β-Ocimene, a Key Floral and Foliar Volatile Involved in Multiple Interactions between Plants and Other Organisms



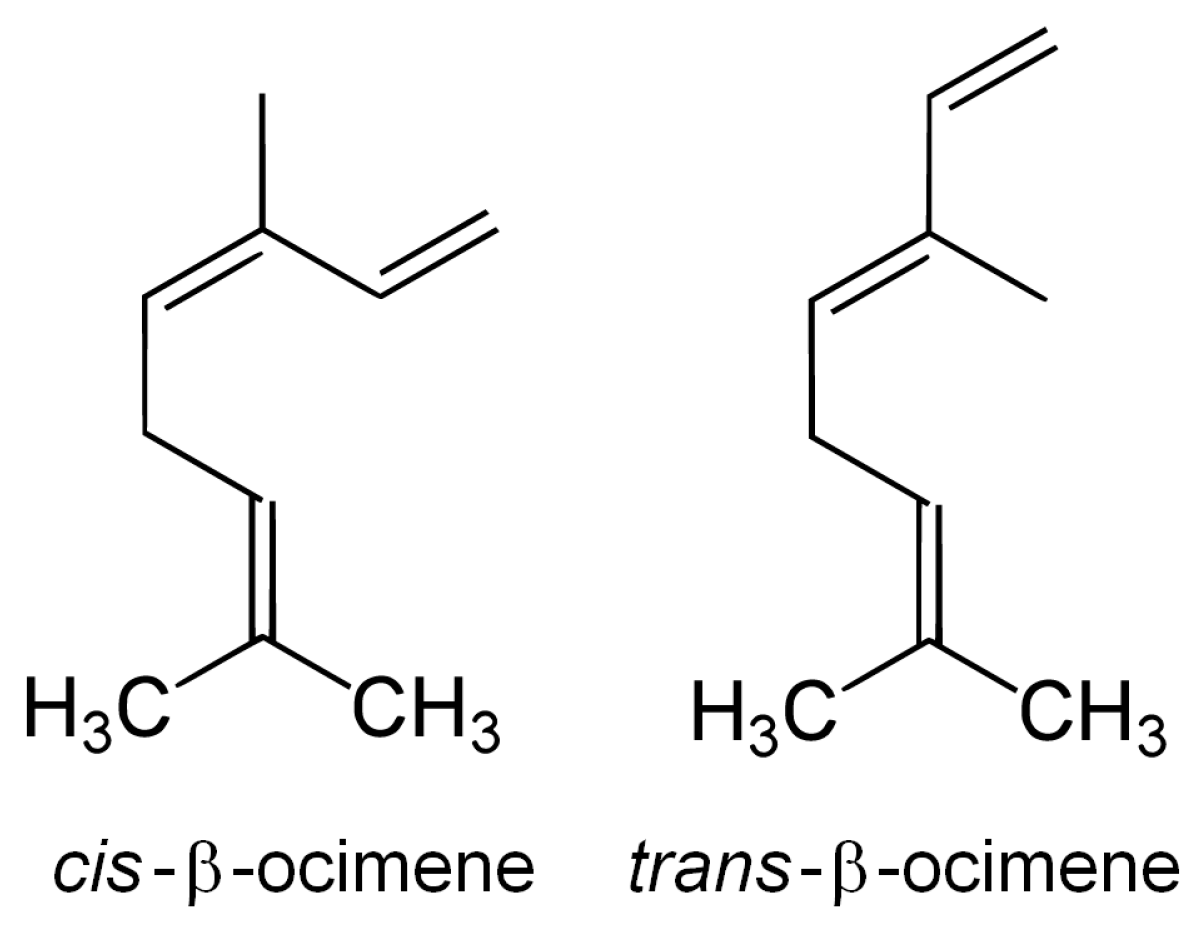
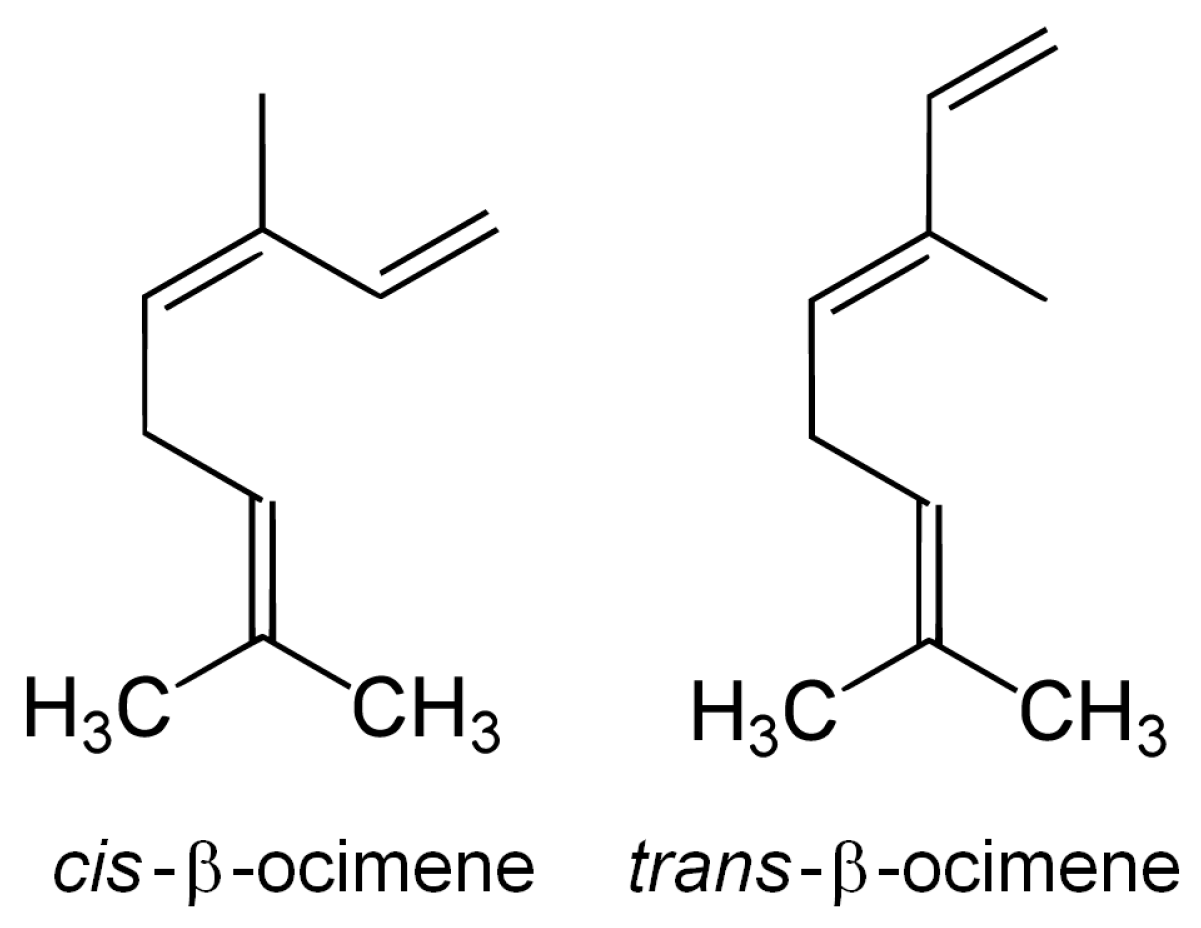{kind=link}
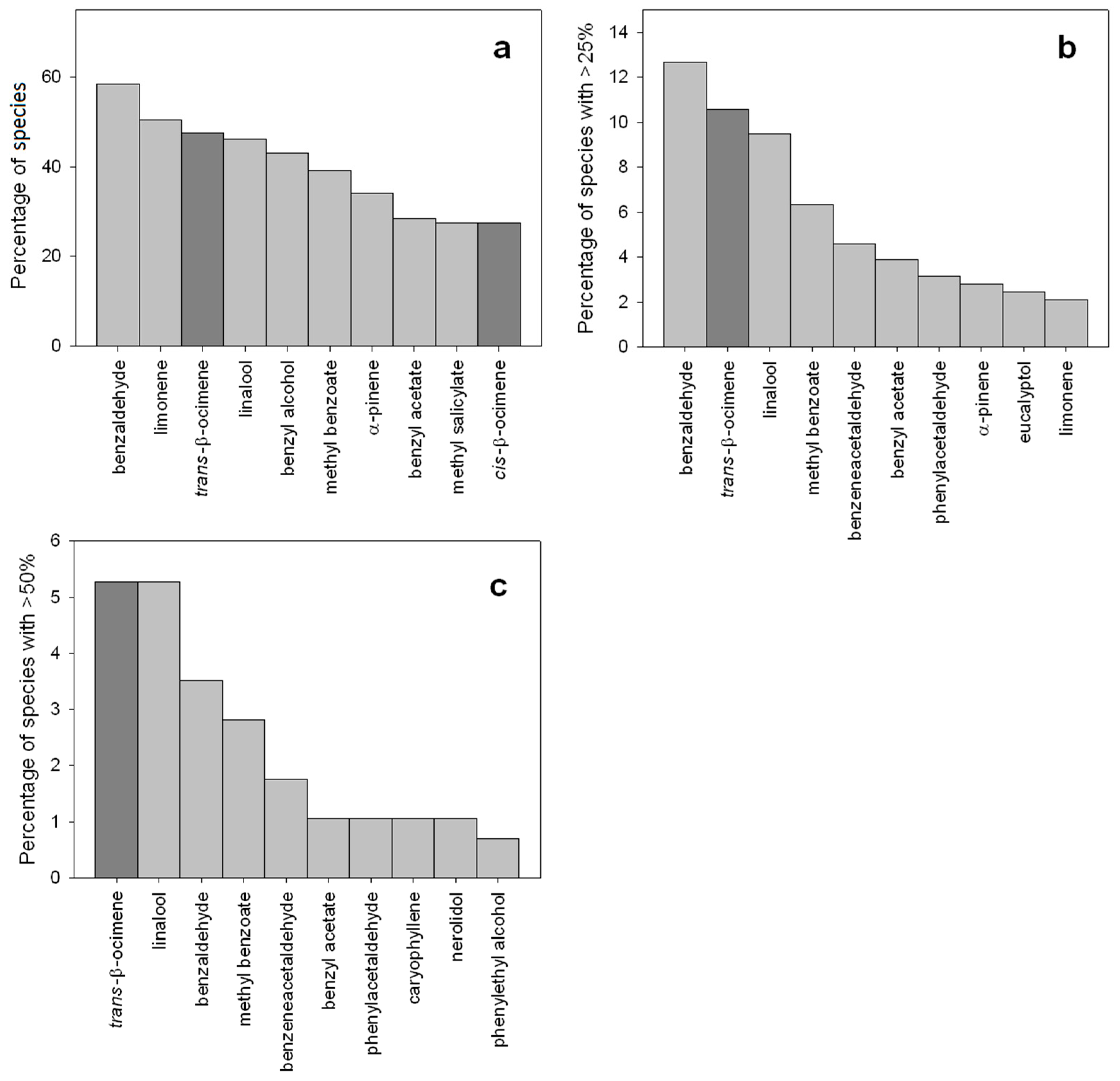{kind=link}
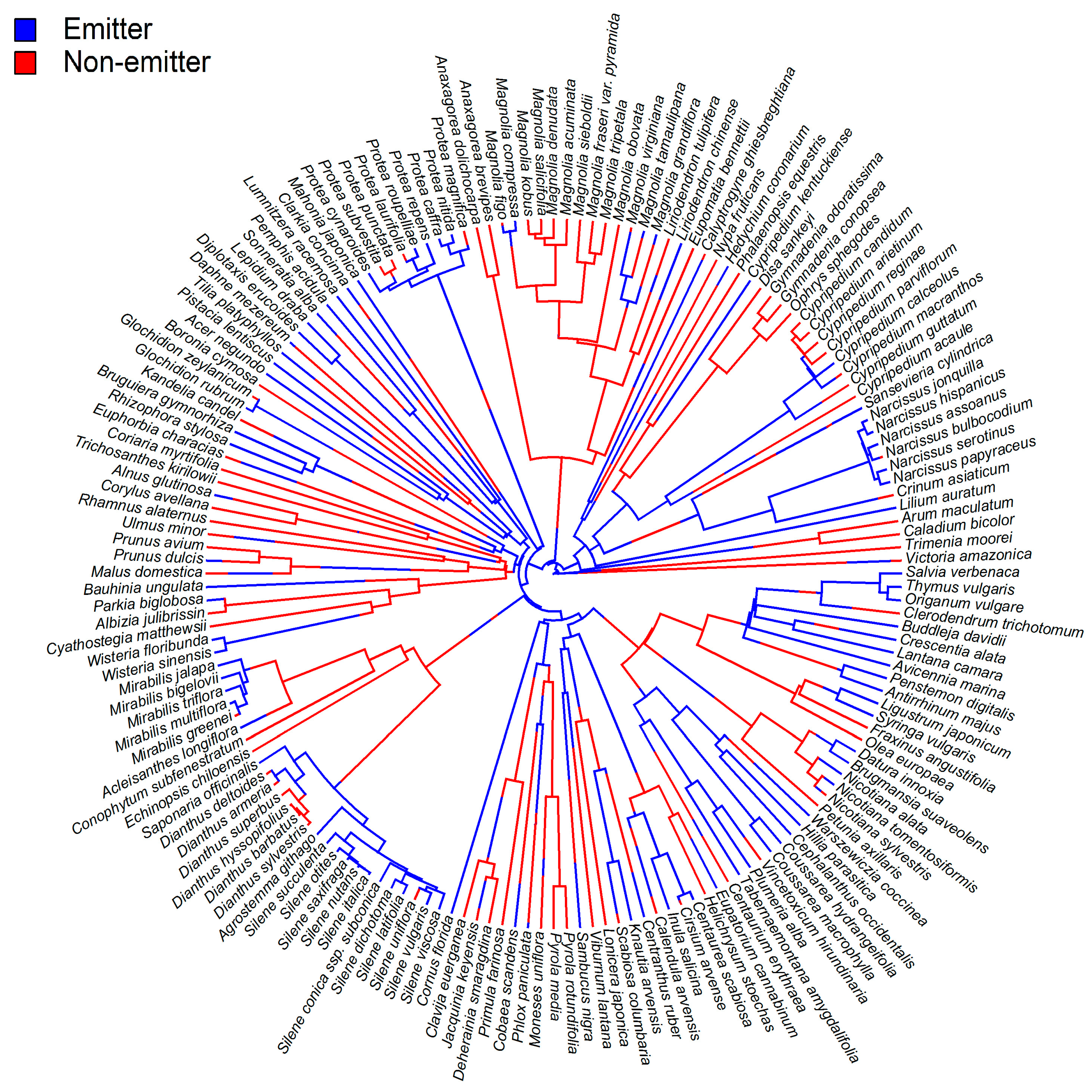{kind=link}
Abstract
:1. Introduction
2. Biosynthesis of β-Ocimene
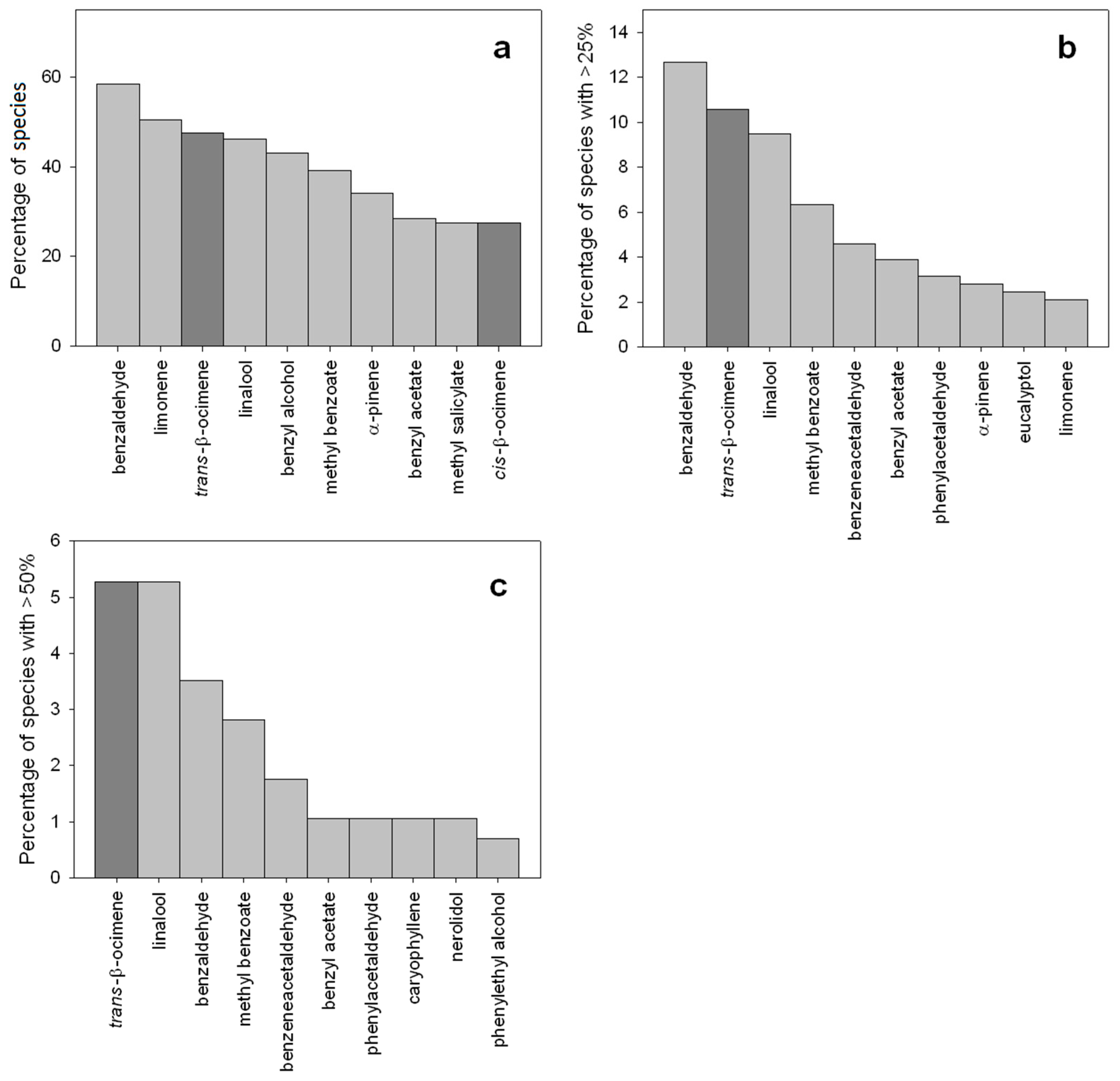
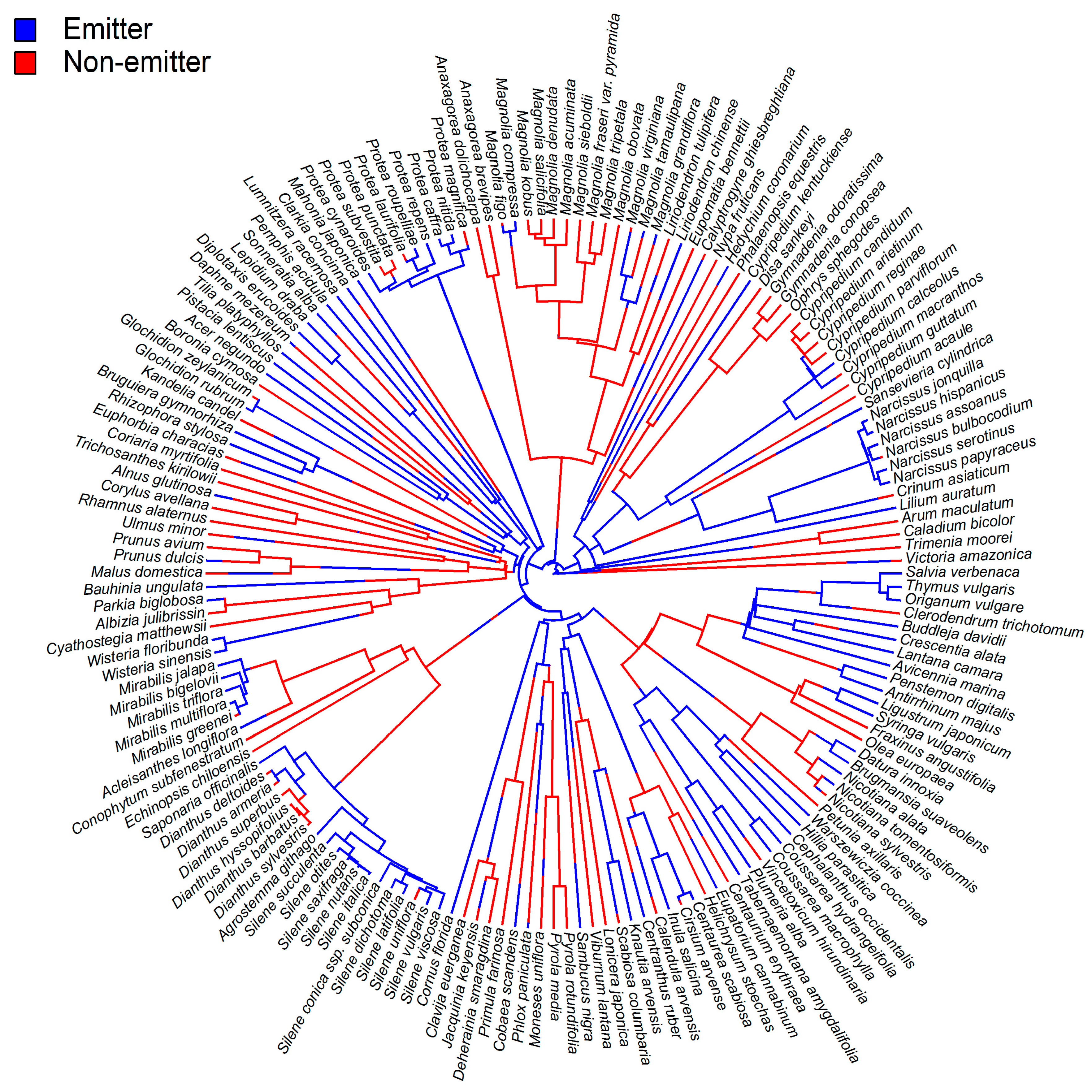
3. Functions of β-Ocimene in Flowers
4. Biological Functions of β-Ocimene Emissions from Non-Floral Tissues
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Knudsen, J.T.; Eriksson, R.; Gershenzon, J.; Ståhl, B. Diversity and Distribution of Floral Scent. Bot. Rev. 2006, 72, 1–120. [Google Scholar] [CrossRef]
- Negre-Zakharov, F.; Long, M.C.; Dudareva, N. Floral scents and fruit aromas inspired by nature. In Plant-Derived Natural Products; Osbourn, A.E., Lanzotti, V., Eds.; Springer: Berlin, Germany, 2009; pp. 405–431. [Google Scholar]
- Grison-pigé, L.; Bessière, J.-M.; Hossaert-McKey, M. Specific attraction of fig-pollinating wasps: Role of volatile compounds released by tropical figs. J. Chem. Ecol. 2002, 28, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.A.; Lutmerding, A.; Dudareva, N.; Smith, B.H. Intensity and the ratios of compounds in the scent of snapdragon flowers affect scent discrimination by honeybees (Apis mellifera). J. Comp. Physiol. A 2005, 191, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, J.T.; Tollsten, L. Floral scent in bat-pollinated plants: A case of convergent evolution. Bot. J. Linn. Soc. 1995, 119, 45–57. [Google Scholar] [CrossRef]
- Von Helversen, O.; Winkler, L.; Bestmann, H.J. Sulphur-containing “perfumes” attract flower-visiting bats. J. Comp. Physiol. A 2000, 186, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.D.; Jürgens, A. Convergent evolution of carrion and faecal scent mimicry in fly-pollinated angiosperm flowers and a stinkhorn fungus. S. Afr. J. Bot. 2010, 76, 796–807. [Google Scholar] [CrossRef]
- Grison, L.; Edwards, A.A.; Hossaert-McMey, M. Interspecies variation in floral fragrances emitted by tropical Ficus species. Phytochemistry 1999, 52, 1293–1299. [Google Scholar] [CrossRef]
- Farré-Armengol, G.; Filella, I.; Llusià, J.; Peñuelas, J. Deciphering the biotic and climatic factors that determine floral scents: A meta-analysis of floral volatile emissions. Perspect. Plant Ecol. Evol. Syst. 2017. submitted. [Google Scholar]
- Pichersky, E.; Noel, J.P.; Dudareva, N. Biosynthesis of plant volatiles: Nature’s diversity and ingenuity. Science 2006, 311, 808–811. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E. Sites of synthesis, biochemistry and functional role of plant volatiles. S. Afr. J. Bot. 2010, 76, 612–631. [Google Scholar] [CrossRef]
- Dudareva, N.; Pichersky, E. Biochemical and Molecular Genetic Aspects of Floral Scents. Plant Physiol. 2000, 122, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Pichersky, E.; Gershenzon, J. Biochemistry of Plant Volatiles. Plant Physiol. 2004, 135, 1893–1902. [Google Scholar] [CrossRef] [PubMed]
- Muhlemann, J.K.; Klempien, A.; Dudareva, N. Floral volatiles: From biosynthesis to function. Plant Cell Environ. 2014, 37, 1936–1949. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, J.; Köllner, T.G.; Gershenzon, J. Monoterpene and sesquiterpene synthases and the origin of terpene skeletal diversity in plants. Phytochemistry 2009, 70, 1621–1637. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Martin, D.; Kish, C.M.; Kolosova, N.; Gorenstein, N.; Fäldt, J.; Miller, B.; Bohlmann, J. (E)-β-Ocimene and Myrcene Synthase Genes of Floral Scent Biosynthesis in Snapdragon: Function and Expression of Three Terpene Synthase Genes of a New Terpene Synthase Subfamily. Plant Cell 2003, 15, 1227–1241. [Google Scholar] [CrossRef] [PubMed]
- Navia-Giné, W.G.; Yuan, J.S.; Mauromoustakos, A.; Murphy, J.B.; Chen, F.; Korth, K.L. Medicago truncatula (E)-β-ocimene synthase is induced by insect herbivory with corresponding increases in emission of volatile ocimene. Plant Physiol. Biochem. 2009, 47, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.-I.; Ozawa, R.; Kugimiya, S.; Takabayashi, J.; Bohlmann, J. Herbivore-Induced Defense Response in a Model Legume. Two-Spotted Spider Mites Induce Emission of (E)-β-Ocimene and Transcript Accumulation of (E)-β-Ocimene Synthase in Lotus japonicus. Plant Physiol. 2004, 135, 1976–1983. [Google Scholar] [CrossRef] [PubMed]
- Faldt, J.; Arimura, G.; Gershenzon, J.; Takabayashi, J.; Bohlmann, J. Functional identification of AtTPS03 as (E)-β-ocimene synthase: A monoterpene synthase catalyzing jasmonate- and wound-induced volatile formation in Arabidopsis thaliana. Planta 2003, 216, 745–751. [Google Scholar] [PubMed]
- Gerlach, G.; Schill, R. Composition of Orchid Scents Attracting Euglossine Bees. Bot. Acta 1991, 104, 379–384. [Google Scholar] [CrossRef]
- Borg-Karlson, A.-K.; Valterová, I.; Nilsson, L.A. Volatile compounds from flowers of six species in the family Apiaceae: Bouquets for different pollinators? Phytochemistry 1994, 35, 111–119. [Google Scholar] [CrossRef]
- Okamoto, T.; Kawakita, A.; Kato, M. Interspecific variation of floral scent composition in Glochidion and its association with host-specific pollinating seed parasite (Epicephala). J. Chem. Ecol. 2007, 33, 1065–1081. [Google Scholar] [CrossRef] [PubMed]
- Dufaÿ, M.; Hossaert-McKey, M.; Anstett, M.C. When leaves act like flowers: How dwarf palms attract their pollinators. Ecol. Lett. 2003, 6, 28–34. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Tollsten, L. Trends in floral scent chemistry in pollination syndromes: Floral scent composition in moth-pollinated taxa. Bot. J. Linn. Soc. 1993, 113, 263–284. [Google Scholar] [CrossRef]
- Andersson, S.; Nilsson, L.A.A.; Groth, I.; Bergstrom, G. Floral scents in butterfly-pollinated plants: Possible convergence in chemical composition. Bot. J. Linn. Soc. 2002, 140, 129–153. [Google Scholar] [CrossRef]
- Filella, I.; Primante, C.; Llusià, J.; Martín González, A.M.; Seco, R.; Farré-Armengol, G.; Rodrigo, A.; Bosch, J.; Peñuelas, J. Floral advertisement scent in a changing plant-pollinators market. Sci. Rep. 2013, 3, 3434. [Google Scholar] [CrossRef] [PubMed]
- Schiestl, F.P.; Schl, P.M. Floral Isolation, Specialized Pollination, and Pollinator Behavior in Orchids. Annu. Rev. Entomol. 2009, 54, 425–446. [Google Scholar] [CrossRef] [PubMed]
- Granero, A.M.; Guerra Sanz, J.M.; Egea Gonzalez, F.J.; Martinez Vidal, J.L.; Dornhaus, A.; Ghani, J.; Serrano, A.R.; Chittka, L. Chemical compounds of the foraging recruitment pheromone in bumblebees. Naturwissenschaften 2005, 92, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Pecetti, L.; Tava, A.; Felicioli, A.; Pinzauti, M.; Piano, E. Effect of three volatile compounds from lucerne flowers on their attractiveness towards pollinators. Bull. Insectol. 2002, 55, 21–27. [Google Scholar]
- Farré-Armengol, G.; Filella, I.; Llusia, J.; Peñuelas, J. Floral volatile organic compounds: Between attraction and deterrence of visitors under global change. Perspect. Plant Ecol. Evol. Syst. 2013, 15, 56–67. [Google Scholar] [CrossRef]
- Bergström, G.; Dobson, H.E.M.; Groth, I. Spatial fragrance patterns within the flowers of Ranunculus acris (Ranunculaceae). Plant Syst. Evol. 1995, 195, 221–242. [Google Scholar] [CrossRef]
- Morinaga, S.I.; Kumano, Y.; Ota, A.; Yamaoka, R.; Sakai, S. Day–night fluctuations in floral scent and their effects on reproductive success in Lilium auratum. Popul. Ecol. 2009, 51, 187–195. [Google Scholar] [CrossRef]
- Dötterl, S.; Jahreiß, K.; Jhumur, U.S.; Jürgens, A. Temporal variation of flower scent in Silene otites (Caryophyllaceae): A species with a mixed pollination system. Bot. J. Linn. Soc. 2012, 169, 447–460. [Google Scholar] [CrossRef]
- Okamoto, T.; Kawakita, A.; Kato, M. Floral adaptations to nocturnal moth pollination in Diplomorpha (Thymelaeaceae). Plant Species Biol. 2008, 23, 192–201. [Google Scholar] [CrossRef]
- Dötterl, S.; Wolfe, L.M.; Jürgens, A. Qualitative and quantitative analyses of flower scent in Silene latifolia. Phytochemistry 2005, 66, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Effmert, U.; Große, J.; Röse, U.S.R.; Ehrig, F.; Kägi, R.; Piechulla, B. Volatile composition, emission pattern, and localization of floral scent Emission in Mirabilis jalapa (Nyctaginaceae). Am. J. Bot. 2005, 92, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Saona, C.; Parra, L.; Quiroz, A.; Isaacs, R. Variation in highbush blueberry floral volatile profiles as a function of pollination status, cultivar, time of day and flower part: Implications for flower visitation by bees. Ann. Bot. 2011, 107, 1377–1390. [Google Scholar] [CrossRef] [PubMed]
- Raguso, R.A.; Levin, R.A.; Foose, S.E.; Holmberg, M.W.; McDade, L.A. Fragrance chemistry, nocturnal rhythms and pollination “syndromes” in Nicotiana. Phytochemistry 2003, 63, 265–284. [Google Scholar] [CrossRef]
- Hoballah, M.E.; Stuurman, J.; Turlings, T.C.J.; Guerin, P.M.; Connétable, S.; Kuhlemeier, C. The composition and timing of flower odour emission by wild Petunia axillaris coincide with the antennal perception and nocturnal activity of the pollinator Manduca sexta. Planta 2005, 222, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Negre, F.; Kish, C.M.; Boatright, J.; Underwood, B.; Shibuya, K.; Wagner, C.; Clark, D.G.; Dudareva, N. Regulation of Methylbenzoate Emission after Pollination in Snapdragon and Petunia Flowers. Plant Cell 2003, 15, 2992–3006. [Google Scholar] [CrossRef] [PubMed]
- Schiestl, F.P.; Ayasse, M. Post-pollination emission of a repellent com-pound in a sexually deceptive orchid: A new mechanism for maximisingreproductive success? Oecologia 2001, 126, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Muhlemann, J.K.; Waelti, M.O.; Widmer, A.; Schiestl, F.P. Postpollination Changes in Floral Odor in Silene latifolia: Adaptive Mechanisms for Seed-Predator Avoidance? J. Chem. Ecol. 2006, 32, 1855–1860. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Endo, T.; Fujii, H.; Hara, M.; Omura, M. Isolation and characterization of (E)-β-ocimene and 1,8 cineole synthases in Citrus unshiu Marc. Plant Sci. 2005, 168, 987–995. [Google Scholar] [CrossRef]
- Kárpáti, Z.; Knaden, M.; Reinecke, A.; Hansson, B.S. Intraspecific Combinations of Flower and Leaf Volatiles Act Together in Attracting Hawkmoth Pollinators. PLoS ONE 2013, 8, e72805. [Google Scholar] [CrossRef] [PubMed]
- Farré-Armengol, G.; Filella, I.; Llusia, J.; Primante, C.; Peñuelas, J. Enhanced emissions of floral volatiles by Diplotaxis erucoides (L.) in response to folivory and florivory by Pieris brassicae (L.). Biochem. Syst. Ecol. 2015, 63, 51–58. [Google Scholar] [CrossRef]
- Theis, N.; Kesler, K.; Adler, L.S. Leaf herbivory increases floral fragrance in male but not female Cucurbita pepo subsp. texana (Cucurbitaceae) flowers. Am. J. Bot. 2009, 96, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Kessler, A.; Halitschke, R.; Poveda, K. Herbivory-mediated pollinator limitation: Negative impacts of induced volatiles on plant–pollinator interactions. Ecology 2011, 92, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Kessler, A.; Halitschke, R. Testing the potential for conflicting selection on floral chemical traits by pollinators and herbivores: Predictions and case study. Funct. Ecol. 2009, 23, 901–912. [Google Scholar] [CrossRef]
- Junker, R.R.; Loewel, C.; Gross, R.; Dötterl, S.; Keller, A.; Blüthgen, N. Composition of epiphytic bacterial communities differs on petals and leaves. Plant Biol. 2011, 13, 918–924. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Sanchez-moreiras, A.M.; Abel, C.; Sohrabi, R.; Lee, S.; Gershenzon, J.; Tholl, D. The major volatile organic compound emitted from Arabidopsis thaliana flowers, the sesquiterpene (E)-β-caryophyllene, is a defense against a bacterial pathogen. New Phytol. 2012, 193, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Raguso, R.A. Why are some floral nectars scented? Ecology 2004, 85, 1486–1494. [Google Scholar] [CrossRef]
- Carter, C.; Thornburg, R.W. Is the nectar redox cycle a floral defense against microbial attack? Trends Plant Sci. 2004, 9, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Adler, L.S. The ecological significance of toxic nectar. Oikos 2000, 91, 409–420. [Google Scholar] [CrossRef]
- Basim, E.; Basim, H.; Özcan, M. Antibacterial activities of Turkish pollen and propolis extracts against plant bacterial pathogens. J. Food Eng. 2006, 77, 992–996. [Google Scholar] [CrossRef]
- Dobson, H.E.M.; Bergström, G. The ecology and evolution of pollen odors. Plant Syst. Evol. 2000, 222, 63–87. [Google Scholar] [CrossRef]
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Kessler, D.; Baldwin, I.T. Making sense of nectar scents: The effects of nectar secondary metabolites on floral visitors of Nicotiana attenuata. Plant J. 2007, 49, 840–854. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.N.; Mishra, A.K.; Tripathi, S. Antibacterial potential of plant volatile oils: A review potential. Proc. Natl. Acad. Sci. India Sect. B Biol. 2011, 81, 23–68. [Google Scholar]
- Utama, I.M.S.; Wills, R.B.H.; Ben-Yehoshua, S.; Kuek, C. In Vitro Efficacy of Plant Volatiles for Inhibiting the Growth of Fruit and Vegetable Decay Microorganisms. J. Agric. Food Chem. 2002, 50, 6371–6377. [Google Scholar] [CrossRef] [PubMed]
- Hansen, U.; Seufert, G. Terpenoid emission from Citrus sinensis (L.) OSBECK under drought stress. Phys. Chem. Earth 1999, 42, 681–687. [Google Scholar] [CrossRef]
- Wang, Y.F.; Owen, S.M.; Li, Q.J.; Peñuelas, J. Monoterpene emissions from rubber trees (Hevea brasiliensis) in a changing landscapeand climate: Chemical speciation andenvironmental control. Glob. Chang. Biol. 2007, 13, 2270–2282. [Google Scholar] [CrossRef]
- Rostelien, T.; Borg-Karlson, A.K.; Mustaparta, H. Selective receptor neurone responses to E-β-ocimene, β-myrcene, E,E-α-farnesene and homo-farnesene in the moth Heliothis virescens, identified by gas chromatography linked to electrophysiology. J. Comp. Physiol. A 2000, 186, 833–847. [Google Scholar] [CrossRef] [PubMed]
- Kariyat, R.R.; Mauck, K.E.; Balogh, C.M.; Stephenson, A.G.; Mescher, M.C.; De Moraes, C.M. Inbreeding in horsenettle (Solanum carolinense) alters night-time volatile emissions that guide oviposition by Manduca sexta moths. Proc. R. Soc. B Biol. Sci. 2013, 280, 20130020. [Google Scholar] [CrossRef] [PubMed]
- Loughrin, J.H.; Manukian, A.; Heath, R.R.; Turlings, T.C.; Tumlinson, J.H. Diurnal cycle of emission of induced volatile terpenoids by herbivore-injured cotton plants. Proc. Natl. Acad. Sci. USA 1994, 91, 11836–11840. [Google Scholar] [CrossRef] [PubMed]
- Miresmailli, S.; Gries, R.; Gries, G.; Zamarc, R.H.; Isman, M.B. Herbivore-induced plant volatiles allow detection of Trichoplusia ni (Lepidoptera: Noctuidae) infestation on greenhouse tomato plants. Pest Manag. Sci. 2010, 66, 916–924. [Google Scholar] [CrossRef] [PubMed]
- Copolovici, L.; Kännastea, A.; Remmel, T.; Niinemets, Ü. Volatile organic compound emissions from Alnus glutinosa under interacting drought and herbivory stresses. Environ. Exp. Bot. 2014, 100, 55–63. [Google Scholar] [CrossRef]
- Miresmailli, S.; Gries, R.; Gries, G.; Zamarc, R.H.; Isman, M.B. Population density and feeding duration of cabbage looper larvae on tomato plants alter the levels of plant volatile emissions. Pest Manag. Sci. 2011, 68, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Pickett, J.A.; Rasmussen, H.B.; Woodcock, C.M.; Matthes, M.; Napier, J.A. Plant stress signalling: Understanding and exploiting plant–plant interactions. Biochem. Soc. Trans. 2003, 31, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Birkett, M.A.; Campbell, C.A.M.; Chamberlain, K.; Guerrieri, E.; Hick, A.J.; Martin, J.L.; Matthes, M.; Napier, J.A.; Pettersson, J.; Pickett, J.A.; et al. New roles for cis-jasmone as an insect semiochemical and in plant defense. Proc. Natl. Acad. Sci. USA 2000, 97, 9329–9334. [Google Scholar] [CrossRef] [PubMed]
- De Moraes, C.M.; Lewis, W.J.; Paré, P.W.; Alborn, H.T.; Tumlinson, J.H. Herbivore-infested plants selectively attract parasitoids. Nature 1998, 393, 570–573. [Google Scholar] [CrossRef]
- De Moraes, C.; Mescher, M.C.; Tumlinson, J.H. Caterpillar-induced nocturnal plant volatiles repel conspecific females. Nature 2001, 410, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.; Ozawa, R.; Shimoda, T.; Nishioka, T.; Boland, W.; Takabayashi, J. Herbivory-induced volatiles elicit defence genes in lima bean leaves. Nature 2000, 406, 512–515. [Google Scholar] [PubMed]
- Arimura, G.; Ozawa, R.; Nishioka, T.; Boland, W.; Koch, T.; Kühnemann, F.; Takabayashi, J. Herbivore-induced volatiles induce the emission of ethylene in neighboring lima bean plants. Plant J. 2002, 29, 87–98. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farré-Armengol, G.; Filella, I.; Llusià, J.; Peñuelas, J. β-Ocimene, a Key Floral and Foliar Volatile Involved in Multiple Interactions between Plants and Other Organisms. Molecules 2017, 22, 1148. https://doi.org/10.3390/molecules22071148
Farré-Armengol G, Filella I, Llusià J, Peñuelas J. β-Ocimene, a Key Floral and Foliar Volatile Involved in Multiple Interactions between Plants and Other Organisms. Molecules. 2017; 22(7):1148. https://doi.org/10.3390/molecules22071148
Chicago/Turabian StyleFarré-Armengol, Gerard, Iolanda Filella, Joan Llusià, and Josep Peñuelas. 2017. "β-Ocimene, a Key Floral and Foliar Volatile Involved in Multiple Interactions between Plants and Other Organisms" Molecules 22, no. 7: 1148. https://doi.org/10.3390/molecules22071148
APA StyleFarré-Armengol, G., Filella, I., Llusià, J., & Peñuelas, J. (2017). β-Ocimene, a Key Floral and Foliar Volatile Involved in Multiple Interactions between Plants and Other Organisms. Molecules, 22(7), 1148. https://doi.org/10.3390/molecules22071148