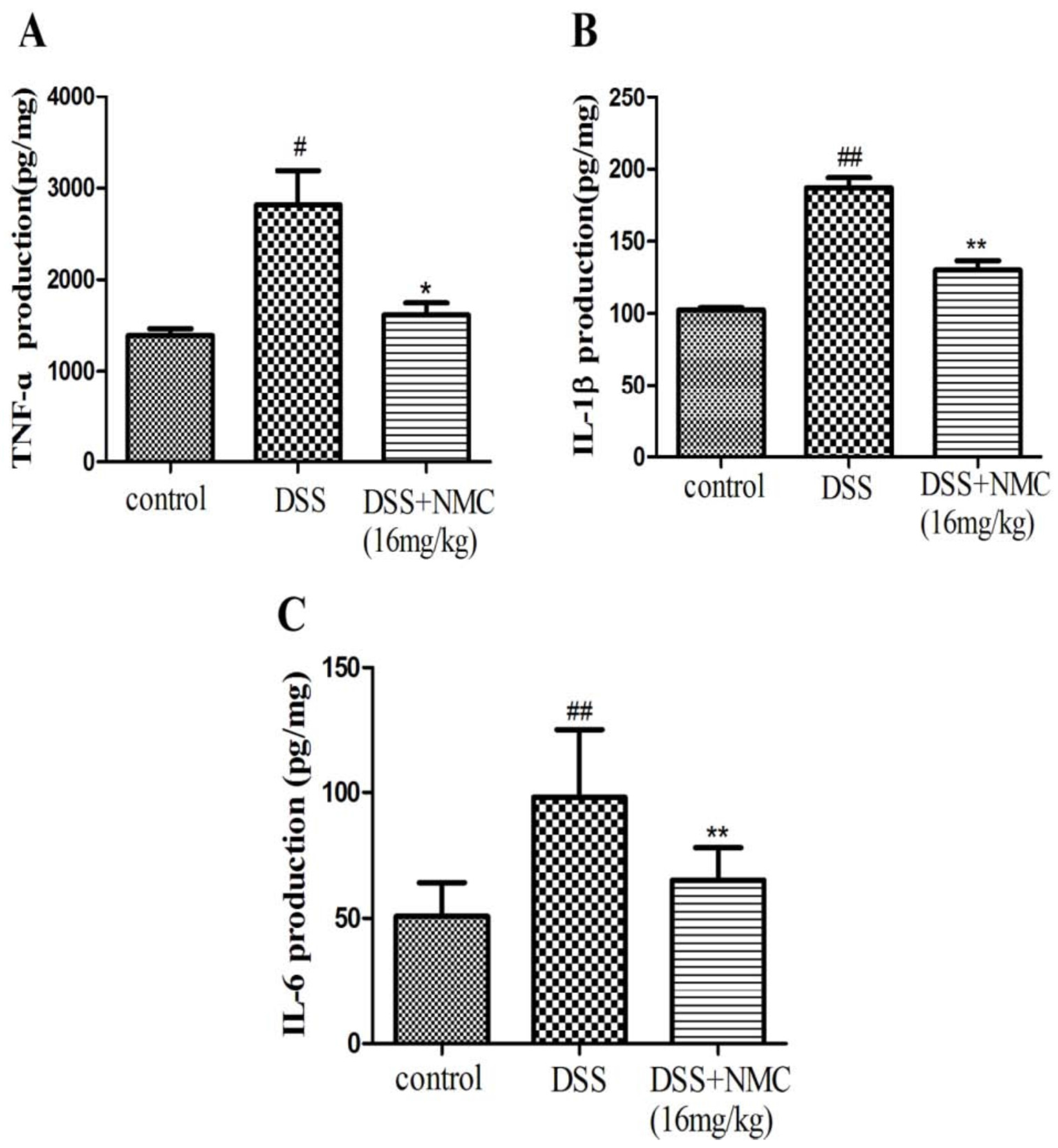
2.4. NMC Reduced the Pro-Inflammatory Cytokines in the Colon
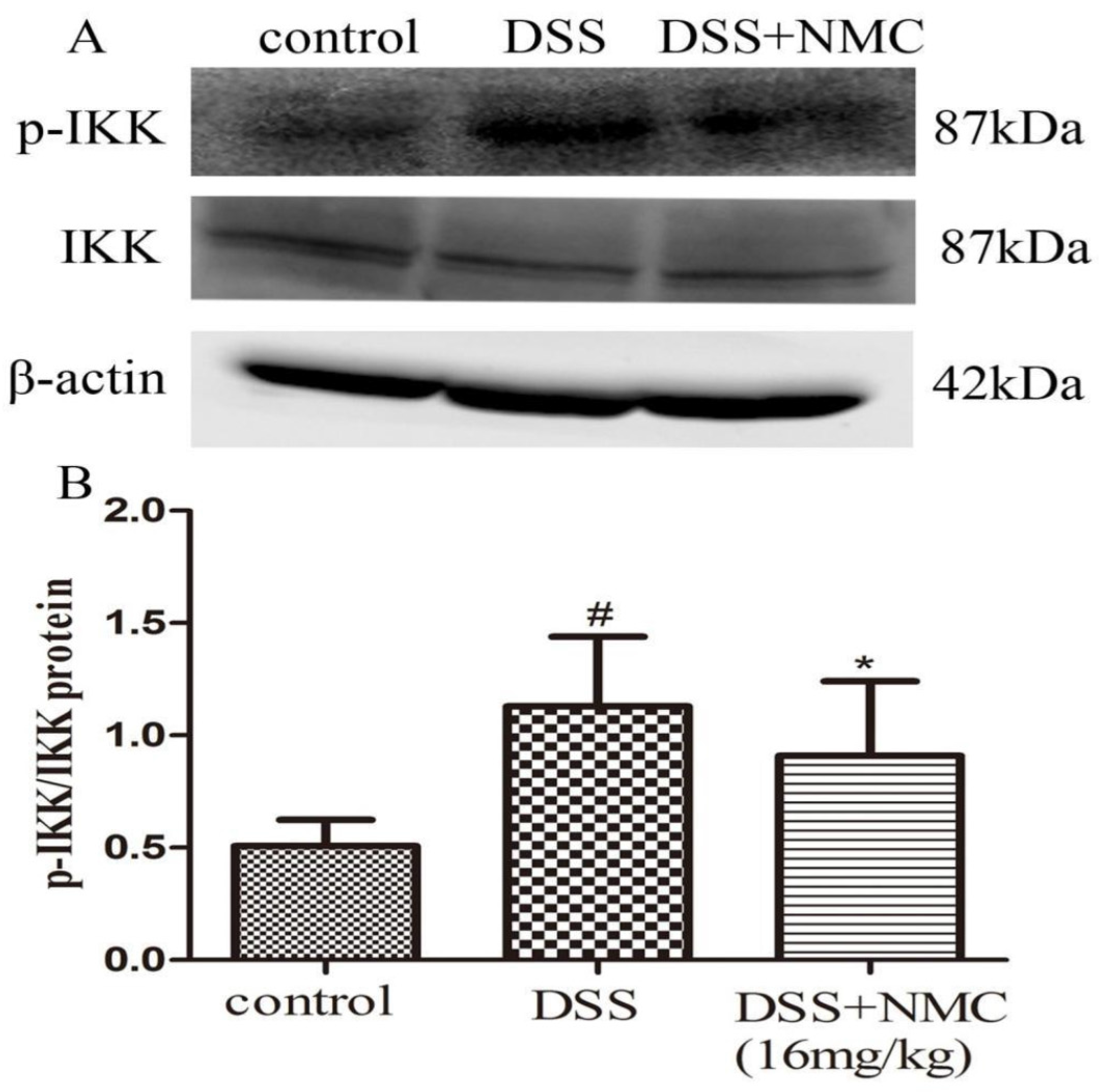
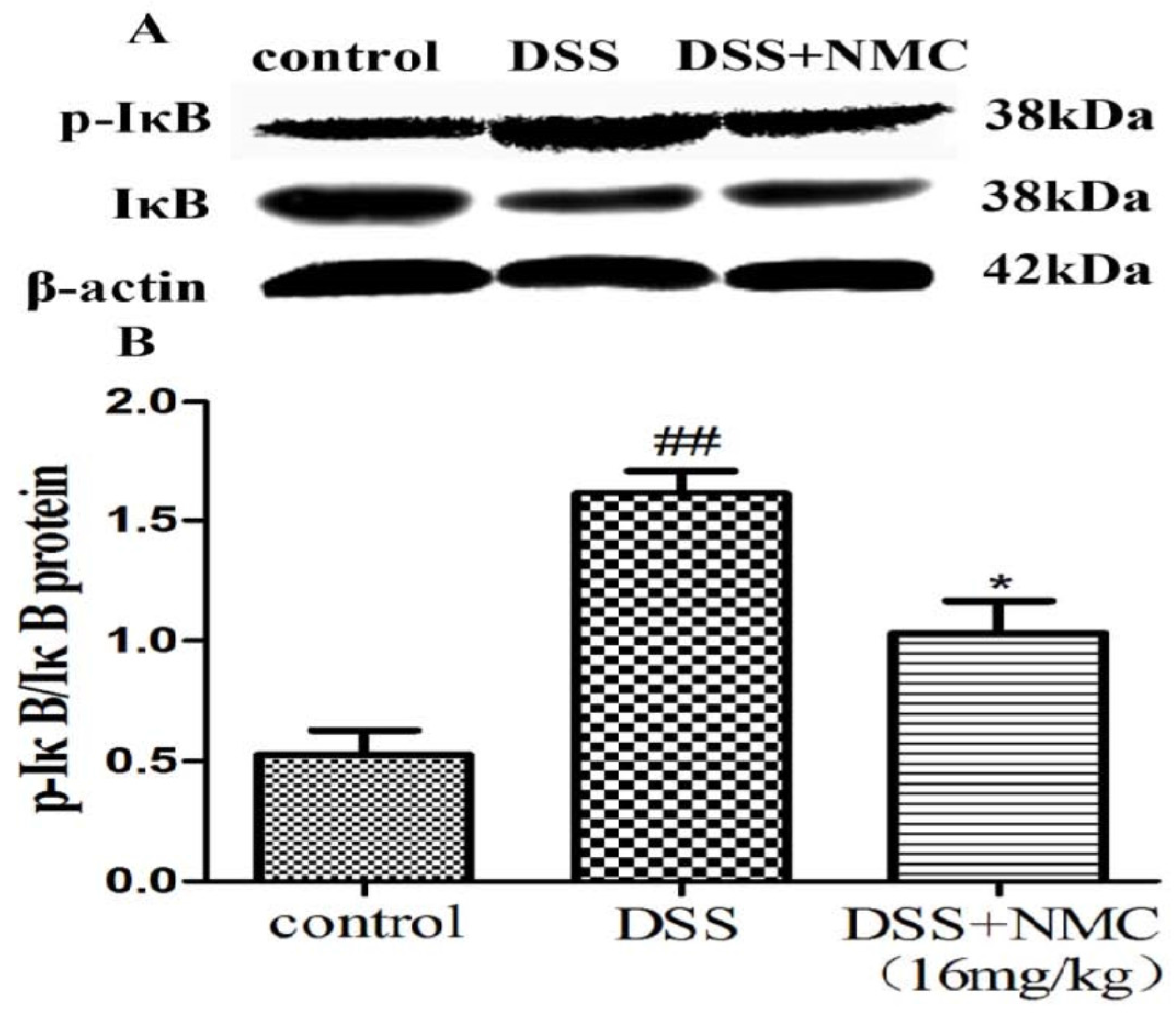
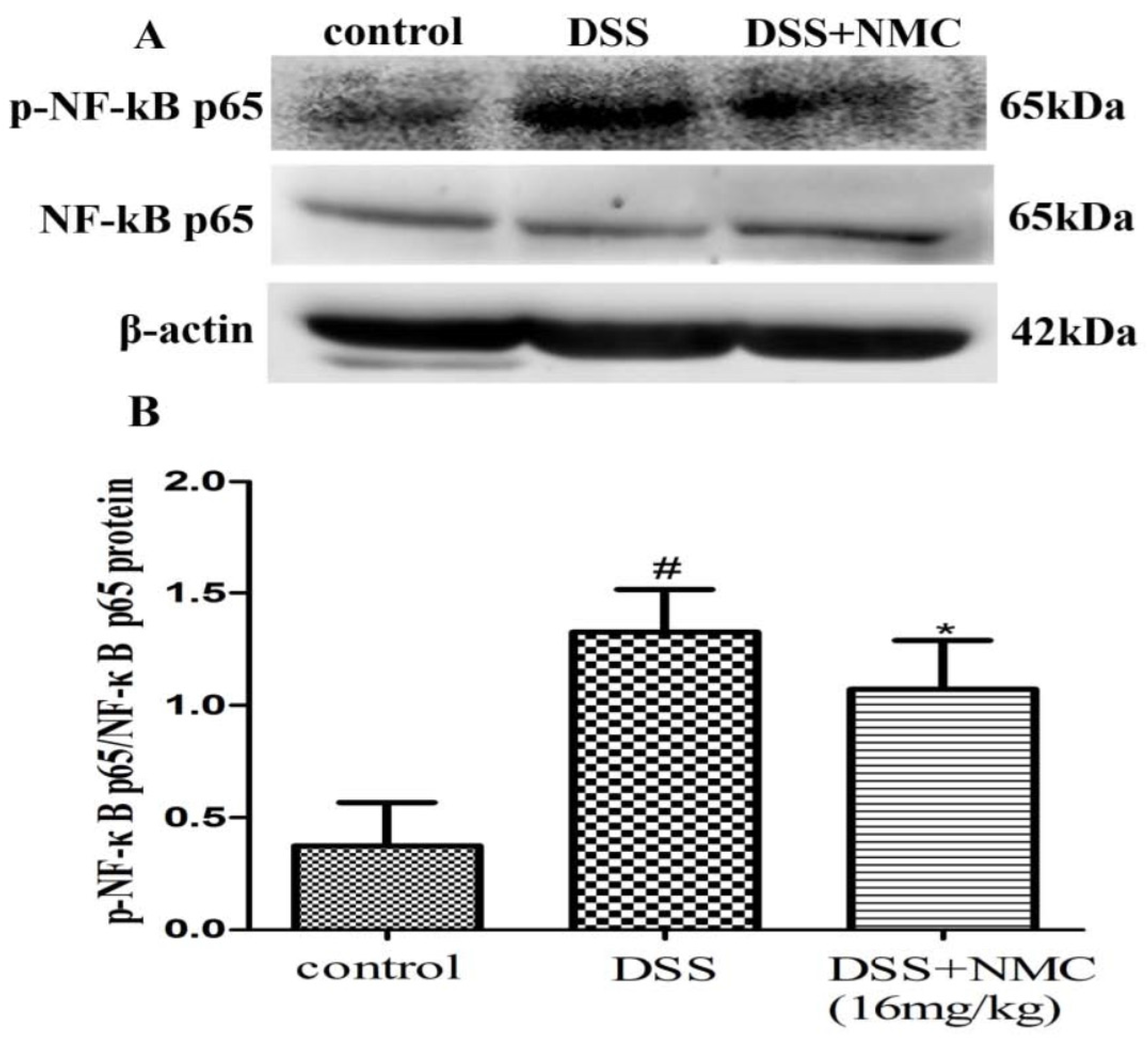
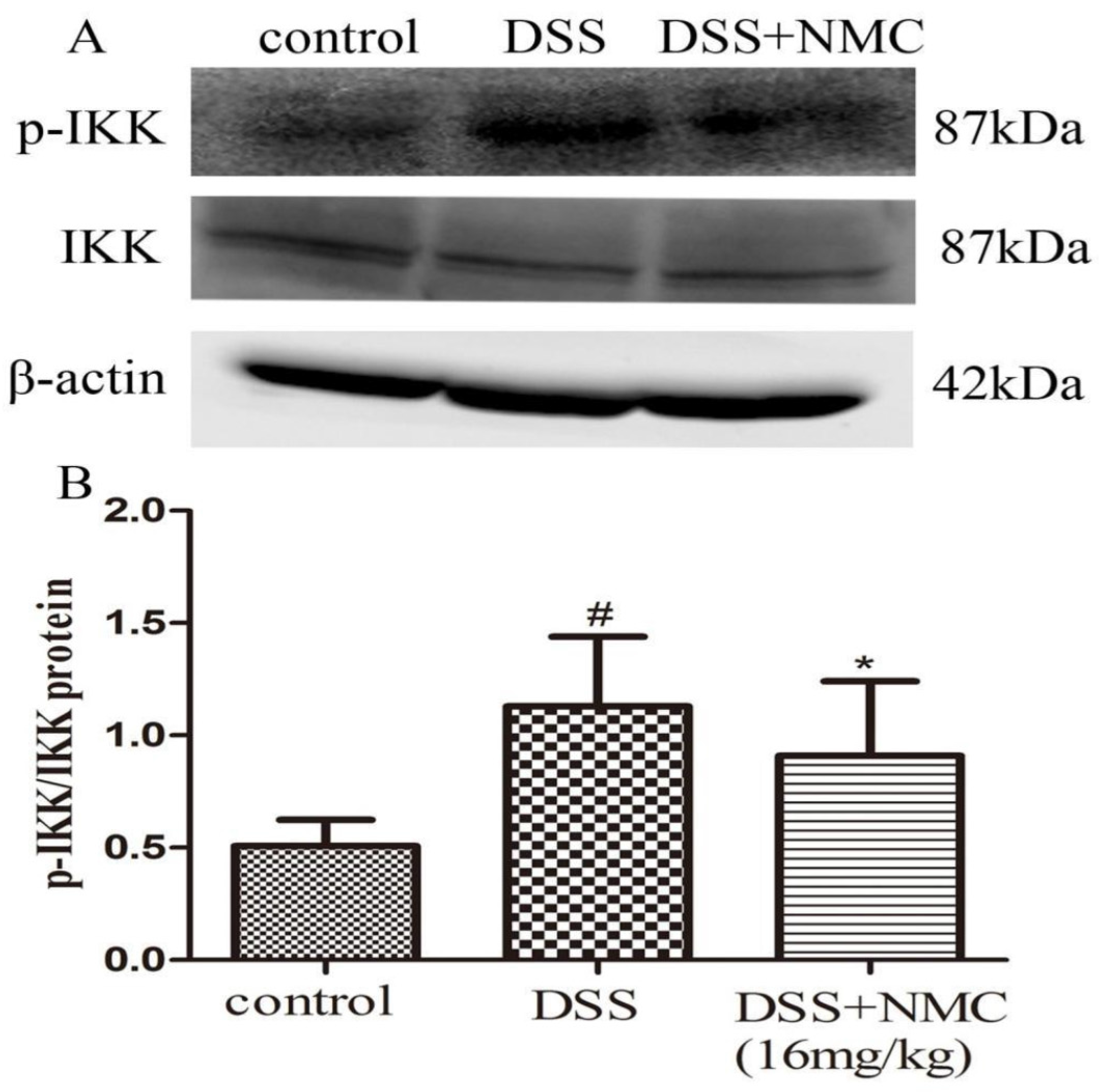
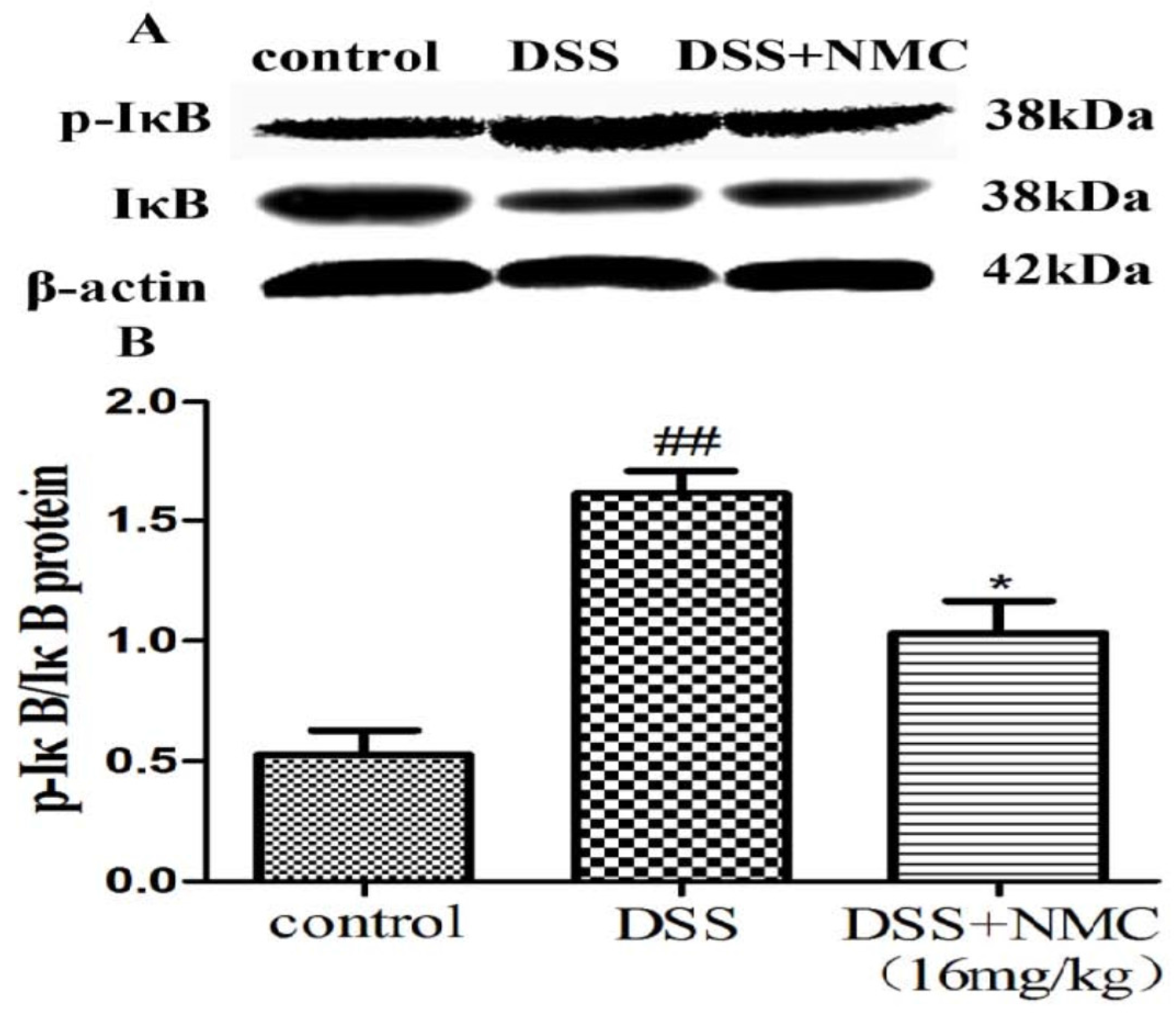
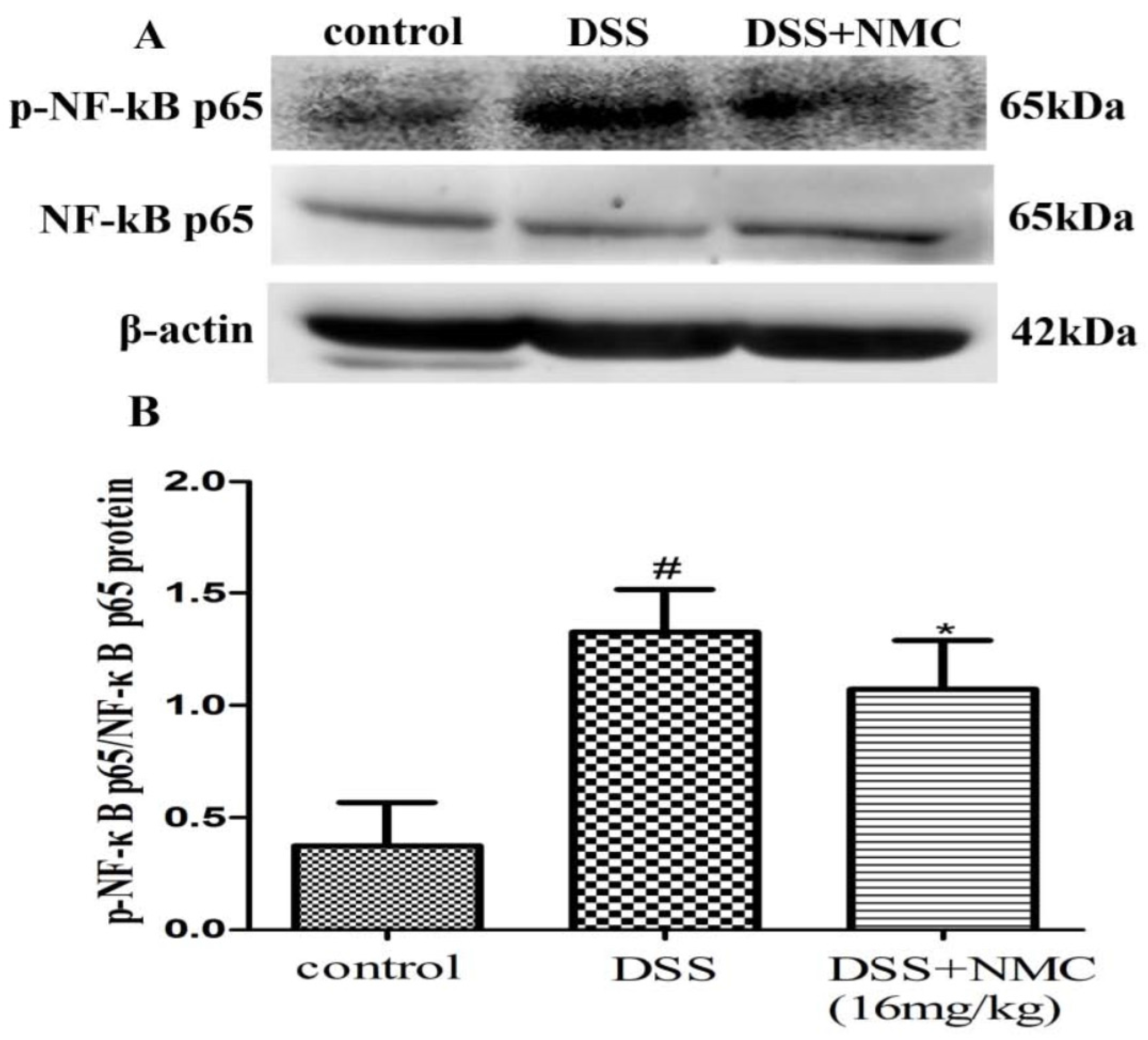
2.5. NMC Suppressed the Expression of p-NF-κB p65, IκB and IκB Kinase (IKK) in DSS-Induced Colitis
3. Discussion
4. Materials and Methods
4.1. Drugs and Reagents
4.2. Animals
4.3. Induction of Colitis and Experimental Design
4.4. Assessment of Disease Severity
4.5. Histopathological Evaluation of Colon
4.6. Myeloperoxidase (MPO) Activity Assay
4.7. Cytokine Analysis by ELISA
4.8. Western Blot Analysis
4.9. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Abraham, B.P.; Ahmed, T.; Ali, T. Inflammatory Bowel Disease: Pathophysiology and Current Therapeutic Approaches. Curr. Pharm. Des. 2017, 239, 115–146. [Google Scholar]
- Bae, S.I.; Kim, Y.S. Colon cancer screening and surveillance in inflammatory bowel disease. Clin. Endosc. 2014, 47, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Pasadhika, S.; Rosenbaum, J.T. Crohn’s Disease and Ulcerative Colitis. Intraocular Inflamm. 2016. [Google Scholar] [CrossRef]
- Loddo, I.; Romano, C. Inflammatory Bowel Disease: Genetics, Epigenetics, and Pathogenesis. Front. Immunol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Abraham, C.; Ruslan, M. Interactions between the host innate immune system and microbes in inflammatory bowel disease. Gastroenterology 2011, 140, 1729–1737. [Google Scholar] [CrossRef] [PubMed]
- De Zoeten, E.F.; Fuss, I.J. Cytokines and inflammatory Bowel Disease. Pediatr. Inflamm. Bowel Dis. 2013. [Google Scholar] [CrossRef]
- Dijkstra, G.; Moshage, H.; Van Dullemen, H.M.; de Jager-Krikken, A.; Tiebosch, A.T.; Kleibeuker, J.H.; Jansen, P.L.; van Goor, H. Expression of nitric oxide synthases and formation of nitrotyrosine and reactive oxygen species in inflammatory bowel disease. J. Pathol. 1998, 86, 416–421. [Google Scholar] [CrossRef]
- Shin, E.K.; Kwon, H.-S.; Kim, Y.H.; Shin, H.-K.; Kim, J.-K. Chrysin, a natural flavone, improves murine inflammatory bowel diseases. Biochem. Biophys. Res. Commun. 2009, 381, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Perše, M.; Cerar, A. Dextran Sodium Sulphate Colitis Mouse Model: Traps and Tricks. J. Biomed. Biotechnol. 2012, 2012, 718617. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Berstadb, A. Experimental colitis in animal models. Scand. J. Gastroenterol. 1992, 27, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Waldner, M.J.; Neurath, M.F. Chemically induced mouse models of colitis. Curr. Protoc. Pharmacol. 2009, 5, 1–15. [Google Scholar]
- Dothel, G.; Vasina, V.; Barbara, G.; De Ponti, F. Animal models of chemically induced intestinal inflammation: Predictivity and ethical issues. Pharmacol. Ther. 2013, 139, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Low, D.; Nguyen, D.D.; Mizoguchi, E. Animal models of ulcerative colitis and their application in drug research. Drug Des. Devel. Ther. 2013, 7, 1341–1357. [Google Scholar] [PubMed]
- Sha, T.; Igaki, K.; Yamasaki, M.; Watanabe, T.; Tsuchimori, N. Establishment and validation of a new semi-chronic dextran sulfate sodium-induced model of colitis in mice. Int. Immunopharm. 2013, 15, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Hall, L.J.; Faivre, E.; Quinlan, A.; Shanahan, F.; Nally, K.; Melgar, S. Induction and activation of adaptive immune populations during acute and chronic phases of a murine model of experimental colitis. Dig. Dis. Sci. 2011, 56, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Lautenschläger, C.; Schmidt, C.; Fischer, D.; Stallmach, A. Drug delivery strategies in the therapy of inflammatory bowel disease. Adv. Drug Deliv. Rev. 2014, 71, 58–76. [Google Scholar] [CrossRef] [PubMed]
- Boido, C.C.; Tasso, B.; Boido, V.; Sparatore, F. Cytisine derivatives as ligands for neuronal nicotine receptors and with various pharmacological activities. Il Farm. 2003, 58, 265–277. [Google Scholar] [CrossRef]
- Vakhitova, Y.V.; Farafontova, E.I.; Zainullina, L.F.; Vakhitov, V.A.; Tsypysheva, I.P.; Yunusov, M.S. Search for (–)-Cytisine Derivatives As Potential Inhibitors of NF-κB and STAT1. Rus. J. Bioorgan. Chem. 2015, 41, 297–304. [Google Scholar] [CrossRef]
- Okayasu, I.; Hatakeyama, S.; Yamada, M.; Ohkusa, T.; Inagaki, Y.; Nakaya, R. A novel method in the induction of reliable experimental acute and chronic ulcerative colitis in mice. Gastroenterology 1990, 98, 694–702. [Google Scholar] [CrossRef]
- Pandurangan, A.K.; Ismail, S.; Saadatdoust, Z.; Esa, N.M. Allicin Alleviates Dextran Sodium Sulfate- (DSS-) Induced Ulcerative Colitis in BALB/c Mice. Oxid. Med. Cell. Longev. 2015. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Hong, T.; Dong, M.; Meng, Y.; Mu, J. Protective effect of myricetin in dextran sulphate sodium-induced murine ulcerative colitis. Mol. Med. Rep. 2013, 7, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Atreya, I.; Atreya, R.; Neurath, M.F. NF-κB in inflammatory bowel disease. J. Int. Med. 2008, 263, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Kiron, M.; Das, M.D.; Biancone, L. Is IBD an Autoimmune Disorder? Inflamm. Bowel Dis. 2006, 14, S97–S101. [Google Scholar]
- Faubion, W.A., Jr.; Loftus, E.V., Jr.; Harmsen, W.S.; Zinsmeister, A.R.; Sandborn, W.J. The natural history of corticosteroid therapy for inflammatory bowel disease: A population-based study. Gastroenterology 2001, 121, 255–260. [Google Scholar] [CrossRef]
- Meier, J.; Sturm, A. Current treatment of ulcerative colitis. World J. Gastroenterol. 2011, 17, 3204–3212. [Google Scholar] [PubMed]
- Beaugerie, L.; Brousse, N.; Bouvier, A.M.; Colombel, J.F.; Lemann, M.; Cosnes, J.; Hébuterne, X.; Cortot, A.; Bouhnik, Y.; Gendre, J.P.; et al. Lymphoproliferative disorders in patients r,eceiving thiopurines for inflammatory bowel disease: A prospective observational cohort study. Lancet 2009, 374, 1617–1625. [Google Scholar] [CrossRef]
- Campregher, C.; Gasche, C. Aminosalicylates. Best Pract. Res. Clin. Gastroenterol. 2011, 25, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Maloy, K.J.; Powrie, F. Intestinal homeostasis and its breakdown in inflammatory bowel disease. Nature 2011, 474, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Li, Y.R. Oxidative stress and redox signaling mechanisms of inflammatory bowel disease: Updated experimental and clinical evidence. Exp. Biol. Med. 2012, 237, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Matuszyk, A.; Ceranowicz, P.; Warzecha, Z.; Cieszkowski, J.; Ceranowicz, D.; Gałazka, K.; Bonior, J.; Jaworek, J.; Bartus, K.; Gil, K.; et al. Exogenous Ghrelin Accelerates the Healing of Acetic Acid-Induced Colitis in Rats. Int. J. Mol. Sci. 2016, 17, 1455. [Google Scholar] [CrossRef] [PubMed]
- Fina, D.; Pallone, F. What Is the Role of Cytokines and Chemokines in IBD? Pathophysiology 2008, 14, S117–S118. [Google Scholar]
- Asakura, H. Proinflammatory cytokines in IBD. J. Gastroenterol. 1999, 34, 149–151. [Google Scholar] [CrossRef] [PubMed]
- Biasi, F.; Leonarduzzi, G.; Oteiza, P.I.; Poli, G. Inflammatory bowel disease: Mechanisms, redox considerations, and therapeutic targets. Antioxid. Redox Signal. 2013, 19, 1711–1747. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Muñoz, F.; Dominguez-Lopez, A.; Yamamoto-Furusho, J.K. Role of cytokines in inflammatory bowel disease. World J. Gastroenterol. 2008, 14, 4280–4288. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Lin, Q.; Yang, T.; Zeng, L.; Shi, L.; Chen, Y.; Luo, F. Oat β-glucan ameliorates dextran sulfate sodium (DSS)-induced ulcerative colitis in mice. Food Funct. 2015, 6, 3454–3463. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Liu, W.; Jin, B.; Geng, J.; Li, J.; Ding, H.; Wu, X.; Xu, Q.; Sun, Y.; Gao, J. Asiatic acid ameliorates dextran sulfate sodium-induced murine experimental colitis via suppressing mitochondria-mediated NLRP3 inflammasome activation. Int. Immunopharm. 2015, 24, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Fichtner-Feigl, S.; Fuss, I.J.; Preiss, J.C.; Strober, W.; Kitani, A. Treatment of murine Th1- and Th2-mediated inflammatory bowel disease with NF-kappa B decoy oligonucleotides. J. Clin. Investig. 2005, 115, 3057–3071. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.L.; Fan, H.Y.; Yang, M.Y.; Zhang, Z.K.; Liu, K. Therapeutic effect of a hydroxynaphthoquinone fraction on dextran sulfate sodium-induced ulcerative colitis. World J. Gastroenterol. 2014, 20, 15310–15318. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, A.S., Jr. The NF-κB and IκB proteins: New discoveries and insights. Annu. Rev. Immunol. 1996, 14, 649–681. [Google Scholar] [CrossRef] [PubMed]
- La Ferla, K.; Seegert, D.; Schreiber, S. Activation of NF-kB in intestinal epitheial cells by E. coli strains isolated from the colonic mucosa of IBD patients. Int. J. Colorectal Dis. 2004, 19, 334–342. [Google Scholar] [PubMed]
- Tsang, S.W.; Ip, S.P.; Wu, J.C.Y.; Ng, S.C.; Yung, K.K.L.; Bian, Z.X. A Chinese medicinal formulation ameliorates dextran sulfate sodium-induced experimental colitis by suppressing the activity of nuclear factor-kappaB signaling. J. Ethnopharmacol. 2015, 162, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.D.; Kang, S.H.; Bang, K.S.; Chang, Y.N.; Lee, J.H.; Jin, J.S. Glycyrrhetic Acid Ameliorates Dextran Sulfate Sodium-Induced Ulcerative Colitis in Vivo. Molecules 2016, 21, 523. [Google Scholar] [CrossRef] [PubMed]
- Cooper, H.S.; Murphy, S.N.; Shah, R.S.; Sedergran, D.J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Investig. 1993, 69, 238–249. [Google Scholar] [PubMed]
- Kim, H.; Im, J.P.; Kim, J.S.; Kang, J.S.; Lee, W.J. Alloferon Alleviates Dextran Sulfate Sodium-induced Colitis. Immune Netw. 2015, 15, 135–141. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphological Changes | ||||
|---|---|---|---|---|
| Groups | Inflammation Severity (0–3) | Injury Extent (0–3) | Crypt Damage (0–4) | Involved Colon (0–4) |
| Control | 0 | 0 | 0 | 0 |
| DSS | 2–3 | 3 | 3 | 2 |
| 5-ASA+DSS | 2 | 1–2 | 2 | 1 |
| NMC(1 mg/kg)+DSS | 3 | 2–3 | 3 | 2 |
| NMC(4 mg/kg)+DSS | 1–2 | 2 | 2 | 1 |
| NMC(16 mg/kg)+DSS | 1 | 2 | 1 | 1 |
| Score | Weight Loss (%) | Stool Consistency | Occult/Gross Bleeding |
|---|---|---|---|
| 0 1 2 3 4 | None 1–5 5–10 10–20 >20 | Normal Loose stool Diarrhea | Negative Positive Gross bleeding |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiao, Y.-F.; Lu, M.; Zhao, Y.-P.; Liu, N.; Niu, Y.-T.; Niu, Y.; Zhou, R.; Yu, J.-Q. N-Methylcytisine Ameliorates Dextran-Sulfate-Sodium-Induced Colitis in Mice by Inhibiting the Inflammatory Response. Molecules 2018, 23, 510. https://doi.org/10.3390/molecules23030510
Jiao Y-F, Lu M, Zhao Y-P, Liu N, Niu Y-T, Niu Y, Zhou R, Yu J-Q. N-Methylcytisine Ameliorates Dextran-Sulfate-Sodium-Induced Colitis in Mice by Inhibiting the Inflammatory Response. Molecules. 2018; 23(3):510. https://doi.org/10.3390/molecules23030510
Chicago/Turabian StyleJiao, Yan-Fang, Min Lu, Yu-Ping Zhao, Ning Liu, Ya-Ting Niu, Yang Niu, Ru Zhou, and Jian-Qiang Yu. 2018. "N-Methylcytisine Ameliorates Dextran-Sulfate-Sodium-Induced Colitis in Mice by Inhibiting the Inflammatory Response" Molecules 23, no. 3: 510. https://doi.org/10.3390/molecules23030510
APA StyleJiao, Y.-F., Lu, M., Zhao, Y.-P., Liu, N., Niu, Y.-T., Niu, Y., Zhou, R., & Yu, J.-Q. (2018). N-Methylcytisine Ameliorates Dextran-Sulfate-Sodium-Induced Colitis in Mice by Inhibiting the Inflammatory Response. Molecules, 23(3), 510. https://doi.org/10.3390/molecules23030510