Evaluation of Strategies to Produce Highly Porous Cross-Linked Aggregates of Porcine Pancreas Lipase with Magnetic Properties


 and
and 
Abstract

1. Introduction
2. Results and Discussion
2.1. Influence of Co-Feeders in the Preparation of CLEAs
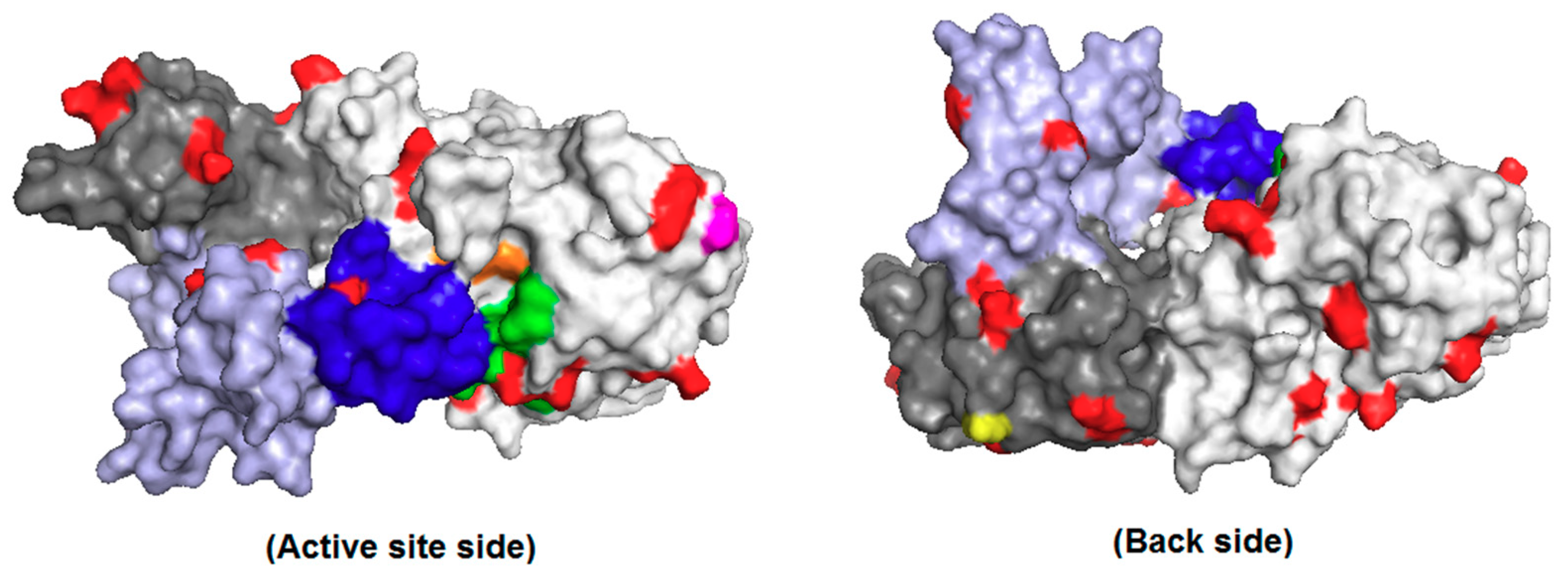
2.2. Treatment of PPL Surface
2.3. Preparation of Magnetic CLEAs Using Starch in the Aggregation and Crosslinking Steps
2.4. Influence of Time of Glutaraldehyde Treatment in the Preparation of PPL CLEAs
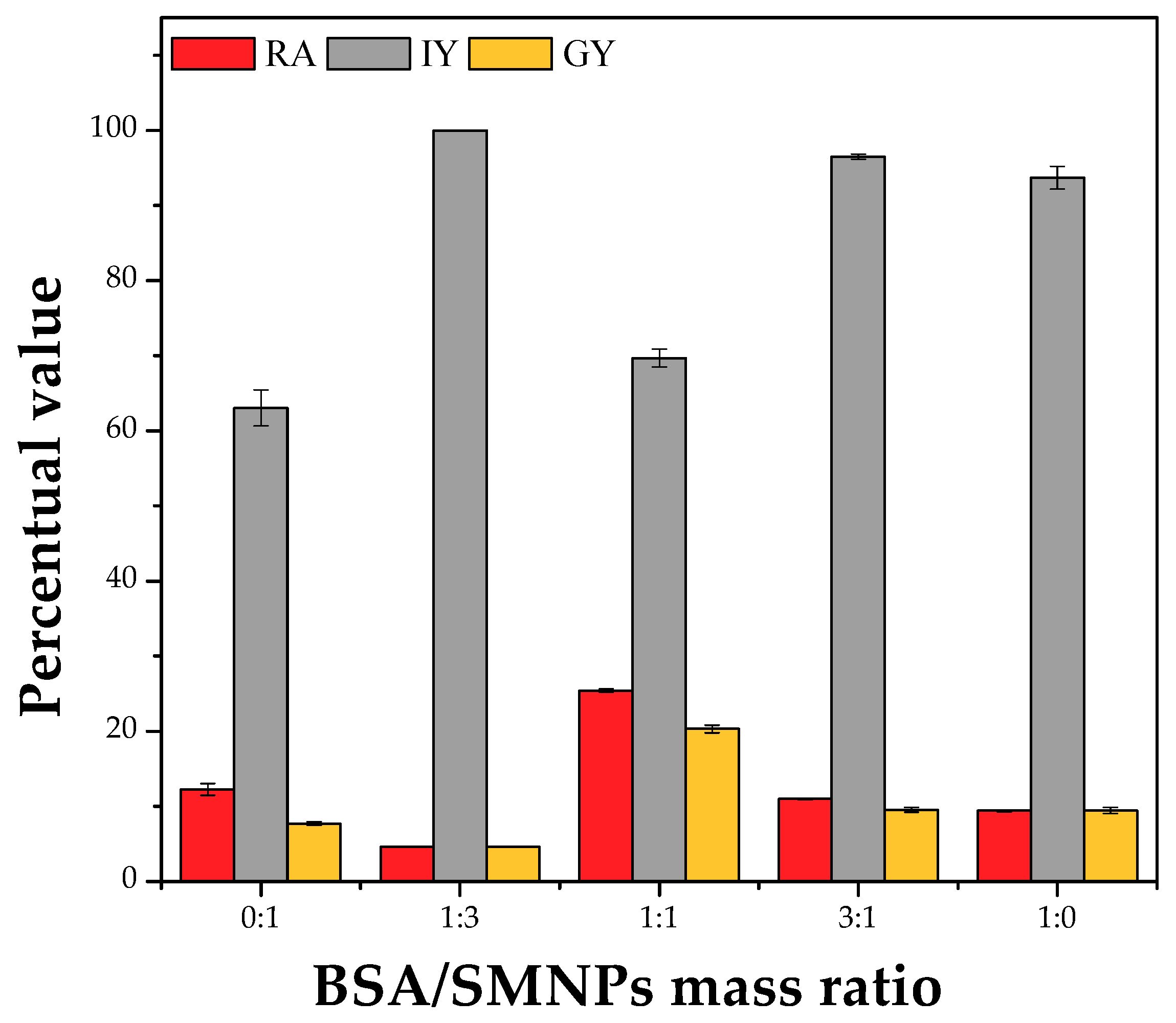
2.5. Selection of Additives for Preparation of PPL CLEAs Using 0.8% (w/v) Starch as Pore-Making Agent
2.6. Characterization of Free and Immobilized PPL
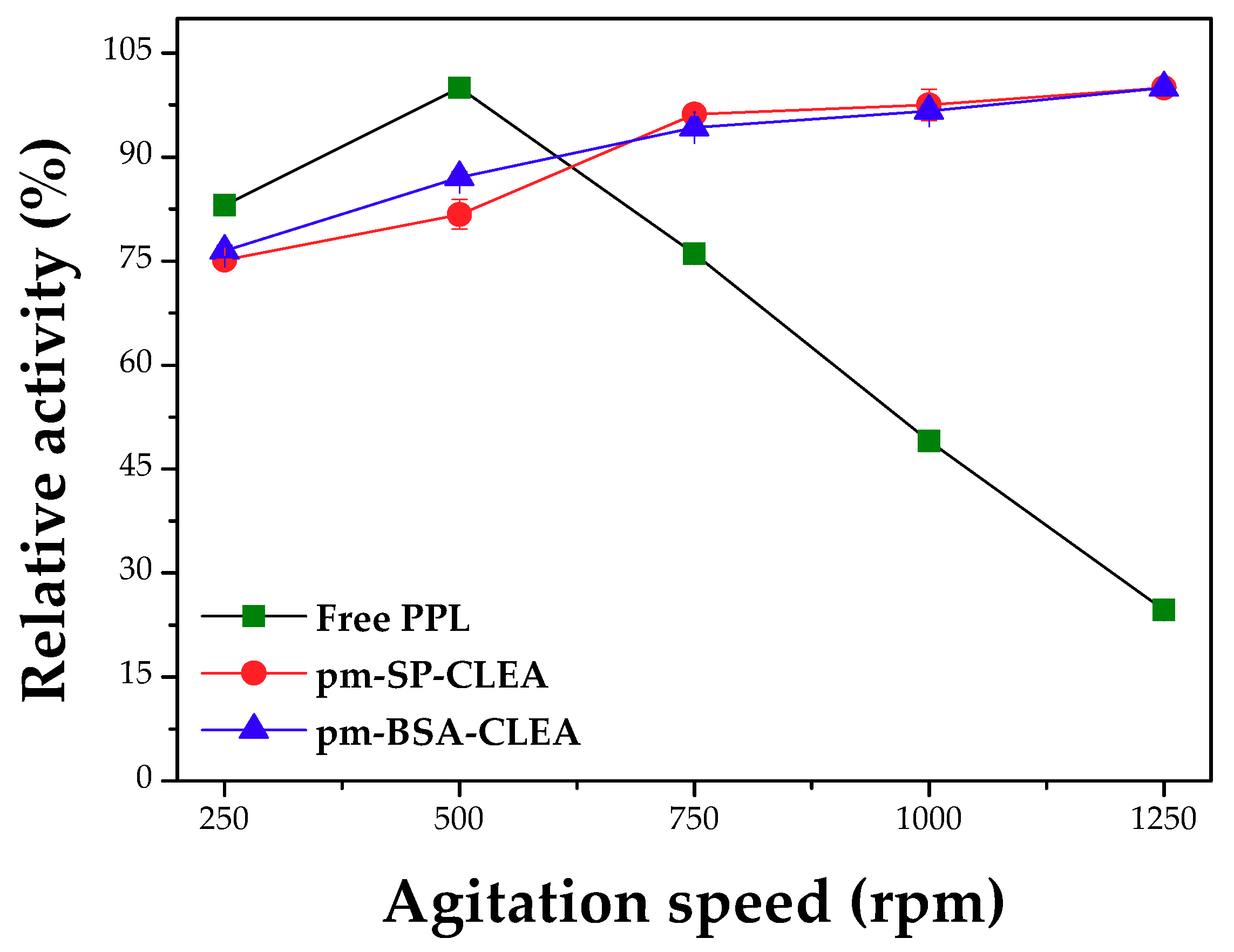
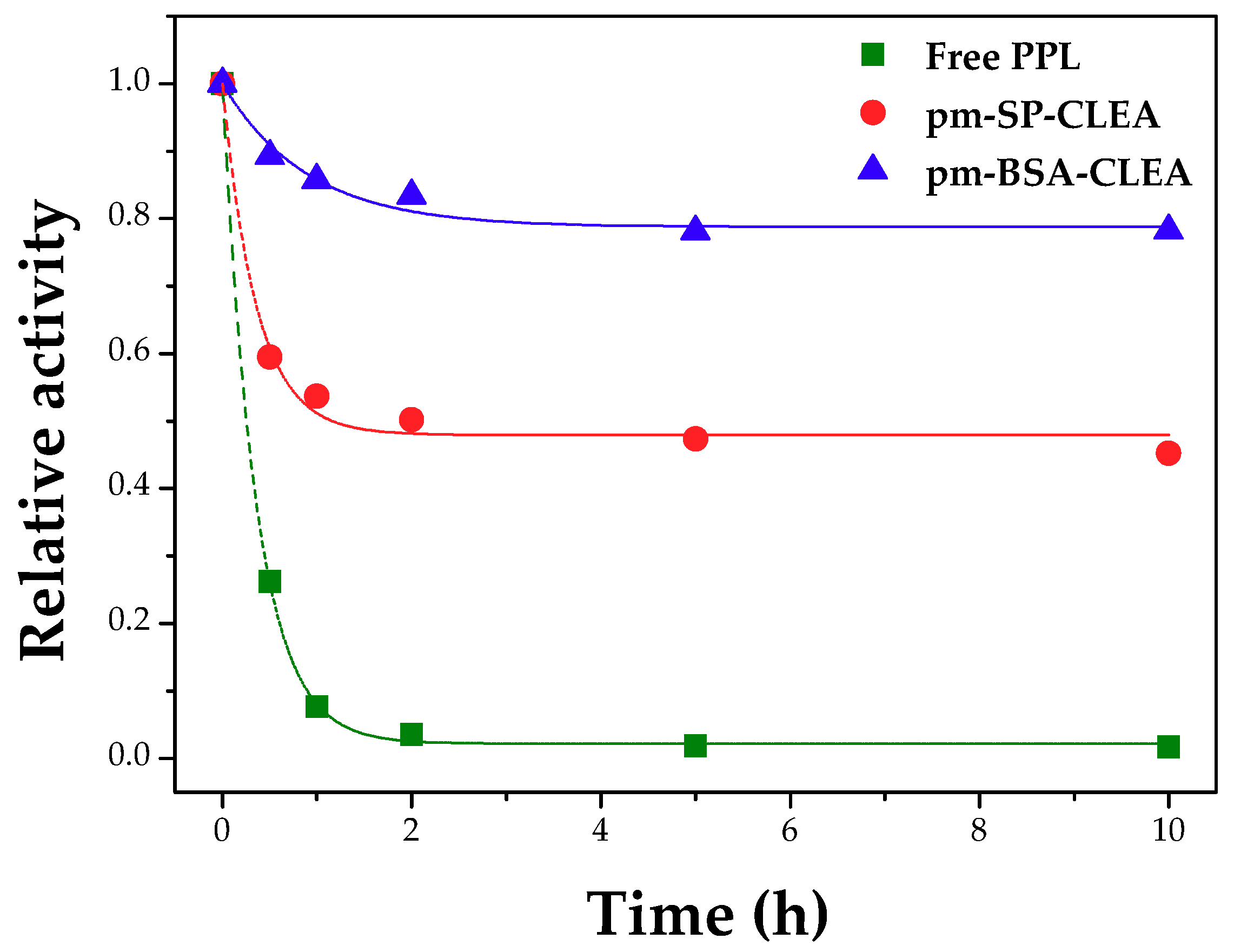
Effect of pH, Temperature and Agitation Rate on Hydrolytic Activity and Thermal Stability
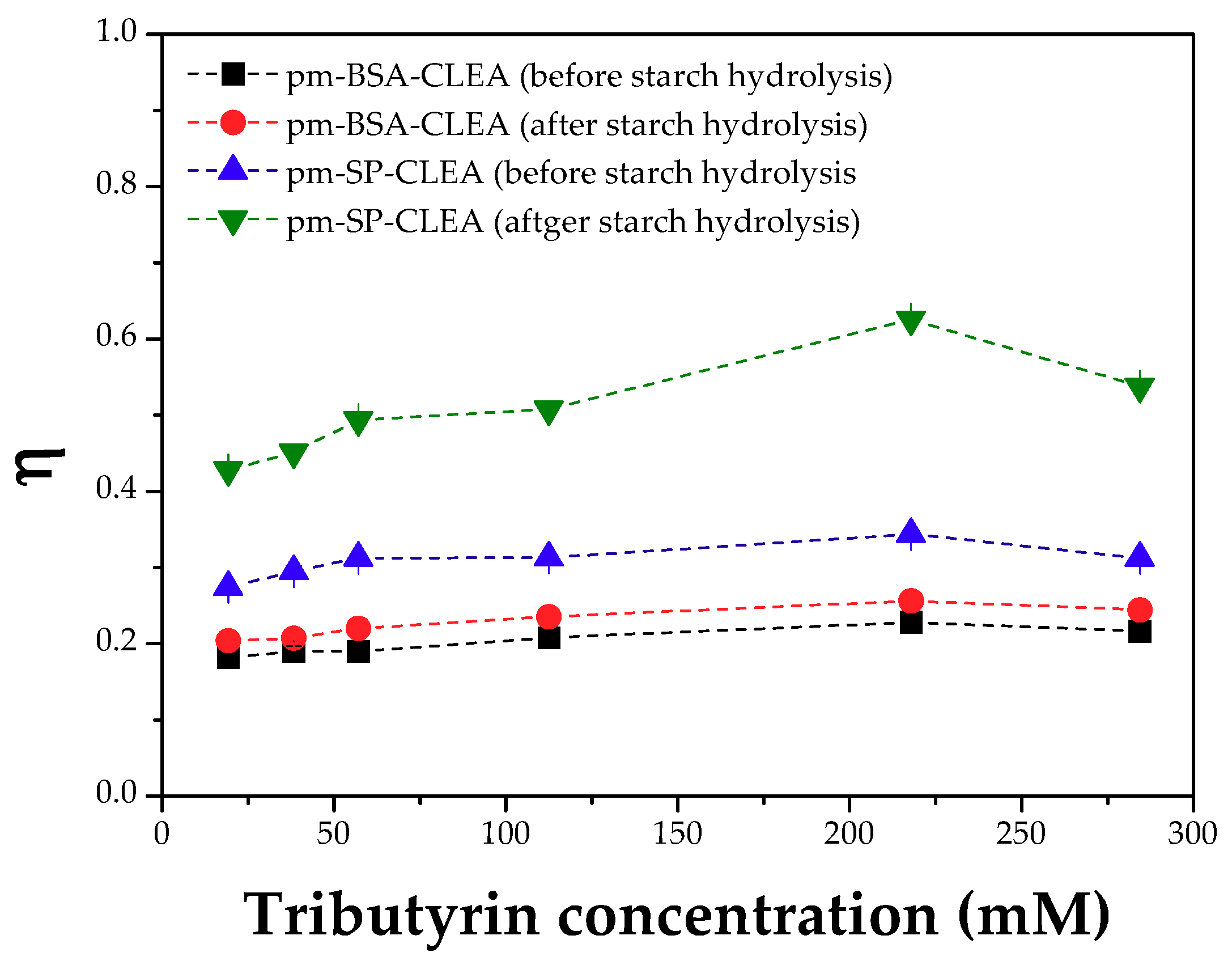
2.7. Determination of Effectiveness Factors
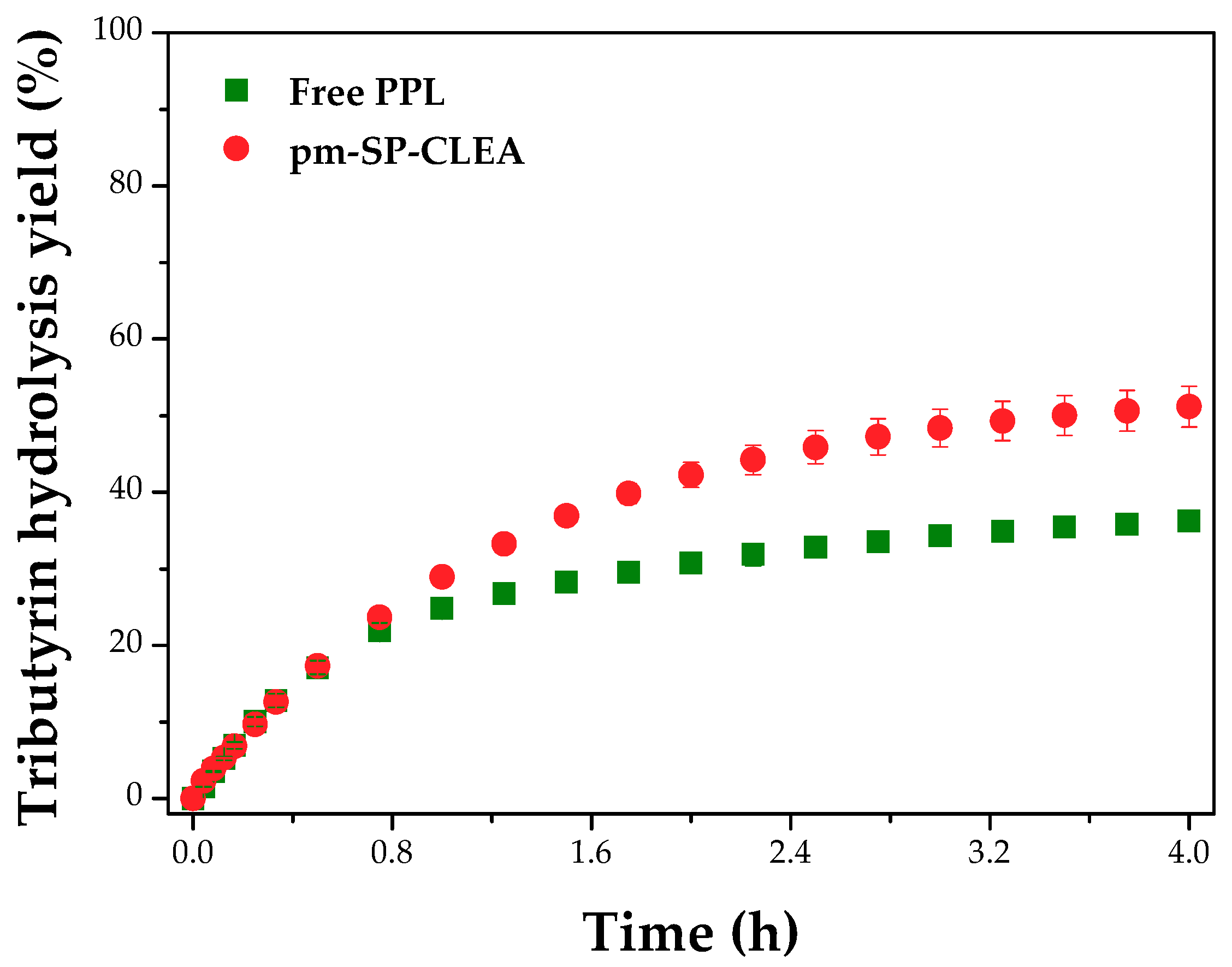
2.8. Evaluation of Tributyrin Hydrolysis and Biocatalyst Reuse
3. Materials and Methods
3.1. General Procedure of CLEA Preparation
3.2. Preparation of Porous Magnetic CLEAs (pm-CLEAs)
3.3. Protein Assay
3.4. Standard Activity Assay
3.5. Chemical or Physical Modification of the Lipase Surface
3.6. Characterization of Free and Immobilized PPL
Effect of pH and Temperature on PPL Hydrolytic Activity and Thermal Stability
3.7. Determination of Effectiveness Factor
3.8. Biocatalyst Reuse in Hydrolysis of Tributyrin
3.9. Scanning Electron Microscopy with Field Emission Gun (SEM-FEG) of PPL CLEAs
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Castro, H.F.; Mendes, A.A.; Santos, J.C.; Aguiar, C.L. Modificação de óleos e gorduras por biotransformação. Quim. Nova 2004, 27, 146–156. [Google Scholar] [CrossRef]
- Jaeger, K.E.; Eggert, T. Lipases for biotechnology. Curr. Opin. Biotechnol. 2002, 13, 390–397. [Google Scholar] [CrossRef]
- Kapoor, M.; Gupta, M.N. Lipase promiscuity and its biochemical applications. Process Biochem. 2012, 47, 555–569. [Google Scholar] [CrossRef]
- Sharma, R.; Chisti, Y.; Banerjee, U.C. Production, purification, characterization, and applications of lipases. Biotechnol. Adv. 2001, 19, 627–662. [Google Scholar] [CrossRef]
- He, W.S.; Zhu, H.; Chen, Z.Y. Plant Sterols: Chemical and enzymatic structural modifications and effects on their cholesterol-lowering activity. J. Agric. Food Chem. 2018, 66, 3047–3062. [Google Scholar] [CrossRef] [PubMed]
- Hasan, F.; Shah, A.A.; Hameed, A. Industrial applications of microbial lipases. Enzyme Microb. Technol. 2006, 39, 235–251. [Google Scholar] [CrossRef]
- Jamil, F.; Al-Haj, L.; Al-Muhtaseb, A.H.; Al-Hinai, M.A.; Baawain, M.; Rashid, U.; Ahmad, M.N.M. Current scenario of catalysts for biodiesel production: A critical review. Rev. Chem. Eng. 2018, 34, 267–297. [Google Scholar] [CrossRef]
- Mendes, A.A.; Oliveira, P.C.; Castro, H.F. Properties and biotechnological applications of porcine pancreatic lipase. J. Mol. Catal. B Enzym. 2012, 78, 119–134. [Google Scholar] [CrossRef]
- Pandey, A.; Benjamin, S.; Soccol, C.R.; Nigam, P.; Krieger, N.; Soccol, V.T. The realm of microbial lipases in biotechnology. Biotechnol. Appl. Biochem. 1999, 29, 119–131. [Google Scholar] [PubMed]
- SÁ, A.G.A.; Meneses, A.C.; Araújo, P.H.H.; Oliveira, D. A review on enzymatic synthesis of aromatic esters used as flavor ingredients for food, cosmetics and pharmaceuticals industries. Trends Food Sci. Technol. 2017, 69, 95–105. [Google Scholar] [CrossRef]
- Sarmah, N.; Revathi, D.; Sheelu, G.; Yamuna Rani, K.; Sridhar, S.; Mehtab, V.; Sumana, C. Recent advances on sources and industrial applications of lipases. Biotechnol. Prog. 2018, 34, 5–28. [Google Scholar] [CrossRef] [PubMed]
- Talekar, S.; Shah, V.; Patil, S.; Nimbalkar, M. Porous cross-linked enzyme aggregates (p-CLEAs) of Saccharomyces cerevisiae invertase. Catal. Sci. Technol. 2012, 2, 1575–1579. [Google Scholar] [CrossRef]
- Byun, H.G.; Eom, T.K.; Jung, W.K.; Kim, S.K. Lipase catalyzed production of monoacylglycerols by the esterification of fish oil fatty acids with glycerol. Biotechnol. Bioprocess Eng. 2007, 12, 491–496. [Google Scholar] [CrossRef]
- Caballero, V.; Bautista, F.M.; Campelo, J.M.; Luna, D.; Marinas, J.M.; Romero, A.A.; Hidalgo, J.M.; Luque, R.; Macario, A.; Giordano, G. Sustainable preparation of a novel glycerol-free biofuel by using pig pancreatic lipase: Partial 1,3-regiospecific alcoholysis of sunflower oil. Process Biochem. 2009, 44, 334–342. [Google Scholar] [CrossRef]
- Ramos, M.D.; Miranda, L.P.; Giordano, R.L.C.; Fernandez-Lafuente, R.; Kopp, W.; Tardioli, P.W. 1,3-Regiospecific ethanolysis of soybean oil catalyzed by crosslinked porcine pancreas lipase aggregates. Biotechnol. Prog. 2018, 34, 910–920. [Google Scholar] [CrossRef] [PubMed]
- Vescovi, V.; Santos, J.B.C.; Tardioli, P.W. Porcine pancreatic lipase hydrophobically adsorbed on octyl-silica: A robust biocatalyst for syntheses of xylose fatty acid esters. Biocatal. Biotransform. 2017, 35, 298–305. [Google Scholar] [CrossRef]
- DiCosimo, R.; McAuliffe, J.; Poulose, A.J.; Bohlmann, G. Industrial use of immobilized enzymes. Chem. Soc. Rev. 2013, 42, 6437–6474. [Google Scholar] [CrossRef] [PubMed]
- Adlercreutz, P. Immobilisation and application of lipases in organic media. Chem. Soc. Rev. 2013, 42, 6406–6436. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, O.; Torres, R.; Ortiz, C.; Berenguer-Murcia, Á.; Rodrigues, R.C.; Fernandez-Lafuente, R. Heterofunctional supports in enzyme immobilization: From traditional immobilization protocols to opportunities in tuning enzyme properties. Biomacromolecules 2013, 14, 2433–2462. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Galan, C.; Berenguer-Murcia, Á.; Fernandez-Lafuente, R.; Rodrigues, R.C. Potential of different enzyme immobilization strategies to improve enzyme performance. Adv. Synth. Catal. 2011, 353, 2885–2904. [Google Scholar] [CrossRef]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzyme Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Ortiz, C.; Berenguer-Murcia, Á.; Torres, R.; Fernández-Lafuente, R. Modifying enzyme activity and selectivity by immobilization. Chem. Soc. Rev. 2013, 42, 6290–6307. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.C.S.; Barbosa, O.; Ortiz, C.; Berenguer-Murcia, A.; Rodrigues, R.C.; Fernandez-Lafuente, R. Importance of the support properties for immobilization or purification of enzymes. ChemCatChem 2015, 7, 2413–2432. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Pelt, S. Enzyme immobilisation in biocatalysis: Why, what and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [PubMed]
- Xavier Malcata, F.; Reyes, H.R.; Garcia, H.S.; Hill, C.G.; Amundson, C.H. Immobilized lipase reactors for modification of fats and oils-A review. J. Am. Oil Chem. Soc. 1990, 67, 890–910. [Google Scholar] [CrossRef]
- Bastida, A.; Sabuquillo, P.; Armisen, P.; Fernández-Lafuente, R.; Huguet, J.; Guisán, J.M. A single step purification, immobilization, and hyperactivation of lipases via interfacial adsorption on strongly hydrophobic supports. Biotechnol. Bioeng. 1998, 58, 486–493. [Google Scholar] [CrossRef]
- Cipolatti, E.P.; Pinto, M.C.C.; Robert, J.M.; Silva, T.P.; Beralto, T.C.; Santos, J.G.F.; Castro, R.P.V.; Fernandez-Lafluente, R.; Manoel, E.A.; Pinto, J.C.; et al. Pilot-scale development of core-shell polymer supports for the immobilization of recombinant lipase B from Candida antarctica and their application in the production of ethyl esters from residual fatty acids. J. Appl. Polym. Sci. 2018, 46727, 1–13. [Google Scholar] [CrossRef]
- Mendes, A.A.; Castro, H.F.; Andrade, G.S.S.S.; Tardioli, P.W.; Giordano, R.D.L.C. Preparation and application of epoxy-chitosan/alginate support in the immobilization of microbial lipases by covalent attachment. React. Funct. Polym. 2013, 73, 160–167. [Google Scholar] [CrossRef]
- Palomo, J.M.; Muñoz, G.; Fernández-Lorente, G.; Mateo, C.; Fernández-Lafuente, R.; Guisán, J.M. Interfacial adsorption of lipases on very hydrophobic support (octadecyl–Sepabeads): Immobilization, hyperactivation and stabilization of the open form of lipases. J. Mol. Catal. B Enzym. 2002, 19, 279–286. [Google Scholar] [CrossRef]
- Paula, A.V.; Urioste, D.; Santos, J.C.; Castro, H.F. Porcine pancreatic lipase immobilized on polysiloxane–polyvinyl alcohol hybrid matrix: Catalytic properties and feasibility to mediate synthesis of surfactants and biodiesel. J. Chem. Technol. Biotechnol. 2007, 82, 281–288. [Google Scholar] [CrossRef]
- Rios, N.S.; Pinheiro, M.P.; Lima, M.L.B.; Freire, D.M.G.; Silva, I.J.; Rodríguez-Castellón, E.; Sant’Ana, H.B.; Macedo, A.C.; Gonçalves, L.R.B. Pore-expanded SBA-15 for the immobilization of a recombinant Candida antarctica lipase B: Application in esterification and hydrolysis as model reactions. Chem. Eng. Res. Des. 2018, 129, 12–24. [Google Scholar] [CrossRef]
- Vescovi, V.; Giordano, R.L.C.; Mendes, A.A.; Tardioli, P.W. Immobilized lipases on functionalized silica particles as potential biocatalysts for the synthesis of fructose oleate in an organic solvent/water system. Molecules 2017, 22, 212. [Google Scholar] [CrossRef] [PubMed]
- Brady, L.; Brzozowski, A.M.; Derewenda, Z.S.; Dodson, E.; Dodson, G.; Tolley, S.; Turkenburg, J.P.; Christiansen, L.; Huge-Jensen, B.; Norskov, L.; et al. A serine protease triad forms the catalytic centre of a triacylglycerol lipase. Nature 1990, 343, 767. [Google Scholar] [CrossRef] [PubMed]
- Miled, N.; Beisson, F.; Caro, J.; Caro, A.; Arondel, V.; Verger, R. Interfacial catalysis by lipases. J. Mol. Catal. B Enzym. 2001, 11, 165–171. [Google Scholar] [CrossRef]
- Schmid, R.D.; Verger, R. Lipases: Interfacial enzymes with attractive applications. Angew. Chem. Inst. Ed. 1998, 37, 1608–1633. [Google Scholar] [CrossRef]
- Verger, R. Interfacial activation’ of lipases: Facts and artifacts. Trends Biotechnol. 1997, 15, 32–38. [Google Scholar] [CrossRef]
- Manoel, E.A.; Santos, J.C.S.; Freire, D.M.G.; Rueda, N.; Fernandez-Lafuente, R. Immobilization of lipases on hydrophobic supports involves the open form of the enzyme. Enzyme Microb. Technol. 2015, 71. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Rantwijk, F.; Sheldon, R.A. Cross-Linked enzyme aggregates: A simple and effective method for the immobilization of Penicillin Acylase. Org. Lett. 2000, 2, 1361–1364. [Google Scholar] [CrossRef] [PubMed]
- Mateo, C.; Palomo, J.M.; Langen, L.M.; Rantwijk, F.; Sheldon, R.A. A new, mild cross-linking methodology to prepare cross-linked enzyme aggregates. Biotechnol. Bioeng. 2004, 86, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L.; Illanes, A.; Pessela, B.C.C.; Abian, O.; Fernández-Lafuente, R.; Guisán, J.M. Encapsulation of crosslinked penicillin G acylase aggregates in lentikats: Evaluation of a novel biocatalyst in organic media. Biotechnol. Bioeng. 2004, 86, 558–562. [Google Scholar] [CrossRef] [PubMed]
- Magro, L.; Hertz, P.F.; Fernandez-Lafuente, R.; Klein, M.P.; Rodrigues, R.C. Preparation and characterization of a Combi-CLEAs from pectinases and cellulases: A potential biocatalyst for grape juice clarification. RSC Adv. 2016, 6, 27242–27251. [Google Scholar] [CrossRef]
- Kim, M.H.; Park, S.; Kim, Y.H.; Won, K.; Lee, S.H. Immobilization of formate dehydrogenase from Candida boidinii through cross-linked enzyme aggregates. J. Mol. Catal. B Enzym. 2013, 97, 209–214. [Google Scholar] [CrossRef]
- López-Gallego, F.; Betancor, L.; Hidalgo, A.; Alonso, N.; Fernández-Lafuente, R.; Guisán, J.M. Co-aggregation of Enzymes and Polyethyleneimine: A simple method to prepare stable and immobilized derivatives of glutaryl acylase. Biomacromolecules 2005, 6, 1839–1842. [Google Scholar] [CrossRef] [PubMed]
- Samoylova, Y.V.; Sorokina, K.N.; Piligaev, A.V.; Parmon, V.N. Preparation of stable cross-linked enzyme aggregates (CLEAs) of a Ureibacillus thermosphaericus esterase for application in malathion removal from wastewater. Catalysts 2018, 8, 154. [Google Scholar] [CrossRef]
- Cui, J.D.; Liu, R.L.; Li, L.B. A facile technique to prepare cross-linked enzyme aggregates of bovine pancreatic lipase using bovine serum albumin as an additive. Korean J. Chem. Eng. 2016, 33, 610–615. [Google Scholar] [CrossRef]
- Guauque Torres, M.P.; Foresti, M.L.; Ferreira, M.L. Effect of different parameters on the hydrolytic activity of cross-linked enzyme aggregates (CLEAs) of lipase from Thermomyces lanuginosa. Biochem. Eng. J. 2013, 72, 18–23. [Google Scholar] [CrossRef]
- Mafra, A.C.O.; Beltrame, M.B.; Ulrich, L.G.; Giordano, R.L.C.; Ribeiro, M.P.A.; Tardioli, P.W. Combined CLEAs of invertase and soy protein for economically feasible conversion of sucrose in a fed-batch reactor. Food Bioprod. Process. 2018, 110, 145–157. [Google Scholar] [CrossRef]
- Mahmod, S.S.; Yusof, F.; Jami, M.S.; Khanahmadi, S. Optimizing the preparation conditions and characterization of a stable and recyclable cross-linked enzyme aggregate (CLEA)-protease. Bioresour. Bioprocess. 2016. [Google Scholar] [CrossRef]
- Tirunagari, H.; Basetty, S.; Rode, H.B.; Fadnavis, N.W. Crosslinked enzyme aggregates (CLEA) of phytase with soymilk proteins. J. Biotechnol. 2018, 282, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Jana, A.K.; Kumar, S.; Jana, M.M. Solid state fermentation with recovery of Amyloglucosidase from extract by direct immobilization in cross linked enzyme aggregate for starch hydrolysis. Biocatal. Agric. Biotechnol. 2015, 4, 486–492. [Google Scholar] [CrossRef]
- Wilson, L.; Fernández-Lorente, G.; Fernández-Lafuente, R.; Illanes, A.; Guisán, J.M.; Palomo, J.M. CLEAs of lipases and poly-ionic polymers: A simple way of preparing stable biocatalysts with improved properties. Enzyme Microb. Technol. 2006, 39, 750–755. [Google Scholar] [CrossRef]
- Araujo-Silva, R.; Mafra, A.; Rojas, M.; Kopp, W.; Giordano, R.; Fernandez-Lafuente, R.; Tardioli, P. Maltose production using starch from cassava bagasse catalyzed by cross-linked β-amylase aggregates. Catalysts 2018, 8, 170. [Google Scholar] [CrossRef]
- Cruz, J.; Barbosa, O.; Rodrigues, R.C.; Fernandez-Lafuente, R.; Torres, R.; Ortiz, C. Optimized preparation of CALB-CLEAs by response surface methodology: The necessity to employ a feeder to have an effective crosslinking. J. Mol. Catal. B Enzym. 2012, 80, 7–14. [Google Scholar] [CrossRef]
- Mafra, A.C.O.; Kopp, W.; Beltrame, M.B.; Giordano, R.L.C.; Ribeiro, M.P.A.; Tardioli, P.W.; Lima Camargo Giordano, R.; Arruda Ribeiro, M.P.; Tardioli, P.W. Diffusion effects of bovine serum albumin on cross-linked aggregates of catalase. J. Mol. Catal. B Enzym. 2016, 133, 107–116. [Google Scholar] [CrossRef]
- Shah, S.; Sharma, A.; Gupta, M.N. Preparation of cross-linked enzyme aggregates by using bovine serum albumin as a proteic feeder. Anal. Biochem. 2006, 351, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.D.; Jia, S.R. Optimization protocols and improved strategies of cross-linked enzyme aggregates technology: Current development and future challenges. Crit. Rev. Biotechnol. 2015, 35, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Jia, C.; Qi, W.; Yu, Q.; Peng, X.; Su, R.; He, Z. Porous-CLEAs of papain: Application to enzymatic hydrolysis of macromolecules. Bioresour. Technol. 2011, 102, 3541–3545. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Lin, T.; Feng, Y.; Tan, Z.; Jia, S. Preparation of spherical cross-linked lipase aggregates with improved activity, stability and reusability characteristic in water-in-ionic liquid microemulsion. J. Chem. Technol. Biotechnol. 2017, 92, 1785–1793. [Google Scholar] [CrossRef]
- Jafari Khorshidi, K.; Lenjannezhadian, H.; Jamalan, M.; Zeinali, M. Preparation and characterization of nanomagnetic cross-linked cellulase aggregates for cellulose bioconversion. J. Chem. Technol. Biotechnol. 2016, 91, 539–546. [Google Scholar] [CrossRef]
- Kopp, W.; Costa, T.P.; Pereira, S.C.; Jafelicci, M.; Giordano, R.C.; Marques, R.F.C.; Araújo-Moreira, F.M.; Giordano, R.L.C. Easily handling penicillin G acylase magnetic cross-linked enzymes aggregates: Catalytic and morphological studies. Process Biochem. 2014, 49, 38–46. [Google Scholar] [CrossRef]
- Kumar, V.V.; Sivanesan, S.; Cabana, H. Magnetic cross-linked laccase aggregates—Bioremediation tool for decolorization of distinct classes of recalcitrant dyes. Sci. Total Environ. 2014, 487, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Peirce, S.; Russo, M.E.; Isticato, R.; Fernández-Lafuente, R.; Salatino, P.; Marzocchella, A. Structure and activity of magnetic cross-linked enzyme aggregates of bovine carbonic anhydrase as promoters of enzymatic CO2 capture. Biochem. Eng. J. 2017, 127, 188–195. [Google Scholar] [CrossRef]
- Guauque Torres, M.P.; Foresti, M.L.; Ferreira, M.L. CLEAs of Candida antarctica lipase B (CALB) with a bovine serum albumin (BSA) cofeeder core: Study of their catalytic activity. Biochem. Eng. J. 2014, 90, 36–43. [Google Scholar] [CrossRef]
- Rhim, J.W.; Gennadiosb, A.; Wellerc, C.L.; Cezeiratd, C.; Hanna, M.A. Soy protein isolate–dialdehyde starch films. Ind. Crops Prod. 1998, 8, 195–203. [Google Scholar] [CrossRef]
- Hermoso, J.; Pignol, D.; Kerfelec, B.; Crenon, I.; Chapus, C.; Fontecilla-camps, J.C. Lipase activation by nonionic detergents. J. Biol. Chem. 1996, 271, 18007–18016. [Google Scholar] [CrossRef] [PubMed]
- Dalal, S.; Kapoor, M.; Gupta, M.N. Preparation and characterization of combi-CLEAs catalyzing multiple non-cascade reactions. J. Mol. Catal. B Enzym. 2007, 44, 128–132. [Google Scholar] [CrossRef]
- Hirata, D.B.; Albuquerque, T.L.; Rueda, N.; Virgen-Ortíz, J.J.; Tacias-Pascacio, V.G.; Fernandez-Lafuente, R. Evaluation of different immobilized lipases in transesterification reactions using tributyrin: Advantages of the heterofunctional octyl agarose beads. J. Mol. Catal. B Enzym. 2016, 133, 117–123. [Google Scholar] [CrossRef]
- Hirata, D.B.; Albuquerque, T.L.; Rueda, N.; Sánchez-Montero, J.M.; Garcia-Verdugo, E.; Porcar, R.; Fernandez-Lafuente, R. Advantages of heterofunctional octyl supports: Production of 1,2-dibutyrin by specific and selective hydrolysis of tributyrin catalyzed by immobilized lipases. Chem. Select 2016, 1, 3259–3270. [Google Scholar] [CrossRef]
- Virgen-Ortíz, J.J.; Tacias-Pascacio, V.G.; Hirata, D.B.; Torrestiana-Sanchez, B.; Rosales-Quintero, A.; Fernandez-Lafuente, R. Relevance of substrates and products on the desorption of lipases physically adsorbed on hydrophobic supports. Enzyme Microb. Technol. 2017, 96, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beisson, F.; Tiss, A.; Rivière, C.; Verger, R. Methods for lipase detection and assay: A critical review. Eur. J. Lipid Sci. Technol. 2000, 102, 133–153. [Google Scholar] [CrossRef]
- Snyder, S.L.; Sobocinski, P.Z. An improved 2,4,6-trinitrobenzenesulfonic acid method for the determination of amines. Anal. Biochem. 1975, 64, 284–288. [Google Scholar] [CrossRef]
- Cipolatti, E.P.; Valério, A.; Henriques, R.O.; Moritz, D.E.; Ninow, J.L.; Freire, D.M.G.; Manoel, E.A.; Fernandez-Lafuente, R.; De Oliveira, D. Nanomaterials for biocatalyst immobilization-state of the art and future trends. RSC Adv. 2016, 6, 104675–104692. [Google Scholar] [CrossRef]
- Baniukevic, J.; Hakki Boyaci, I.; Goktug Bozkurt, A.; Tamer, U.; Ramanavicius, A.; Ramanaviciene, A. Magnetic gold nanoparticles in SERS-based sandwich immunoassay for antigen detection by well oriented antibodies. Biosens. Bioelectron. 2013, 43, 281–288. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CLEAs of PPL | RA a ± σ (%) | IY b ± σ (%) | GY c ± σ (%) |
|---|---|---|---|
| PPL CLEA d | 1.46 ± 0.08 | 65.25 ± 3.52 | 0.93 ± 0.003 |
| PPL BSA-CLEA e | 9.40 ± 0.11 | 93.69 ± 1.50 | 9.45 ± 0.39 |
| PPL SP-CLEA f | 22.81 ± 1.16 | 51.42 ± 0.35 | 10.72 ± 0.34 |
| PPL SMNPs-CLEA g | 12.27 ± 0.77 | 63.04 ± 2.43 | 7.72 ± 0.24 |
| PPL BSA-SMNPs-CLEA h | 25.43 ± 0.21 | 69.69 ± 1.2 | 20.30 ± 0.52 |
| PPL SP-SMNPs-CLEA i | 32.84 ± 1.76 | 45.25 ± 1.07 | 16.40 ± 1.12 |
| PPL Treatment with | RA ± σ (%) | IY ± σ (%) |
|---|---|---|
| PEI | 7.17 ± 0.32 | 99.50 ± 1.00 |
| Dodecyl aldehyde | 20.08 ± 0.72 | 99.00 ± 2.00 |
| PEI followed by Dodecyl aldehyde | 25.64 ± 1.37 | 99.60 ± 2.00 |
| Additives | Recovered Activities of Magnetic CLEAs (RA ± σ (%)) | |
|---|---|---|
| Before Starch Hydroysis with α-Amylase | After Starch Hydrolysis with α-Amylase | |
| SMNPs a | 24.46 ± 0.24 | 54.01 ± 0.64 |
| BSA + SMNPs b | 28.18 ± 0.67 | 66.55 ± 0.01 |
| SP + SMNPs c | 36.77 ± 0.38 | 81.02 ± 0.16 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guimarães, J.R.; Giordano, R.d.L.C.; Fernandez-Lafuente, R.; Tardioli, P.W. Evaluation of Strategies to Produce Highly Porous Cross-Linked Aggregates of Porcine Pancreas Lipase with Magnetic Properties. Molecules 2018, 23, 2993. https://doi.org/10.3390/molecules23112993
Guimarães JR, Giordano RdLC, Fernandez-Lafuente R, Tardioli PW. Evaluation of Strategies to Produce Highly Porous Cross-Linked Aggregates of Porcine Pancreas Lipase with Magnetic Properties. Molecules. 2018; 23(11):2993. https://doi.org/10.3390/molecules23112993
Chicago/Turabian StyleGuimarães, José Renato, Raquel de Lima Camargo Giordano, Roberto Fernandez-Lafuente, and Paulo Waldir Tardioli. 2018. "Evaluation of Strategies to Produce Highly Porous Cross-Linked Aggregates of Porcine Pancreas Lipase with Magnetic Properties" Molecules 23, no. 11: 2993. https://doi.org/10.3390/molecules23112993
APA StyleGuimarães, J. R., Giordano, R. d. L. C., Fernandez-Lafuente, R., & Tardioli, P. W. (2018). Evaluation of Strategies to Produce Highly Porous Cross-Linked Aggregates of Porcine Pancreas Lipase with Magnetic Properties. Molecules, 23(11), 2993. https://doi.org/10.3390/molecules23112993