
Continuous Gradient Temperature Raman Spectroscopy of Fish Oils Provides Detailed Vibrational Analysis and Rapid, Nondestructive Graphical Product Authentication


Abstract
1. Introduction
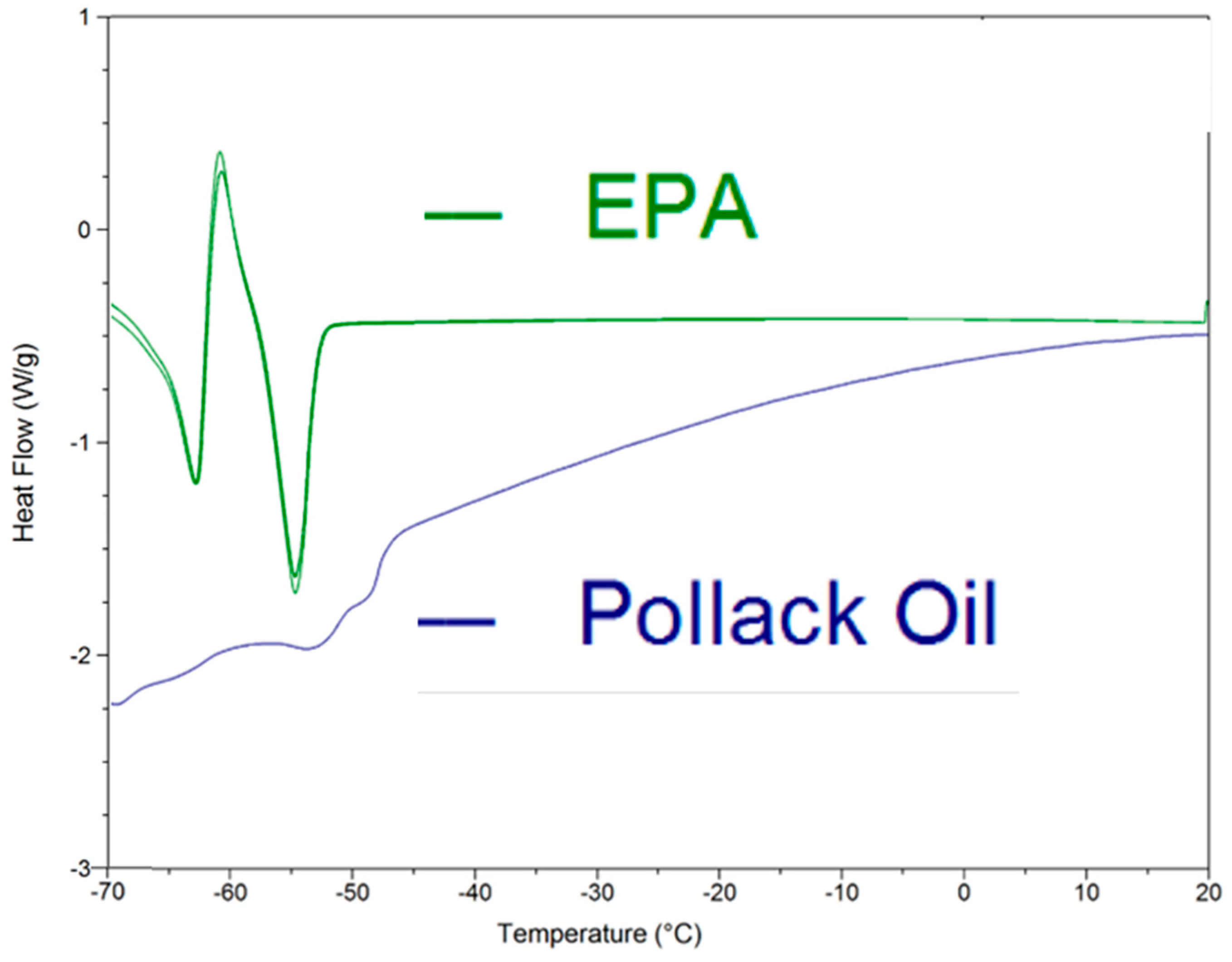
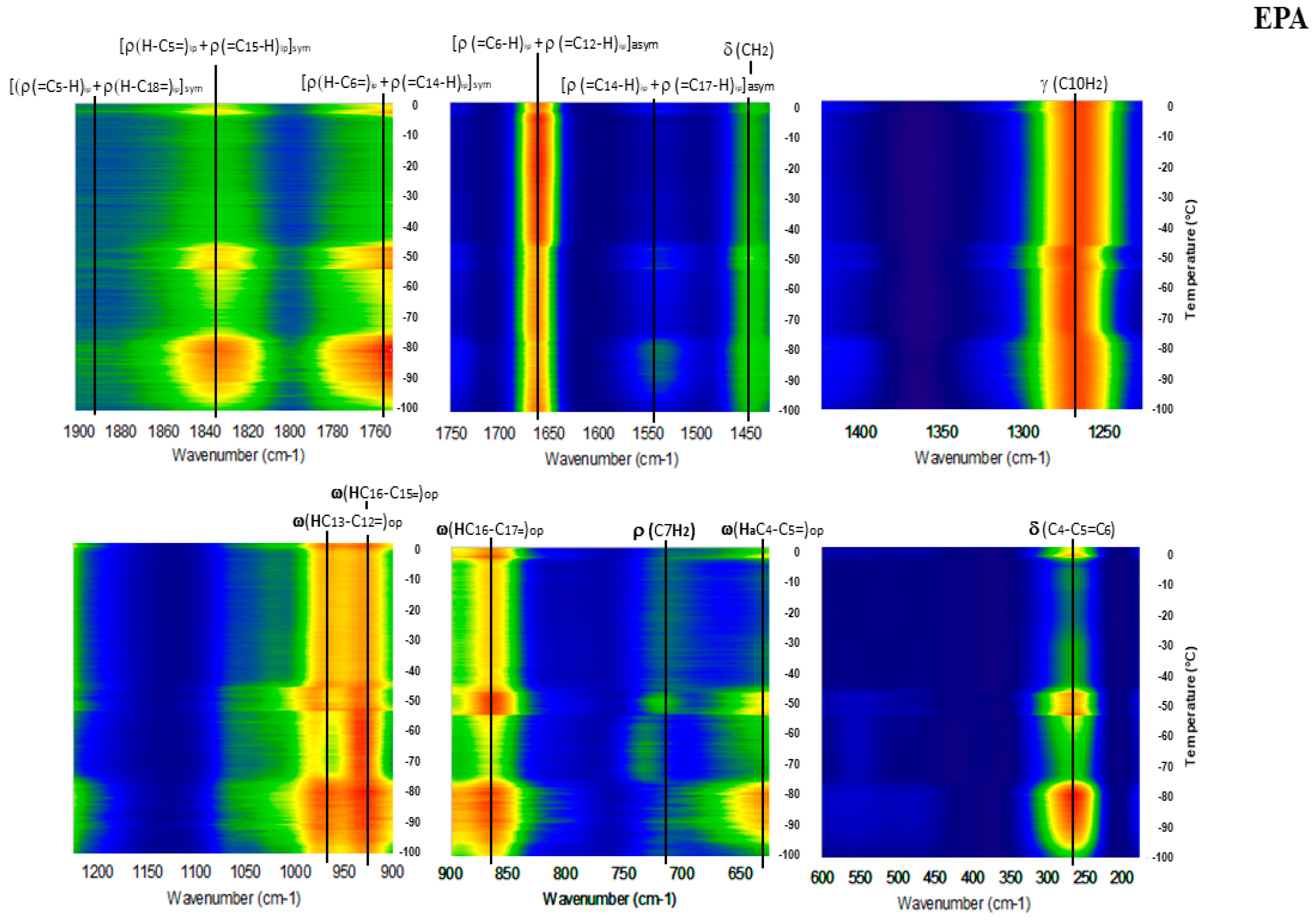
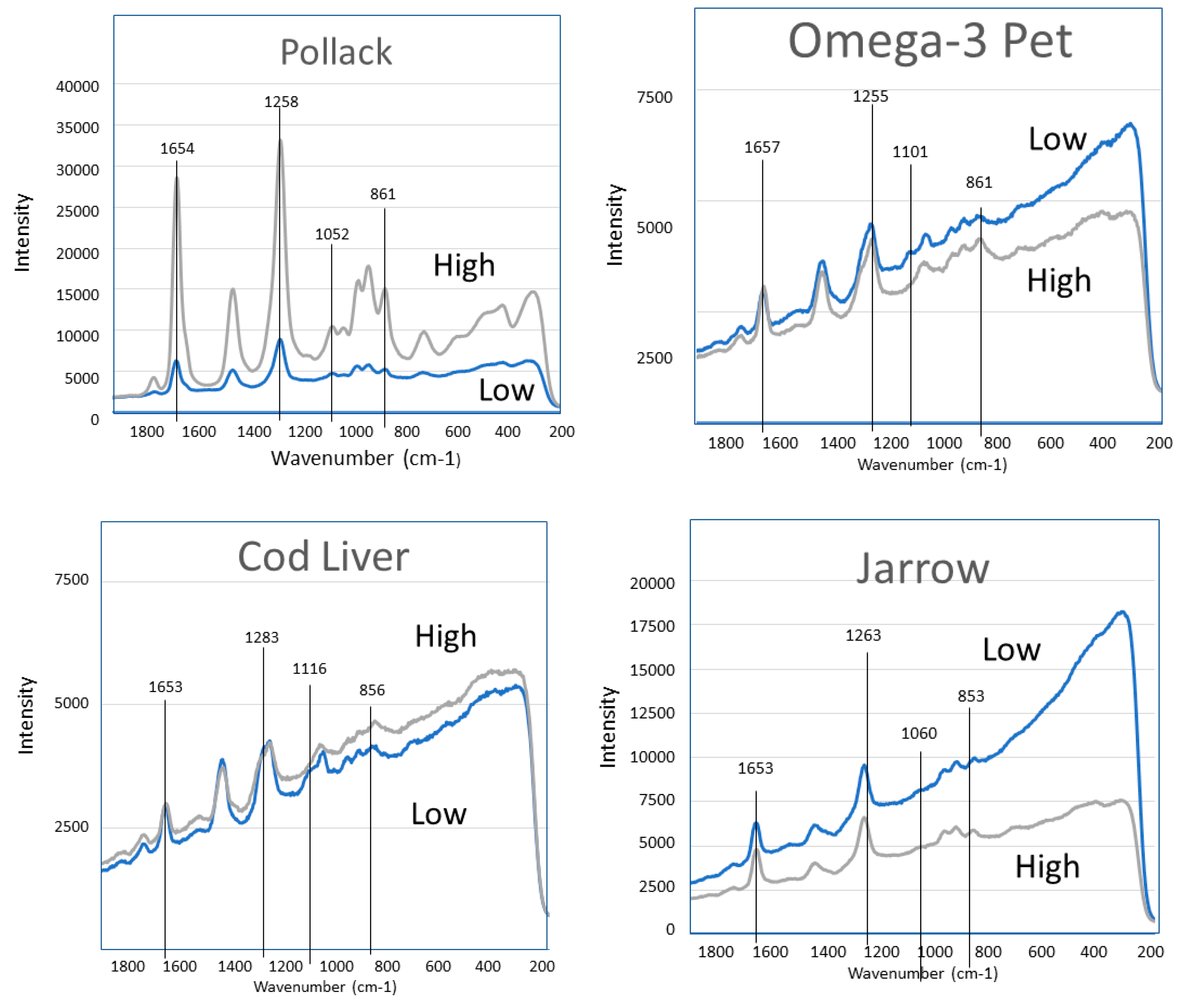
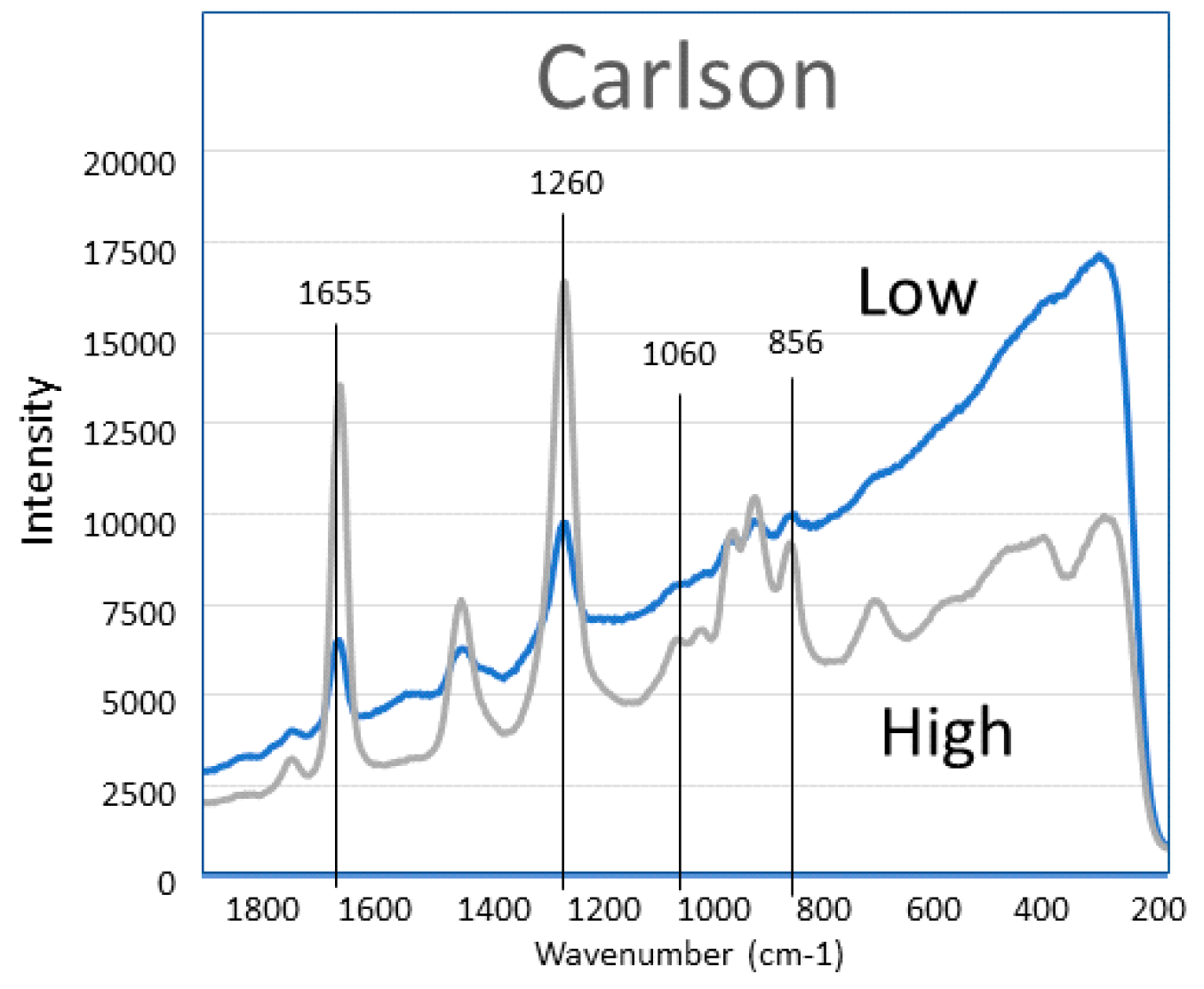
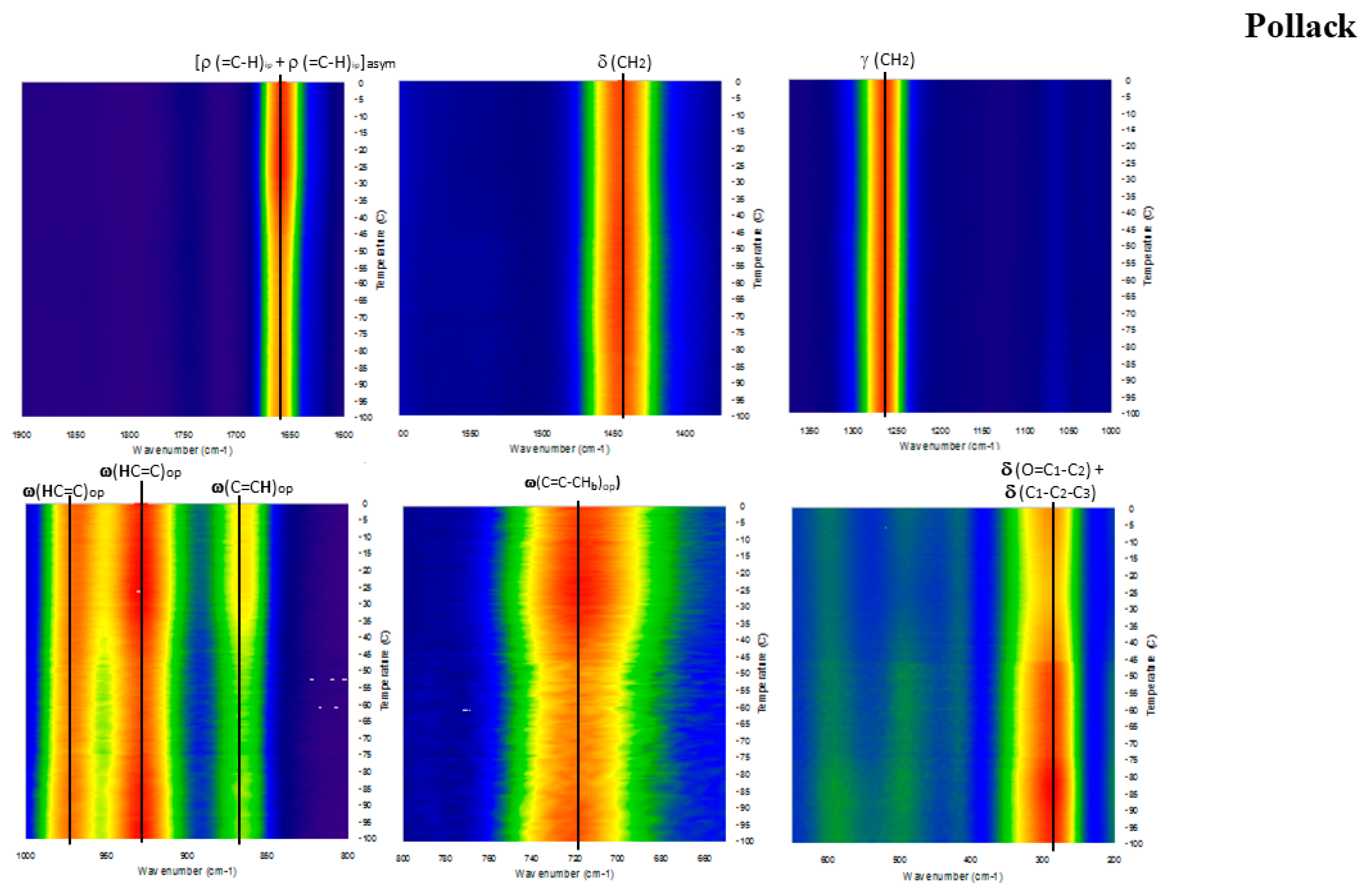
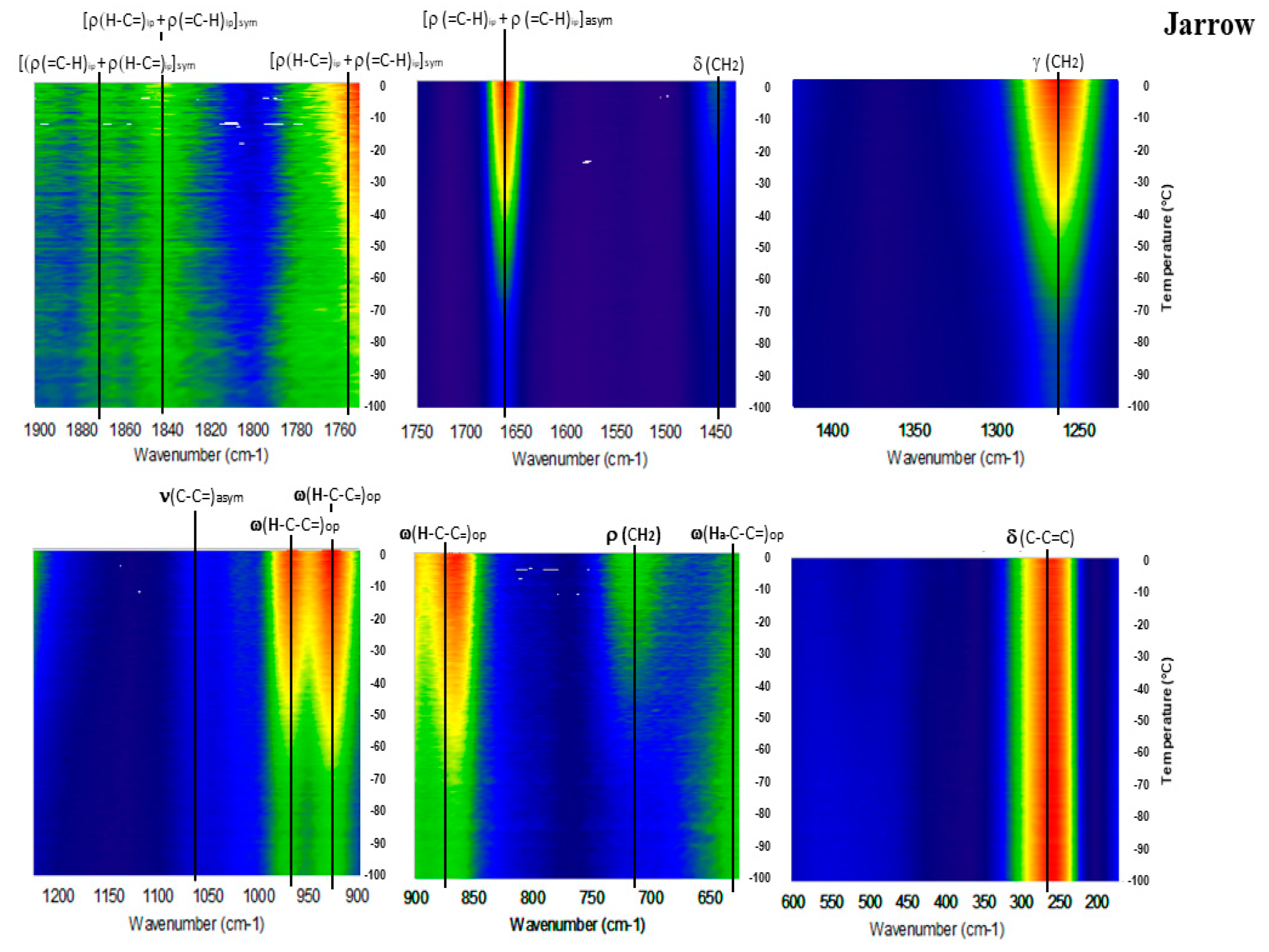
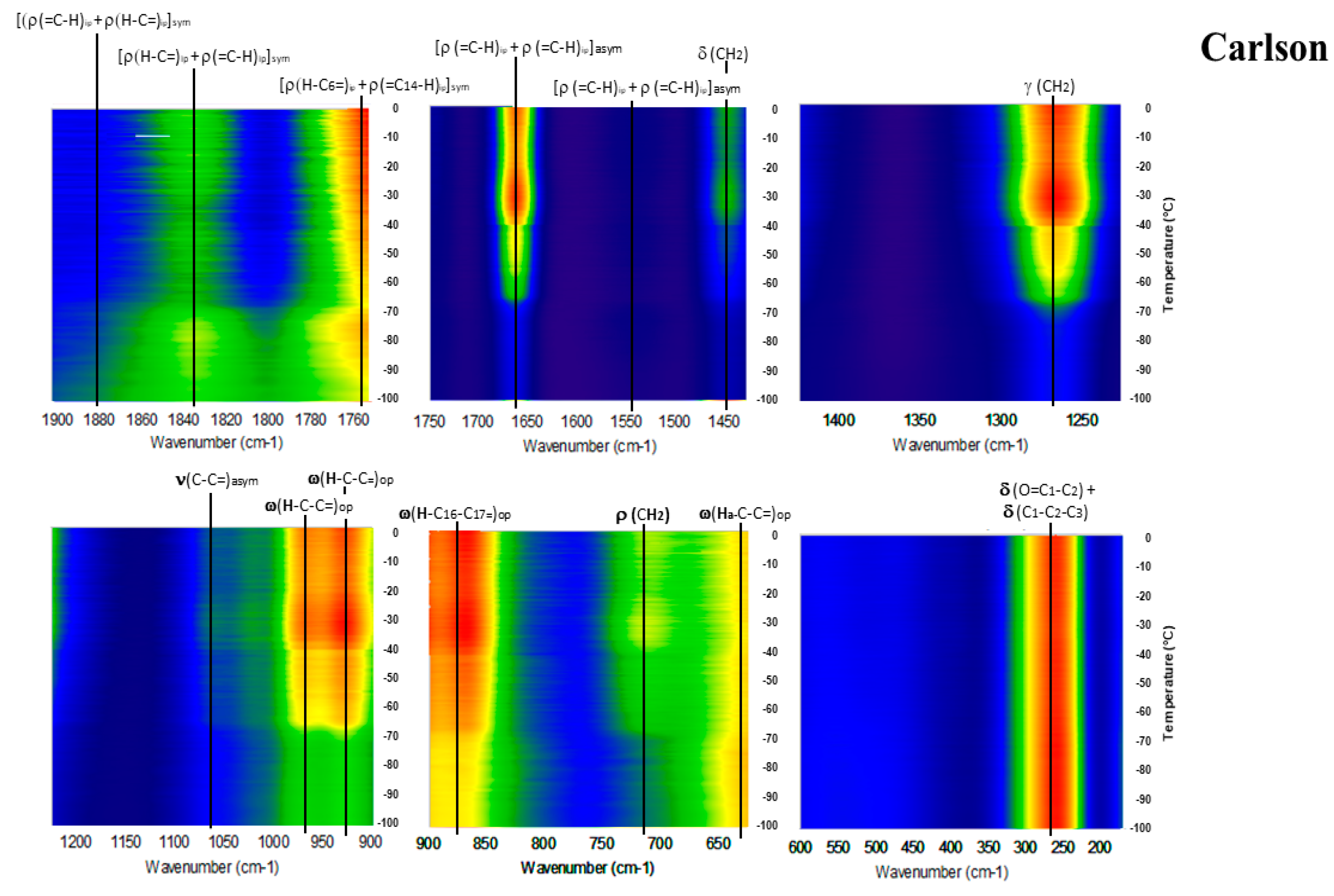
2. Results
3. Discussion
Detailed Spectroscopy of EPA and DHA
4. Materials and Methods
- Cis-5,8,11,14,17 Eicosapentaenoic acid 98%, Sigma-Aldrich Inc., St. Louis, MO, USA. (EPA)
- Wiley’s Finest Alaskan Pollack oil concentrate was donated by Wiley’s Finest Inc. Coshocton, OH, USA. Liquid form, specified as >90% TAG. DHA 189 mg; EPA 289 mg; other omega-3 56 mg. (PO)
- Omega-3 Pet, Nordic Naturals, Inc. Watsonville, CA, USA; purchased from a retail vendor. Anchovy and sardine oil, liquid form, specified as TAG. DHA 100 mg; EPA 170 mg; other omega-3 40 mg. (O3P)
- Pet Cod Liver Oil, Nordic Naturals, Inc. Watsonville, CA, USA; purchased from a retail vendor. Wild Arctic Cod liver oil, liquid form, specified as TAG. DHA 120 mg; EPA 80 mg; other omega-3 50 mg. (CLO).
- Max DHA fish oil, purchased directly from Jarrow Formulas, Inc. Los Angeles, CA, USA. Encapsulated fish oil concentrated in DHA by molecular distillation. Unspecified fish source. DHA 600 mg; EPA and other omega-3 not listed but is within range 50–250 mg. (JW).
- Carlson Maximum Omega 2000 fish oil was donated by J.R. Carlson Laboratories, Arlington Heights, IL, USA. Encapsulated Norwegian fish oil concentrate from anchovy, sardine, mackerel. Reported (but not specified) as TAG. DHA 190 mg; EPA 480 mg; other omega-3 96 mg. (CN).
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gawrisch, K.; Soubias, O. Structure and dynamics of polyunsaturated hydrocarbon chains in lipid bilayers—significance for GPCR function. Chem. Phys. Lipids 2008, 153, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.C.; Niu, S.-L.; Litman, B.J. Quantifying the differential effects of DHA and DPA on the early events in visual signal transduction. Chem. Phys. Lipids 2012, 165, 393–400. [Google Scholar] [CrossRef]
- Janssen, C.I.; Kiliaan, A.J. Long-chain polyunsaturated fatty acids (LCPUFA) from genesis to senescence: The influence of LCPUFA on neural development, aging, and neurodegeneration. Prog. Lipid Res. 2014, 53, 1–17. [Google Scholar] [CrossRef]
- Crawford, M.A.; Broadhurst, C.L.; Cunnane, S.; Marsh, D.E.; Schmidt, W.F.; Brand, A.; Ghebremeskel, K. Nutritional armor in evolution: Docosahexaenoic acid as a determinant of neural, evolution and hominid brain development. Mil. Med. 2014, 179, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Brenna, J.T.; Carlson, S.E. Docosahexaenoic acid and human brain development: Evidence that a dietary supply is needed for optimal development. J. Hum. Evol. 2014, 77, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Crawford, M.A.; Thabet, M.; Wang, Y. An introduction to a theory on the role of π-electrons of docosahexaenoic acid in brain function. Oilseed Fat Crop Lipid 2018, 25. [Google Scholar] [CrossRef]
- Crawford, M.A.; Thabet, M.; Wang, Y.; Broadhurst, C.L.; Schmidt, W.F. A theory on the role of π-electrons of docosahexaenoic acid in brain Function. Oilseed Fat Crop Lipid 2018, 25. [Google Scholar] [CrossRef]
- Moriguchi, T.; Lim, S.-Y.; Greiner, R.; Lefkowitz, W.; Loewke, J.; Hoshiba, J.; Salem, N., Jr. Effects of an n-3-deficient diet on brain, retina, and liver fatty acyl composition in artificially reared rats. J. Lipid Res. 2004, 45, 1437–1445. [Google Scholar] [CrossRef]
- Niu, S.L.; Mitchell, D.C.; Lim, S.Y.; Wen, Z.M.; Kim, H.; Salem, N.; Litman, B.J. Reduced G protein-coupled signaling efficiency in retinal rod outer segments in response to n-3 fatty acid deficiency. J. Biol. Chem. 2004, 279, 31098–31104. [Google Scholar] [CrossRef]
- SanGiovanni, J.P.; Chew, E.Y. The role of omega-3 long-chain polyunsaturated fatty acids in health and disease of the retina. Prog. Retinal Eye Res. 2005, 24, 87–138. [Google Scholar] [CrossRef]
- Dyall, S.C.; Michael-Titus, A.T. Neurological benefits of omega-3 fatty acids. Neuromol. Med. 2008, 10, 219–235. [Google Scholar] [CrossRef] [PubMed]
- Olsen, E.F.; Rukke, E.-O.; Flåtten, A.; Isaksson, T. Quantitative determination of saturated, monounsaturated, and polyunsaturated fatty acids in pork adipose tissue with non-destructive Raman spectroscopy. Meat Sci. 2007, 76, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Bekhit, M.Y.; Grung, B.; Mjøs, S.A. Determination of omega-3 fatty acids in fish oil supplements using vibrational spectroscopy and chemometric methods. Appl. Spectrosc. 2014, 68, 1190–1200. [Google Scholar] [CrossRef] [PubMed]
- Toledo, D.A.; Roque, N.R.; Teixeira, L.; Milán-Garcés, E.A.; Carneiro, A.B.; Almeida, M.R.; Andrade, G.F.; Martins, J.S.; Pinho, R.R.; Freire-de-Lima, G.C.; et al. Lipid body organelles within the parasite Trypanosoma cruzi: A role for intracellular arachidonic acid metabolism. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Killeen, D.P.; Marshall, S.N.; Burgess, E.J.; Gordon, K.C.; Perry, N.B. Raman spectroscopy of fish oil capsules: Polyunsaturated fatty acid quantitation plus detection of ethyl esters and oxidation. J. Agric. Food Chem. 2017, 65, 3551–3558. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Han, L.; Yang, Z.; Xu, L.; Liu, X. Technical note: Characterization of lipid constitution in Fourier transform infrared spectra and spectroscopic discrimination of animal-derived feedstuffs from different species. J. Anim. Sci. 2017, 95, 2794–2800. [Google Scholar] [PubMed]
- Broadhurst, C.L.; Schmidt, W.F.; Kim, M.S.; Nguyen, J.K.; Qin, J.; Chao, K.; Bauchan, G.L.; Shelton, D.R. Continuous gradient temperature Raman spectroscopy of n-6DPA and DHA from −100 to 20 °C. Chem. Phys. Lipids 2016, 200, 1–10. [Google Scholar] [CrossRef]
- Broadhurst, C.L.; Schmidt, W.F.; Kim, M.S.; Nguyen, J.K.; Qin, J.; Chao, K.; Bauchan, G.L.; Shelton, D.R. Continuous gradient temperature Raman spectroscopy of oleic and linoleic acids from −100 to 50 °C. Lipids 2016, 51, 1289–1302. [Google Scholar] [CrossRef]
- Broadhurst, C.L.; Schmidt, W.F.; Nguyen, J.K.; Qin, J.; Chao, K.; Aubuchon, S.R.; Kim, M.S. Continuous gradient temperature Raman spectroscopy and differential scanning calorimetry of N-3DPA and DHA from −100 to 10 °C. Chem. Phys. Lipids 2017, 204, 94–102. [Google Scholar] [CrossRef]
- Broadhurst, C.L.; Schmidt, W.F.; Nguyen, J.K.; Qin, J.; Chao, K.; Kim, M.S. Continuous gradient temperature Raman spectroscopy from −100 to 40 °C yields new molecular models of arachidonic acid and 2-arachidonoyl-1-stearoyl-sn-glycero-3-phosphocholine. Prostaglandins Leuk. Essent. Fat. Acids 2017, 127, 6–15. [Google Scholar] [CrossRef]
- Broadhurst, C.L.; Schmidt, W.F.; Nguyen, J.K.; Qin, J.; Chao, K.; Kim, M.S. Gradient temperature Raman spectroscopy of fatty acids with one to six double bonds identifies specific carbons and provides systematic three dimensional structures. J. Biophys. Chem. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Siddiqui, N.; Sim, J.; Silwood, C.J.L.; Toms, H.; Iles, R.I.; Grootveld, M. Multicomponent analysis of encapsulated marine oil supplements using high-resolution 1H and 13C NMR techniques. J. Lipid Res. 2003, 44, 2406–2427. [Google Scholar] [CrossRef] [PubMed]
- Aursand, M.; Standal, I.B.; Axelson, D.E. High-resolution 13C nuclear magnetic resonance spectroscopy pattern recognition of fish oil capsules. J. Agric. Food Chem. 2007, 55, 38–47. [Google Scholar] [CrossRef]
- Araujo, P.; Zhu, H.; Breivik, J.F.; Hjelle, J.I.; Zeng, Y. Determination and structural elucidation of TAGs in krill oil by chromatographic techniques. Lipids 2014, 49, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhou, H.; Zhang, Y.; Shen, Y.; Su, H.; Jin, J.; Jin, Q.; Wang, X. Characterization of positional distribution of fatty acids and TAG molecular composition of marine fish oils rich in omega-3 polyunsaturated fatty acids. BioMed Res. Int. 2018. [Google Scholar] [CrossRef]
- Karunathilaka, S.R.; Mossoba, M.M.; Chung, J.K.; Haile, E.A.; Srigley, C.T. Rapid prediction of fatty acid content in marine oil omega-3 dietary supplements using a portable Fourier transform infrared (FTIR) device and partial least-squares regression (PLSR) analysis. J. Agric. Food Chem. 2017, 65, 224–233. [Google Scholar] [CrossRef]
- Pi, F.; Kaneko, F.; Shinzawa, H.; Suzuki, M.; Iwahashi, M.; Ozaki, Y. Temperature dependence of structure and dynamic properties of oleic acid γ and α phases studied by FTIR spectroscopy. Bull. Chem. Soc. Japan 2011, 84, 403–412. [Google Scholar] [CrossRef]
- Chao, K.; Dhakal, S.; Qin, J.; Peng, Y.; Schmidt, W.F.; Kim, M.S.; Chan, D.E. A spatially offset Raman spectroscopy method for non-destructive detection of gelatin-encapsulated powders. Sensors 2017, 17, 618. [Google Scholar] [CrossRef]
- Rohman, A.; Man, Y.B.C. Analysis of cod liver oil adulteration using Fourier transform infrared (FTIR) spectroscopy. J. Am. Oil Chem. Soc. 2009, 86, 1149–1153. [Google Scholar] [CrossRef]
- Rohman, A.; Man, Y.B.C. Authentication analysis of cod liver oil from beef fat using fatty acid composition and FTIR spectra. Food Add. Contam. A 2011, 28, 1469–1474. [Google Scholar] [CrossRef]
- Inoue, T.; Hisatsugu, Y.; Yamamoto, R.; Suzuki, M. Solid–liquid phase behavior of binary fatty acid mixtures 1. Oleic acid/stearic acid and oleic acid/behenic acid mixtures. Chem. Phys. Lipids 2004, 127, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Bouzidi, L.; Narine, S.S. Relationships between molecular structure and kinetic and thermodynamic controls in lipid systems. Part III. Crystallization and phase behavior of 1-palmitoyl-2,3-stearoyl-sn-glycerol (PSS) and tristearoylglycerol (SSS) binary system. Chem. Phys. Lipids 2012, 165, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Bayés-García, L.; Calvet, T.; Cuevas-Diarte, M.À.; Ueno, S.; Sato, K. Phase behavior of binary mixture systems of saturated-unsaturated mixed-acid TAGs: Effects of glycerol structures and chain-chain interactions. J. Phys. Chem. B 2015, 119, 4417–4427. [Google Scholar] [CrossRef] [PubMed]
- Brenna, J.T.; Salem, N. Jr.; Sinclair, A.J.; Cunnane, S.C.; International Society for the Study of Fatty Acids and Lipids, ISSFAL. Alpha-linolenic acid supplementation and conversion to n-3 long-chain polyunsaturated fatty acids in humans. Prostag. Leukotr. Ess. 2009, 80, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Park, H.G.; Lawrence, P.; Engel, M.G.; Kothapalli, K.; Brenna, J.T. Metabolic fate of docosahexaenoic acid (DHA; 22:6n-3) in human cells: Direct retroconversion of DHA to eicosapentaenoic acid (20:5n-3) dominates over elongation to tetracosahexaenoic acid (24:6n-3). FEBS Lett. 2016, 590, 3188–3194. [Google Scholar] [CrossRef] [PubMed]
- Chilton, F.H.; Dutta, R.; Reynolds, L.M.; Sergeant, S.; Mathias, R.A.; Seeds, M.C. Precision nutrition and omega-3 polyunsaturated fatty acids: A case for personalized supplementation approaches for the prevention and management of human diseases. Nutrients 2017, 9, 1165. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Ogaki, T.; Sato, K. Crystallization and transformation mechanisms of α βand γ polymorphs of ultra-pure oleic acid. J. Am. Oil. Chem. Soc. 1985, 62, 1600–1604. [Google Scholar] [CrossRef]
- Ueno, S.; Miyazaki, A.; Yano, J.; Furukawa, Y.; Suzuki, M.; Sato, K. Polymorphism of linoleic acid (cis-9, cis-12-octadecadienoic acid) and α-linoleic acid (cis-9, cis-12, cis-15-octadecatrienoic acid). Chem. Phys. Lipids 2000, 107, 169–178. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DHA (cm−1) | EPA (cm−1) | Spectral Assignments | ||||
|---|---|---|---|---|---|---|
| −100 to −50 °C | −50 to −10 °C | −100 to −30 °C | −50 to −10 °C | |||
| DHA a,b | EPA a,b | |||||
| 1851 | 1849 | 1849 | [ρ(H-Ca=) + ρ(=Cb-H)]sym | 7,17 | 5,15 | |
| 1784 | 1782 | 1782 | [ρ(=Ca-H) + ρ(H-Cb=)]sym | 8,16 | 6,14 | |
| 1759 | 1757 | 1757 | [ρ(H-Ca=) + ρ(=Cb-H)]sym | 7,14 | 5,12 | |
| DHA a,b | EPA a,b | |||||
| 1698 | 1698 | 1698 | 1698 | [ρ(H-Ca=) + ρ(=Cb-H)]asym | 10,16 | 8,14 |
| 1644 | 1644 | 1647 | 1647 | [ρ(=Ca-H) + ρ(H-Cb=)]asym | 8,14 | 6,12 |
| 1565 | 1568 | 1568 | [ρ(=Ca-H) + ρ(H-Cb=)]asym | 16,19 | 14,17 | |
| 1478 | 1478 | 1481 | 1481 | [ρ(H-Ca=) + ρ(=Cb-H)]asym | 10,13 | 8,11 |
| 1437 | 1437 | 1437 | 1437 | δ(CH2) | ||
| DHA a | EPA a | |||||
| 1408 | 1392 | δ(CaH2) | 9 | 7 | ||
| 1341 | 1341 | 1338 | 1338 | γ(CaH2) | 12 | 10 |
| 1263 | 1263 | 1263 | γ(CaH2) | 9 | 7 | |
| 1234 | 1234 | 1235 | 1235 | γ(CaH2) | 15 | 13 |
| DHA a,b | EPA a,b | |||||
| 1094 | 1094 | 1088 | 1088 | ν(Ca-Cb=)asym | 21,20 | 19,18 |
| 1049 | 1049 | ν(Ca-Cb=)asym | 3,4 | |||
| 972 | 972 | ω(H-Ca-Cb=)op | 12,11 | 10,9 | ||
| 958 | 958 | 958 | ρ(CaH2) | 9 | 7 | |
| 914 | 914 | 914 | 914 | ω(H-Ca-Cb=)op | 6,7 | 4,5 |
| DHA a,b | EPA a,b | |||||
| 851 | 851 | 851 | 851 | ω(=Ca-CbHα)op | 14,15 | 12,13 |
| 827 | 827 | ν(Ca-Cb=)asym | 18,19 | 16,17 | ||
| 754 | 754 | 754 | 754 | ρ(CaH2) | 12 | 12,11 |
| 715 | 715 | ω(HαCa-Cb=)op | 6,5 | 4,5 | ||
| 696 | 696 | 696 | ρ(CaH2) | 6 | 4 | |
| 651 | ω(H-C5=C6)op | 3,4 | 4 | |||
| DHA a,b,c | EPA a,b,c | |||||
| 563 | 544 | δ(Ca-Cb=Cc) + δ(Ca=Cb-Cc) | 12,11,10 + 8,7,6 | 10,9,8 + 6,5,4 | ||
| 464 | 464 | 473 | 473 | δ(CaCbCc) | 2,3,4 | 2,3,4 |
| 415 | 406 | 406 | δ(O=Ca-Cb) + δ(Ca-Cb=Cc) | 1,2 + 1,2,3 | 1,2 + 1,2,3 | |
| 265 | 265 | 252 | 252 | δ(Ca-Cb=Cc) | 3,4,5 | 4,5,6 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Broadhurst, C.L.; Schmidt, W.F.; Qin, J.; Chao, K.; Kim, M.S. Continuous Gradient Temperature Raman Spectroscopy of Fish Oils Provides Detailed Vibrational Analysis and Rapid, Nondestructive Graphical Product Authentication. Molecules 2018, 23, 3293. https://doi.org/10.3390/molecules23123293
Broadhurst CL, Schmidt WF, Qin J, Chao K, Kim MS. Continuous Gradient Temperature Raman Spectroscopy of Fish Oils Provides Detailed Vibrational Analysis and Rapid, Nondestructive Graphical Product Authentication. Molecules. 2018; 23(12):3293. https://doi.org/10.3390/molecules23123293
Chicago/Turabian StyleBroadhurst, C. Leigh, Walter F. Schmidt, Jianwei Qin, Kuanglin Chao, and Moon S. Kim. 2018. "Continuous Gradient Temperature Raman Spectroscopy of Fish Oils Provides Detailed Vibrational Analysis and Rapid, Nondestructive Graphical Product Authentication" Molecules 23, no. 12: 3293. https://doi.org/10.3390/molecules23123293
APA StyleBroadhurst, C. L., Schmidt, W. F., Qin, J., Chao, K., & Kim, M. S. (2018). Continuous Gradient Temperature Raman Spectroscopy of Fish Oils Provides Detailed Vibrational Analysis and Rapid, Nondestructive Graphical Product Authentication. Molecules, 23(12), 3293. https://doi.org/10.3390/molecules23123293