Time-dependent Inhibition of CYP2C8 and CYP2C19 by Hedera helix Extracts, A Traditional Respiratory Herbal Medicine

 and
and 
Abstract
:1. Introduction
2. Results
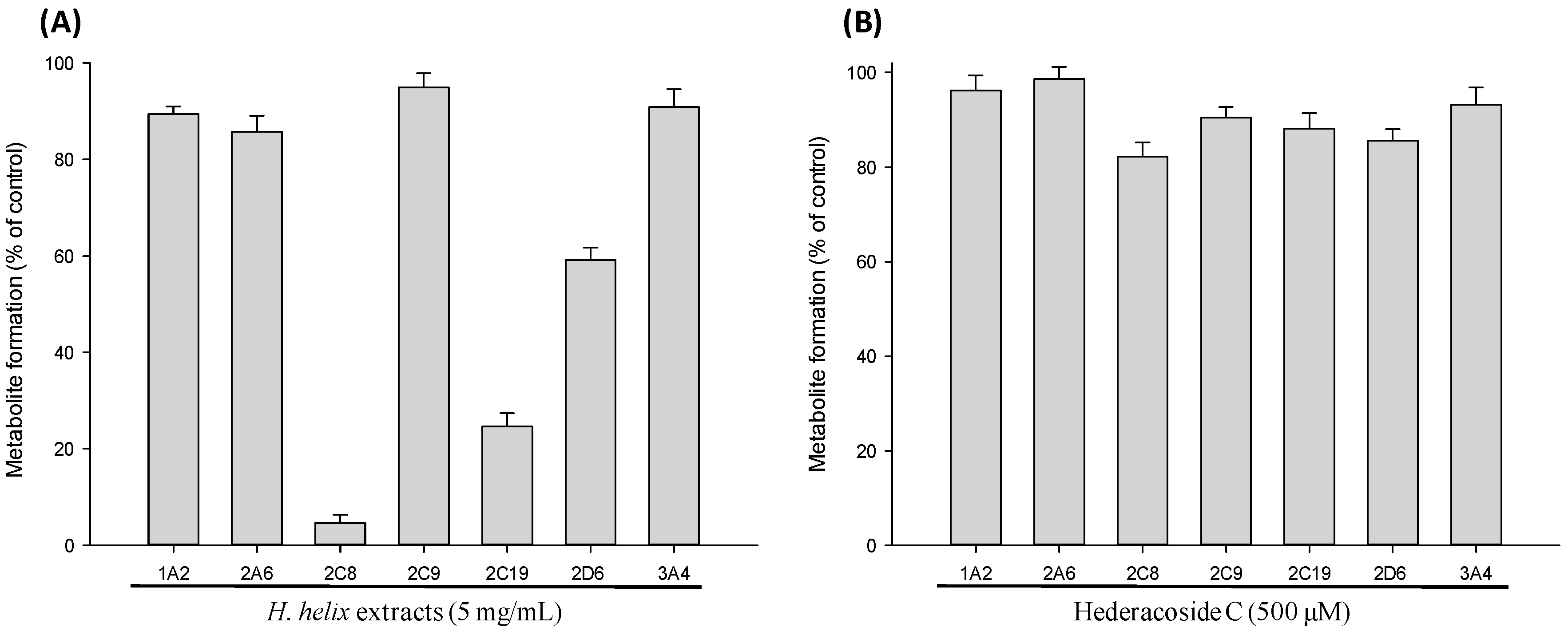
2.1. CYP Inhibition Assay in Pooled Human Liver Microsomes
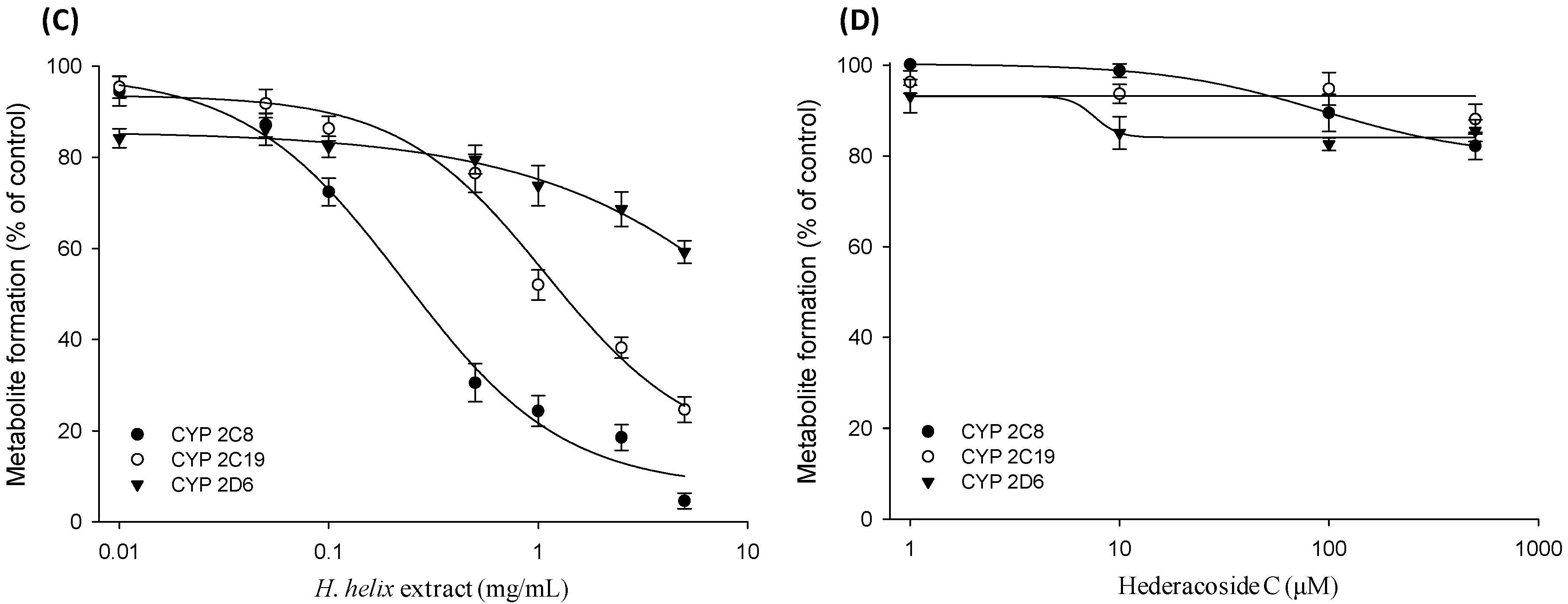
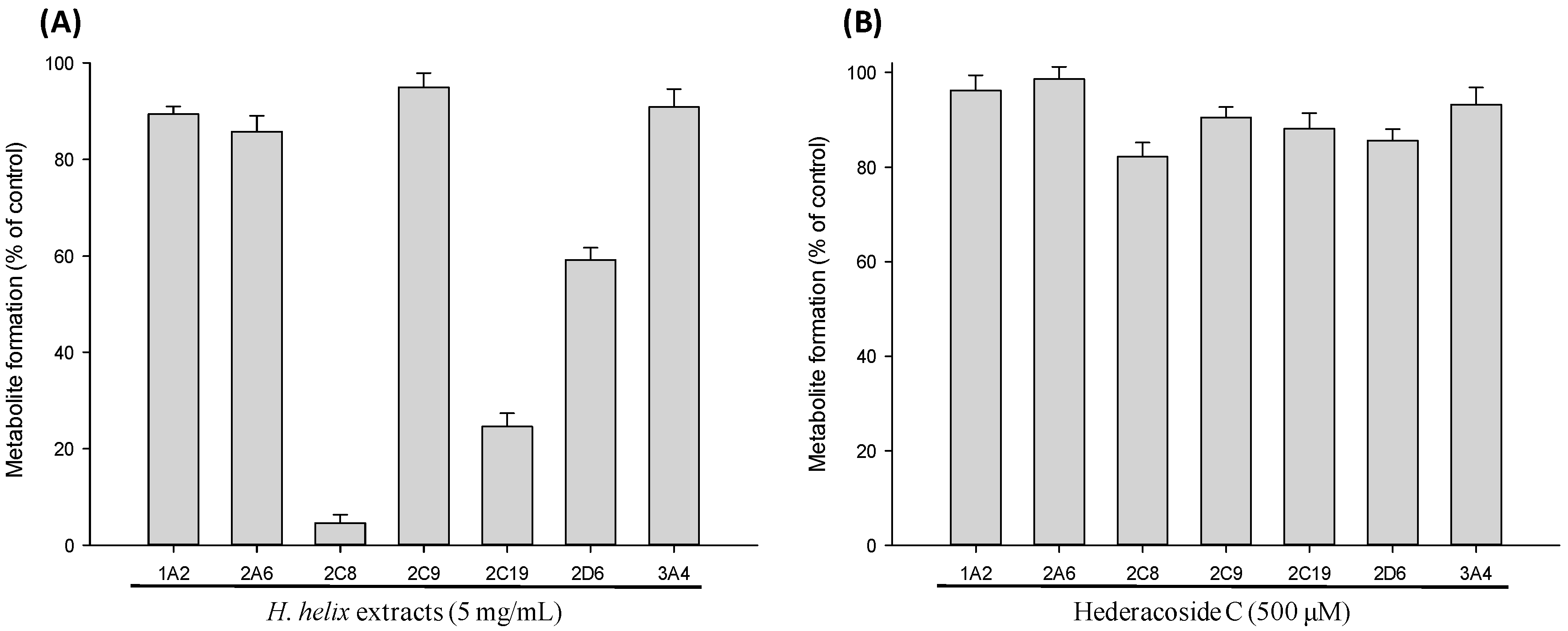
2.2. CYP Inhibition Assay in cDNA-Expressed Recombinant CYP Isozymes
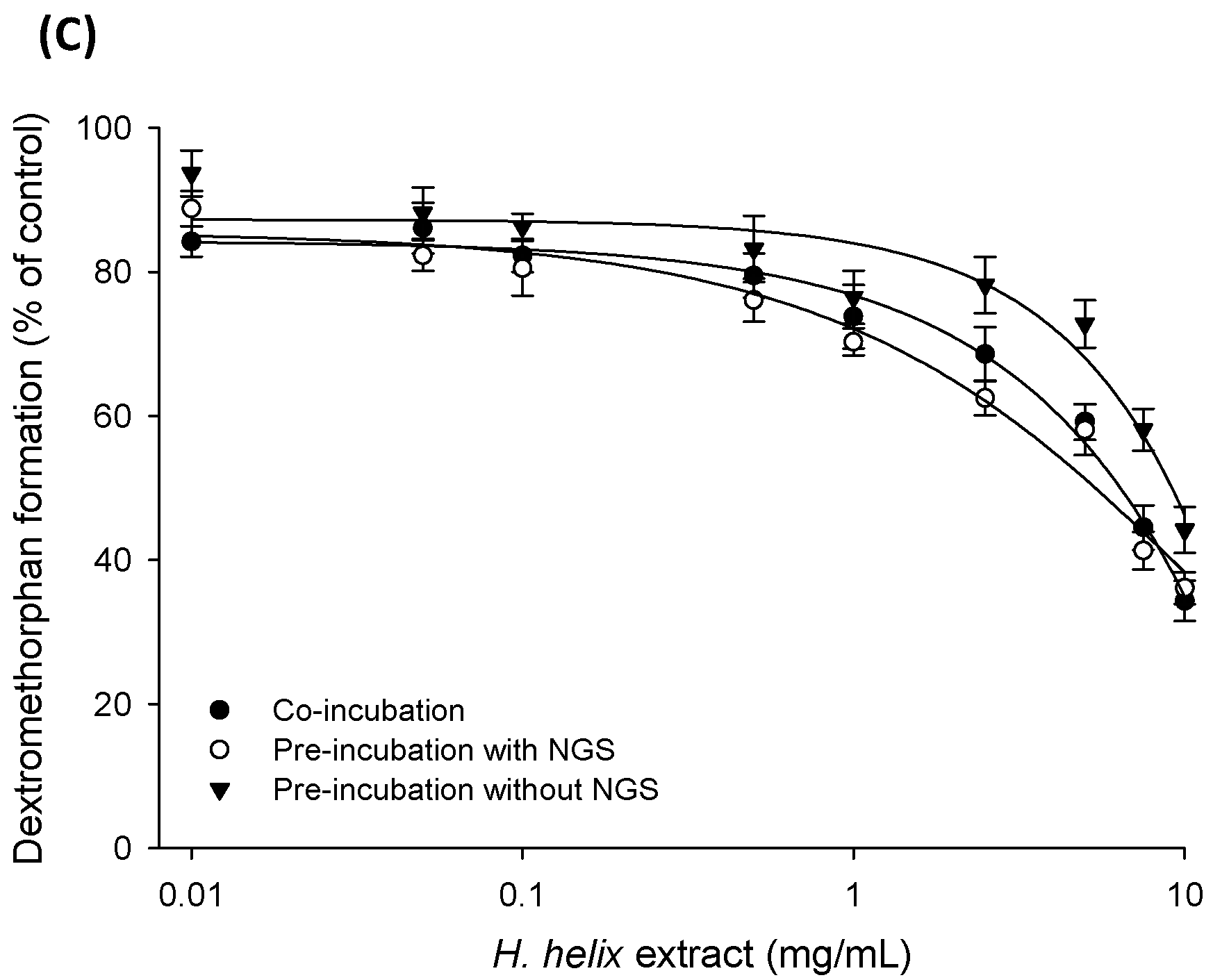
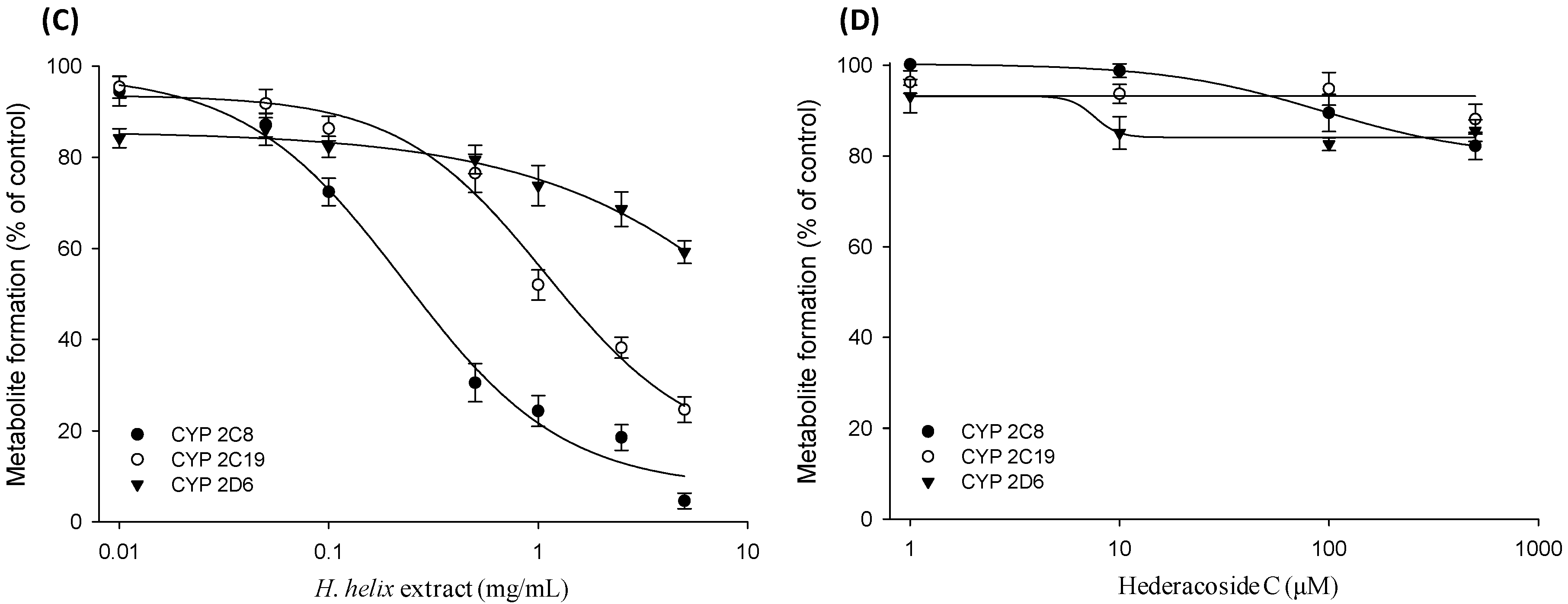
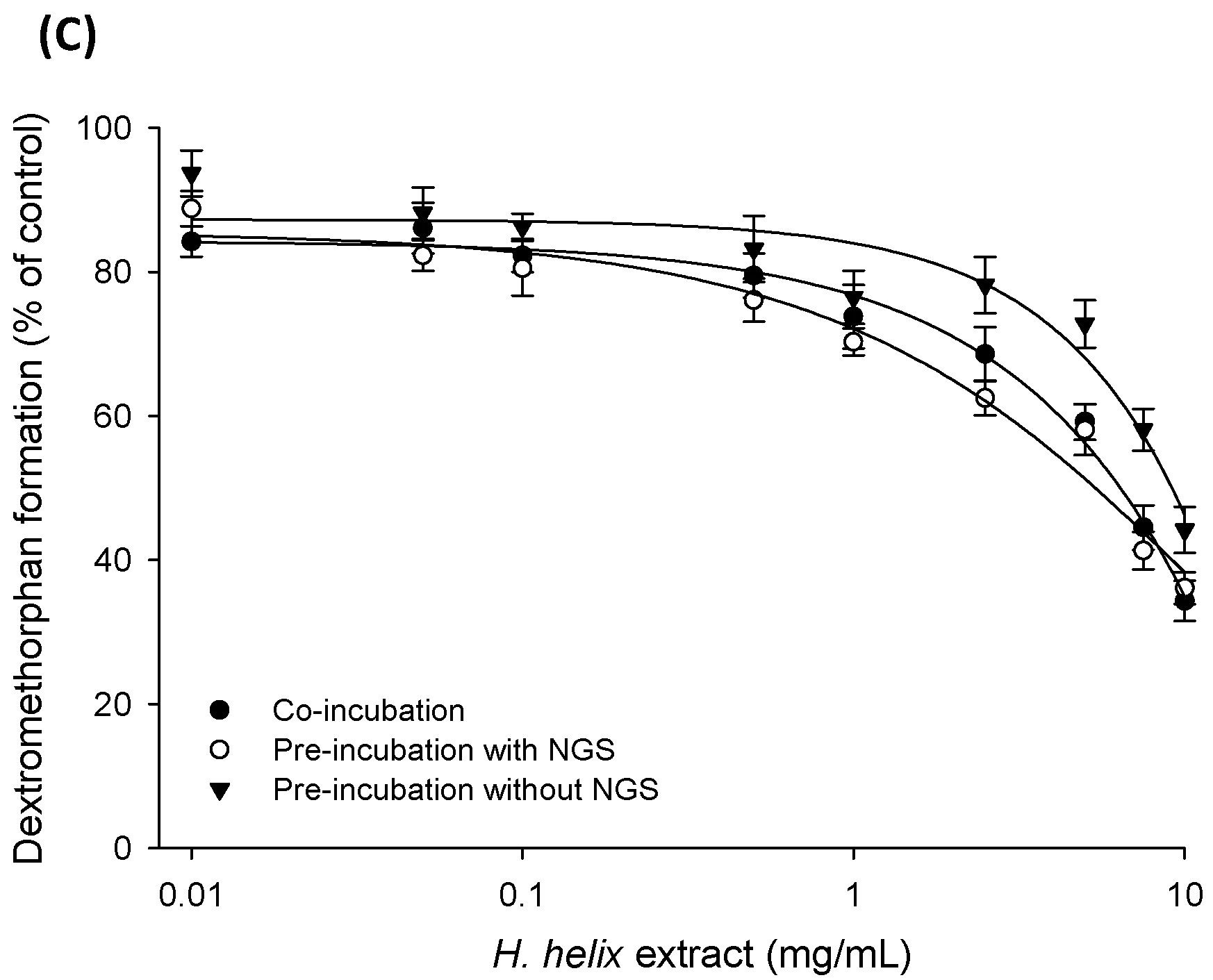
2.3. Time-Dependent Inhibition (TDI) and Mechanism-based Inactivation (MBI) Tests with Recombinant CYP2C8, CYP2C19 and CYP2D6 Isozymes
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
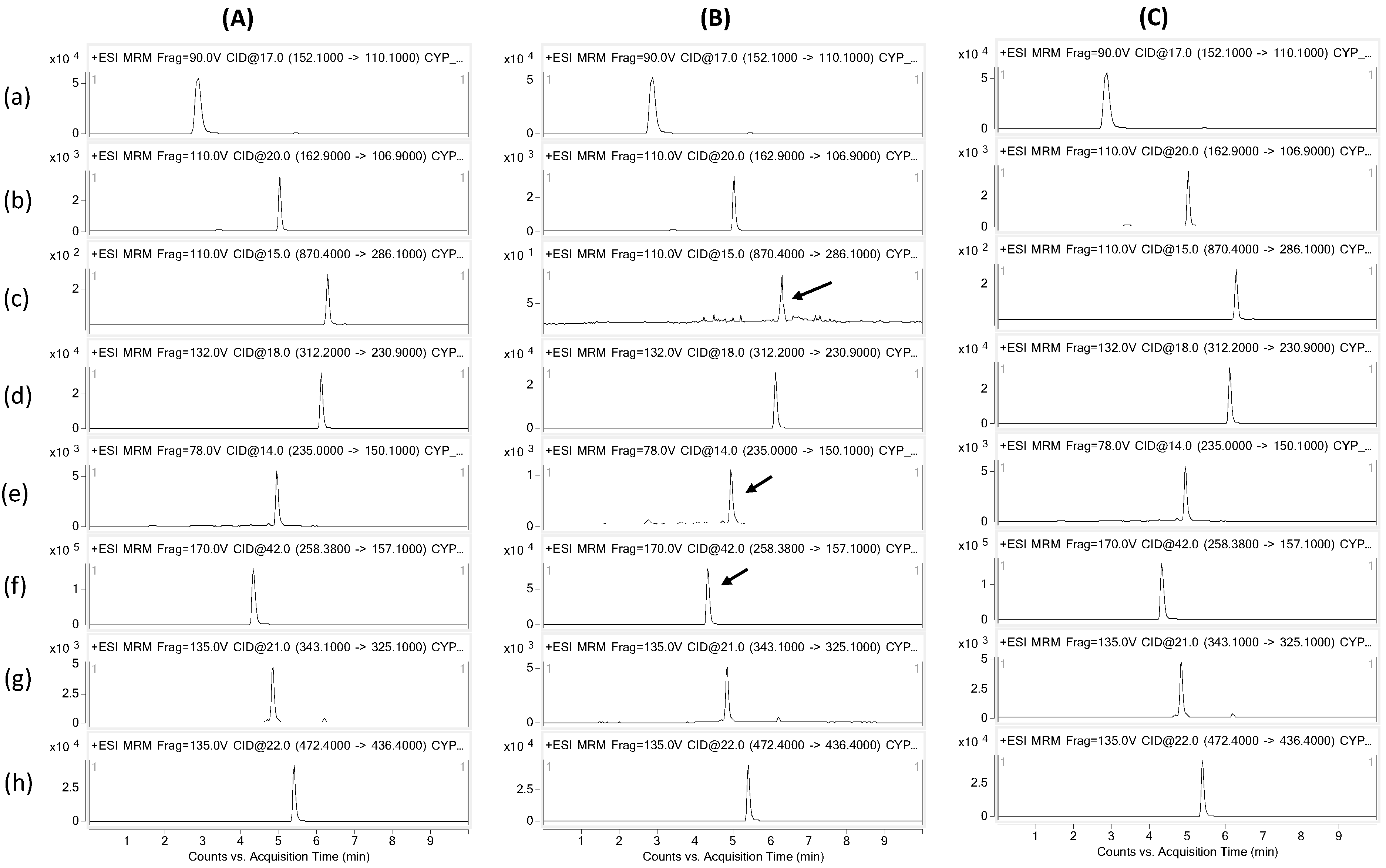
4.2. CYP Inhibition Assay
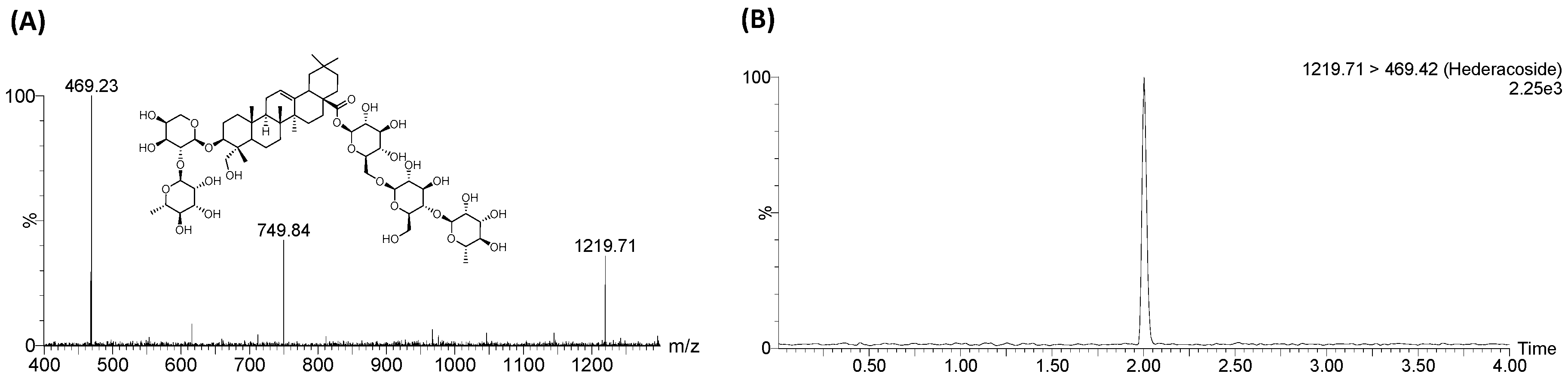
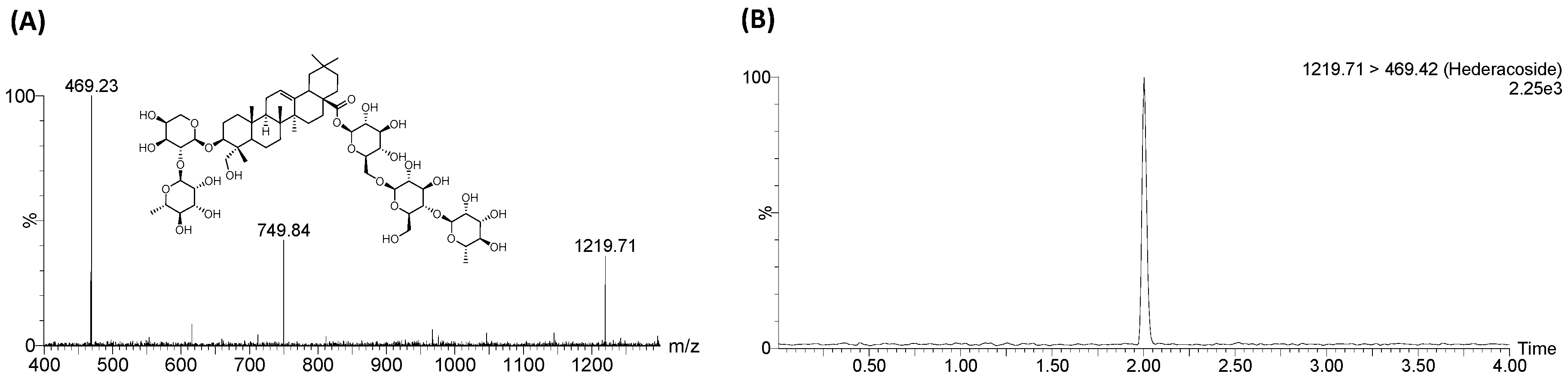
4.3. Sample Preparation
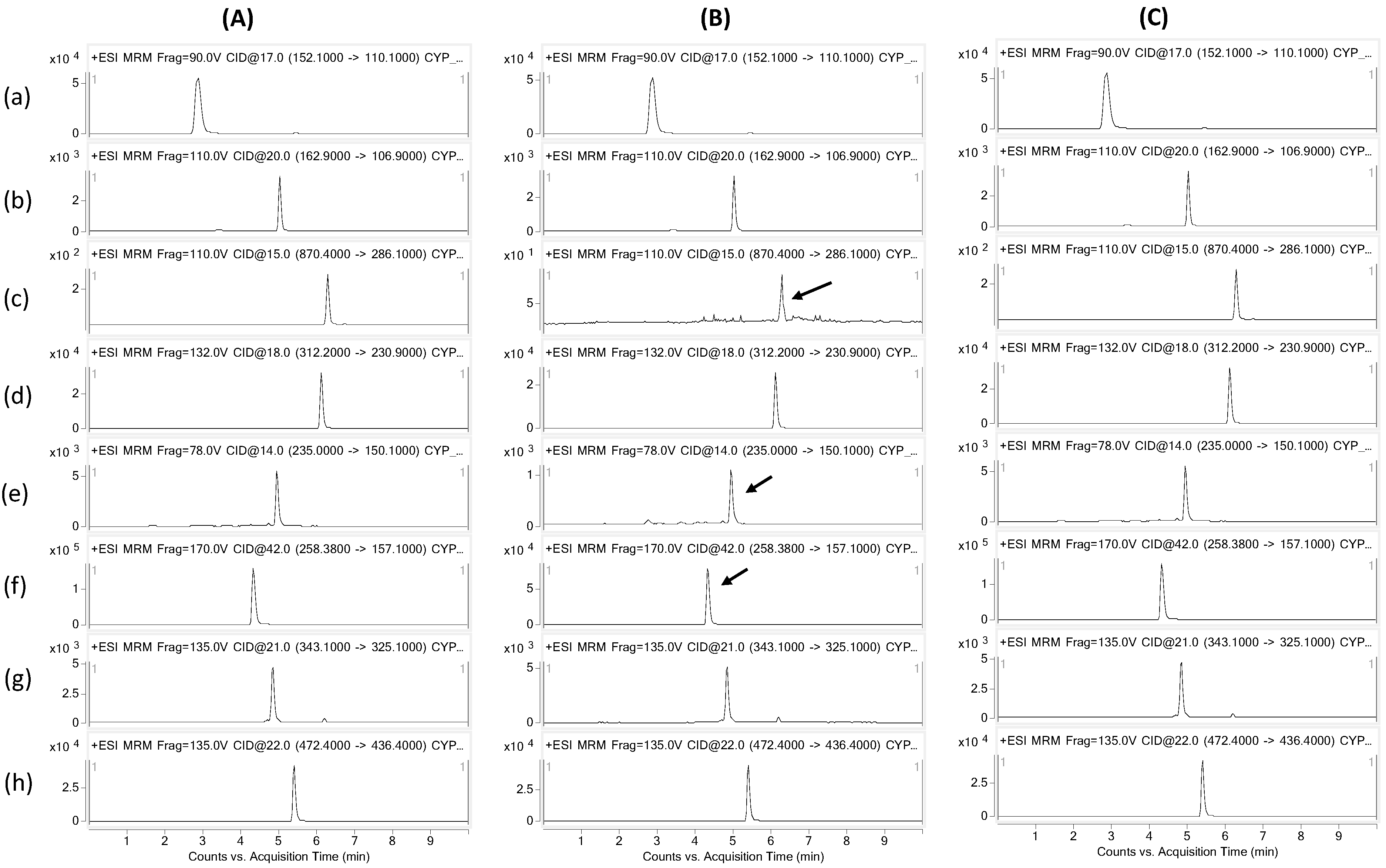
4.4. LC-MS/MS Analysis
4.5. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Natural Medicines Comprehensive Database. Available online: http://naturaldatabase.therapeuticresearch.com/home.aspx?cs=&s=ND (accessed on 2 July 2017).
- Cioaca, C.; Margineanu, C.; Cucu, V. The saponins of Hedera helix with antibacterial activity. Pharmazie 1978, 33, 609–610. [Google Scholar] [PubMed]
- Wolf, A.; Gosens, R.; Meurs, H.; Haberlein, H. Pre-treatment with α-hederin increases β-adrenoceptor mediated relaxation of airway smooth muscle. Phytomedicine 2011, 18, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Stauss-Grabo, M.; Atiye, S.; Warnke, A.; Wedemeyer, R.S.; Donath, F.; Blume, H.H. Observational study on the tolerability and safety of film-coated tablets containing ivy extract (Prospan® Cough Tablets) in the treatment of colds accompanied by coughing. Phytomedicine 2011, 18, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Fazio, S.; Pouso, J.; Dolinsky, D.; Fernandez, A.; Hernandez, M.; Clavier, G.; Hecker, M. Tolerance, safety and efficacy of Hedera helix extract in inflammatory bronchial diseases under clinical practice conditions: a prospective, open, multicentre postmarketing study in 9657 patients. Phytomedicine 2009, 16, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Trute, A.; Gross, J.; Mutschler, E.; Nahrstedt, A. In vitro antispasmodic compounds of the dry extract obtained from Hedera helix. Planta Med. 1997, 63, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Capasso, A.; Pinto, A.; Mascolo, N.; Capasso, F. Reduction of agonist-induced contractions of guinea-pig isolated ileum by flavonoids. Phytother. Res. 1991, 5, 85–87. [Google Scholar] [CrossRef]
- Hegener, O.; Prenner, L.; Runkel, F.; Baader, S.L.; Kappler, J.; Haberlein, H. Dynamics of β2-adrenergic receptor−ligand complexes on living cells. Biochemistry 2004, 43, 6190–6199. [Google Scholar] [CrossRef] [PubMed]
- Sieben, A.; Prenner, L.; Sorkalla, T.; Wolf, A.; Jakobs, D.; Runkel, F.; Haberlein, H. α-Hederin, but not hederacoside c and hederagenin from hedera helix, affects the binding behavior, dynamics, and regulation of β2-adrenergic receptors. Biochemistry 2009, 48, 3477–3482. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.U.; Choi, M.S.; Kim, I.S.; Kim, S.H.; Yoo, H.H. An ultra-high-performance liquid chromatography-tandem mass spectrometric method for the determination of hederacoside C, a drug candidate for respiratory disorder, in rat plasma. J. Pharmaceut. Biomed. 2016, 129, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.U.; Choi, M.S.; Choe, K.; Yoo, H.H. Interactions between herbs and antidiabetics: an overview of the mechanisms, evidence, importance, and management. Arch. Pharm. Res. 2015, 37, 1281–1298. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.U.; Choi, M.S.; Kim, I.S.; Luo, Z.; Xue, Y.; Yao, G.; Zhang, Y.; Yoo, H.H. In Vitro Assessment of CYP-Mediated Drug Interactions for Kinsenoside, an Antihyperlipidemic Candidate. Molecules 2016, 21, 800. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.I.; Maffei, M. Plant Specialized Metabolism: Genomics, Biochemistry, and Biological Functions, 1st ed.; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Assessment report on Hedera helix, L., folium, Committee on Herbal Medicinal Products (HMPC), European Medicines Agency (EMA). EMA/HMPC/586887/2014. Available online: http://www.ema.europa.eu/docs/en_GB/document_library/Herbal_HMPC_assessment_report/2016/01/WC500199887.pdf (accessed on 24 November 2015).
- Liu, J.; Liu, Y.P.; Bullock, P.; KlaasseN, C.D. Suppression of liver cytochrome P450 by α-hederin: relevance to hepatoprotection. Toxicol. Appl. Pharm. 1995, 134, 124–131. [Google Scholar] [CrossRef]
- Jeong, H.G. Suppression of constitutive and inducible cytochrome P450 gene expression by α-hederin in mice. IUBMB Life 1998, 46, 1019–1026. [Google Scholar] [CrossRef]
- Nebert, D.W.; Russell, D.W. Clinical importance of the cytochromes P450. Lancet 2002, 360, 1155–1162. [Google Scholar] [CrossRef]
- Nomeir, A.A.; Palamanda, J.R.; Favreau, L. Identification of CYP mechanism-based inhibitors. In Optimization in Drug Discovery: In Vitro Methods, 1st ed.; Humana Press: Totowa, NJ, USA, 2004; pp. 245–262. ISBN 978-1-62703-742-6. [Google Scholar]
- Obach, R.S.; Walsky, R.L.; Venkatakrishnan, K. Mechanism-based inactivation of human cytochrome p450 enzymes and the prediction of drug-drug interactions. Drug Metab. Dispos. 2007, 35, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Riley, R.J.; Grime, K.; Weaver, R. Time-dependent CYP inhibition. Expert Opin. Drug Met. 2007, 3, 51–66. [Google Scholar] [CrossRef] [PubMed]
- Stresser, D.M.; Mao, J.; Kenny, J.R.; Jones, B.C.; Grime, K. Exploring concepts of in vitro time-dependent CYP inhibition assays. Expert Opin. Drug Met. 2014, 10, 157–174. [Google Scholar] [CrossRef] [PubMed]
- Orr, S.T.; Ripp, S.L.; Ballard, T.E.; Henderson, J.L.; Scott, D.O.; Obach, R.S.; Sun, H.; Kalgutkar, A.S. Mechanism-based inactivation (MBI) of cytochrome P450 enzymes: structure–activity relationships and discovery strategies to mitigate drug–drug interaction risks. J. Med. Chem. 2012, 55, 4896–4933. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration (FDA), Drug Development and Drug Interactions: Table of Substrates, Inhibitors and Inducers. Available online: http://www.fda.gov/Drugs/DevelopmentApprovalProcess/DevelopmentResources/DrugInteractionsLabeling/ucm093664.htm (accessed on 11 July 2017).
- Grimm, S.W.; Einolf, H.J.; Hall, S.D.; He, K.; Lim, H.K.; Ling, K.H.J.; Lu, C.; Nomeir, A.A.; Seibert, E.; Skordos, K.W.; et al. The conduct of in vitro studies to address time-dependent inhibition of drug-metabolizing enzymes: a perspective of the pharmaceutical research and manufacturers of America. Drug Metab. Dispos. 2009, 37, 1355–1370. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.U.; Choe, K.; Yoo, H.H. Review on a traditional herbal medicine, Eurycoma longifolia jack (Tongkat Ali): its traditional uses, chemistry, evidence-based pharmacology and toxicology. Molecules 2016, 21, 331. [Google Scholar] [CrossRef] [PubMed]
- Kamel, A.; Harriman, S. Inhibition of cytochrome P450 enzymes and biochemical aspects of mechanism-based inactivation (MBI). Drug Discov. Today Technol. 2013, 10, e177–e189. [Google Scholar] [CrossRef] [PubMed]
- Brahmi, Z.; Katho, T.; Hatsumata, R.; Hiroi, A.; Miyakawa, N.; Yakou, E.; Sugaya, K.; Onose, J.I.; Naoki, A.B.E. Effective Cytochrome P450 (CYP) Inhibitors Isolated from Tarragon (Artemisia dracunculus). Biosci. Biotechnol. Biochem. 2012, 76, 1028–1031. [Google Scholar] [CrossRef] [PubMed]
- Taesotikul, T.; Dumrongsakulchai, W.; Wattanachai, N.; Navinpipat, V.; Somanabandhu, A.; Tassaneeyakul, W.; Tassaneeyakul, W. Inhibitory effects of Phyllanthus amarus and its major lignans on human microsomal cytochrome P450 activities: evidence for CYP3A4 mechanism-based inhibition. Drug Metab. Pharmacokinet. 2011, 26, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Hellum, B.H.; Liang, A.; Nilsen, O.G. Inhibitory mechanisms of human CYPs by three alkaloids isolated from traditional chinese herbs. Phytother. Res. 2015, 29, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.M.; Kim, I.S.; Rehman, S.U.; Choe, K.; Yoo, H.H. In Vitro Evaluation of the Effects of Eurycoma longifolia Extract on CYP-Mediated Drug Metabolism. Evid. Based Complement. Alternat. Med. 2015, 631329. [Google Scholar] [CrossRef]
Sample Availability: Not available. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P450 Isozyme | PROBE Substrate | Substrate Conc. (µM) | Metabolite Monitored | Precursor-Ion (m/z) | Daughter-Ion (m/z) |
|---|---|---|---|---|---|
| CYP1A2 | Phenacetin | 40 | Acetaminophen | 152.1 | 110.1 |
| CYP2A6 | Coumarin | 2.5 | 7-OH-coumarin | 162.9 | 106.9 |
| CYP2C8 | Paclitaxel | 10 | 6-OH-paclitaxel | 870.4 | 286.1 |
| CYP2C9 | Diclofenac | 10 | 4-OH-diclofenac | 312.2 | 230.9 |
| CYP2C19 | Mephenytoin | 160 | 4-OH-mephenytoin | 235.0 | 150.1 |
| CYP2D6 | Dextromethorphan | 5 | Dextrorphan | 258.3 | 157.1 |
| CYP3A4 | Midazolam | 2.5 | 1-OH-midazolam | 343.1 | 325.1 |
| Internal Standard | - | Terfenadine | 472.4 | 436.4 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rehman, S.U.; Kim, I.S.; Choi, M.S.; Kim, S.H.; Zhang, Y.; Yoo, H.H. Time-dependent Inhibition of CYP2C8 and CYP2C19 by Hedera helix Extracts, A Traditional Respiratory Herbal Medicine. Molecules 2017, 22, 1241. https://doi.org/10.3390/molecules22071241
Rehman SU, Kim IS, Choi MS, Kim SH, Zhang Y, Yoo HH. Time-dependent Inhibition of CYP2C8 and CYP2C19 by Hedera helix Extracts, A Traditional Respiratory Herbal Medicine. Molecules. 2017; 22(7):1241. https://doi.org/10.3390/molecules22071241
Chicago/Turabian StyleRehman, Shaheed Ur, In Sook Kim, Min Sun Choi, Seung Hyun Kim, Yonghui Zhang, and Hye Hyun Yoo. 2017. "Time-dependent Inhibition of CYP2C8 and CYP2C19 by Hedera helix Extracts, A Traditional Respiratory Herbal Medicine" Molecules 22, no. 7: 1241. https://doi.org/10.3390/molecules22071241
APA StyleRehman, S. U., Kim, I. S., Choi, M. S., Kim, S. H., Zhang, Y., & Yoo, H. H. (2017). Time-dependent Inhibition of CYP2C8 and CYP2C19 by Hedera helix Extracts, A Traditional Respiratory Herbal Medicine. Molecules, 22(7), 1241. https://doi.org/10.3390/molecules22071241