
Anti-Onchocerca and Anti-Caenorhabditis Activity of a Hydro-Alcoholic Extract from the Fruits of Acacia nilotica and Some Proanthocyanidin Derivatives


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Dereplication of the Active Fractions and Isolation of Catechin-3-O-Gallate as the Major Constituent
2.2. Nematocidal Activity against C. elegans
2.3. Nematocidal Activity against Onchocerca ochengi
2.4. Cytotoxicity against Caco-2 Cells
2.5. Acute Mammalian Toxicity
3. Discussion
4. Materials and Methods
4.1. General Instrumentation for Phytochemical Analysis and Purification
4.2. Chemicals and Plant Material
4.3. Extraction, Bioassay-Guided Fractionation and Isolation of Catechin-3-O-Gallate
4.3.1. Hydro-Alcoholic Extraction from A. nilotica Fruits
4.3.2. Fractionation of Hydro-Alcoholic Extract of A. nilotica Fruits
4.4. Nematocidal Activity of the Pure Compounds
4.4.1. Caenorhabditis elegans Strains and Monoxenic and Axenic Culture Conditions
4.4.2. In Vitro Screening: Lethality Testing Using Caenorhabditis elegans
4.4.3. Onchocerca ochengi Isolation and Culture Conditions
4.4.4. In Vitro Assays: Lethality of Onchocerca ochengi
4.5. In Vitro Cytotoxicity and CC50 Determination
4.6. Acute Oral Toxicity on Rats
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Organisation Mondiale de la Santé (OMS). Agir Pour Réduire L’Impact des Maladies Tropicales Négligées: Premier Rapport de L’OMS Sur les Maladies Tropicales Négligées; OMS: Genève, Switzerlands, 2011; pp. 124–129. [Google Scholar]
- Zoure, H.; Noma, M.; Tekle, A.; Amazigo, U.; Diggle, P.; Giorgi, E.; Remme, J.H. The geographic distribution of onchocerciasis in the 20 participating countries of the African Programme for Onchocerciasis Control: 2. pre-control endemicity levels and estimated number infected. Parasites Vectors 2014, 7, 326. [Google Scholar] [CrossRef] [PubMed]
- Tanya, V.N.; Wandji, S.; Kamgno, J.; Achukwi, D.M.; Enyong, P.A.I. Recent Advances in Onchocerciasis Research and Implication for Control; Cameroon Academy of Sciences: Yaounde, Cameroon, 2013; p. 200. [Google Scholar]
- Ali, M.M.; Mukhtar, M.M.; Baraka, O.Z.; Homeida, M.M.; Kheir, M.M.; Mackenzie, C.D. Immunocompetence may be important in the effectiveness of Mectizan (ivermectin) in the treatment of human onchocerciasis. Acta Trop. 2002, 84, 49–53. [Google Scholar] [CrossRef]
- Awadzi, K.; Boakye, D.A.; Edwards, G.; Opoku, N.O.; Attah, S.K.; Osei-Atweneboana, M.Y.; Lazdins-Helds, J.K.; Ardrey, A.E.; Addy, E.T.; Quartey, B.T.; et al. An investigation of persistent microfilaridermias despite multiple treatments with ivermectin, in two onchocerciasis-endemic foci in Ghana. Ann. Trop. Med. Parasitol. 2004, 98, 231–249. [Google Scholar] [CrossRef] [PubMed]
- Osei-Atweneboana, M.Y.; Eng, J.K.L.; Boakye, D.A.; Gyapong, J.O.; Prichard, R.K. Prevalence and intensity of Onchocerca volvulus infection and efficacy of ivermectin in endemic communities in Ghana: A twophase epidemiological study. Lancet 2007, 369, 2021–2029. [Google Scholar] [CrossRef]
- Langworthy, N.G.; Renz, A.; Mackenstedt, U.; Henkle-Dührsen, K.; de Bronsvoort, M.B.; Tanya, V.N.; Donnelly, M.J.; Trees, A.J. Macrofilaricidal activity of tetracycline against the filarial nematode Onchocerca ochengi: Elimination of Wolbachia precedes worm death and suggests a dependent relationship. Proc. Biol. Sci. 2000, 267, 1063–1069. [Google Scholar] [CrossRef] [PubMed]
- Townson, S.; Tagboto, S.; McGarry, H.F.; Egerton, G.L.; Taylor, M.J. Onchocerca parasites and Wolbachia endosymbionts: Evaluation of a spectrum of antibiotic types for activity against Onchocerca gutturosa in vitro. Filarial J. 2006, 5, 4. [Google Scholar] [CrossRef] [PubMed]
- Büttner, D.W.; Wanji, S.; Bazzocchi, C.; Bain, O.; Fischer, P. Obligatory symbiotic Wolbachia endobacteriae are absent from Loa loa. Filarial J. 2003, 2, 10. [Google Scholar] [CrossRef] [PubMed]
- Trees, A.J. Onchocerca ochengi: Mimic, Model, or Modulator of O. volvulus? Parasitol. Today 1992, 8, 337–339. [Google Scholar] [CrossRef]
- Ndjonka, D.; Rapado, L.N.; Silber, A.M.; Liebau, E.; Wrenger, C. Natural Products as a source for treating neglected parasitic diseases. Int. J. Mol. Sci. 2013, 14, 3395–3439. [Google Scholar] [CrossRef] [PubMed]
- Nyasse, B.; Ngantchou, I.; Nono, J.J.; Schneider, B. Antifilarial activity in vitro of polycarpol and 3-O-acetyl aleuritolic acid from cameroonian medicinal plants against Onchocercagutturosa. Nat. Prod. Res. 2006, 20, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Ndjonka, D.; Abladam, E.D.; Djafsia, B.; Ajonina-Ekoti, I.; Achukwi, M.D.; Liebau, E. Anthelmintic activity of phenolic acids from the axlewood tree Anogeissus leiocarpus on the filarial nematode Onchocerca ochengi and drug-resistant strains of the free-living nematode Caenorhabditis elegans. J Helminthol. 2014, 88, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Cho-Ngwa, F.; Abongwa, M.; Ngemenya, M.N.; Nyongbela, K.D. Selective activity of extracts of Margaritaria discoidea and Homalium africanum on Onchocerca ochengi. BMC Complement. Altern. Med. 2010, 10, 62. [Google Scholar] [CrossRef] [PubMed]
- Katiki, L.M.; Ferreira, J.F.; Gonzalez, J.M.; Zajac, A.M.; Lindsay, D.S.; Chagas, A.C.; Amarante, A.F. Anthelmintic effect of plant extracts containing condensed and hydrolyzable tannins on Caenorhabditis elegans, and their antioxidant capacity. Vet. Parasitol. 2012, 18, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Ndjonka, D.; Agyare, C.; Luersen, K.; Djafsia, B.; Achukwi, D.; Nukenine, E.N.; Hensel, A.; Liebau, E. In Vitro activity of Cameroonian and Ghanaian medicinal plants on parasitic (Onchocerca ochengi) and free-living (Caenorhabditis elegans) nematodes. J. Helminthol. 2011, 85, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Ndjonka, D.; Ajonina-Ekoti, I.; Djafsia, B.; Luersen, K.; Abladam, E.; Liebau, E. Anogeissus leiocarpus extract on the parasite nematode Onchocerca ochengi and on drug resistant mutantstrains of the free-living nematode Caenorhabditis elegans. Vet. Parasitol. 2012, 190, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, H.; Reider, K.; Franke, K.; Wessjohann, L.A.; Keiser, J.; Dagne, E.; Arnold, N. Characterization of constituents and anthelmintic properties of Hagenia abyssinica. Sci. Pharm. 2012, 80, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Waterman, C.; Smith, R.A.; Pontiggia, L.; DerMarderosian, A. Anthelmintic screening of Sub-Saharan African plants used in traditional medicine. J. Ethnopharmacol. 2010, 127, 755–759. [Google Scholar] [CrossRef] [PubMed]
- Metuge, J.A.; Nyongbela, K.D.; Mbah, J.A.; Samje, M.; Fotso, G.; Babiaka, S.B.; Cho-Ngwa, F. Anti-Onchocerca activity and phytochemical analysis of an essential oil from Cyperus articulatus L. BMC Complement. Altern. Med. 2014, 14, 223. [Google Scholar] [CrossRef] [PubMed]
- Metuge, J.A.; Babiaka, S.B.; Mbah, J.A.; Ntie-Kang, F.; Ayimele, G.A.; Cho-Ngwa, F. Anti-onchocerca Metabolites from Cyperus articulatus: Isolation, In Vitro Activity and In Silico. ‘Drug-Likeness’. Nat. Prod. Bioprospect. 2014, 4, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Samje, M.; Metuge, J.; Mbah, J.; Nguesson, B.; Cho-ngwa, F. In Vitro anti-Onchocerca ochengi activities of extracts and chromatographic fractions of Craterispermum laurinum and Morinda lucida. BMC Complement. Altern. Med. 2014, 14, 325. [Google Scholar] [CrossRef] [PubMed]
- Eyog, M.O.; Ndoye, O.; Kengue, J.; Awono, A. Les Fruitiers Forestiers Comestibles du Cameroun; International Plant Genetic Resources Institute (IPGRI): Cotonou, Benin, 2006; p. 220. [Google Scholar]
- Solomon-Wisdom, G.O.; Shittu, G.A. In Vitro antimicrobial and phytochemical activities of Acacia nilotica leaf extract. J. Med. Plants Res. 2010, 4, 1232–1234. [Google Scholar]
- Ali, A.; Akhtar, N.; Khan, B.A.; Khan, M.S.; Rasul, A.; Khalid, N.; Waseem, K.; Mahmood, T.; Ali, L. Acacia nilotica: A plant of multipurpose medicinal uses. J. Med. Plants Res. 2012, 6, 1492–1496. [Google Scholar]
- Maldini, M.; Montoro, P.; Hamed, A.I.; Mahalel, U.; Oleszek, W.; Stochmal, A.; Piacente, S. Strong antioxidant phenolics from Acacia nilotica: Profiling by ESI-MS and qualitative-quantitative determination by LC-ESI-MS. J. Pharm. Biomed. Anal. 2011, 56, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Malan, E. Derivatives of (+)-catechin-5-gallate from the bark of Acacia nilotica. Phytochemistry 1991, 30, 2737–2739. [Google Scholar] [CrossRef]
- Singh, R.; Singh, B.; Singh, S.; Kumar, N.; Kumar, S.; Arora, S. Anti-free radical activities of kaempferol isolated from Acacia nilotica (L.) Willd. Ex. Del. Toxicol. In Vitro 2008, 22, 1965–1970. [Google Scholar] [CrossRef] [PubMed]
- Badar, N.; Iqbal, Z.; Khan, M.N.; Akhtar, M.S. In Vitro and In Vivo anthelmintic activity of Acacia nilotica (L.) willd. Ex Delile bark and leaves. Pak. Vet. J. 2011, 31, 185–191. [Google Scholar]
- Ferreira, L.E.; Castro, P.M.; Chagas, A.C.; França, S.C.; Beleboni, R.O. In vitro anthelmintic activity of aqueous leaf extract of Annona muricata L. (Annonaceae) against Haemonchus contortus from sheep. Exp. Parasitol. 2013, 134, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.L.; Cai, Y.; Davies, A.P.; Lewis, J.R. 1H and 13C NMR Assignments of some green tea polyphenols. Magn. Reson. Chem. 1996, 34, 887–890. [Google Scholar] [CrossRef]
- Ramanandraibe, V.; Grellier, P.; Martin, M.T.; Deville, A.; Joyeau, R.; Ramanitrahasimbola, D.; Mouray, E.; Rasoanaive, P.; Mambu, L. Antiplasmodial phenolic compounds from Piptadenia pervillei. Planta Med. 2008, 74, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Kahiya, C.; Mukaratirwa, S.; Thamsborg, S.M. Effects of Acacia nilotica and Acacia karoo diets on Haemonchus contortus infection in goats. Vet. Parasitol. 2003, 115, 265–274. [Google Scholar] [CrossRef]
- Ahmadu, A.; Abdulkarim, A.; Grougnet, R.; Myrianthopoulos, V.; Tillequin, F.; Magiatis, P.; Skaltsounis, A.L. Two new peltogynoids from Acacia nilotica Delile with kinase inhibitory activity. Planta Med. 2010, 76, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Geary, T.G.; Sangster, N.C.; Thompson, D.P. Frontiers in anthelmintic pharmacology. Vet. Parasitol. 1999, 84, 275–295. [Google Scholar] [CrossRef]
- Katiki, L.M.; Ferreira, J.F.; Zajac, A.M.; Masler, C.; Lindsay, D.S.; Carolina, A.; Chagas, A.C.; Amarante, A.F. Caenorhabditis elegans as a model to screen plant extracts and compounds as natural anthelmintics for veterinary use. Vet. Parasitol. 2011, 182, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Koné, W.M.; Vargas, M.; Keiser, J. Anthelmintic activity of medicinal plants used in Côte d’Ivoire for treating parasitic diseases. Parasitol. Res. 2012, 110, 2351–2362. [Google Scholar] [CrossRef] [PubMed]
- Manolaraki, F.; Sotiraki, S.; Stefanakis, A.; Skampardonis, V.; Volanis, M.; Hoste, H. Anthelmintic activity of some Mediterranean browse plants against parasitic nematodes. Parasitology 2010, 137, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jie, G.; Zhang, J.; Zhao, B. Significant longevity-extending effects of EGCG on Caenorhabditis elegans under stress. Free Radic. Biol. Med. 2009, 46, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Mukai, D.; Matsuda, A.E.N.; Yoshioka, A.E.Y. Potential anthelmintics: Polyphenols from the tea plant Camellia sinensis L. are lethally toxic to Caenorhabditis elegans. J. Nat. Med. 2008, 62, 155–159. [Google Scholar] [PubMed]
- Yamasaki, T.; Sato, M.; Mori, T.; Mohamed, A.S.A.; Fujii, K.; Tsukioka, J. Toxicity of tannins towards the free-living nematode Caenorhabditis elegans and the brine shrimp Artemia salina. J. Nat. Toxins 2002, 11, 165–171. [Google Scholar] [PubMed]
- Martin, R.J. Modes of Action of Anthelmintic Drugs. Vet. J. 1997, 31, 11–34. [Google Scholar] [CrossRef]
- Culetto, E.; Baylis, H.A.; Richmond, J.E.; Jones, A.K.; Fleming, J.T.; Squire, M.D.; Lewis, J.A.; Sattelle, D.B. The Caenorhabditis elegans unc-63 gene encodes a levamisole sensitive nicotinic acetylcholine. J. Biol. Chem. 2004, 279, 42476–42483. [Google Scholar] [CrossRef] [PubMed]
- Köhler, P. The biochemical basis of anthelmintic action and resistance. Int. J. Parasitol. 2001, 31, 336–345. [Google Scholar] [CrossRef]
- Roos, M.H.; Boersema, J.H.; Borgsteede, F.H.; Cornelissen, J.; Taylor, M.; Ruitenberg, E.J. Molecular analysis of selection for benzimidazole resistance in the sheep parasite Haemonchus contortus. Mol. Biochem. Parasitol. 1990, 43, 77–88. [Google Scholar] [CrossRef]
- Lubega, G.W.; Klein, R.D.; Geary, T.G.; Prichard, R.K. Haemonchus contortus: The role of two β-tubulin gene subfamilies in the resistance to benzimidazole anthelmintics. Biochem. Pharmacol. 1994, 47, 1705–1715. [Google Scholar] [CrossRef]
- Driscoll, M.; Dean, E.; Reilly, E.; Bergholz, E.; Chalfie, M. Genetic and Molecular Analysis of a Caenorhabditis elegans β-tubulin that conveys benzimidazole sensitivity. J. Cell Biol. 1989, 109, 2993–3003. [Google Scholar] [CrossRef] [PubMed]
- Yates, D.M.; Portillo, V.; Wolstenholme, A.J. The avermectin receptors of Haemonchus contortus and Caenorhabditis elegans. Int. J. Parasitol. 2003, 33, 1183–1193. [Google Scholar] [CrossRef]
- Laughton, D.L.; Lunt, G.G.; Wolstenholme, A.J. Reporter gene constructs suggest that the Caenorhabditis elegans avermectin receptor beta-subunit is expressed solely in the pharynx. J. Exp. Biol. 1997, 200, 1509–1514. [Google Scholar] [PubMed]
- Dent, J.A.; Smith, M.M.; Vassilatis, D.K.; Avery, L. The genetics of ivermectin resistance in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2000, 97, 2674–2679. [Google Scholar] [CrossRef] [PubMed]
- Chun-Yang, Y. Emerging usage of plant-based coagulants for water and wastewater treatment. Process Biochem. 2010, 45, 1437–1444. [Google Scholar]
- Jeon, J.R.; Kim, E.J.; Kim, Y.M.; Murugesan, K.; Kim, J.H.; Chang, Y.S. Use of grape seed and its natural polyphenol extracts as a natural organic coagulant for removal of cationic dyes. Chemosphere 2009, 77, 1090–1098. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, Z.; Sarwar, M.; Jabbar, A.; Ahmed, S.; Nisa, M.; Sajid, M.S.; Khan, M.N.; Mufti, K.A.; Yaseen, M. Direct and indirect anthelmintic effects of condensed tannins in sheep. Vet. Parasitol. 2007, 144, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.P.; Geary, T.G. The structure and function of helminth surfaces. In Biochemistry and Molecular Biology of Parasites, 1st ed.; Marr, J.J., Ed.; Academic Press: New York, NY, USA, 1995; Volume 1, pp. 203–232. [Google Scholar]
- Organization for Economic Co-operation and Development (OECD). Guideline No. 423. Acute Oral Toxicity in Animals; OECD/OCDE No. 423; OECD: Paris, France, 2001. [Google Scholar]
- Mohan, S.; Thiagarajan, K.; Chandrasekaran, R.; Arul, J. In Vitro protection of biological macromolecules against oxidative stress and in vivo toxicity evaluation of Acacia nilotica (L.) and ethyl gallate in rats. BMC Complement. Altern. Med. 2014, 14, 257. [Google Scholar] [CrossRef] [PubMed]
- Guta, M.; Urga, K.; Assefa, A.; Lemma, H.; Addis, G.; Gemeda, N.; Yirsaw, K.; Mudi, K.; Melaku, D. Antibacterial and acute toxicity study of Acacia nilotica. Ethiop. J. Biol. Sci. 2007, 6, 43–49. [Google Scholar]
- Zumdick, S.; Deters, A.; Hensel, A. In vitro transport of Oligomeric Procyanidins (DP 2 to 4) across monolayers caco-2 cells. Fitoterapia 2012, 83, 1210–1217. [Google Scholar] [CrossRef] [PubMed]
- Huggins, D.J.; Sherman, W.; Tidor, B. Rational Approaches to Improving Selectivity in Drug Design. J. Med. Chem. 2012, 55, 1424–1444. [Google Scholar] [CrossRef] [PubMed]
- Qa’dan, F.; Petereit, F.; Nahrstedt, A. Prodelphinidin trimers and characterization of a proanthocyanidin oligomer from Cistus albidus. Pharmazie 2003, 58, 416–419. [Google Scholar] [PubMed]
- Danne, A.; Petereit, F.; Nahrstedt, A. Flavan-3-ols, prodelphinidins and further polyphenols from Cistus salvifolius. Phytochemistry 1994, 37, 533–538. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [PubMed]
- Sulston, J.; Hodgkin, J. Methods. In The Nematode Caenorhabditis elegans, 13th ed.; Wood, W.B., Ed.; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1988; Volume 17, pp. 587–606. [Google Scholar]
- Chitwood, D.J.; Feldlaufer, M.F. Ecdysteroids in Axenically Propagated Caenorhabditis elegans and medium culture medium. J. Nematol. 1990, 22, 598–607. [Google Scholar] [PubMed]
- Dengg, M.; van Meel, J.C. Caenorhabditis elegans as model system for rapid toxicity assessment of pharmaceutical compounds. J. Pharmacol. Toxicol. Methods 2004, 50, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Zhao, Y.; Wu, Q.; Sun, L.; Ruan, Q.; Chen, Y.; Wang, M.; Duan, J.; Wang, D. Full toxicity assessment of Genkwa Flos and the underlying mechanism in nematode Caenorhabditis elegans. PLoS ONE 2014, 9, e91825. [Google Scholar] [CrossRef] [PubMed]
- Kläger, S.; Whitworth, J.A.G.; Post, R.J.; Chavasse, D.C.; Dowham, M.D. How long do the effects of ivermectin on adult Onchocerca volvulus persist? Trop. Med. Pasatol. 1993, 44, 305–310. [Google Scholar]
- Ajaiyeoba, E.O.; Abiodun, O.O.; Falade, M.O.; Ogbole, N.O.; Ashidi, J.S.; Happi, C.T.; Akinboye, D.O. In Vitro cytotoxicity studies of 20 plants used in Nigerian antimalarial ethnomedicine. Phytomedicine 2006, 13, 295–298. [Google Scholar] [CrossRef] [PubMed]
- European Economic Community. Council Directive 86/609/EEC of 24 November 1986 on the Approximation of Laws, Regulations and Administrative Provisions of the Member States Regarding the Protection of Animals Used for Experimental and Other Scientific Purposes; European Economic Community: Brussels, Belgium, 1986. [Google Scholar]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. PLoS Biol. 2010, 8, e100041. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |
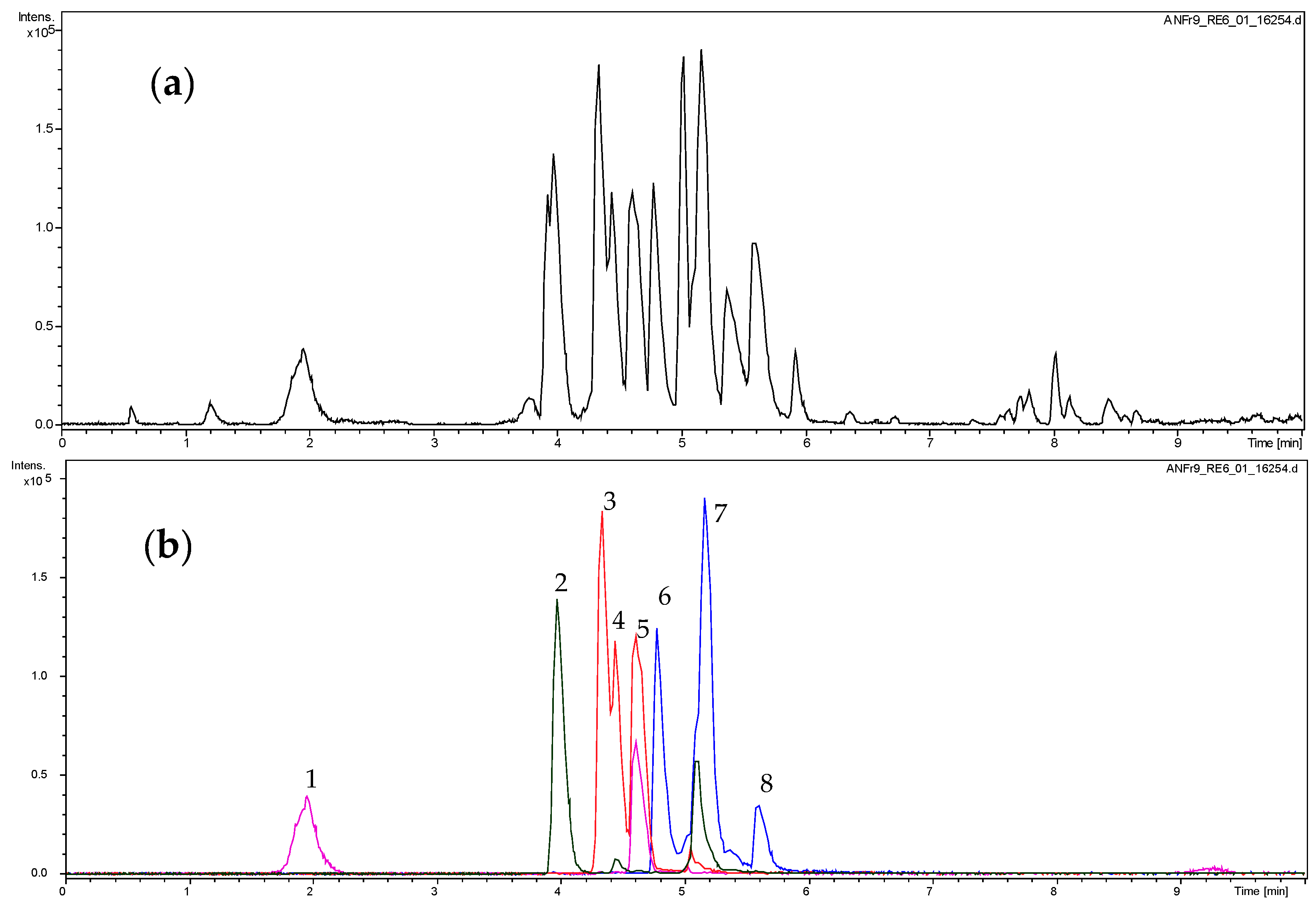
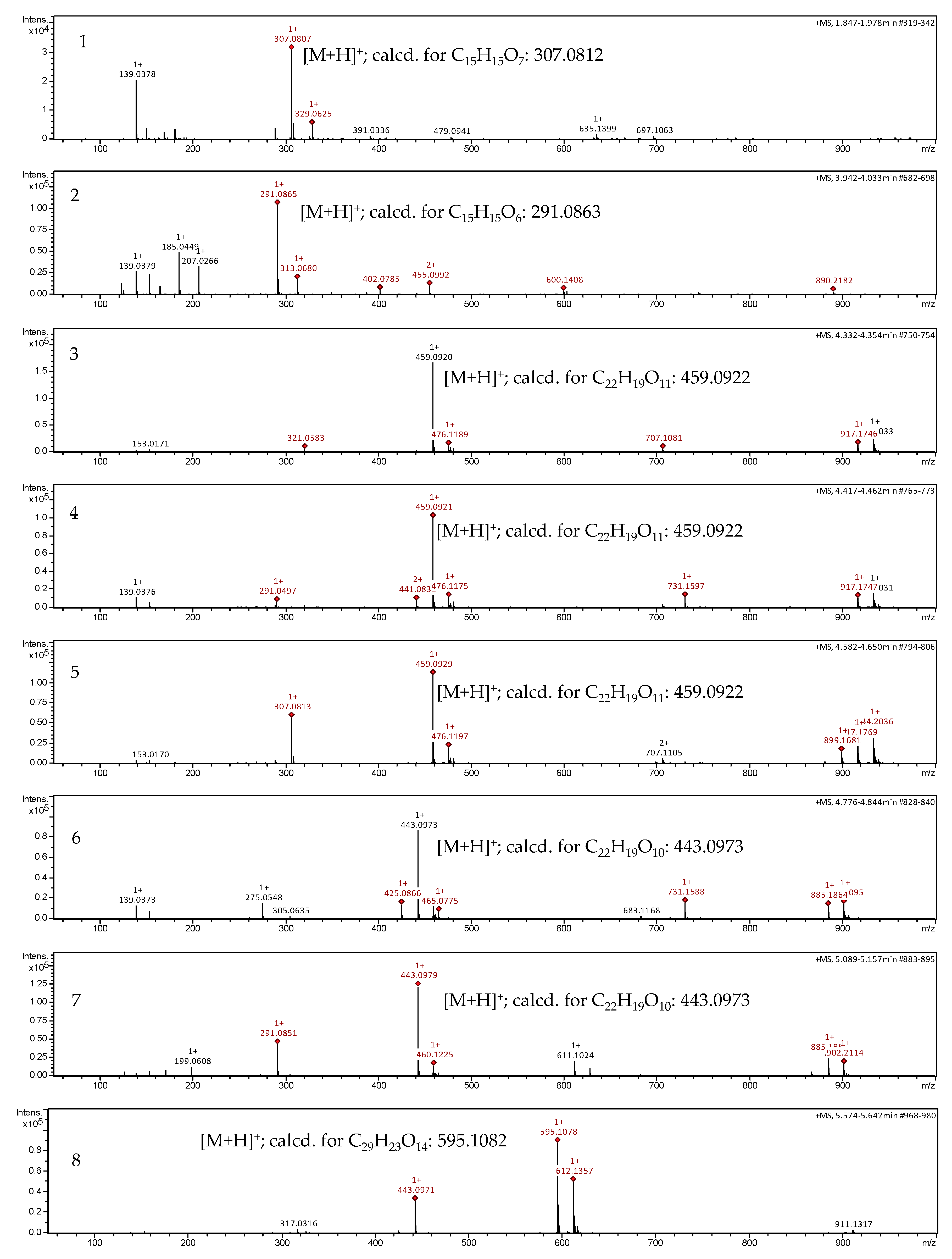
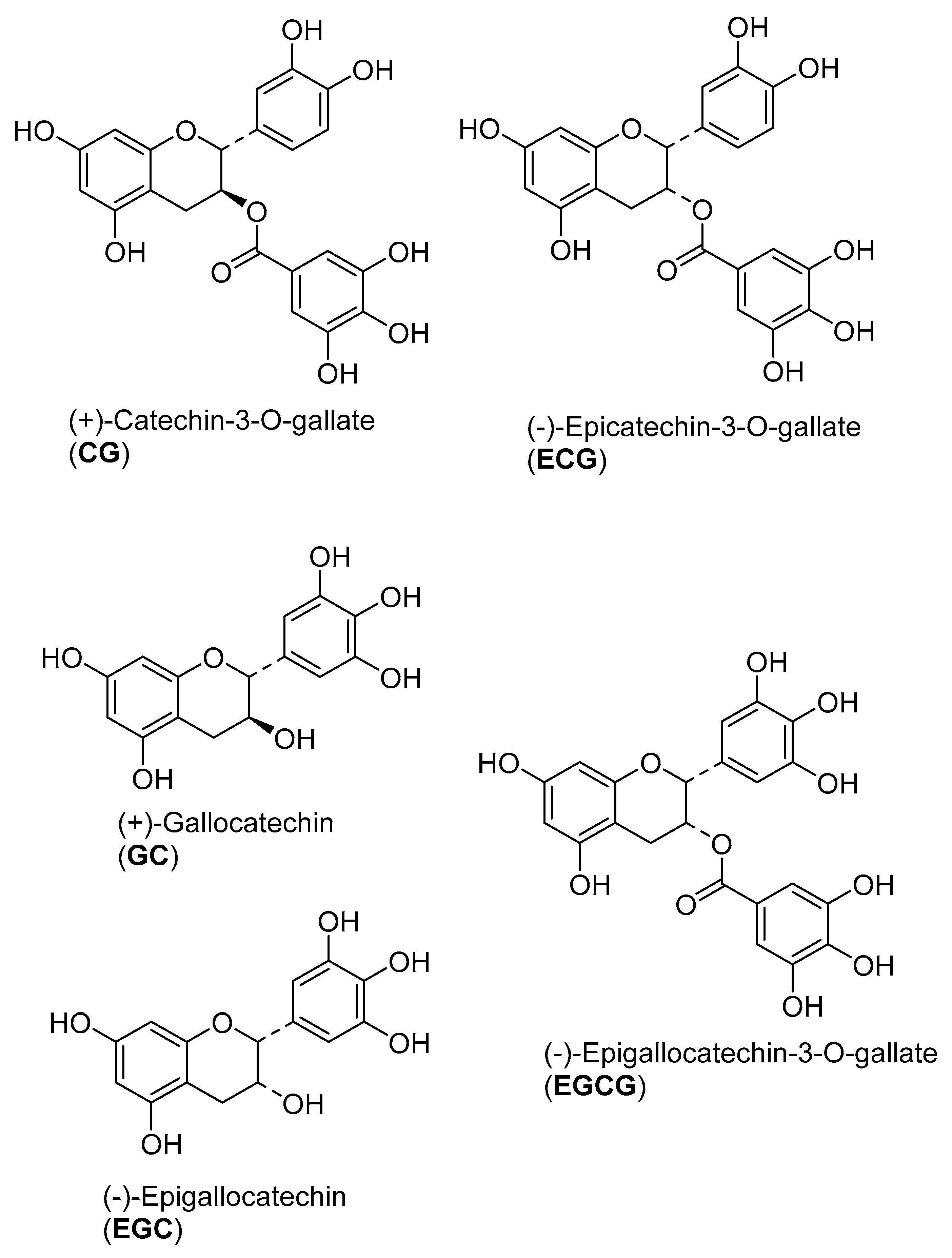

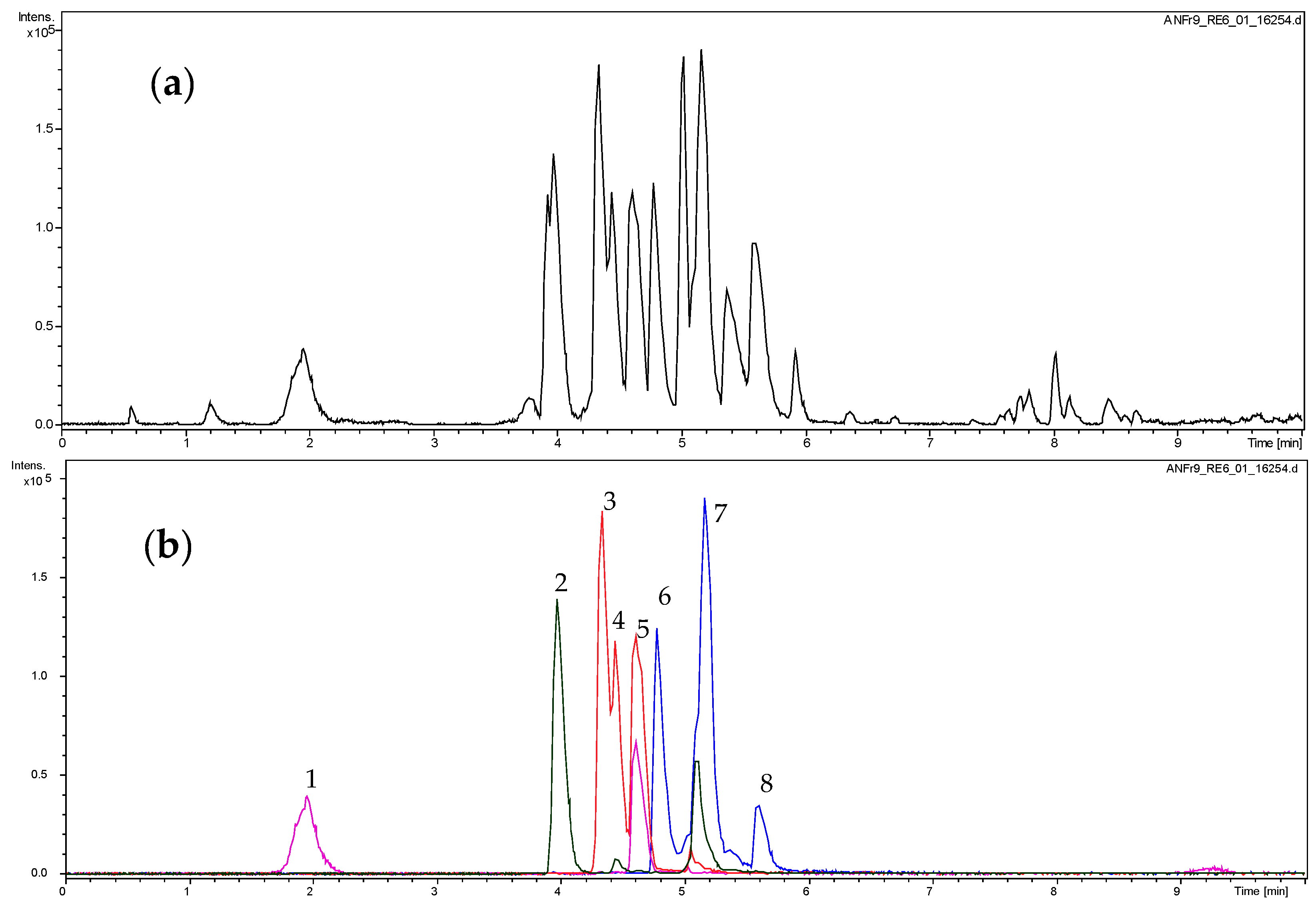{kind=link}
{kind=link}
{kind=link}
| LC50/48 h in μg/mL (µM) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Worms | CE | CG | ECG | GC | EGC | EGCG | Levamisole | Ivermectin | Albendazole |
| Means LC50 ± SD | |||||||||
| O. ochengi males | 11.5 ± 0.1 | 7.6 ± 0.2 (17.2) | 2.1 ± 0.3 (4.7) | 4.2 ± 0.1 (13.6) | 2.1 ± 0.4 (6.8) | 1.2 ± 0.5 (2.6) | 1.0 ± 1.0 (5.1) | 1.2 ± 0.5 (1.3) | 4.2 ± 0.3 (15.7) |
| O. ochengi females | 11.0 ± 0.2 | 4.5 ± 0.3 (10.2) | 2.1 ± 0.6 (4.7) | 5.5 ± 0.3 (18.0) | 3.3 ± 0.6 (10.6) | 1.0 ± 0.5 (2.3) | 2.1 ± 0.1 (10.2) | 1.5 ± 0.3 (1.7) | 1.0 ± 0.2 (3.9) |
| O. ochengi microfilariae | 10.8 ± 0.3 | 4.2 ± 0.1 (9.4) | 1.0 ± 0.4 (2.2) | 3.2 ± 0.3 (10.4) | 3.3 ± 0.5 (10.6) | 1.3 ± 0.2 (2.8) | 1.0 ± 0.3 (5.1) | 1.5 ± 0.0 (1.7) | 2.1 ± 0.5 (7.8) |
| C. elegans WT | 350 ± 1.1 | 90.6 ± 0.07 (204.8) | 33.8 ± 2.7 (2.4) | 74.3 ± 0.3 (242.4) | 62.7 ± 0.2 (204.7) | 49.8 ± 0.8 (108.6) | 1.4 ± 0.2 (7.2) | 1.2 ± 0.1 (1.3) | 7.1 ± 0,2 (26.4) |
| C. elegans ZZ16 | 987.9 ± 0.3 | 95.3 ± 0.1 (215.4) | 39.5 ± 0.9 (76.4) | 65.6 ± 0.2 (214.2) | 92.0 ± 0.3 (301.4) | >100 | >100 | nd | nd |
| C. elegans CB211 | 934.3 ± 1.0 | 96.4 ± 0.1 (217.9) | 41.7 ± 1.2 (89.3) | 21.5 ± 0.3 (70.2) | 83.3 ± 0.6 (271.9) | 26.3 ± 1.3 (57.4) | >100 | nd | nd |
| C. elegans CB3474 | 982.9 ± 0.6 | 95.2 ± 0.3 (215.2) | 19.6 ± 0.8 (44.2) | 83.1 ± 0.1 (271.4) | 53.8 ± 0.9 (175.7) | 35.7 ± 1.0 (77.9) | nd | nd | >100 |
| C. elegans VC722 | 985.5 ± 0.7 | 91.8 ± 0.3 (207.5) | 27.2 ± 1.4 (61.4) | 96.3 ± 0.07 (314.3) | 93.8 ± 0.3 (306.3) | 43.7 ± 2.1 (95.4) | nd | >100 | nd |
| C. elegans DA1316 | 999.3 ± 1.2 | 96.3 ± 0.3 (217.7) | 27.4 ± 1.1 (61.9) | 82.5 ± 0.1 (269.4) | 87.5 ± 0.5 (285.7) | 40.8 ± 0.3 (89.1) | nd | >100 | nd |
| Compounds | Cytotoxicity μg/mL (μM) | Selectivity Index (SI) | |||
|---|---|---|---|---|---|
| Means CC50 ± SD | |||||
| Caco-2 | C. elegans | O. ochengi | |||
| Wild Type | Males | Females | Microfilariae | ||
| CE | 93.2 ± 1.1 | 0.3 | 8.1 | 8.5 | 8.6 |
| CG | 66.3 ± 0.6 (149.9) | 0.7 | 8.7 | 14.7 | 15.8 |
| ECG | 67.6 ± 0.5 (152.8) | 2.0 | 32.2 | 32.2 | 67.6 |
| GC | 66.7 ± 0.7 (217.8) | 0.9 | 15.9 | 12.1 | 20.8 |
| EGC | 47.1 ± 0.5 (153.8) | 0.8 | 22.4 | 14.3 | 14.3 |
| EGCG | 60.9 ± 0.8 (132.9) | 1.2 | 50.8 | 60.9 | 46.9 |
| Levamisole | 27.3 ± 0.8 (31.2) | 18.8 | 27.3 | 13.0 | 27.3 |
| Ivermectin | 28.7 ± 0.7 (140.5) | 23.9 | 23.9 | 19.1 | 19.1 |
| Albendazole | 29.4 ± 0.5 (110.8) | 4.1 | 7 | 29.4 | 14 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dikti Vildina, J.; Kalmobe, J.; Djafsia, B.; Schmidt, T.J.; Liebau, E.; Ndjonka, D. Anti-Onchocerca and Anti-Caenorhabditis Activity of a Hydro-Alcoholic Extract from the Fruits of Acacia nilotica and Some Proanthocyanidin Derivatives. Molecules 2017, 22, 748. https://doi.org/10.3390/molecules22050748
Dikti Vildina J, Kalmobe J, Djafsia B, Schmidt TJ, Liebau E, Ndjonka D. Anti-Onchocerca and Anti-Caenorhabditis Activity of a Hydro-Alcoholic Extract from the Fruits of Acacia nilotica and Some Proanthocyanidin Derivatives. Molecules. 2017; 22(5):748. https://doi.org/10.3390/molecules22050748
Chicago/Turabian StyleDikti Vildina, Jacqueline, Justin Kalmobe, Boursou Djafsia, Thomas J. Schmidt, Eva Liebau, and Dieudonne Ndjonka. 2017. "Anti-Onchocerca and Anti-Caenorhabditis Activity of a Hydro-Alcoholic Extract from the Fruits of Acacia nilotica and Some Proanthocyanidin Derivatives" Molecules 22, no. 5: 748. https://doi.org/10.3390/molecules22050748
APA StyleDikti Vildina, J., Kalmobe, J., Djafsia, B., Schmidt, T. J., Liebau, E., & Ndjonka, D. (2017). Anti-Onchocerca and Anti-Caenorhabditis Activity of a Hydro-Alcoholic Extract from the Fruits of Acacia nilotica and Some Proanthocyanidin Derivatives. Molecules, 22(5), 748. https://doi.org/10.3390/molecules22050748