Identification of Carotenoids and Isoprenoid Quinones from Asaia lannensis and Asaia bogorensis






Abstract
:1. Introduction
2. Results and Discussion
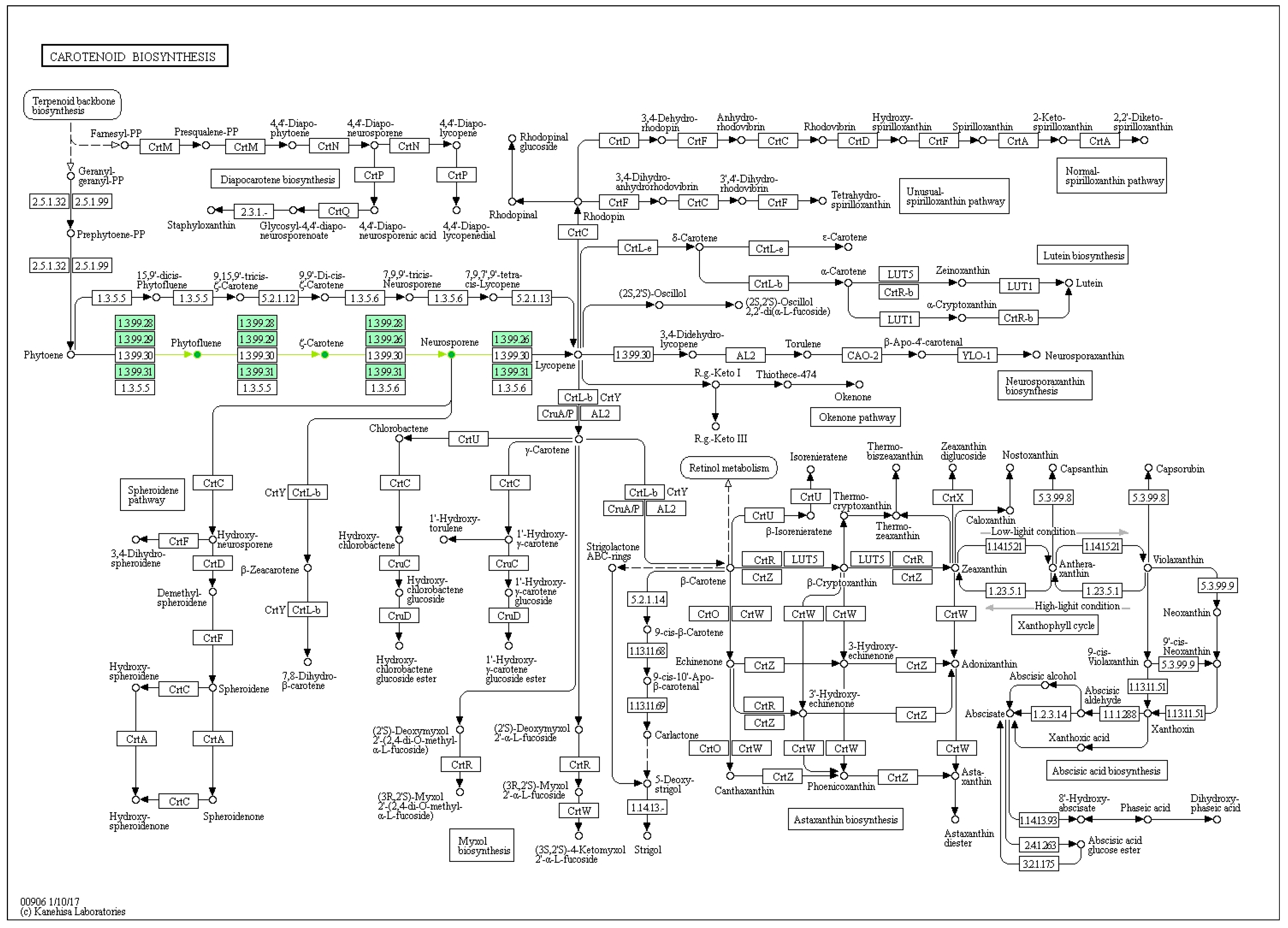
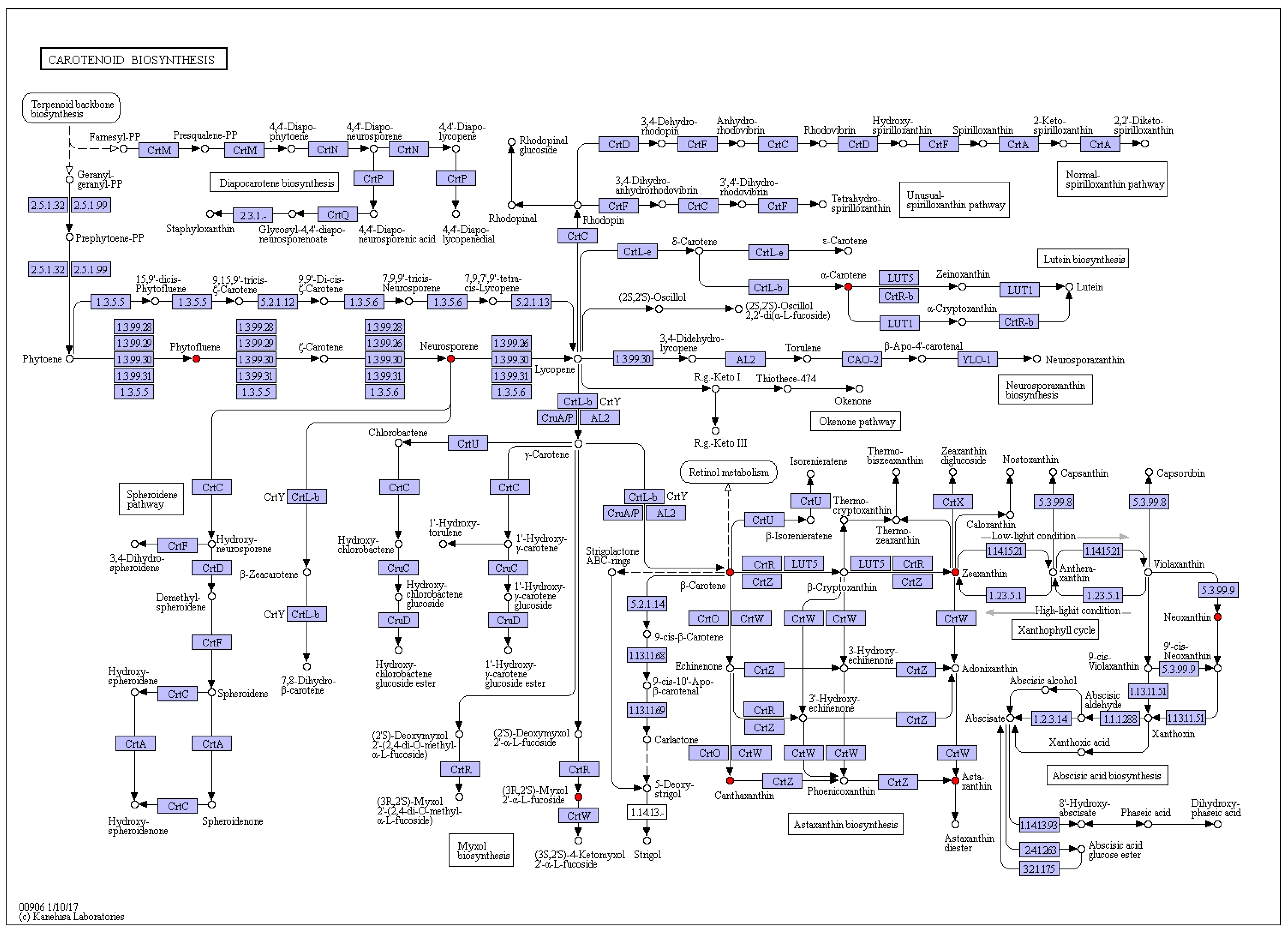
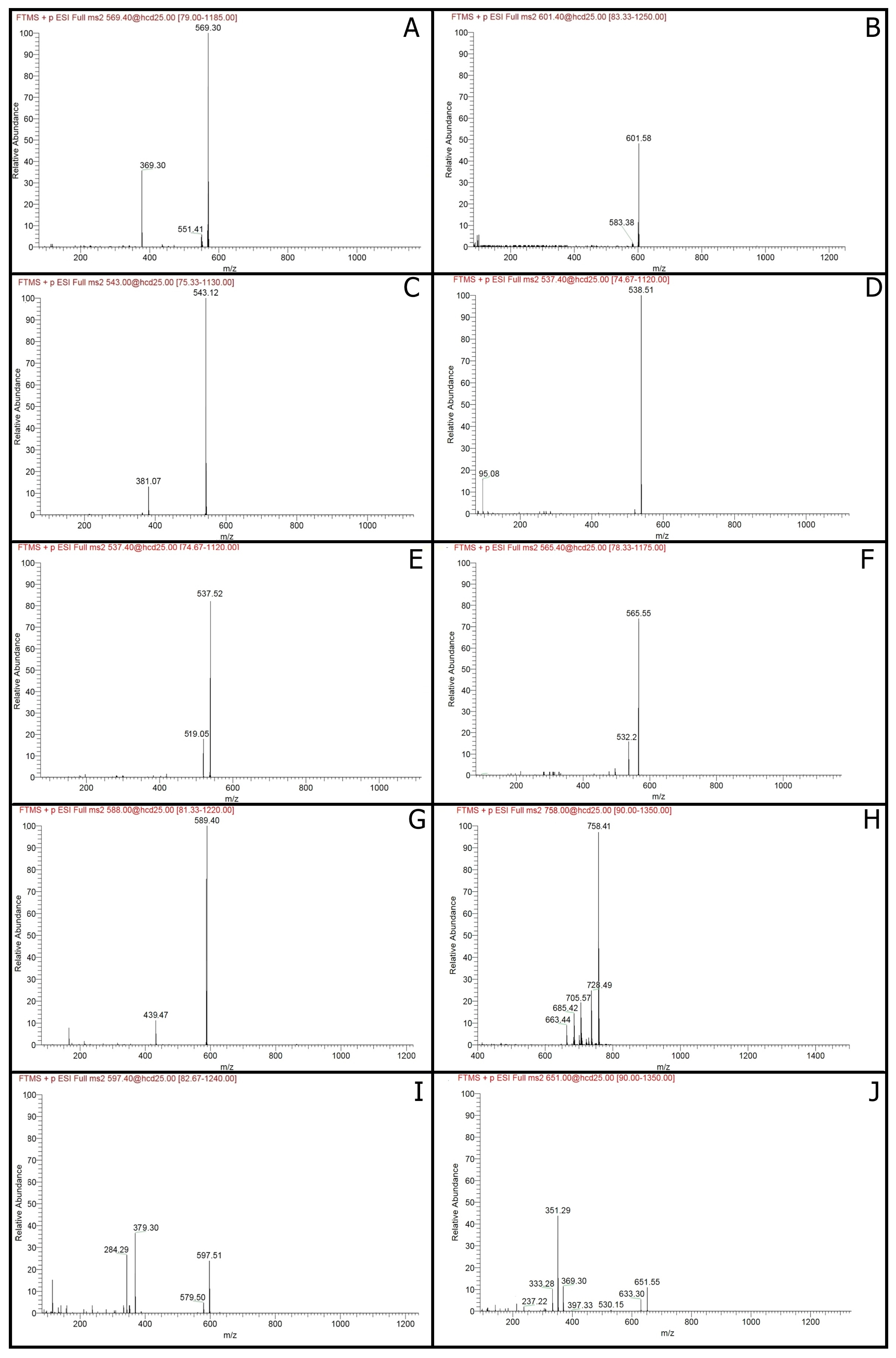
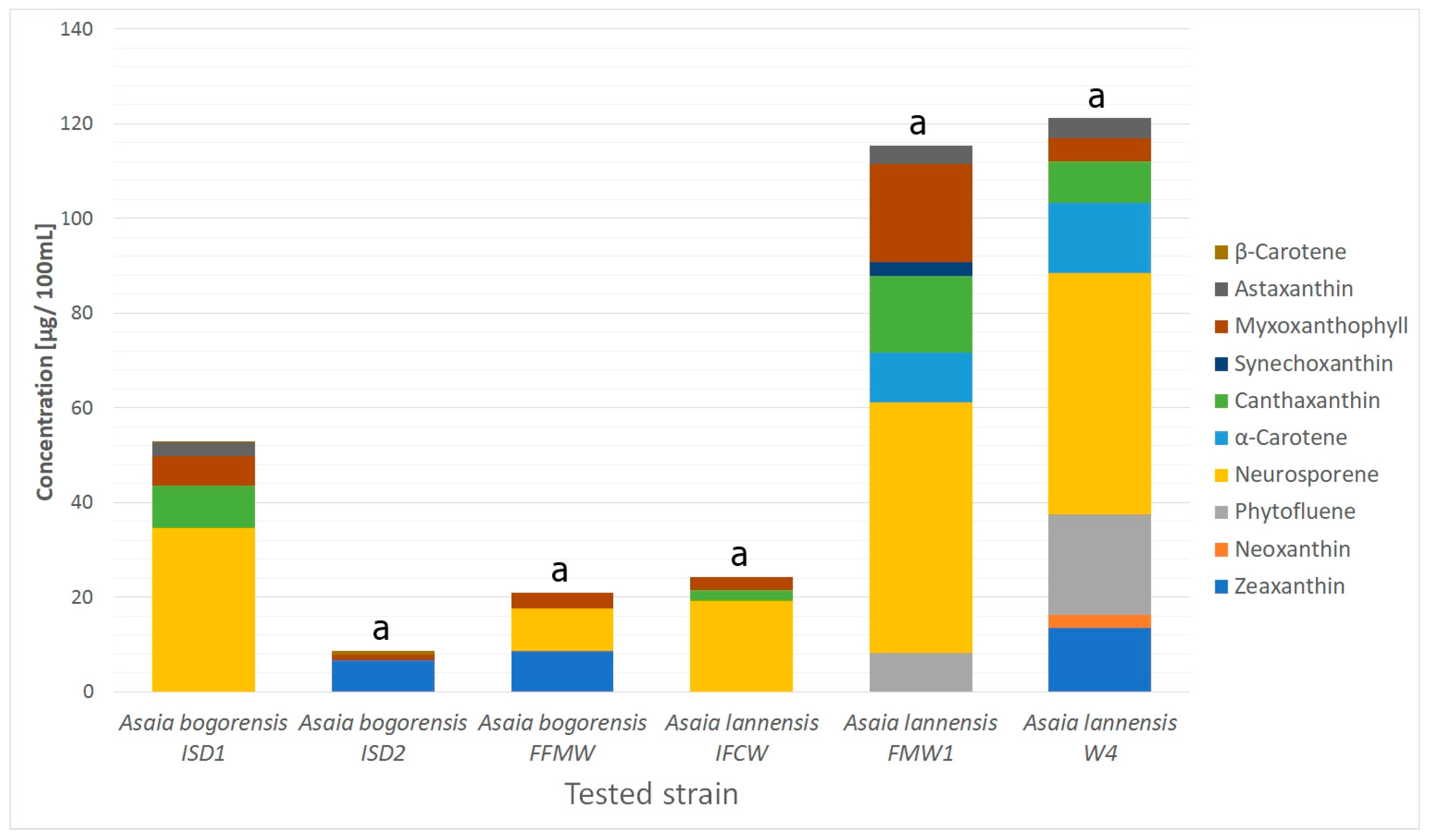
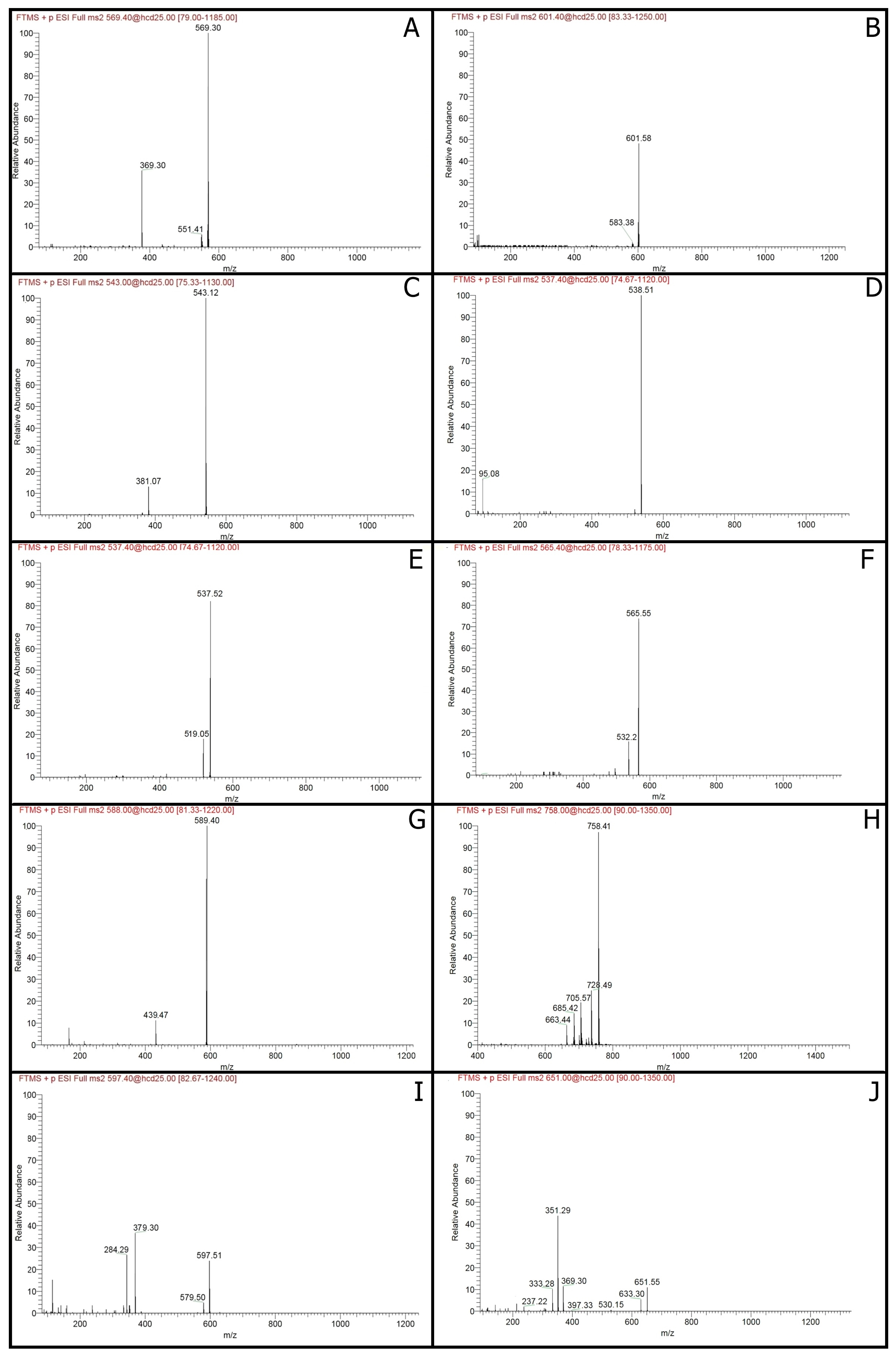
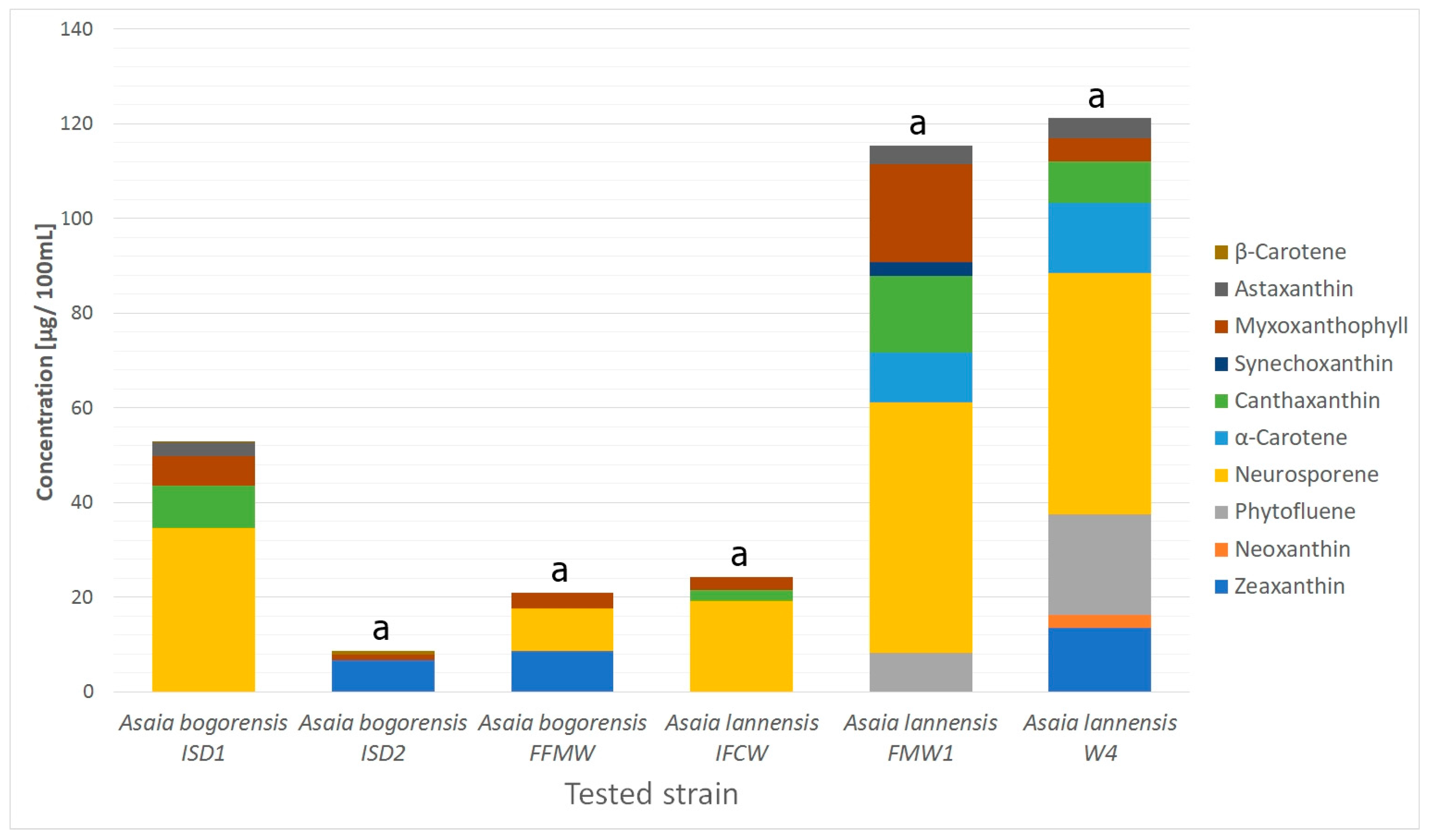
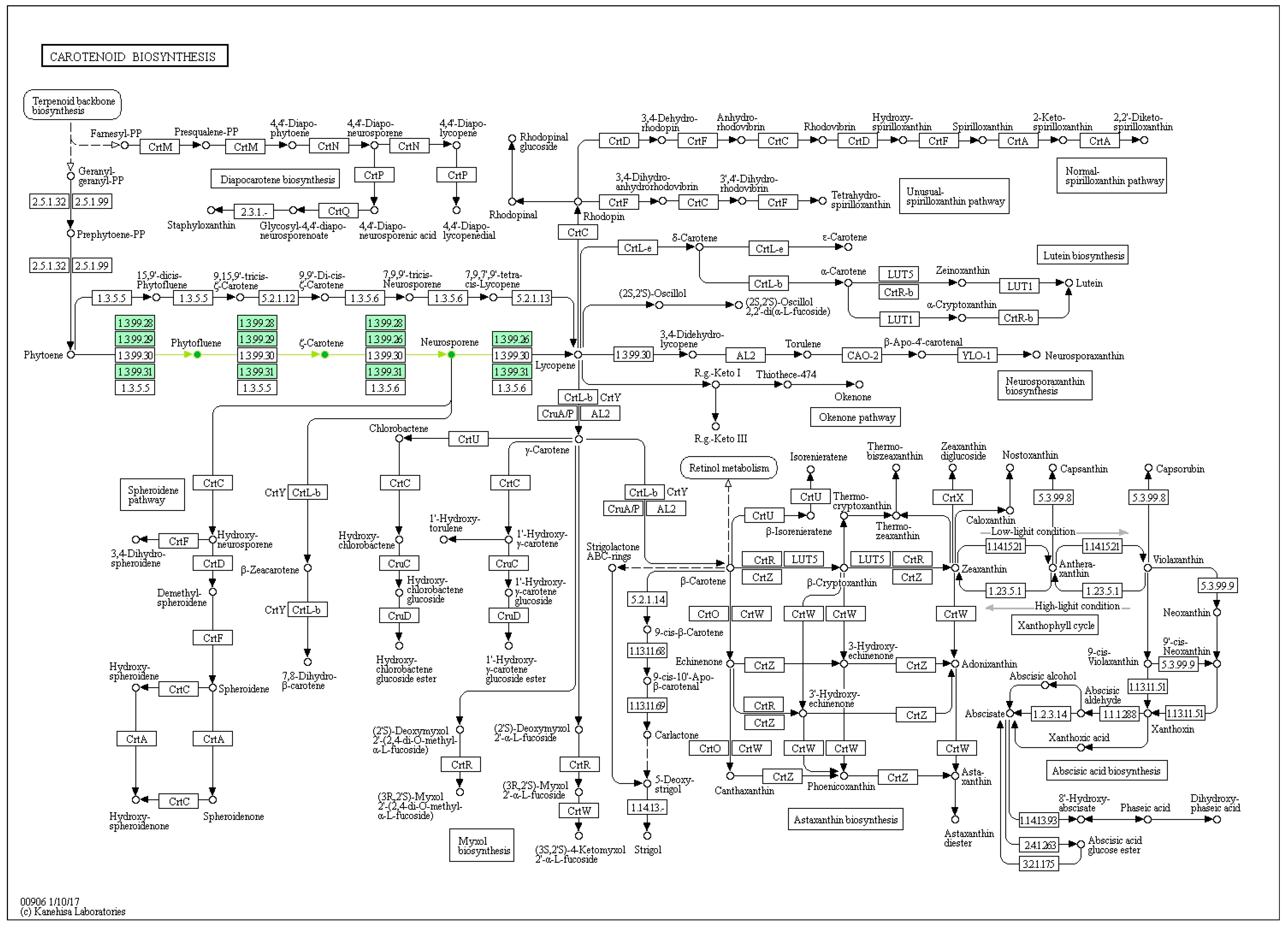
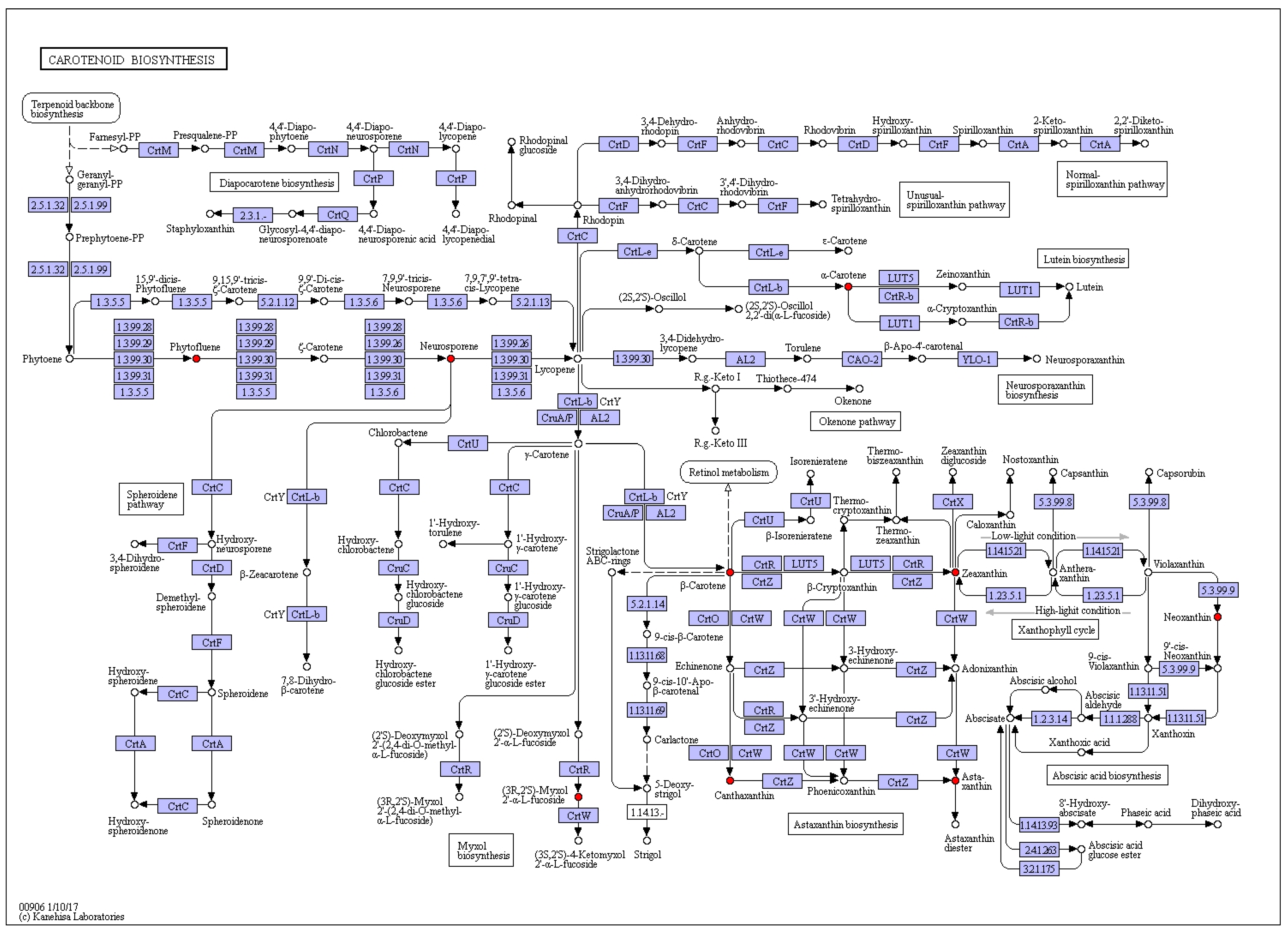
2.1. Carotenoids Identification
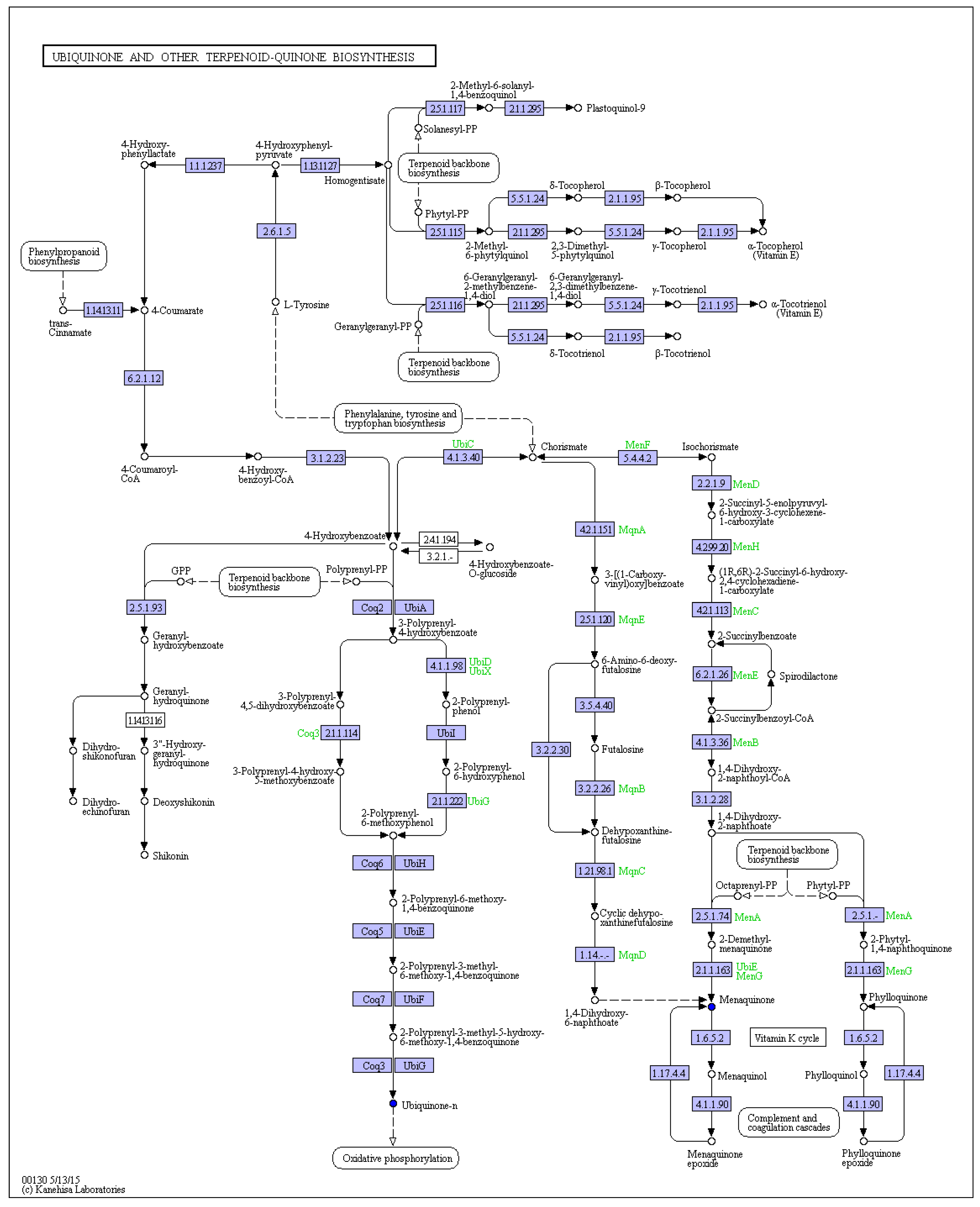
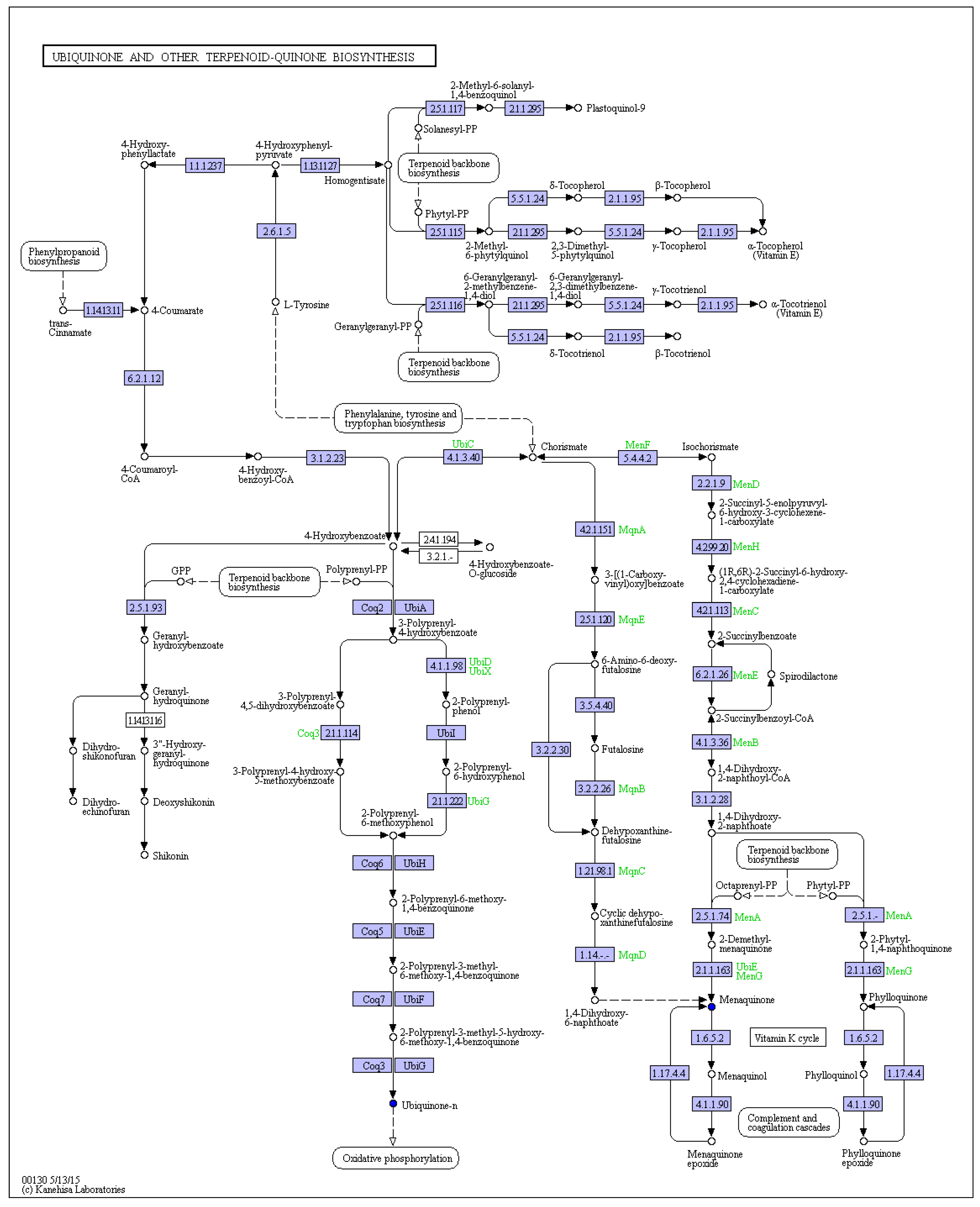
2.2. Isoprenoid Quinones Identification
3. Materials and Methods
3.1. Materials
3.1.1. Bacterial Strains
3.1.2. Chemicals and Standards
3.2. Methods
3.2.1. Culture Conditions
3.2.2. Extraction of Carotenoids
3.2.3. UHPLC-DAD-ESI-MS Analysis
3.3. Statistics
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A
References
- Kirti, K.; Amita, S.; Priti, S.; Kumar, A.M.; Jyoti, S. Colorful world of microbes: Carotenoids and their applications. Adv. Biol. 2014, 2014, 837891. [Google Scholar] [CrossRef]
- Köcher, S.; Breitenbach, J.; Müller, V.; Sandmann, G. Structure, function and biosynthesis of carotenoids in the moderately halophilic bacterium Halobacillus halophilus. Arch. Microbiol. 2009, 191, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, A.; Shizuri, Y.; Hoshino, T.; Sandmann, G. Thermocryptoxanthins: Novel intermediates in the carotenoid biosynthetic pathway of Thermus thermophilus. Arch. Microbiol. 1996, 165, 342–345. [Google Scholar] [CrossRef] [PubMed]
- Fong, N.J.C.; Burgess, M.L.; Barrow, K.D.; Glenn, D.R. Carotenoid accumulation in the psychrotrophic bacterium Arthrobacter agilis in response to thermal and salt stress. Appl. Microbiol. Biotechnol. 2001, 56, 750–756. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Terato, H.; Yamamoto, O. Pigments of Rubrobacter radiotolerans. Arch. Microbiol. 1994, 162, 414–421. [Google Scholar] [CrossRef]
- Lutnaes, B.F.; Oren, A.; Liaaen-Jensen, S. New C(40)-carotenoid acyl glycoside as principal carotenoid in Salinibacter ruber, an extremely halophilic eubacterium. J. Nat. Prod. 2002, 65, 1340–1343. [Google Scholar] [CrossRef] [PubMed]
- Adamiak, J.; Otlewska, A.; Gutarowska, B. Halophilic microbial communities in deteriorated buildings. World J. Microbiol. Biotechnol. 2015, 31, 1489–1499. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.N.V.; Kampe, B.; Röscha, P.; Popp, J. Characterization of carotenoids in soil bacteria and investigation of their photodegradation by UVA radiation via resonance Raman spectroscopy. Analyst 2015, 140, 4584–4593. [Google Scholar] [CrossRef] [PubMed]
- Shindo, K.; Misawa, N. New and rare carotenoids isolated from marine bacteria and their antioxidant activities. Mar. Drugs 2014, 12, 1690–1698. [Google Scholar] [CrossRef] [PubMed]
- Mandelli, F.; Miranda, V.; Rodrigues, E.; Mercadante, A.Z. Identification of carotenoids with high antioxidant capacity produced by extremophile microorganisms. World J. Microbiol. Biotechnol. 2012, 28, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Katsura, K.; Kawasaki, H.; Widyastuti, Y.; Saono, S.; Seki, T.; Uchimura, T.; Komagata, K. Asaia bogorensis gen. nov., sp. nov., an unusual acetic acid bacterium in the α-Proteobacteria. Int. J. Syst. Evolut. Microbiol. 2000, 2, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Fleet, G.H.; Heard, G.M. Lactic acid bacteria associated with wine grapes from several Australian vineyards. Appl. Microbiol. 2006, 100, 712–727. [Google Scholar] [CrossRef] [PubMed]
- Favia, G.; Ricci, I.; Damiani, C.; Raddadi, N.; Crotti, E.; Marzorati, M.; Rizzi, A.; Urso, R.; Brusetti, L.; Borin, S.; et al. Bacteria of the genus Asaia stably associate with Anopheles stephensi, an Asian malarial mosquito vector. Proc. Natl. Acad. Sci. USA 2007, 104, 9047–9051. [Google Scholar] [CrossRef] [PubMed]
- Tuuminen, T.; Roggenkamp, A.; Vuopio-Varkila, J. Comparison of two bacteremic Asaia bogorensis isolates from Europe. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 523–524. [Google Scholar] [CrossRef] [PubMed]
- Carretto, E.; Visiello, R.; Bardaro, M.; Schivazappa, S.; Vailati, F.; Farina, C.; Barbarini, D. Asaia lannensis bacteremia in a ‘needle freak’ patient. Future Microbiol. 2016, 11, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.E.; McCalmont, M.; Xu, J.; Millar, B.C.; Heaney, N. Asaia sp., an unusual spoilage organism of fruit-flavored bottled water. Appl. Environ. Microbiol. 2002, 68, 4130–4131. [Google Scholar] [CrossRef] [PubMed]
- Horsáková, I.; Voldřich, M.; Čeřovskỳ, M.; Sedláčková, P.; Šicenrová, P.; Ulbri, P. Asaia sp. as a bacterium decaying the packaged still fruit beverages. Czech J. Food Sci. 2009, 27, 362–365. [Google Scholar]
- Kręgiel, D.; Otlewska, A.; Antolak, H. Attachment of Asaia bogorensis originating in fruit-flavored water to packaging materials. Biomed. Res. Int. 2014, 2014, 514190. [Google Scholar]
- Antolak, H.; Czyżowska, A.; Kręgiel, D. Black currant (Ribes nigrum L.) and bilberry (Vaccinium myrtillus L.) fruit juices inhibit adhesion of Asaia spp. Biomed. Res. Int. 2016, 2016, 3671306. [Google Scholar] [CrossRef] [PubMed]
- Kawai, M.; Higashiura, N.; Hayasaki, K.; Okamoto, N.; Takami, A.; Hirakawa, H.; Matsushita, K.; Azuma, Y. Complete genome and gene expression analyses of Asaia bogorensis reveal unique responses to culture with mammalian cells as a potential opportunistic human pathogen. DNA Res. 2015, 22, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Maresca, J.A.; Graham, J.E.; Bryant, D.A. The biochemical basis for structural diversity in the carotenoids of chlorophototrophic bacteria. Photosynth. Res. 2008, 97, 121–140. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S.; Maoka, T. Identification and spectroscopic characterization of neurosporene. Biotechnol. Lett. 2015, 37, 2027–2031. [Google Scholar] [CrossRef] [PubMed]
- Moskalenko, A.A.; Makhneva, Z.K. Light-harvesting complexes from purple sulfur bacteria Allochromatium minutissimum assembled without carotenoids. J. Photochem. Photobiol. B 2012, 1, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S.; Maoka, T.; Takasaki, K.; Hanada, S. Carotenoids of Gemmatimonas aurantiaca (Gemmatimonadetes): Identification of a novel carotenoid, deoxyoscillol 2-rhamnoside, and proposed biosynthetic pathway of oscillol 2,2′-dirhamnoside. Microbiology 2010, 156, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.E.; Bryant, D.A. The biosynthetic pathway for myxol-2’ fucoside (myxoxanthophyll) in the cyanobacterium Synechococcus sp. strain PCC 7002. J. Bacteriol. 2009, 191, 3292–3300. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.E.; Lecomte, J.T.; Bryant, D.A. Synechoxanthin, an aromatic C40 xanthophyll that is a major carotenoid in the cyanobacterium Synechococcus sp. PCC 7002. J. Nat. Prod. 2008, 71, 1647–1650. [Google Scholar] [CrossRef] [PubMed]
- Beuttler, H.; Hoffmann, J.; Jeske, M.; Hauer, B.; Schmid, R.D.; Altenbuchner, J.; Urlacher, V.B. Biosynthesis of zeaxanthin in recombinant Pseudomonas putida. Appl. Microbiol. Biotechnol. 2011, 89, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Sajilata, M.G.; Singhal, R.S.; Kamat, M.Y. The carotenoid pigment zeaxanthin—A review. Compr. Rev. Food Sci. Food Saf. 2008, 7, 29–49. [Google Scholar] [CrossRef]
- Choi, S.K.; Matsuda, S.; Hoshino, T.; Peng, X.; Misawa, N. Characterization of bacterial beta-carotene 3,3’-hydroxylases, CrtZ, and P450 in astaxanthin biosynthetic pathway and adonirubin production by gene combination in Escherichia coli. Appl. Microbiol. Biotechnol. 2006, 72, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, Q.; Luo, X.; Fang, C.; Zhang, Q.; Tang, Y. Knockout of crtB or crtI gene blocks the carotenoid biosynthetic pathway in Deinococcus radiodurans R1 and influences its resistance to oxidative DNA-damaging agents due to change of free radicals scavenging ability. Arch. Microbiol. 2007, 188, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Kręgiel, D. Attachment of Asaia lannensis to materials commonly used in beverage industry. Food Control 2013, 32, 537–542. [Google Scholar] [CrossRef]
- Vaara, M. Agents that increase the permeability of the outer membrane. Microbiol. Rev. 1992, 56, 395–411. [Google Scholar] [PubMed]
- Kaiser, P.; Geyer, R.; Surmann, P.; Fuhrmann, H. LC-MS method for screening unknown microbial carotenoids and isoprenoid quinones. J. Microbiol. Methods 2012, 88, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Sutthiwong, N.; Fouillaud, M.; Valla, A.; Caro, Y.; Dufossé, L. Bacteria belonging to the extremely versatile genus Arthrobacter as novel source of natural pigments with extended hue range. Food Res. Int. 2014, 65, 156–162. [Google Scholar] [CrossRef]
- Bidaud, P.; Hébert, L.; Barbey, C.; Appourchaux, A.-C.; Torelli, R.; Sanguinetti, M.; Laugier, C.; Petry, S. Rhodococcus equi’s extreme resistance to hydrogen peroxide is mainly conferred by one of its four catalase genes. PLoS ONE 2012, 7, e42396. [Google Scholar] [CrossRef] [PubMed]
- Berenjian, A.; Mahanama, R.; Talbot, A.; Biffin, R.; Regtop, H.; Valtchev, P.; Kavanagh, J.; Dehghani, F. Efficient media for high menaquinone-7 production: Response surface methodology approach. New Biotechnol. 2011, 28, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Nowicka, B.; Kruk, J. Occurrence, biosynthesis and function of isoprenoid quinones. Biochim. Biophys. Acta 2010, 1797, 1587–1605. [Google Scholar] [CrossRef] [PubMed]
- Walther, B.; Karl, J.P.; Booth, S.L.; Boyaval, P. Menaquinones, bacteria, and the food supply: The relevance of dairy and fermented food products to vitamin K requirements. Adv. Nutr. 2013, 4, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Berenjian, A.; Chan, N.L.; Mahanama, R.; Talbot, A.; Regtop, H.; Kavanagh, J.; Dehghani, F. Effect of biofilm formation by Bacillus subtilis natto on menaquinone-7 biosynthesis. Mol. Biotechnol. 2013, 54, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, M.; Kagihiro, S.; Haruki, M.; Takano, K.; Branda, S.; Kolter, R.; Kanaya, S. Biofilm formation by a Bacillus subtilis strain that produces γ-polyglutamate. Microbiology 2006, 152, 2801–2807. [Google Scholar] [CrossRef] [PubMed]
- Malimas, T.; Yukphan, P.; Takahashi, M.; Kaneyasu, M.; Potacharoen, W.; Tanasupawat, S.; Nakagawa, Y.; Tanticharoen, M.; Yamada, Y. Asaia lannaensis sp. nov., a new acetic acid bacterium in the Alphaproteobacteria. Biosci. Biotechnol. Biochem. 2008, 72, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.D.; Jones, D. Distribution of isoprenoid quinone structural types in bacteria and their taxonomic implication. Microbiol. Rev. 1981, 45, 316–354. [Google Scholar] [PubMed]
- Zhang, H.; Xu, X.; Chen, X.; Yuan, F.; Sun, B.; Xu, Y.; Yang, J.; Sun, D. Complete genome sequence of the cellulose-producing strain Komagataeibacter nataicola RZS01. Sci. Rep. 2017, 7, 4431. [Google Scholar] [CrossRef] [PubMed]
- Antolak, H.; Czyzowska, A.; Sakač, M.; Mišan, A.; Đuragić, O.; Kregiel, D. Phenolic compounds contained in little-known wild fruits as antiadhesive agents against the beverage-spoiling bacteria Asaia spp. Molecules 2017, 22, 1256. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Chen, J.; Jiang, Q.; Wang, X.; Lu, Y.; Gong, L.; Chen, D. Simultaneous determination of three active components in rat plasma by UPLC-MS/MS: Application to pharmacokinetic study after oral administration of Herba Sarcandrae extract. Biomed. Chromatogr. 2017, 31, e3834. [Google Scholar] [CrossRef] [PubMed]
- Rivera, S.M.; Christou, P.; Canela-Garayoa, R. Identification of carotenoids using mass spectrometry. Mass Spectrom. Rev. 2014, 33, 353–372. [Google Scholar] [CrossRef] [PubMed]
- Gutarowska, B.; Celikkol-Aydin, S.; Bonifay, V.; Otlewska, A.; Aydin, E.; Oldham, A.L.; Brauer, I.; Duncan, K.E.; Adamiak, J.; Sunner, J.A.; et al. Metabolomic and high-throughput sequencing analysis—Modern approach for the assessment of biodeterioration of materials from historic buildings. Front. Microbiol. 2015, 6, 979. [Google Scholar] [CrossRef] [PubMed]
- KEGG Mapper. Available online: http://www.genome.jp/kegg/mapper.html (accessed on 21 August 2017).
Sample Availability: Strains of Asaia lannensis and Asaia bogorensis are available from the authors. Samples of the compounds are not available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | tR (min) | λmax (nm) | [M − H]+ m/z | MS/MS Fragments | Compound | Formula | Classification | Compounds Concentration (µg/100 mL) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ISD1 | ISD2 | FFMW | IFCW | FMW1 | W4 | ||||||||
| 1 | 2.62 | 413, 438, 470 | 569.4788 | 551, 369 | Zeaxanthin | C40H56O2 | Carotenoids | nd | 6.6 ± 0.09 | 8.5 ± 0.19 | nd | nd | 13.5 ± 0.61 |
| 2 | 3.58 | 286, 484, 520 | 601.5769 | 583 | Neoxanthin | C40H56O4 | Carotenoids | nd | nd | nd | nd | nd | 2.7 ± 0.19 |
| 3 | 4.76 | 280 | 543.1193 | 381 | Phytofluene | C40H62 | Carotenoids | nd | nd | nd | nd | 8.3 ± 0.22 | 21.3 ± 1.21 |
| 4 | 6.62 | 310, 486, 523 | 538.5083 | 95 | Neurosporene | C40H58 | Carotenoids | 34.6 ± 2.08 | nd | 9.1 ± 0.44 | 19.1 ± 1.63 | 52.9 ± 2.59 | 50.9 ± 3.97 |
| 5 | 9.60 | 320, 486, 523 | 537.5193 | 519 | α-Carotene | C40H56 | Carotenoids | nd | nd | nd | nd | 10.4 ± 0.83 | 14.9 ± 0.63 |
| 6 | 13.87 | 320, 486, 523 | 565.5489 | 532 | Canthaxanthin | C40H52O2 | Carotenoids | 9.0 ± 0.36 | nd | nd | 2.4 ± 0.07 | 16.2 ± 0.77 | 8.7 ± 0.23 |
| 7 | 14.88 | 486 | 589.3875 | 439, 163 | Synechoxanthin | C40H42O4 | Carotenoids | nd | nd | nd | nd | 3.0 ± 0.26 | nd |
| 8 | 15.72 | 300, 486, 509 | 758.4098 | 728, 705, 685, 633 | Myxoxanthophyll | C46H66O8 | Carotenoids | 6.1 ± 0.37 | 1.2 ± 0.03 | 3.2 ± 0.18 | 2.8 ± 0.17 | 20.6 ± 1.56 | 5.0 ± 0.45 |
| 9 | 18.34 | 486 | 597.5089 | 579, 379, 285 | Astaxanthin | C40H52O4 | Carotenoids | 3.0 ± 0.09 | nd | nd | nd | 3.9 ± 0.34 | 4.3 ± 0.27 |
| 10 | 20.07 | 286 | 651.5496 | 633, 397, 369, 351, 333 | Menaquinone-7 | C46H64O2 | Isoprenoid quinones | 3.3 ± 0.12 | 4.5 ± 0.21 | 3.2 ± 0.17 | 4.7 ± 0.22 | 9.8 ± 0.32 | 7.7 ± 0.23 |
| 11 | 21.14 | 450, 470, 486 | 537.3791 | 444, 177 | β-Carotene | C40H56 | Carotenoids | 0.1 ± 0.01 | 0.7 ± 0.03 | nd | nd | nd | nd |
| 12 | 22.56 | 286 | 717.4471 | 575, 187 | Menaquinone-8 | C51H72O2 | Isoprenoid quinones | nd | nd | 0.5 ± 0.01 | 1.0 ± 0.02 | 4.5 ± 0.43 | 3.0 ± 0.12 |
| 13 | 24.92 | 280 | 863.6652 | 197 | Ubiquinone-10 | C59H90O4 | Isoprenoid quinones | 13.4 ± 0.60 | 22.5 ± 1.18 | 21.6 ± 0.95 | 38.1 ± 2.23 | 48.6 ± 4.43 | 6.3 ± 0.25 |
| Total Content | 69.6 ± 3.61 | 35.5 ± 1.55 | 46.1 ± 1.94 | 68.1 ± 4.34 | 178.2 ± 11.74 | 138.3 ± 8.15 | |||||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antolak, H.; Oracz, J.; Otlewska, A.; Żyżelewicz, D.; Kręgiel, D. Identification of Carotenoids and Isoprenoid Quinones from Asaia lannensis and Asaia bogorensis. Molecules 2017, 22, 1608. https://doi.org/10.3390/molecules22101608
Antolak H, Oracz J, Otlewska A, Żyżelewicz D, Kręgiel D. Identification of Carotenoids and Isoprenoid Quinones from Asaia lannensis and Asaia bogorensis. Molecules. 2017; 22(10):1608. https://doi.org/10.3390/molecules22101608
Chicago/Turabian StyleAntolak, Hubert, Joanna Oracz, Anna Otlewska, Dorota Żyżelewicz, and Dorota Kręgiel. 2017. "Identification of Carotenoids and Isoprenoid Quinones from Asaia lannensis and Asaia bogorensis" Molecules 22, no. 10: 1608. https://doi.org/10.3390/molecules22101608
APA StyleAntolak, H., Oracz, J., Otlewska, A., Żyżelewicz, D., & Kręgiel, D. (2017). Identification of Carotenoids and Isoprenoid Quinones from Asaia lannensis and Asaia bogorensis. Molecules, 22(10), 1608. https://doi.org/10.3390/molecules22101608