New Cinchona Oximes Evaluated as Reactivators of Acetylcholinesterase and Butyrylcholinesterase Inhibited by Organophosphorus Compounds




Abstract
:1. Introduction
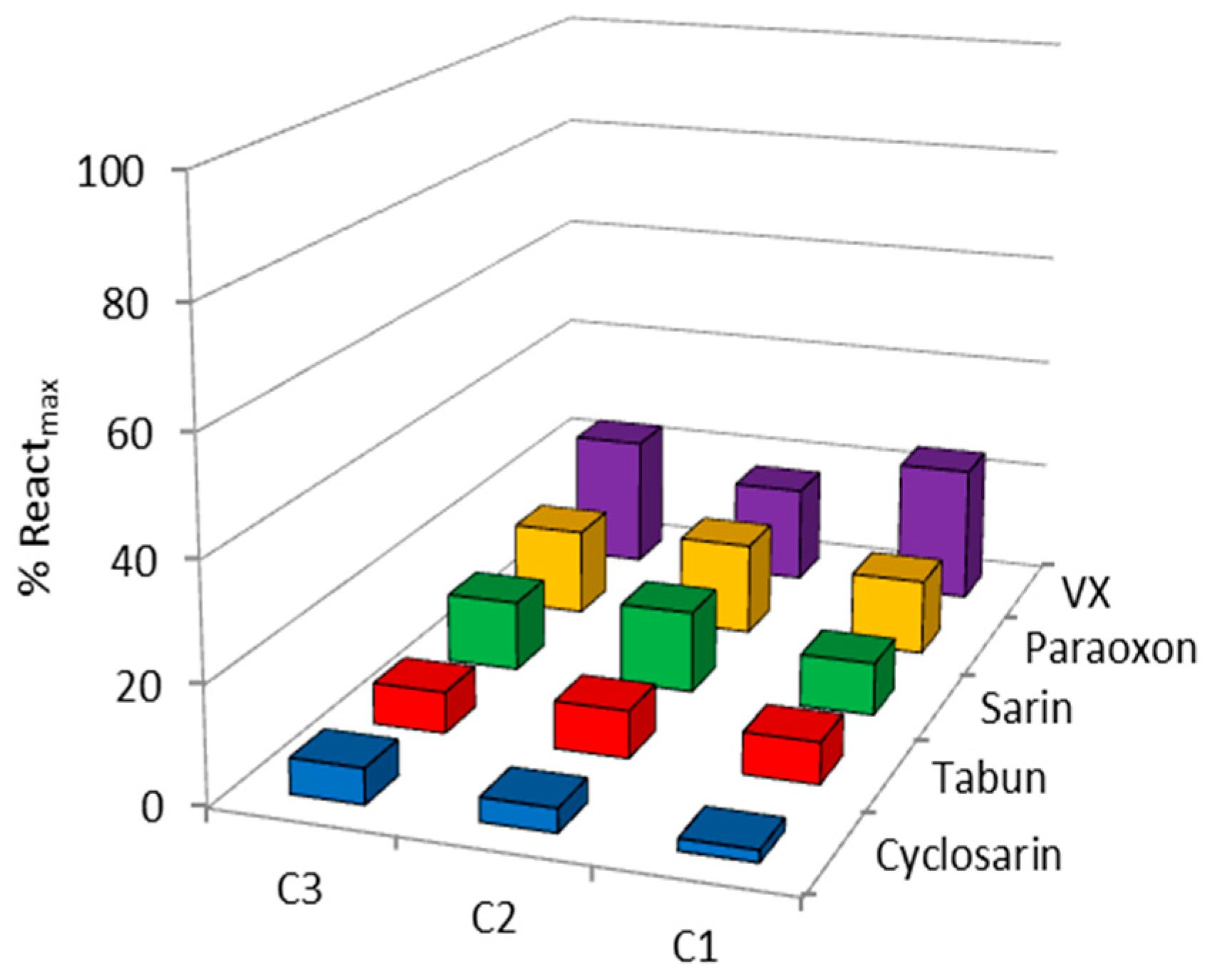
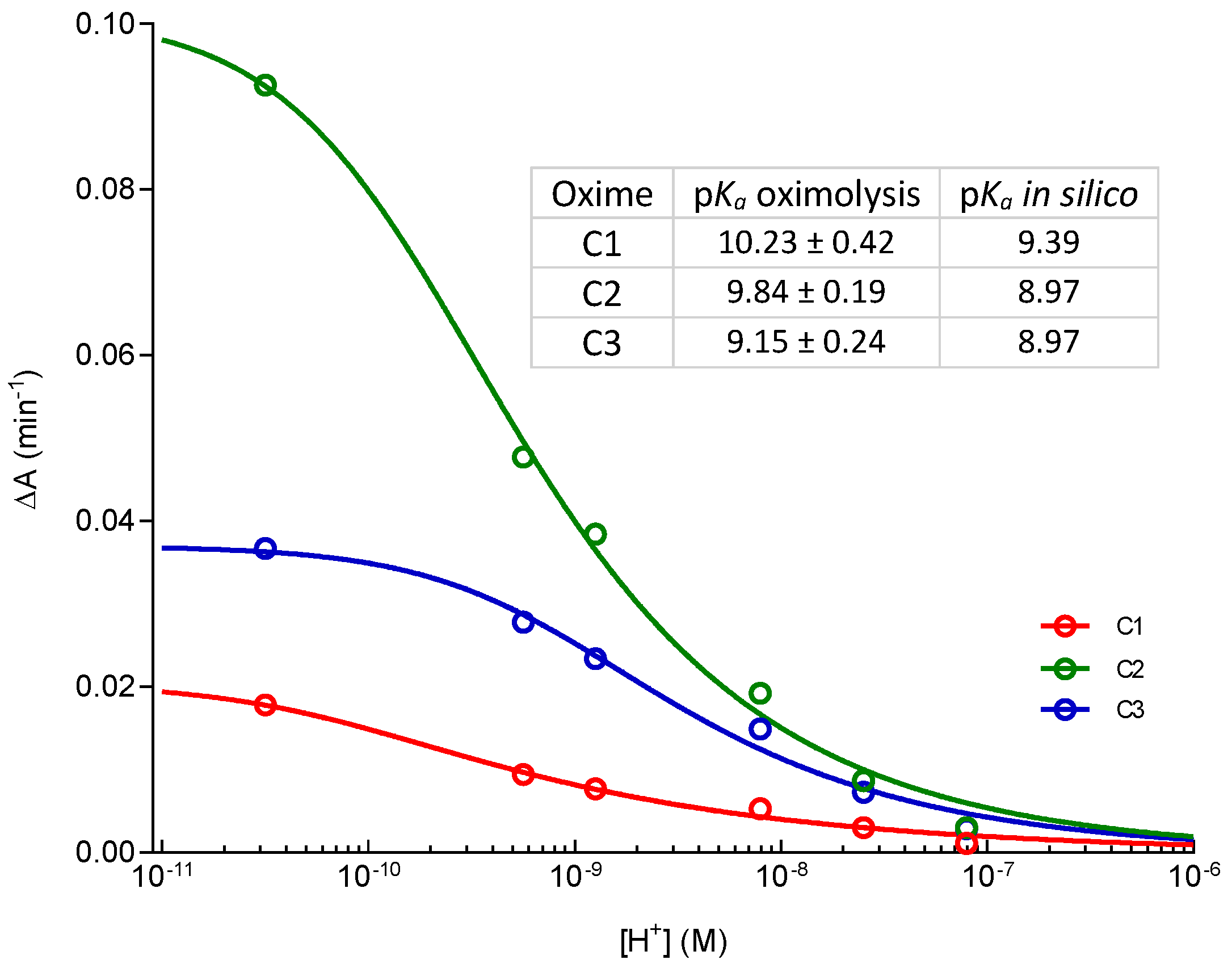
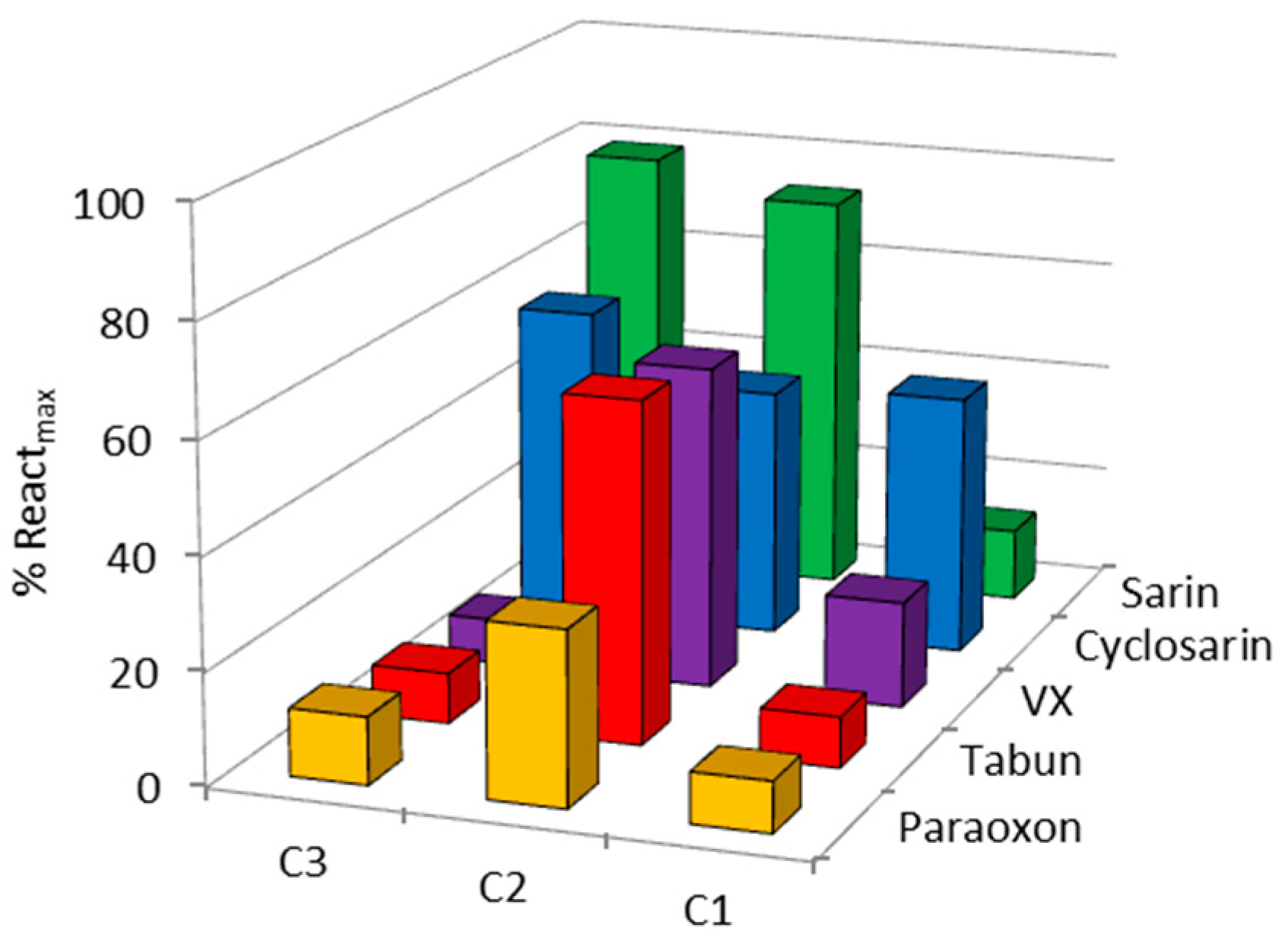
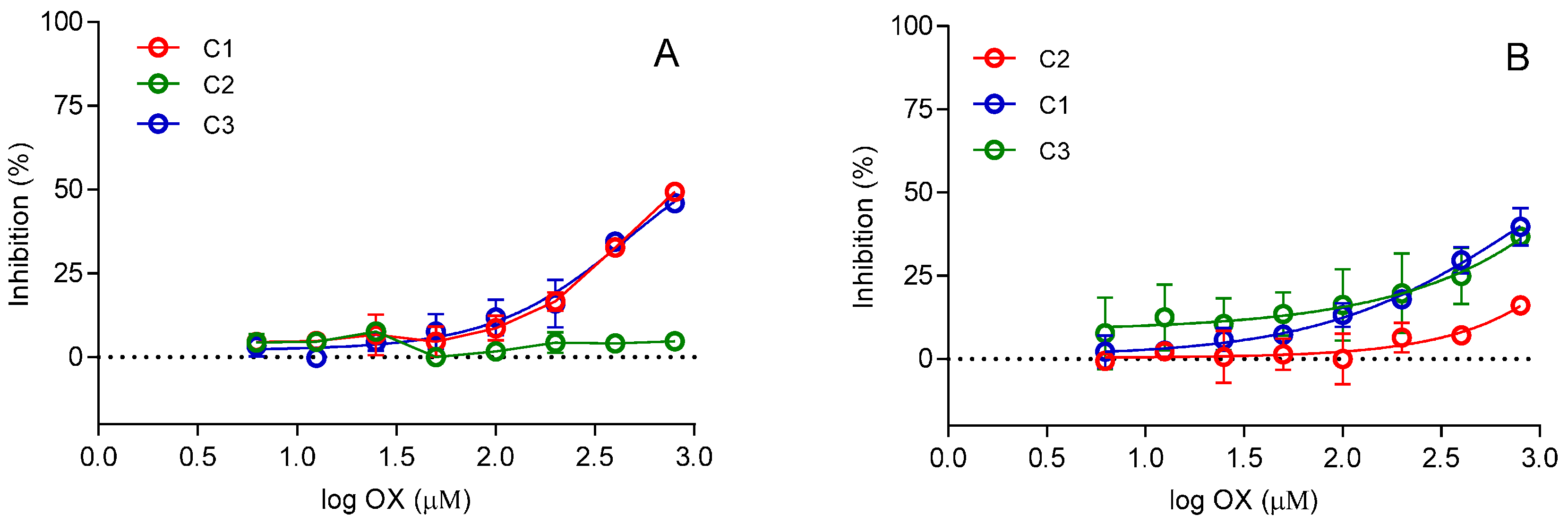
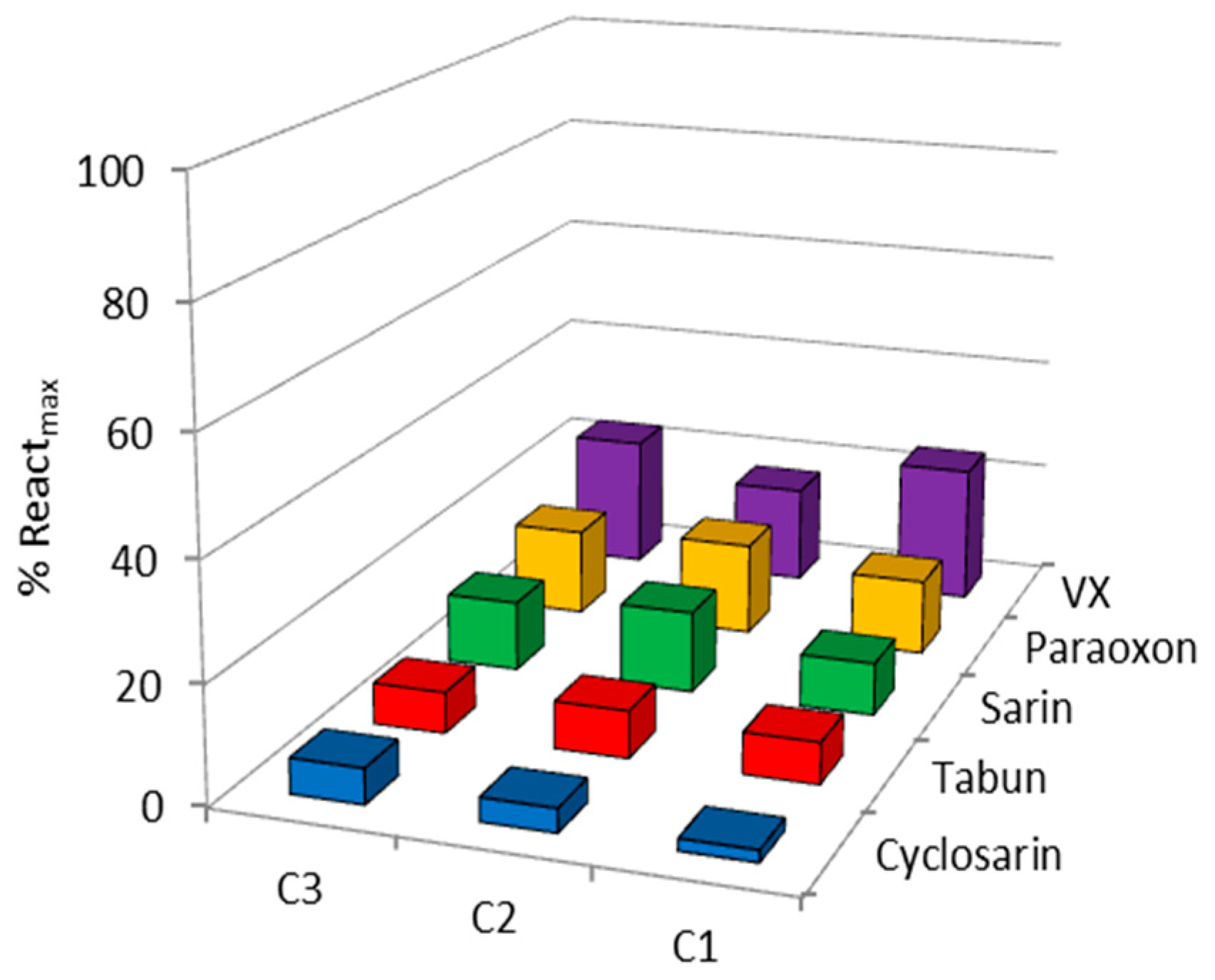
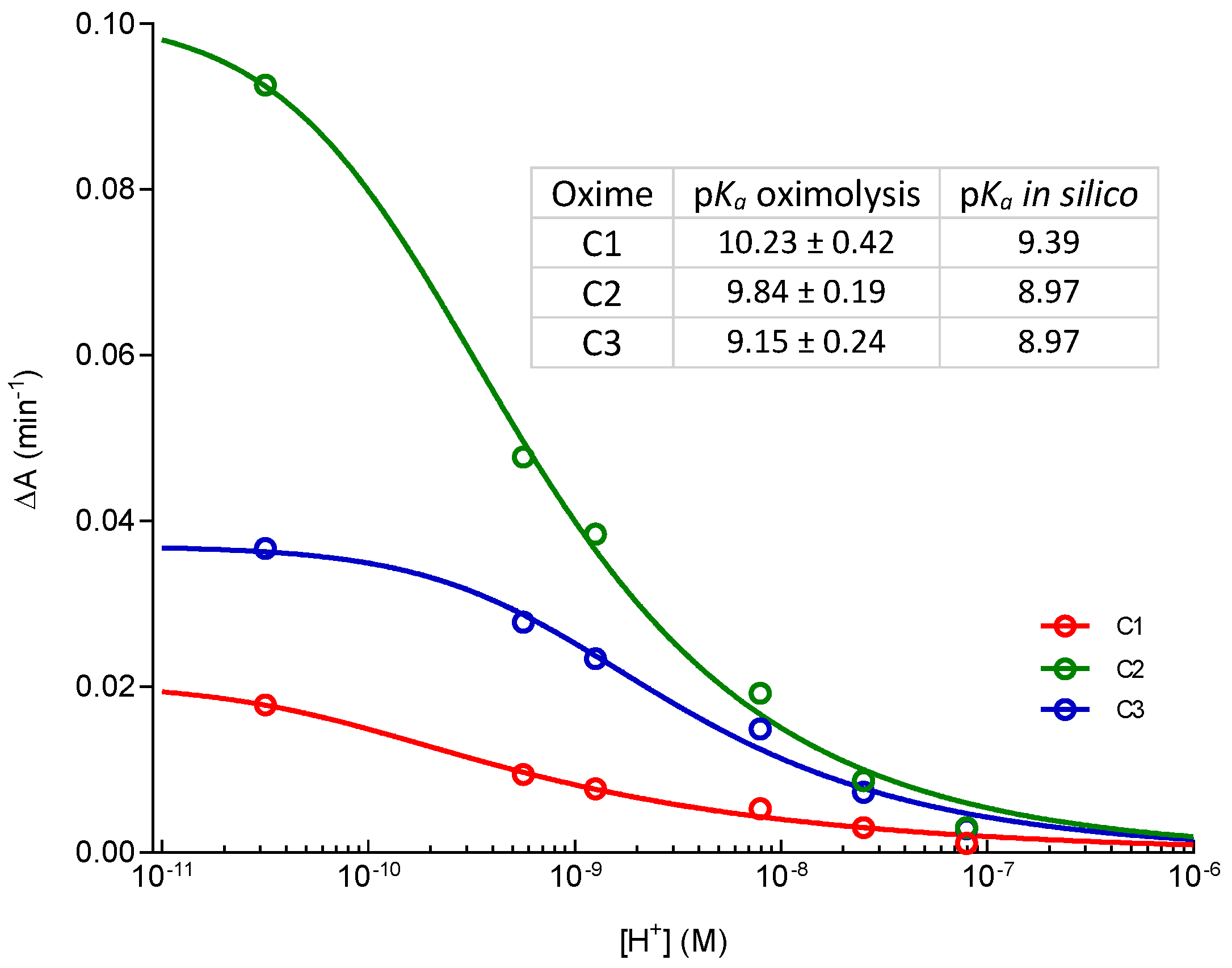
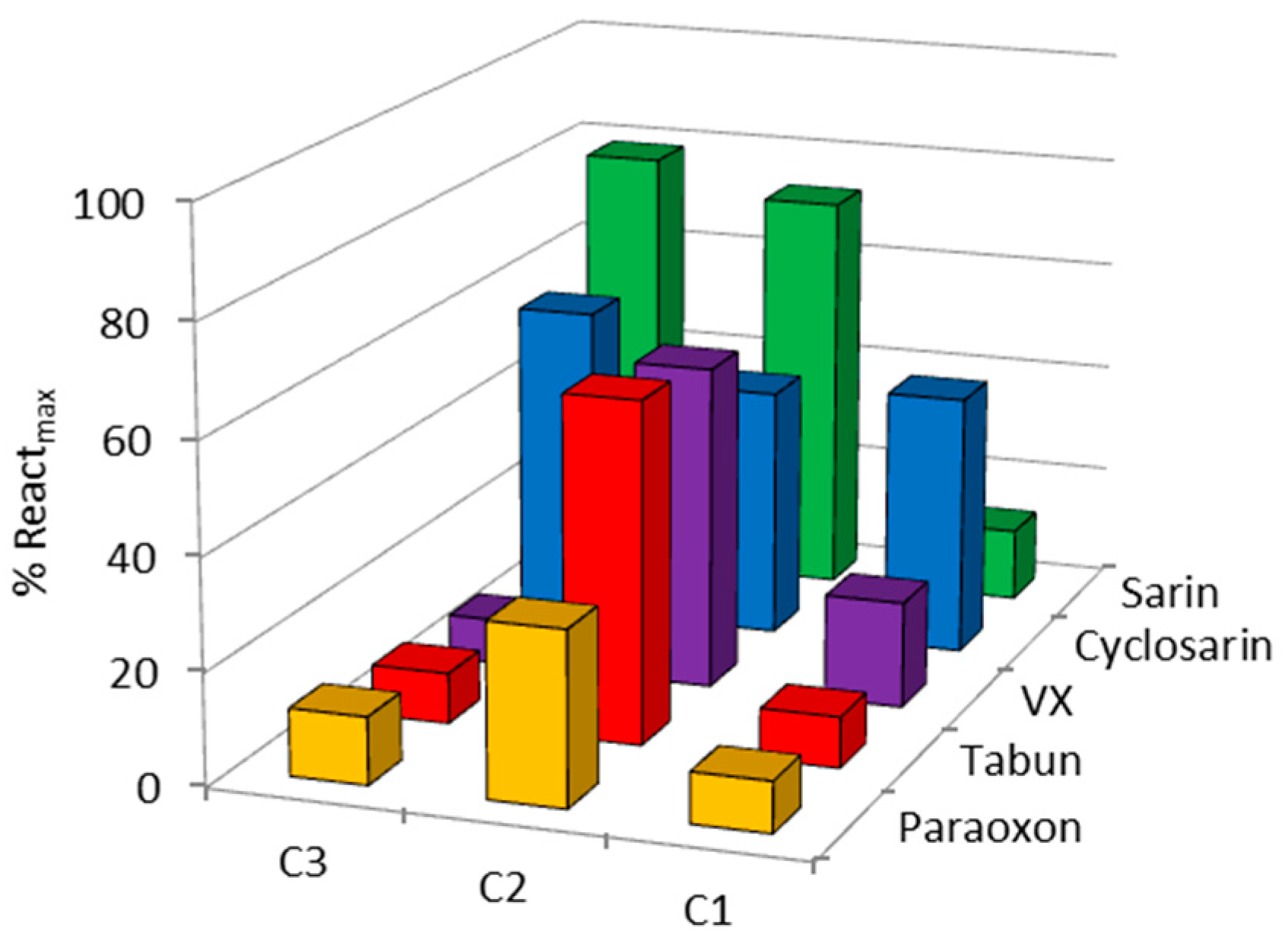
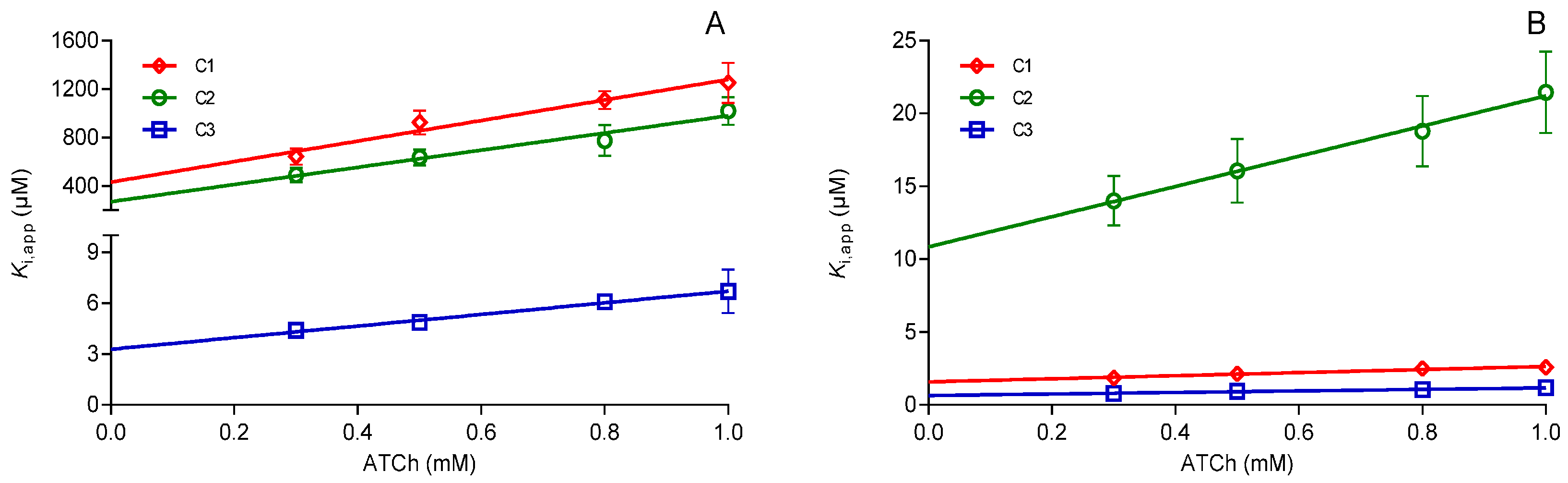
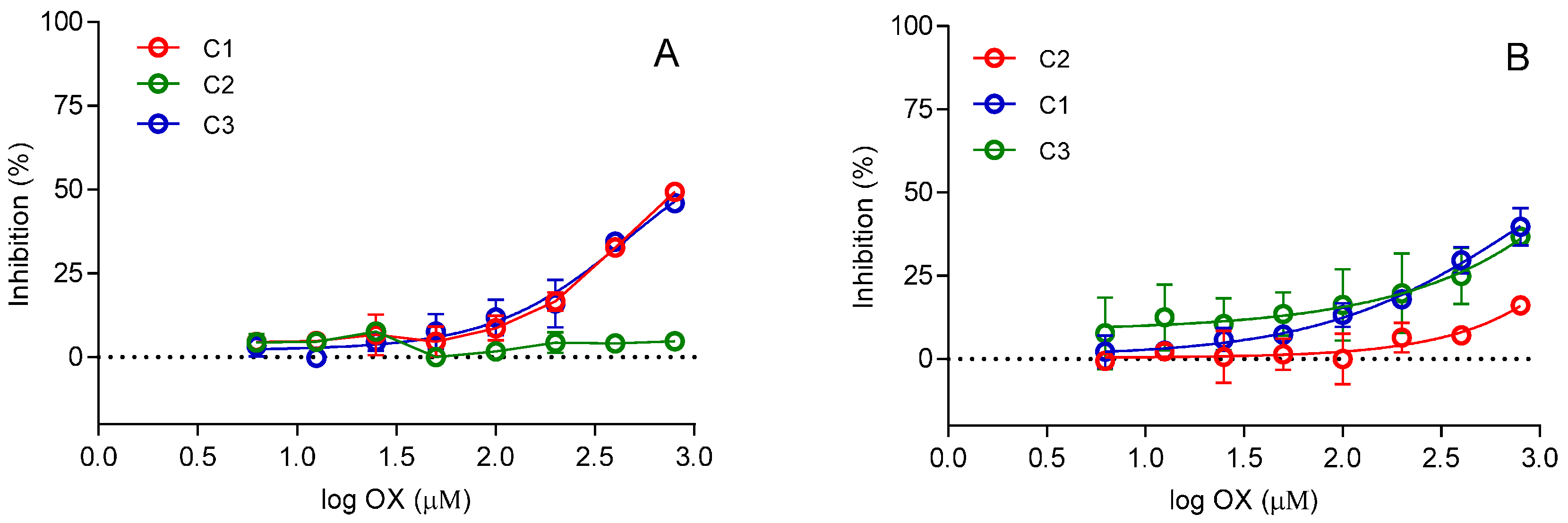
2. Results and Discussion
3. Materials and Methods
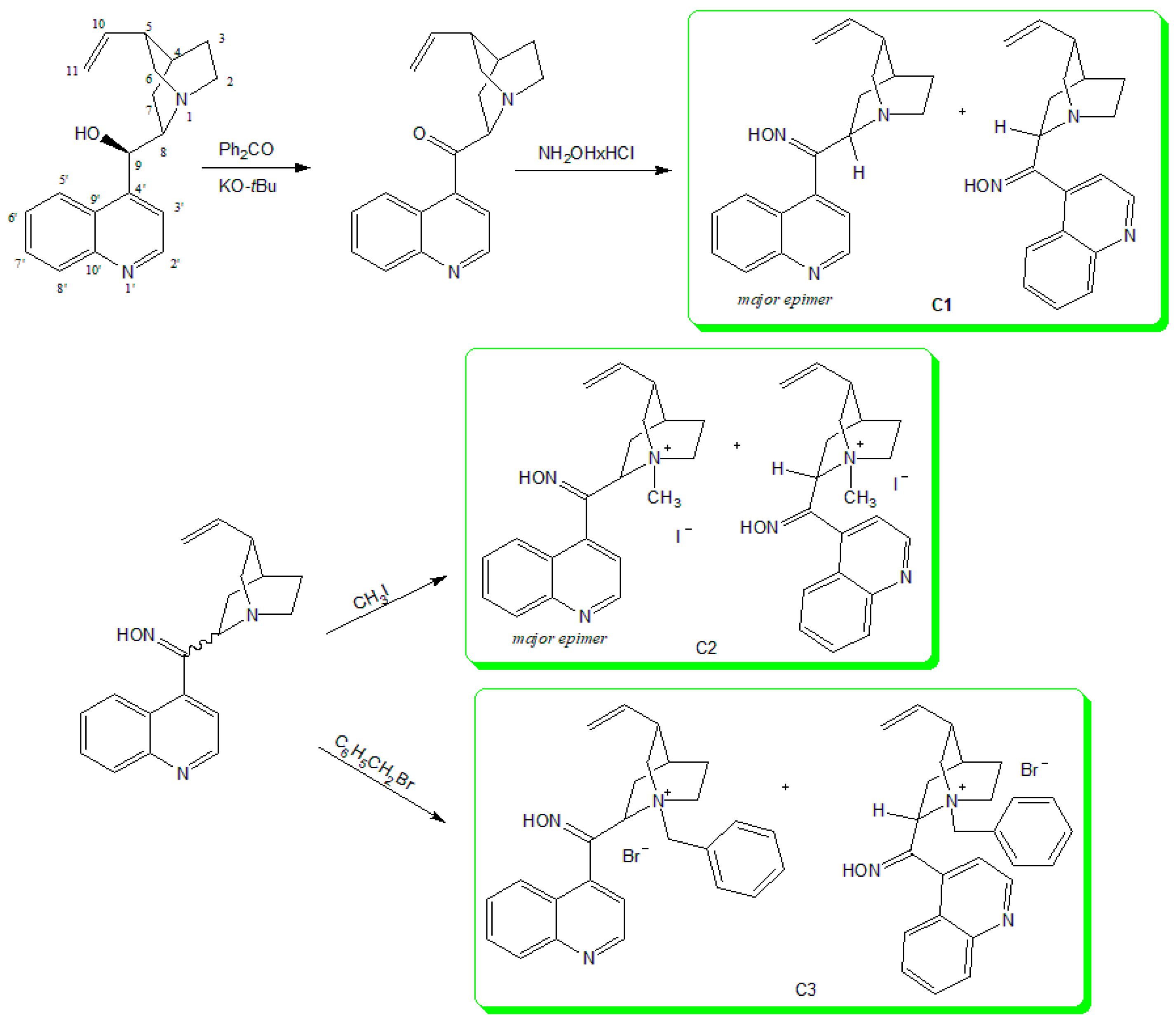
3.1. Synthesis of Cinchona Oximes
3.2. Quantum Chemical Calculations
3.3. Chemicals
3.4. pKa Determination
3.5. Determination of Oxime Inhibition Constants
3.6. Oxime-Assisted Reactivation of OP-Inhibited Cholinesterases
3.7. Cytotoxicity Screening
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bajgar, J. Organophosphates/nerve agent poisoning: Mechanism of action, diagnosis, prophylaxis and treatment. Adv. Clin. Chem. 2004, 38, 151–216. [Google Scholar] [PubMed]
- Stojiljković, M.P.; Jokanović, M. Pyridinium oximes: Rationale for their selection as causal antidotes against organophosphate poisonings and current solutions for auto-injectors. Arh. Hig. Rada Toksikol. 2006, 57, 435–443. [Google Scholar] [PubMed]
- Worek, F.; Thiermann, H.; Wille, T. Oximes in organophosphate poisoning: 60 years of hope and despair. Chem. Biol. Interact. 2016, 259, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Katalinić, M.; Maček Hrvat, N.; Ždarova Karasova, J.; Misik, J.; Kovarik, Z. Translation of in vitro to in vivo pyridinium oxime potential in tabun poisoning. Arh. Ind. Hig. Rada Toxicol. 2015, 66, 285–292. [Google Scholar]
- Kovarik, Z.; Radić, Z.; Berman, H.A.; Simeon-Rudolf, V.; Reiner, E.; Taylor, P. Mutant cholinesterases possessing enhanced capacity for reactivation of their phosphonylated conjugates. Biochemistry 2004, 43, 3222–3229. [Google Scholar] [CrossRef] [PubMed]
- Kovarik, Z.; Čalić, M.; Šinko, G.; Bosak, A.; Berend, S.; Lucić Vrdoljak, A.; Radić, B. Oximes: Reactivators of phosphorylated acetylcholinesterase and antidotes in therapy against tabun poisoning. Chem. Biol. Interact. 2008, 175, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Worek, F.; Thiermann, H. The value of novel oximes for treatment of poisoning by organophosphorus compounds. Pharmacol. Ther. 2013, 139, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Renou, J.; Loiodice, M.; Arboléas, M.; Baati, R.; Jean, L.; Nachon, F.; Renard, P.Y. Tryptoline-3-hydroxypyridinaldoxime conjugates as efficient reactivators of phosphylated human acetyl and butyrylcholinesterases. Chem. Commun. 2014, 50, 3947–3950. [Google Scholar] [CrossRef] [PubMed]
- De Koning, M.C.; van Grol, M.; Noort, D. Peripheral site ligand conjugation to a non-quaternary oxime enchances reactivation of nerve agent-inhibited human acetylcholinesterase. Toxicol. Lett. 2011, 206, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Radić, Z.; Sit, R.K.; Garcia, E.; Zhang, L.; Berend, S.; Kovarik, Z.; Amitai, G.; Fokin, V.V.; Sharpless, K.B.; Taylor, P. Mechanism of interaction of novel uncharged, centrally active reactivators with OP-hAChE conjugates. Chem. Biol. Interact. 2013, 203, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Kovarik, Z.; Maček, N.; Sit, R.K.; Radić, Z.; Fokin, V.V.; Sharpless, K.B.; Taylor, P. Centrally acting oximes in reactivation of tabun-phosphoramidated AChE. Chem. Biol. Interact. 2013, 203, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Mercey, G.; Verdelet, T.; Renou, J.; Kliachyna, M.; Baati, R.; Nachon, F.; Jean, L.; Renard, P.-Y. Reactivators of acetylcholinesterase inhibited by organophosphorus nerve agents. Acc. Chem. Res. 2012, 45, 756–766. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, D.; Fallarero, A.; Shinde, P.; Anju, C.P.; Busygin, I.; Leino, R.; Mohan, C.G.; Vuorela, P. Chemical modifications of Cinchona alkaloids lead to enhanced inhibition of human butyrylcholinesterase. Nat. Prod. Commun. 2014, 9, 455–458. [Google Scholar] [PubMed]
- Nawaz, S.A.; Ayaz, M.; Brandt, W.; Wessjohann, L.A.; Westermann, B. Cation–π and π–π stacking interactions allow selective inhibition of butyrylcholinesterase by modified quinine and cinchonidine alkaloids. Biochem. Biophys. Res. Commun. 2011, 404, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Robins, R.J.; Rhodes, M.J.C. An evaluation of the tautomerism of cinchoninone and quinidinonemade using a combination of 1H-NMR and 13C-NMR spectroscopy. Phytochemistry 1987, 26, 551–556. [Google Scholar] [CrossRef]
- Sit, R.K.; Fokin, V.V.; Amitai, G.; Sharpless, K.B.; Taylor, P.; Radić, Z. Imidazole aldoximes effective in assisting butyrylcholinesterase catalysis of organophosphate detoxification. J. Med. Chem. 2014, 57, 1378–1389. [Google Scholar] [CrossRef] [PubMed]
- Katalinić, M.; Hrvat, N.M.; Baumann, K.; Morasi Piperčić, S.; Makarić, S.; Tomić, S.; Jović, O.; Hrenar, T.; Miličević, A.; Jelić, D.; et al. A comprehensive evaluation of novel oximes in creation of butyrylcholinesterase-based nerve agent bioscavengers. Toxicol. Appl. Pharmacol. 2016, 310, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Kovarik, Z.; Katalinić, M.; Šinko, G.; Binder, J.; Holas, O.; Jung, Y.S.; Musilova, L.; June, D.; Kuča, K. Pseudo-catalytic scavenging: Searching for a suitable reactivator of phosphorylated butyrylcholinesterase. Chem. Biol. Interact. 2010, 187, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Sit, R.K.; Radić, Z.; Gerardi, V.; Zhang, L.; Garcia, E.; Katalinić, M.; Amitai, G.; Kovarik, Z.; Fokin, V.V.; Sharpless, K.B.; et al. New structural scaffolds for centrally acting oxime reactivators of phosphylated cholinesterases. J. Biol. Chem. 2011, 286, 19422–19430. [Google Scholar] [CrossRef] [PubMed]
- Radić, Z.; Dale, T.; Kovarik, Z.; Berend, S.; Garcia, E.; Zhang, L.; Amitai, G.; Green, C.; Radić, B.; Duggan, B.M.; et al. Catalytic detoxification of nerve agent and pesticide organophosphates by butyrylcholinesterase assisted with non-pyridinium oximes. Biochem. J. 2013, 450, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Horn, G.; Wille, T.; Musilek, K.; Kuca, K.; Thiermann, H.; Worek, F. Reactivation kinetics of 31 structurally different bispyridinium oximes with organophosphate-inhibited human butyrylcholinesterase. Arch. Toxicol. 2015, 89, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Čalić, M.; Vrdoljak, A.L.; Radić, B.; Jelić, D.; Jun, D.; Kuča, K.; Kovarik, Z. In vitro and in vivo evaluation of pyridinium oximes: Mode of interaction with acetylcholinesterase, effect on tabun- and soman-poisoned mice and their cytotoxicity. Toxicology 2006, 219, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Kovarik, Z.; Čalić, M.; Bosak, A.; Šinko, G.; Jelić, D. In vitro evaluation of aldoxime interactions with human acetylcholinesterase. Croat. Chem. Acta 2008, 81, 47–57. [Google Scholar]
- Odžak, R.; Čalić, M.; Hrenar, T.; Primožič, I.; Kovarik, Z. Evaluation of newly synthesized monoquaternary pyridinium oximes potency to reactivate tabun-phosphorylated acetylcholinesterase. Toxicology 2007, 233, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Čalić, M.; Bosak, A.; Kuča, K.; Kovarik, Z. Interactions of butane, but-2-ene or xylene-like linked bispyridinium para-aldoximes with native and tabun-inhibited human cholinesterases. Chem. Biol. Interact. 2008, 175, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Simeon-Rudolf, V.; Šinko, G.; Štuglin, A.; Reiner, E. Inhibition of human blood acetylcholinesterase and butyrylcholinesterase by ethopropazine. Croat. Chem. Acta 2001, 74, 173–182. [Google Scholar]
- Kovarik, Z.; Radić, Z.; Berman, H.A.; Simeon-Rudolf, V.; Reiner, E.; Taylor, P. Acetylcholinesterase active centre and gorge conformations analyzed by combinatorial mutations and enantiomeric phosphonates. Biochem. J. 2003, 373, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Katalinić, M.; Rusak, G.; Domaćinović Barović, J.; Šinko, G.; Jelić, D.; Antolović, R.; Kovarik, Z. Structural aspects of flavonoids as inhibitors of human butyrylcholinesterase. Eur. J. Med. Chem. 2010, 45, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Bosak, A.; Gazić Smilović, I.; Vinković, V.; Šinko, G.; Kovarik, Z. Metaproterenol, isoproterenol and their bisdimethylcarbamate derivatives as human cholinesterase inhibitors. J. Med. Chem. 2012, 55, 6716–6723. [Google Scholar] [CrossRef] [PubMed]
- Greig, N.H.; Utsuki, T.; Yu, Q.-S.; Zhu, X.; Holloway, H.W.; Perry, T.A.; Lee, B.; Ingram, D.K.; Lahiri, D.H. A new therapeutic target in Alzheimer’s disease treatment: Attention to butyrylcholinesterase. Curr. Med. Res. Opin. 2001, 17, 2159–2165. [Google Scholar] [CrossRef]
- Organisation for Economic Co-operation and Development (OECD). Guidance Document on Using Cytotoxicity Test to Estimate Starting Doses for Acute Oral Systemic Toxicity; ENV/JM/MONO(2010)20; OECD: Paris, France, 2010. [Google Scholar]
- Pettit, G.R.; Gupta, S.K. Structural biochemistry. Part 8. 9-amino-(9-deoxy)-Cinchona alkaloids. J. Chem. Soc. Perk. T. 1 1968, 10, 1208–1213. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. Gaussian 09, Revision A.02; Gaussian, Inc.: Wallingford, CT, USA, 2009. [Google Scholar]
- Šinko, G.; Čalić, M.; Bosak, A.; Kovarik, Z. Limitation of the Ellman method: Cholinesterase activity measurement in the presence of oximes. Anal. Biochem. 2007, 370, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. New and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Kovarik, Z.; Ciban, N.; Radić, Z.; Simeon-Rudolf, V.; Taylor, P. Active site mutant acetylcholinesterase interactions with 2-PAM, HI-6, and DDVP. Biochem. Biophys. Res. Commun. 2006, 342, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OP | Oxime | kobs (min−1) | Reactmax (%) | t (h) |
|---|---|---|---|---|
| VX | C1 | - | 20 | 24 |
| C2 | 0.0064 | 60 | 24 | |
| C3 | - | 15 | 22 | |
| HI-6 | 0.0356 | 85 | 2.5 | |
| Sarin | C1 | - | 10 | 24 |
| C2 | 0.0024 | 70 | 24 | |
| C3 | 0.0010 | 65 | 22 | |
| HI-6 | 0.0113 | 98 | 24 | |
| Cyclosarin | C1 | 0.0008 | 45 | 24 |
| C2 | 0.0040 | 45 | 24 | |
| C3 | - | 35 | 22 | |
| HI-6 | 0.0260 | 70 | 24 | |
| Paraoxon | C1 | - | 10 | 24 |
| C2 | - | 30 | 24 | |
| C3 | - | 15 | 22 | |
| Obidoxime | 0.0020 | 60 | 24 | |
| Tabun | C1 | - | 10 | 24 |
| C2 | 0.0015 | 60 | 24 | |
| C3 | - | 15 | 22 | |
| K117 | 0.0095 | 67 | 24 |
| Oxime | Concentration (µM) | hAChE | hBChE | hAChE/hBChE |
|---|---|---|---|---|
| Ki (µM) ± S.E. | ||||
| C1 | 100–400 | 432 ± 52 | - | 272 |
| 1–400 | - | 1.59 ± 0.06 | ||
| C2 | 100–400 | 323 ± 77 | - | 29.9 |
| 10–400 | - | 10.8 ± 0.99 | ||
| C3 | 15–100 | 3.72 ± 0.49 | - | 5.8 |
| 0.5–25 | - | 0.64 ± 0.08 | ||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katalinić, M.; Zandona, A.; Ramić, A.; Zorbaz, T.; Primožič, I.; Kovarik, Z. New Cinchona Oximes Evaluated as Reactivators of Acetylcholinesterase and Butyrylcholinesterase Inhibited by Organophosphorus Compounds. Molecules 2017, 22, 1234. https://doi.org/10.3390/molecules22071234
Katalinić M, Zandona A, Ramić A, Zorbaz T, Primožič I, Kovarik Z. New Cinchona Oximes Evaluated as Reactivators of Acetylcholinesterase and Butyrylcholinesterase Inhibited by Organophosphorus Compounds. Molecules. 2017; 22(7):1234. https://doi.org/10.3390/molecules22071234
Chicago/Turabian StyleKatalinić, Maja, Antonio Zandona, Alma Ramić, Tamara Zorbaz, Ines Primožič, and Zrinka Kovarik. 2017. "New Cinchona Oximes Evaluated as Reactivators of Acetylcholinesterase and Butyrylcholinesterase Inhibited by Organophosphorus Compounds" Molecules 22, no. 7: 1234. https://doi.org/10.3390/molecules22071234
APA StyleKatalinić, M., Zandona, A., Ramić, A., Zorbaz, T., Primožič, I., & Kovarik, Z. (2017). New Cinchona Oximes Evaluated as Reactivators of Acetylcholinesterase and Butyrylcholinesterase Inhibited by Organophosphorus Compounds. Molecules, 22(7), 1234. https://doi.org/10.3390/molecules22071234