Chemical Constituents from Apios americana and Their Inhibitory Activity on Tyrosinase

 ,
, 

Abstract
:
1. Introduction
2. Results and Discussion
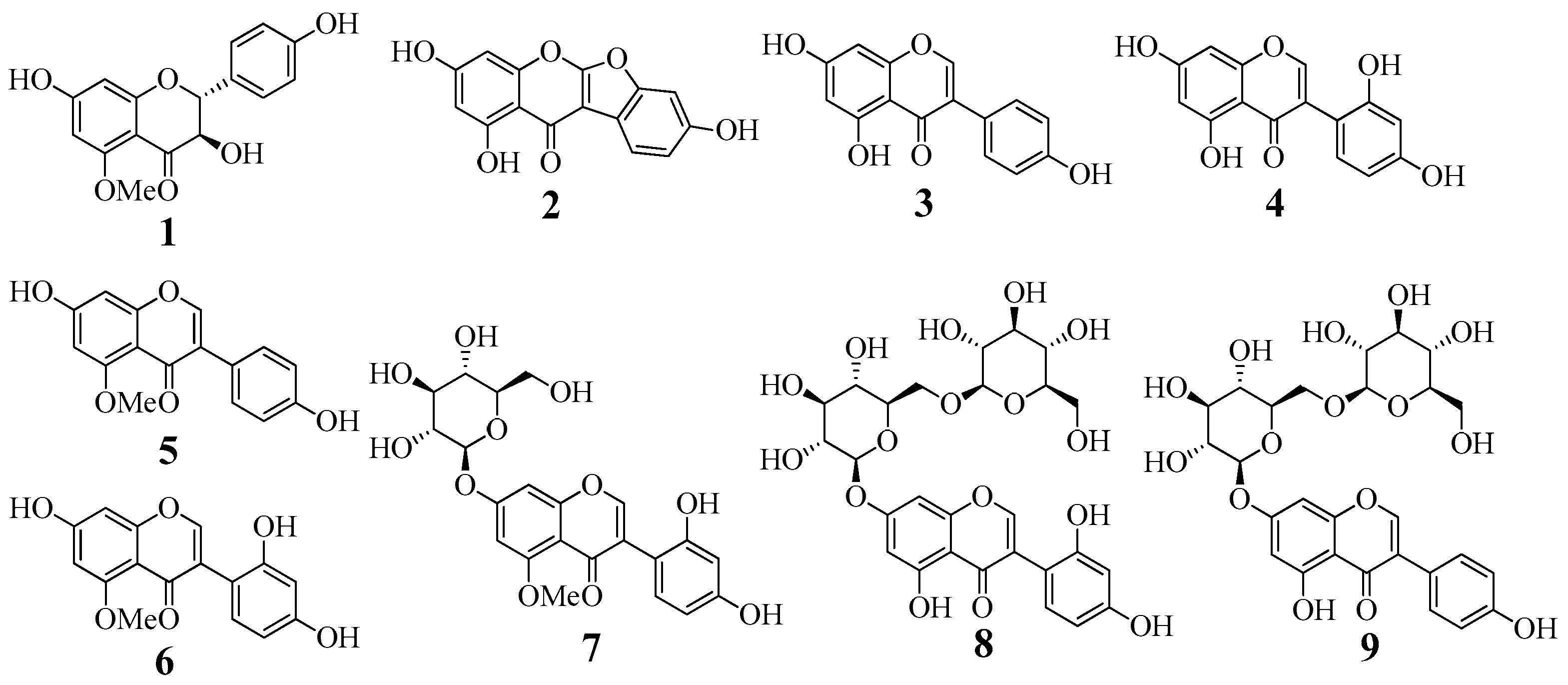
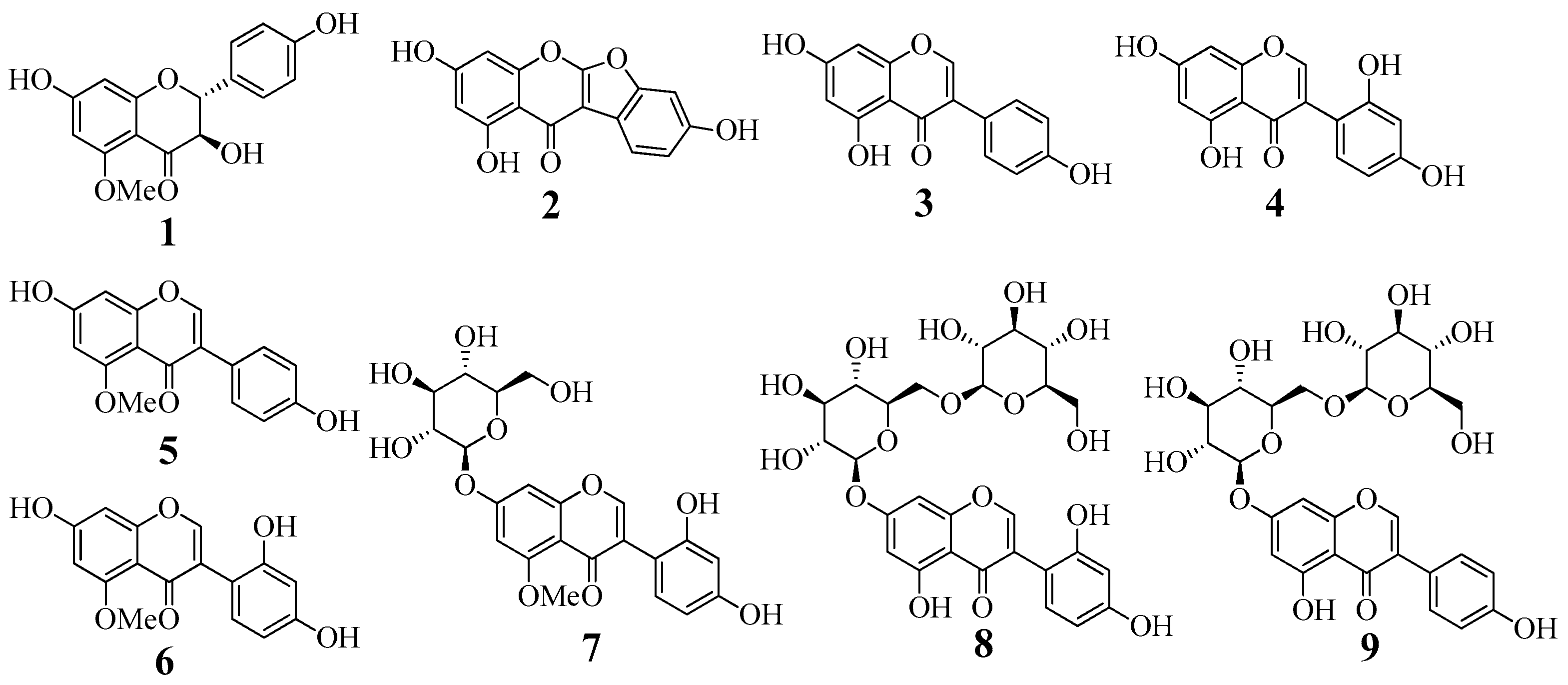
2.1. Isolation and Identification
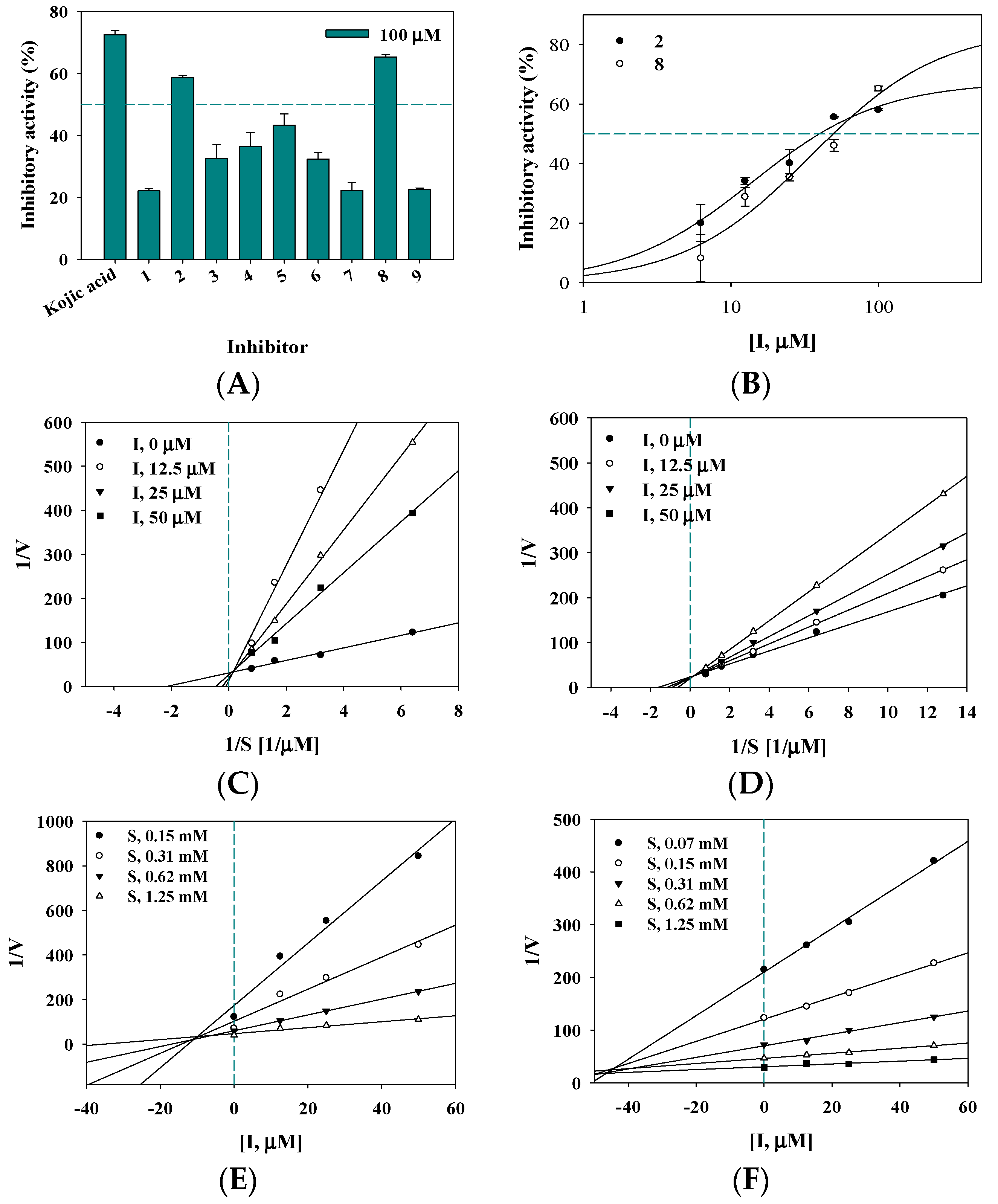
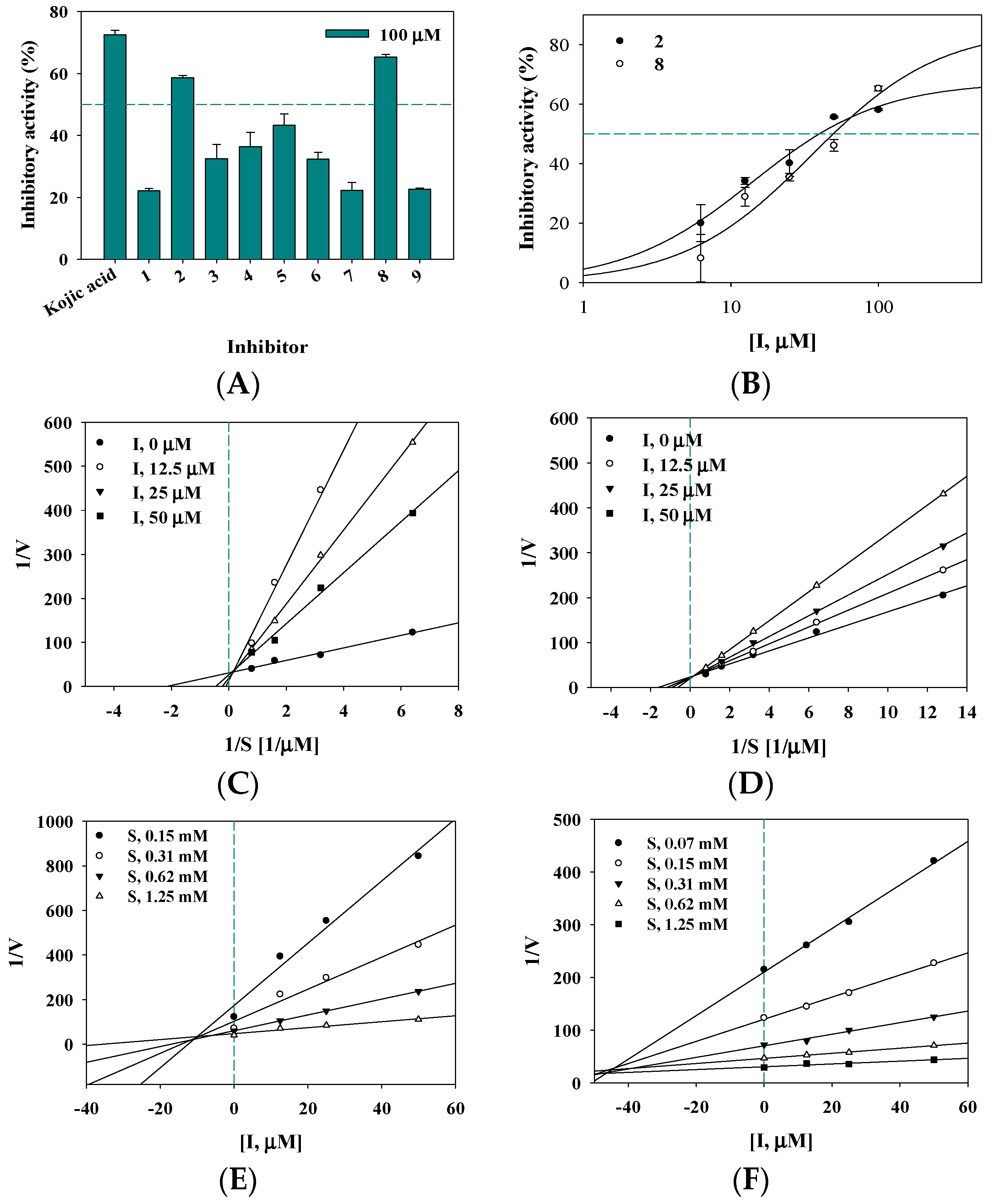
2.2. Inhibitory Activity on Tyrosinase
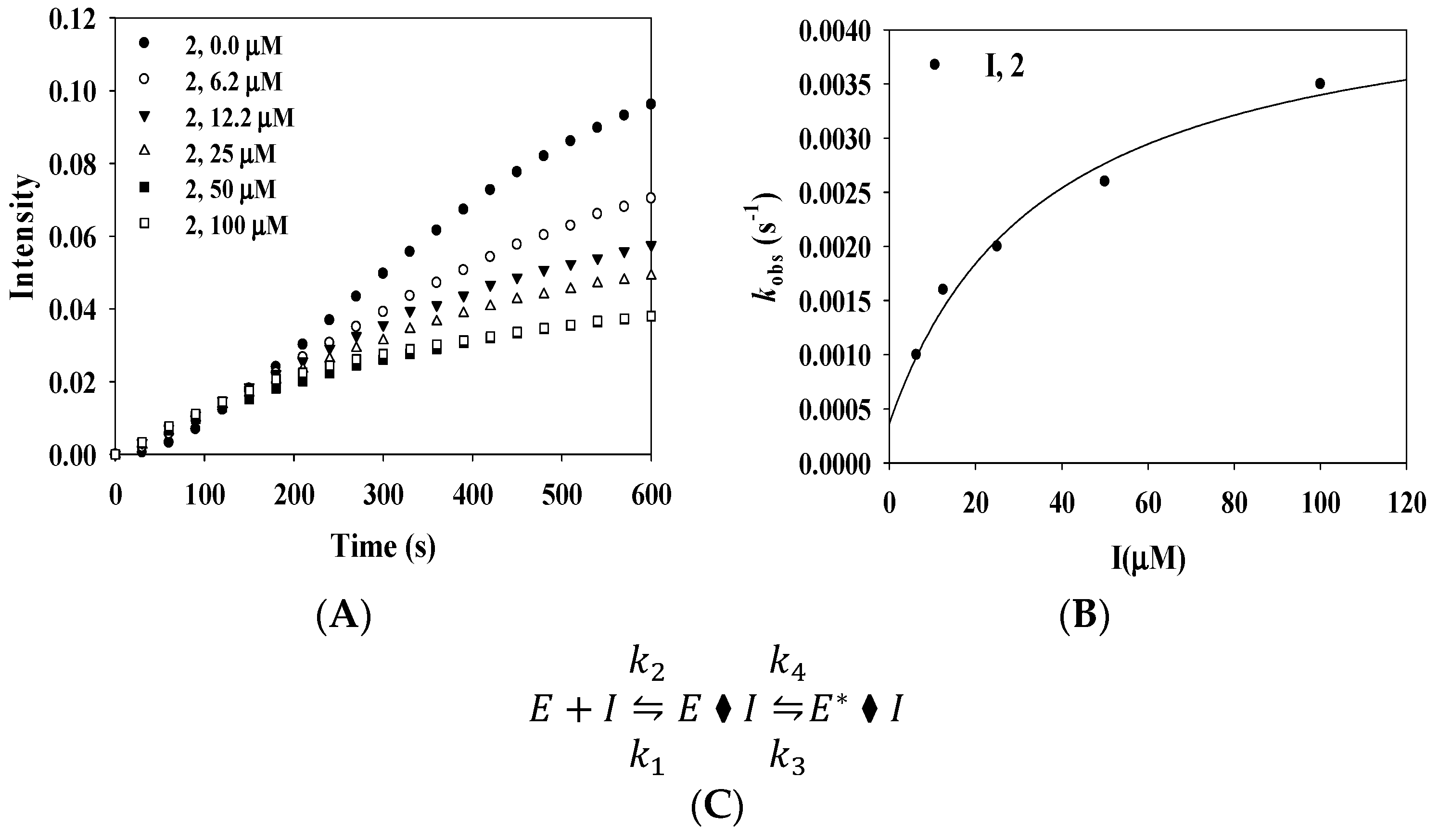
2.3. Enzyme Kinetics
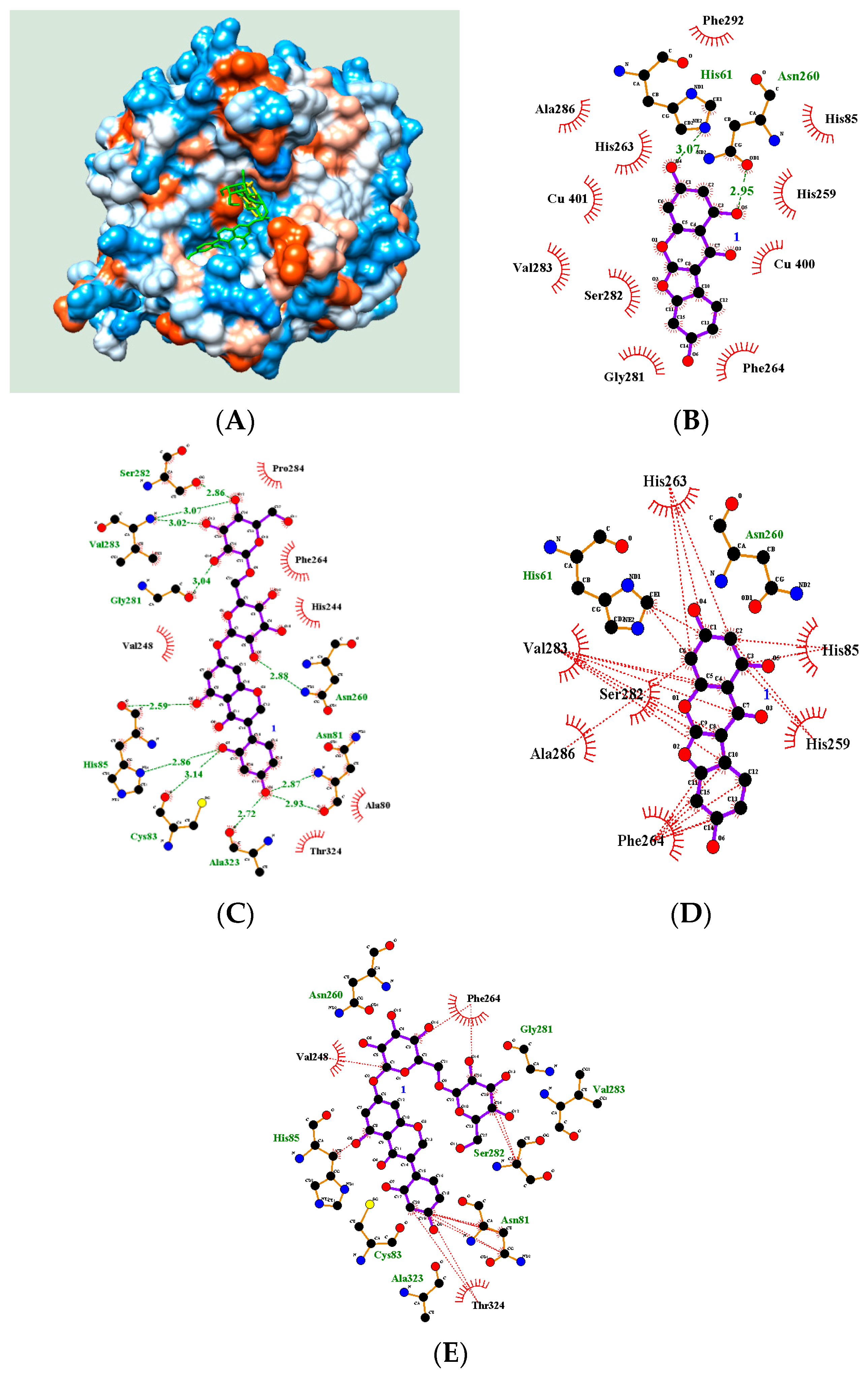
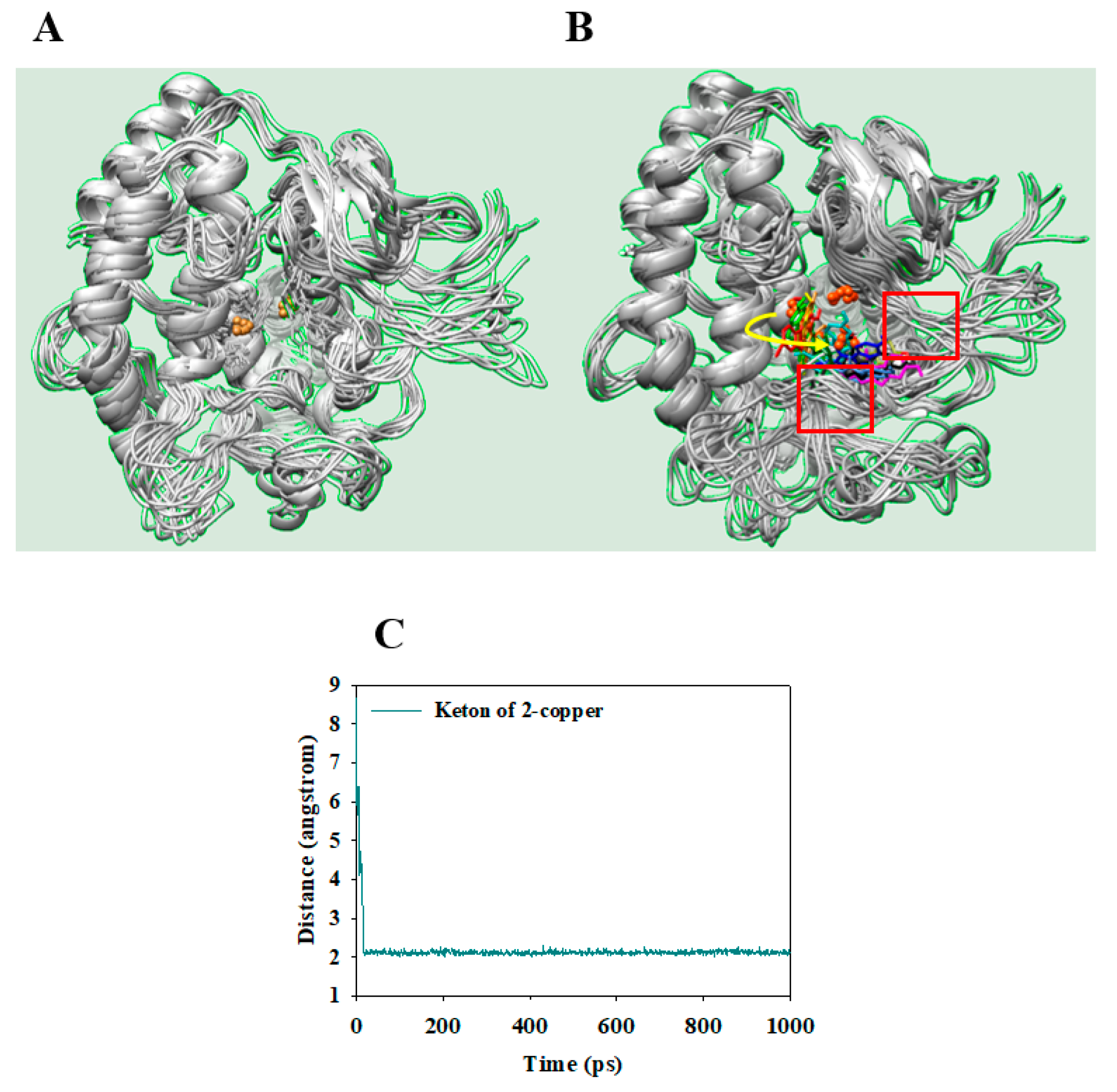
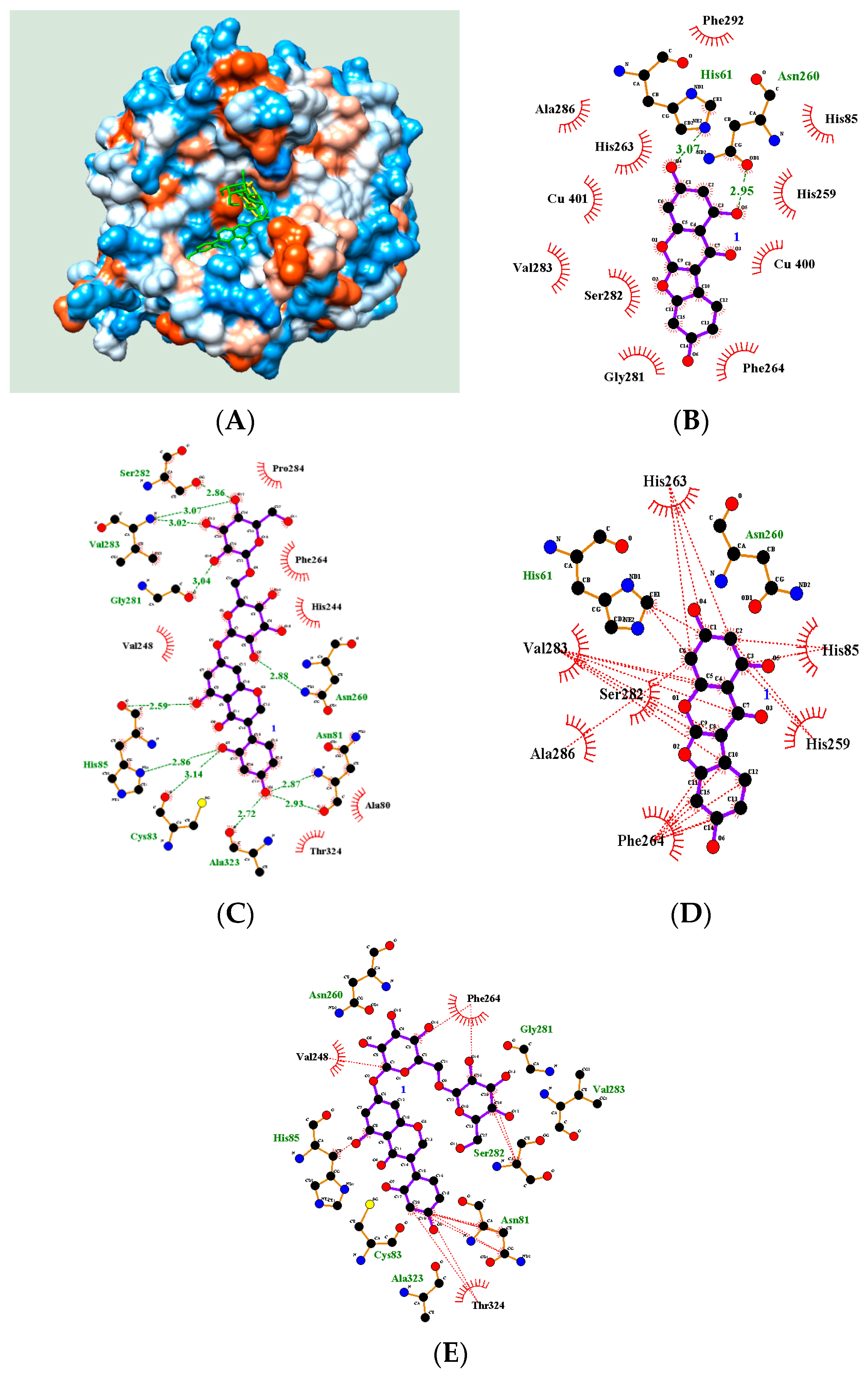
2.4. Molecular Docking
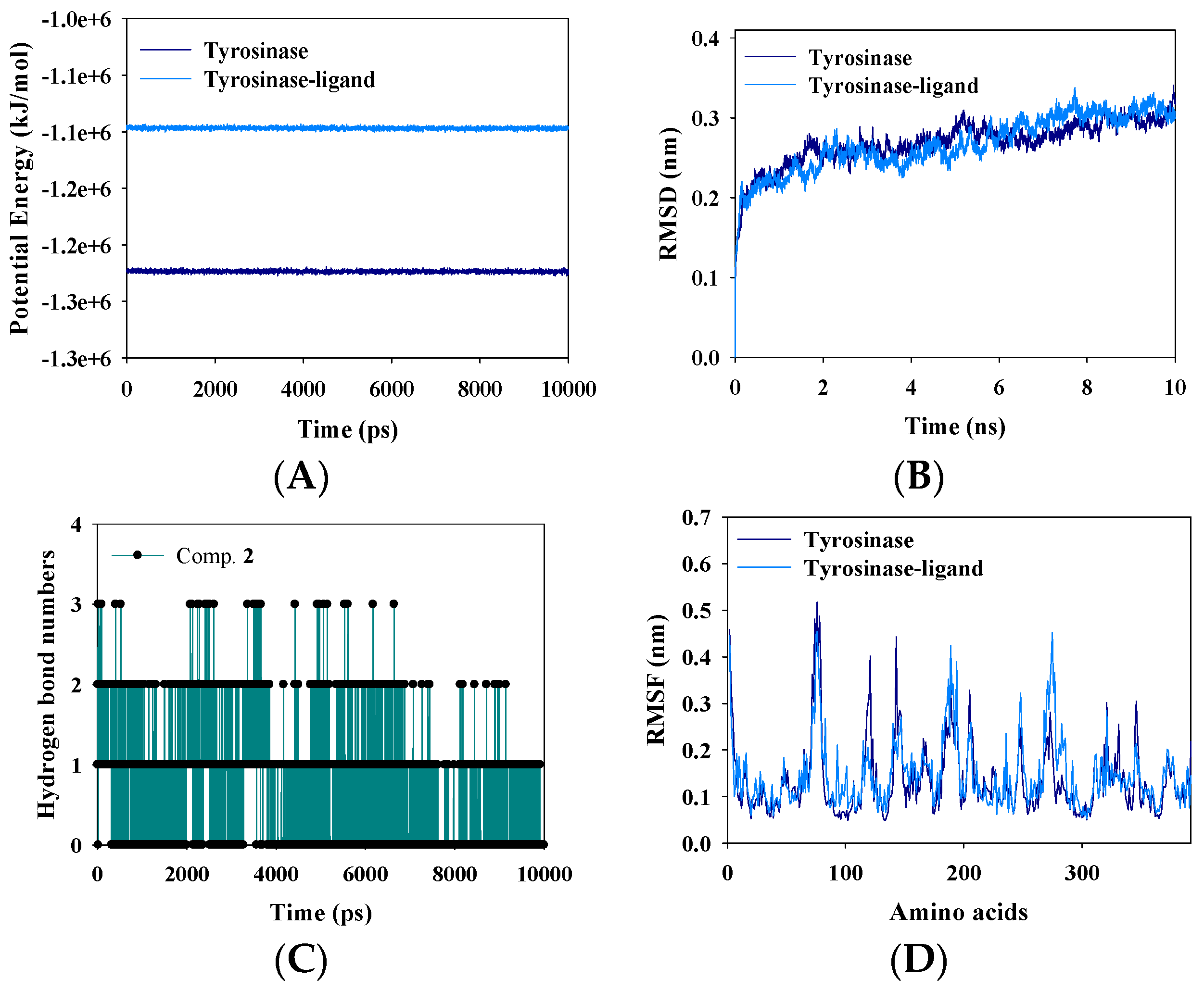
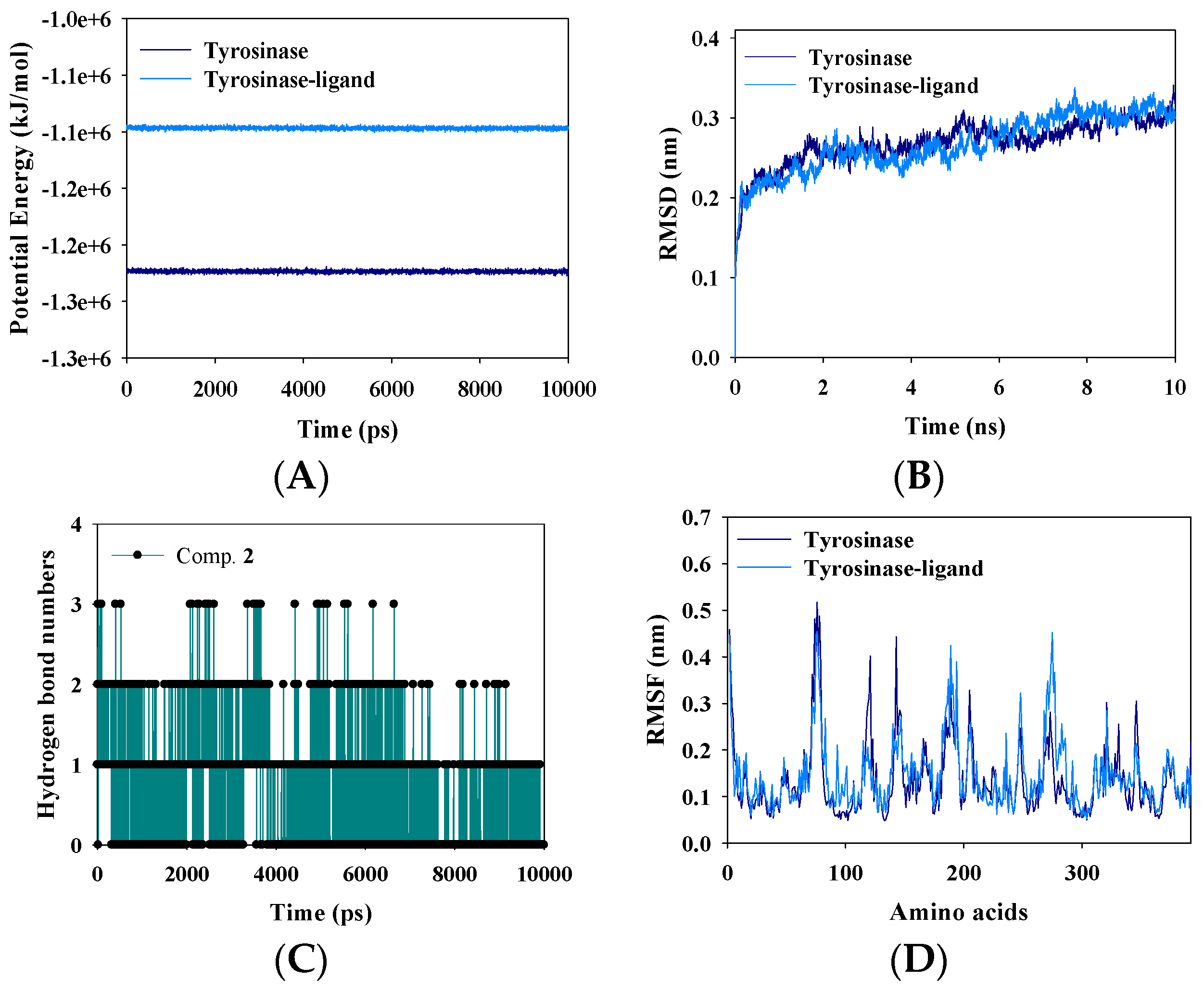
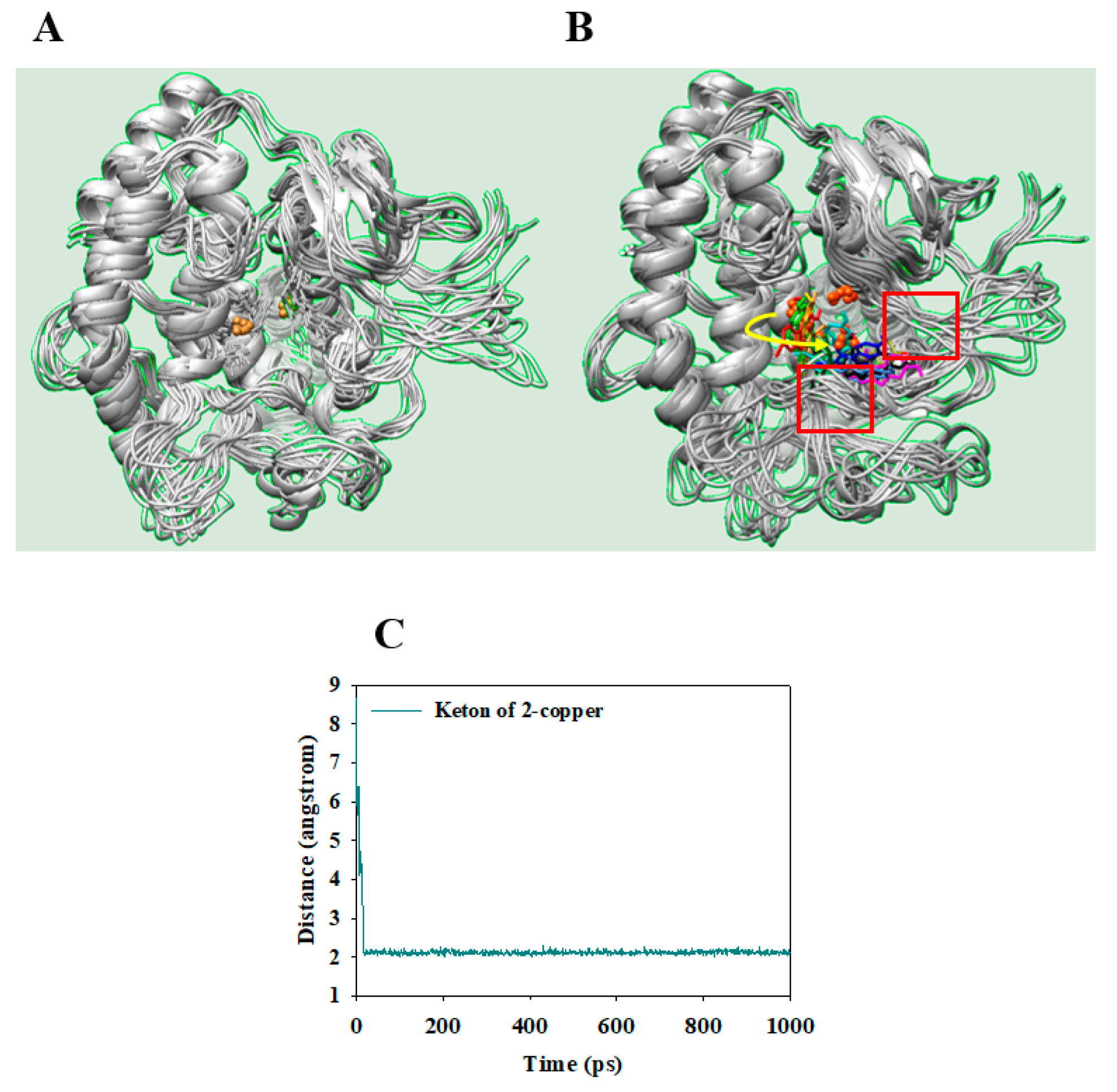
2.5. Molecular Dynamics
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Plant Materials
3.3. Extraction and Isolation
3.4. Enzyme Inhibition Assay
3.5. Molecular Simulations
3.6. Molecular Dynamics
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chen, Z.; Cai, D.; Mou, D.; Yan, Q.; Sun, Y.; Pan, W.; Wan, Y.; Song, H.; Yi, W. Design, synthesis and biological evaluation of hydroxyl- or methoxy-substituted 5-benzylidene (thio) barbiturates as novel tyrosinase inhibitors. Bioorg. Med. Chem. 2014, 22, 3279–3284. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.-H.; Liu, X.; Jia, Y.-L.; Guo, Y.-J.; Wang, Q.; Chen, Q.-X. Inhibitory kinetics of chlorocinnamic acids on mushroom tyrosinase. J. Biosci. Bioeng. 2014, 2, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.-J.; Liu, K.-Y.; Lee, J.; Yang, J.-M.; Qian, G.-Y.; Si, Y.-X.; Park, Y.-D. Effect of hydroxyfallor yellow A on tyrosinase: Intergration of inhibition kinetics with computational simulation. Process Biochem. 2015, 50, 2112–2120. [Google Scholar] [CrossRef]
- Akhtar, M.N.; Sakeh, N.M.; Zareen, S.; Gul, S.; Lo, K.M.; UL-Haq, Z.; Shah, S.A.A.; Ahmad, S. Design and synthesis of chalcone derivatives as potent tyrosinase inhibitors and their structural activity relationship. J. Mol. Struct. 2015, 1085, 97–103. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, G.; Yan, J.; Gong, D. Inhibitory effect of morin on tyrosinase: Insights from spectroscopic and molecular docking studies. Food Chem. 2014, 163, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.K.; Choi, J.-U.; Lee, E.-A.; Park, H.-R. Flaniostatin, a new isoflavonoid glycoside isolated from the leaves of Cudrania tricuspidata as a tryosinase inhibitor. Food Sci. Biotechnol. 2013, 22, 1449–1452. [Google Scholar] [CrossRef]
- Kim, J.H.; Yoon, J.-Y.; Yang, S.Y.; Choi, S.-K.; Kwon, S.J.; Cho, I.S.; Jeong, M.H.; Kim, Y.H.; Choi, G.S. Tyrosinase inhibitory components from Aloe vera and their antiviral activity. J. Enzym. Inhib. Med. Chem. 2016, 32, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.-S.; Ding, H.-Y.; Lin, H.-C. Identifying 6,7,4′-trihydroxyisoflavone as a potent tyrosinase inhibitor. Biosci. Biotechnol. Biochem. 2005, 69, 1999–2001. [Google Scholar] [CrossRef] [PubMed]
- Veitch, N.C. Isoflavonoids of the Leguminosae. Nat. Prod. Rep. 2013, 30, 988–1027. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.-P. Isoflavones: Chemistry, analysis, Functions and effects on health and cancer. Asian Pac. J. Cancer Prev. 2014, 15, 7001–7010. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.-L.; Liu, J.; Zhu, C.; Gao, Y.; Zhao, S.; Liu, W.; Zhang, Y. Interactions between soy isoflavones and other bioactive compounds: A review of their potentially beneficial health effects. Phytochem. Rev. 2015, 14, 459–467. [Google Scholar] [CrossRef]
- Sun, M.; Xie, H.F.; Tang, Y.; Lin, S.-Q.; Sun, S.-N.; Hu, X.-L.; Huang, Y.-X.; Shi, W.; Jian, D. G-protein-coupled estrogen receptor enhances melanogenesis via cAMP-progein kinase (PKA) by upregulating microphthalmia-related transcription factor-tyrosinase in melanoma. J. Steroid Biochem. Mol. Biol. 2017, 165, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Nara, K.; Nihei, K.-I.; Ogasawara, Y.; Koga, H.; Kato, Y. Novel isoflavone diglycoside in groundnut (Apios americana Medik). Food Chem. 2011, 124, 703–710. [Google Scholar] [CrossRef]
- Belamkar, V.; Farmer, A.D.; Weeks, N.T.; Kalberer, S.R.; Blackmon, W.J.; Cannon, S.B. Genomics-assisted characterization of a breeding collection of Apios americana, an edible tuberous legume. Sci. Rep. 2016, 6, 34908. [Google Scholar] [CrossRef] [PubMed]
- Kenmochi, E.; Kabir, S.R.; Ogawa, T.; Naude, R.; Tateno, H.; Hirabayashi, J.; Muramoto, K. Isolation and biochemical characterization of Apios tuber lectin. Molecules 2015, 20, 987–1002. [Google Scholar] [CrossRef] [PubMed]
- Ichige, M.; Fukuda, E.; Miida, S.; Hatta, J.-I.; Misawa, N.; Saito, S.; Fujimaki, T.; Imoto, M.; Shindo, K. Novel isoflavone glucosides in groundnut (Apios americana Medik) and their antiandrogenic activities. J. Agric. Food Chem. 2013, 61, 2183–2187. [Google Scholar] [CrossRef] [PubMed]
- Kaneta, H.; Koda, M.; Saito, S.; Imoto, M.; Kawada, M.; Yamazaki, Y.; Momose, I.; Shindo, K. Biological activites of unique isoflavones prepared from Apios americana Medik. Biosci. Biotech. Biochem. 2016, 80, 774–778. [Google Scholar] [CrossRef] [PubMed]
- Bilia, A.R.; Catalano, S.; Pistelli, L.; Morelli, I. Flavonoids from Pyracantha coccinea roots. Phytochemistry 1993, 33, 1449–1452. [Google Scholar] [CrossRef]
- Do, T.M.L.; Truong, A.V.; Pinnock, T.G.; Pratt, L.M.; Yamamoto, S.; Watarai, H.; Guillanume, D.; Nguyen, K.P.P. New rotenoids and coumaronochromonoids from the aerial part of Boerchaaive recta. Chem. Pharm. Bull. 2013, 61, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Chaipukdee, N.; Kanokmedhakul, S.; Lekphrom, R.; Kanokmedhakul, K. Two new flavanonols from the bark of Akschindlium godefroyanum. Nat. Prod. Res. 2014, 28, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-S.; Kim, D.H.; Lee, J.K.; Lee, J.Y.; Kim, D.H.; Kim, H.K.; Lee, H.-J.; Kim, H.C. Natural ortho-dihydroxyisoflavone derivatives from aged Korean fermented soybean paste as potent tyrosinase and melanin formation inhibitor. Bioorg. Med. Chem. Lett. 2010, 20, 1162–1164. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Cho, C.W.; Tai, B.H.; Yang, S.Y.; Choi, G.-S.; Kang, J.S.; Kim, Y.H. Soluble epoxide hydrolase inhibitory activity of selaginellin derivatives from Selaginella tamariscina. Molecules 2015, 20, 21405–21414. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Ryu, Y.B.; Lee, W.S.; Kim, Y.H. Neuraminidase inhibitory activities of quaternary isoquinoline alkaloids from Corydalis turtschaninovii rhizome. Bioorg. Med. Chem. 2014, 22, 6047–6052. [Google Scholar] [CrossRef] [PubMed]
- Leem, H.H.; Lee, G.Y.; Lee, J.S.; Lee, H.; Kim, J.H.; Kim, Y.H. Soluble epoxide hydrolase inhibitory activity of components from Leonurus japonicas. Int. J. Biol. Macromol. 2017, 10, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, S.-H.; Lee, H.W.; Sun, Y.N.; Jang, W.-H.; Yang, S.-Y.; Jang, H.D.; Kim, Y.H. (−)-Epicatechin derivate from Orostachys japonicas as potential inhibitor of the human butyrylcholinesterase. Int. J. Biol. Macromol. 2016, 91, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inhibitory Activity on Tyrosinase | ||
|---|---|---|
| IC50 (μM) a | Binding Mode (Ki: μM) a | |
| 2 | 39.7 ± 1.5 13.2 ± 1.0 c | Competitive (10.3 ± 0.8) |
| 8 | 50.0 ± 3.7 | Competitive (44.2 ± 1.7) |
| Kojic acid b | 25.2 ± 0.8 | |
| k5(S−1) | k6(S−1) | (μM) | |
|---|---|---|---|
| 2 | 0.0041 | 0.0004 | 35.8 |
| Hydrogen Bonds (Å) | Binding Energy (kcal/mol) | |
|---|---|---|
| 2 | His61(3.07), Asn260(2.95) | −5.87 |
| 8 | Asn81(2.87, 2.93), Cys83(3.14), His85(2.86, 2.59), Gly281(3.04), Val283(3.02, 3.07), Ser282 (2.86), Asn260(2.88) | −4.58 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.H.; Kim, H.Y.; Kang, S.Y.; Kim, J.-B.; Kim, Y.H.; Jin, C.H. Chemical Constituents from Apios americana and Their Inhibitory Activity on Tyrosinase. Molecules 2018, 23, 232. https://doi.org/10.3390/molecules23010232
Kim JH, Kim HY, Kang SY, Kim J-B, Kim YH, Jin CH. Chemical Constituents from Apios americana and Their Inhibitory Activity on Tyrosinase. Molecules. 2018; 23(1):232. https://doi.org/10.3390/molecules23010232
Chicago/Turabian StyleKim, Jang Hoon, Hyo Young Kim, Si Yong Kang, Jin-Baek Kim, Young Ho Kim, and Chang Hyun Jin. 2018. "Chemical Constituents from Apios americana and Their Inhibitory Activity on Tyrosinase" Molecules 23, no. 1: 232. https://doi.org/10.3390/molecules23010232
APA StyleKim, J. H., Kim, H. Y., Kang, S. Y., Kim, J.-B., Kim, Y. H., & Jin, C. H. (2018). Chemical Constituents from Apios americana and Their Inhibitory Activity on Tyrosinase. Molecules, 23(1), 232. https://doi.org/10.3390/molecules23010232