Agrimoniin, an Active Ellagitannin from Comarum palustre Herb with Anti-α-Glucosidase and Antidiabetic Potential in Streptozotocin-Induced Diabetic Rats




Abstract
:1. Introduction
2. Results and Discussion
2.1. Inhibition of α-Glucosidase by C. palustre Extracts
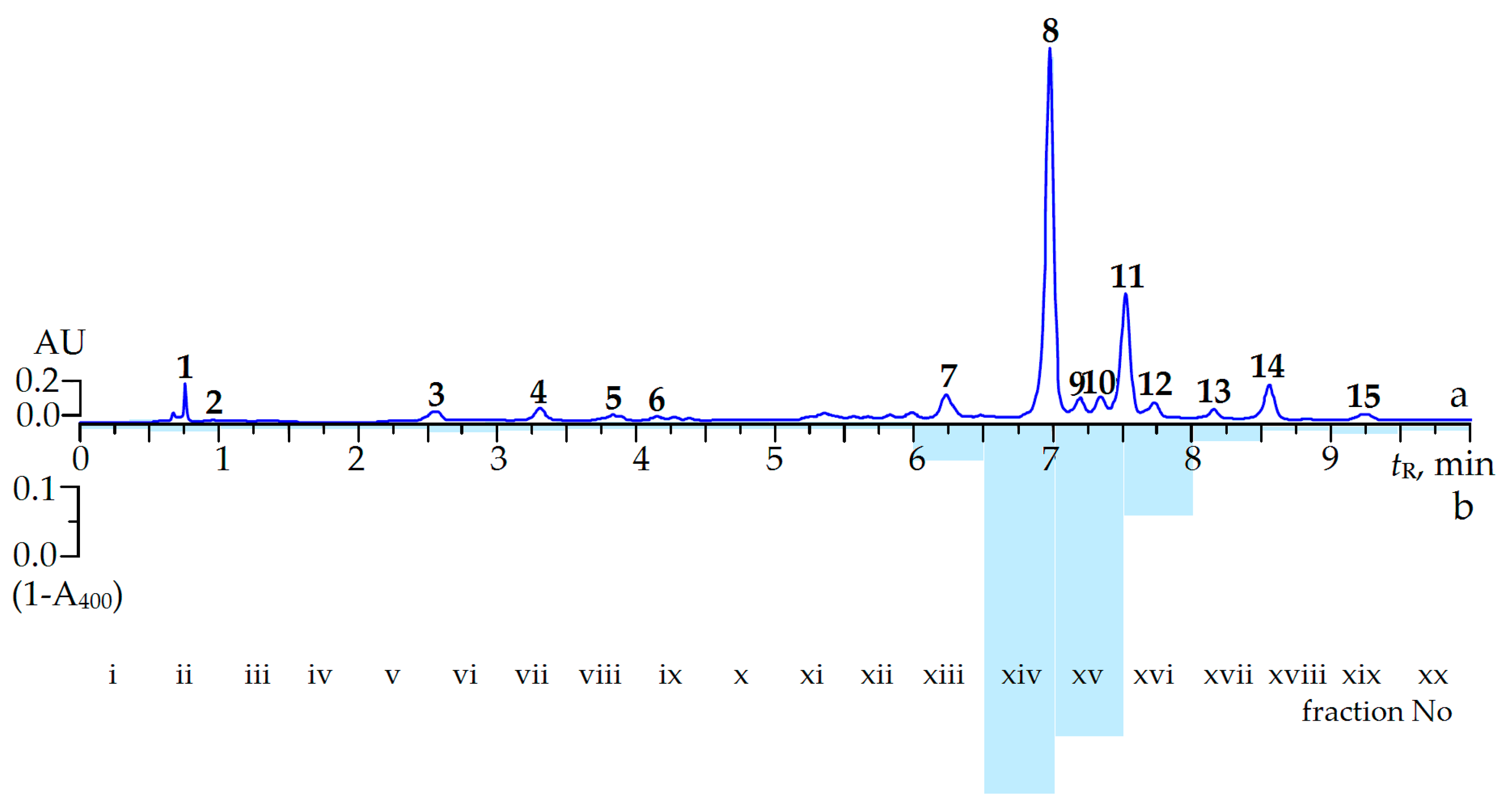
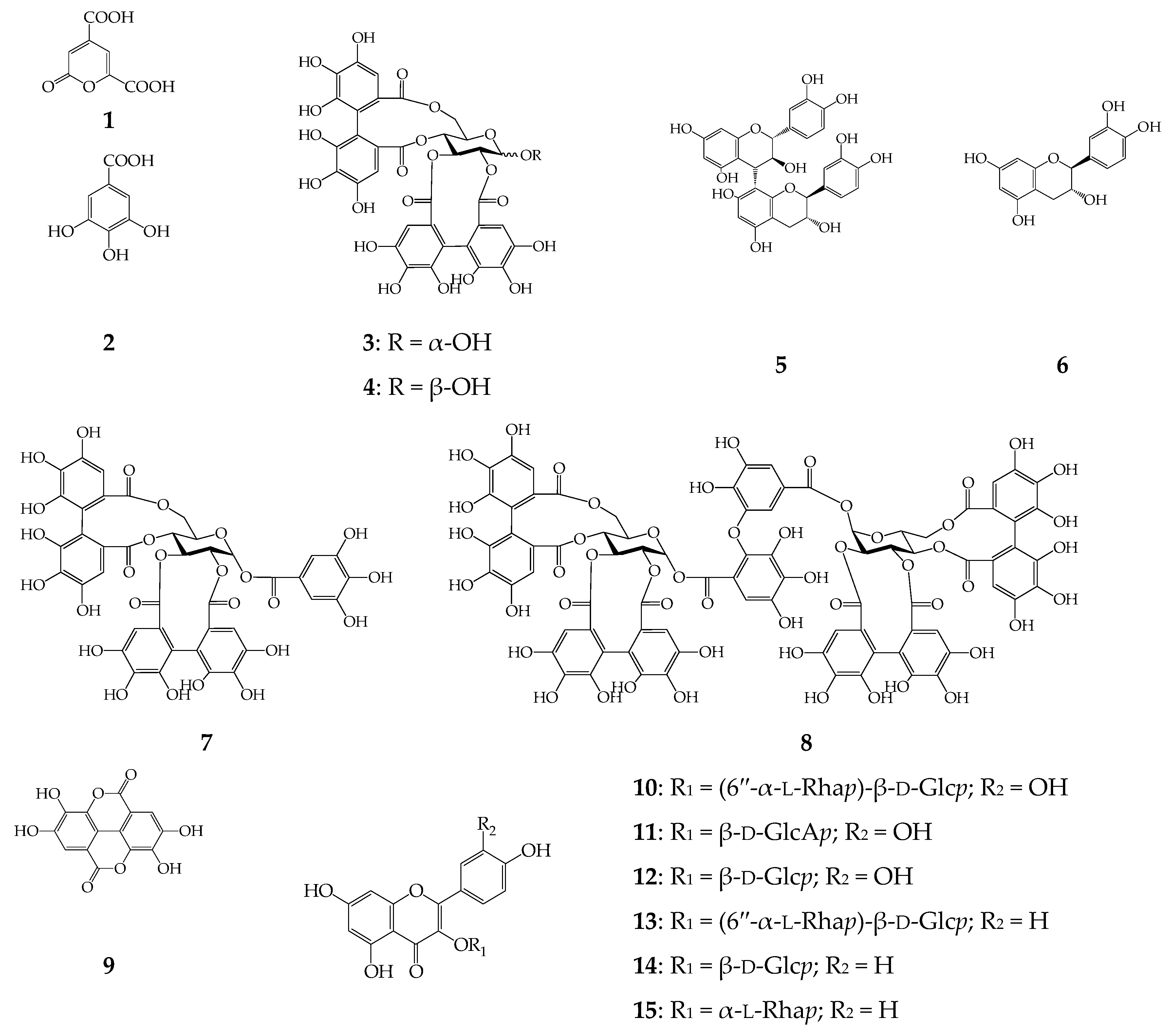
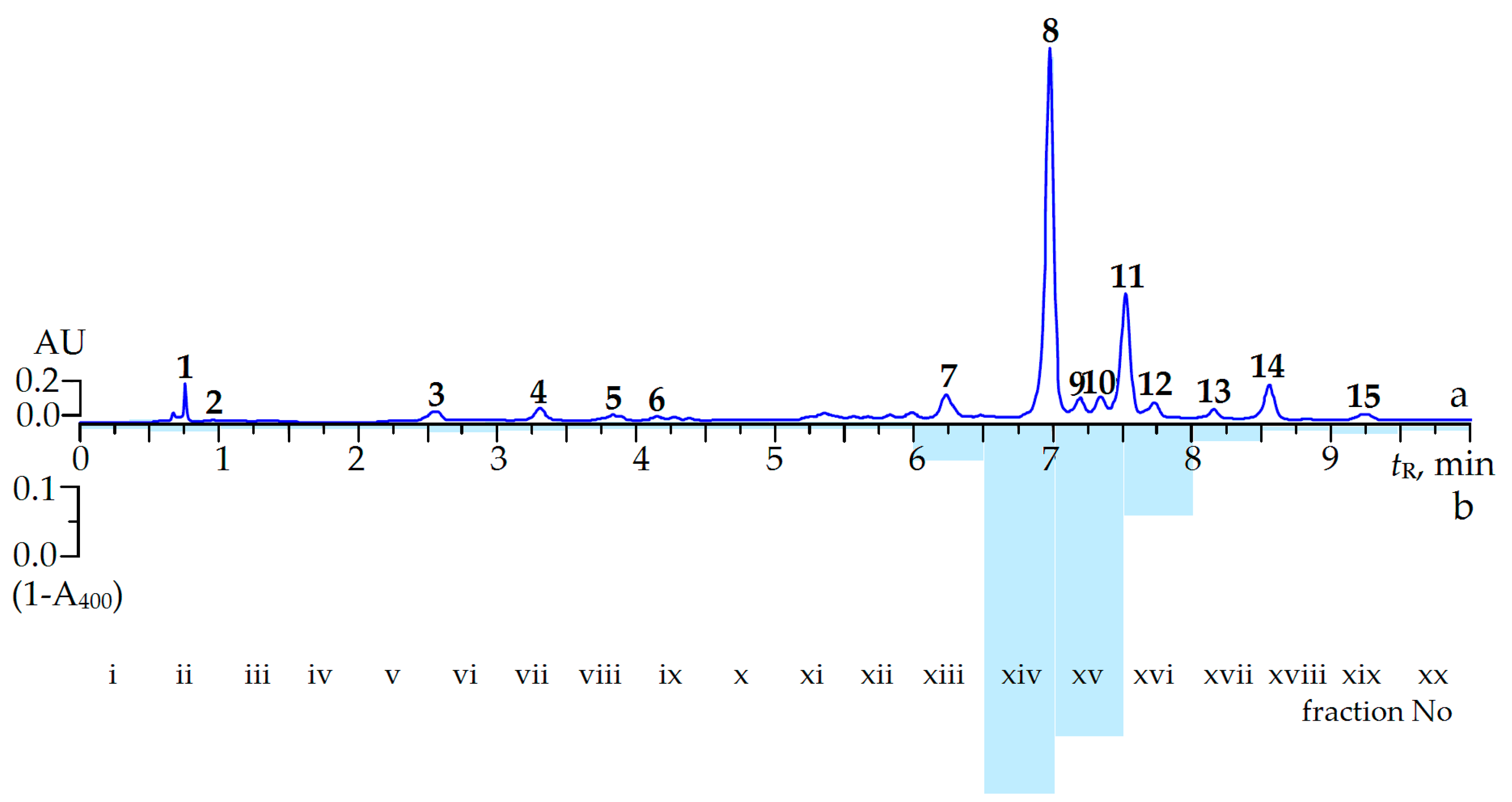
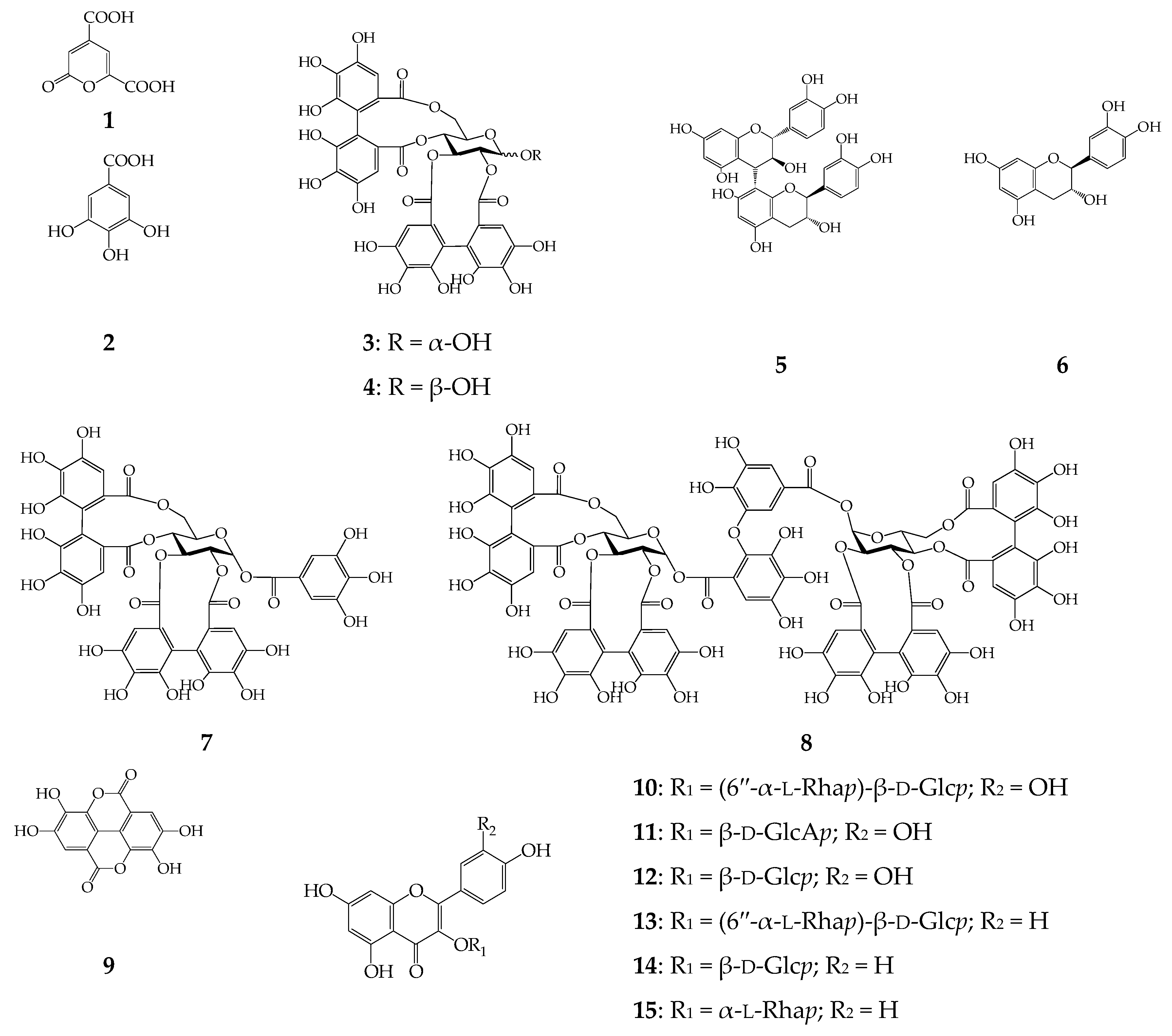
2.2. Screening of α-Glucosidase Inhibitors from C. palustre Herb Extract
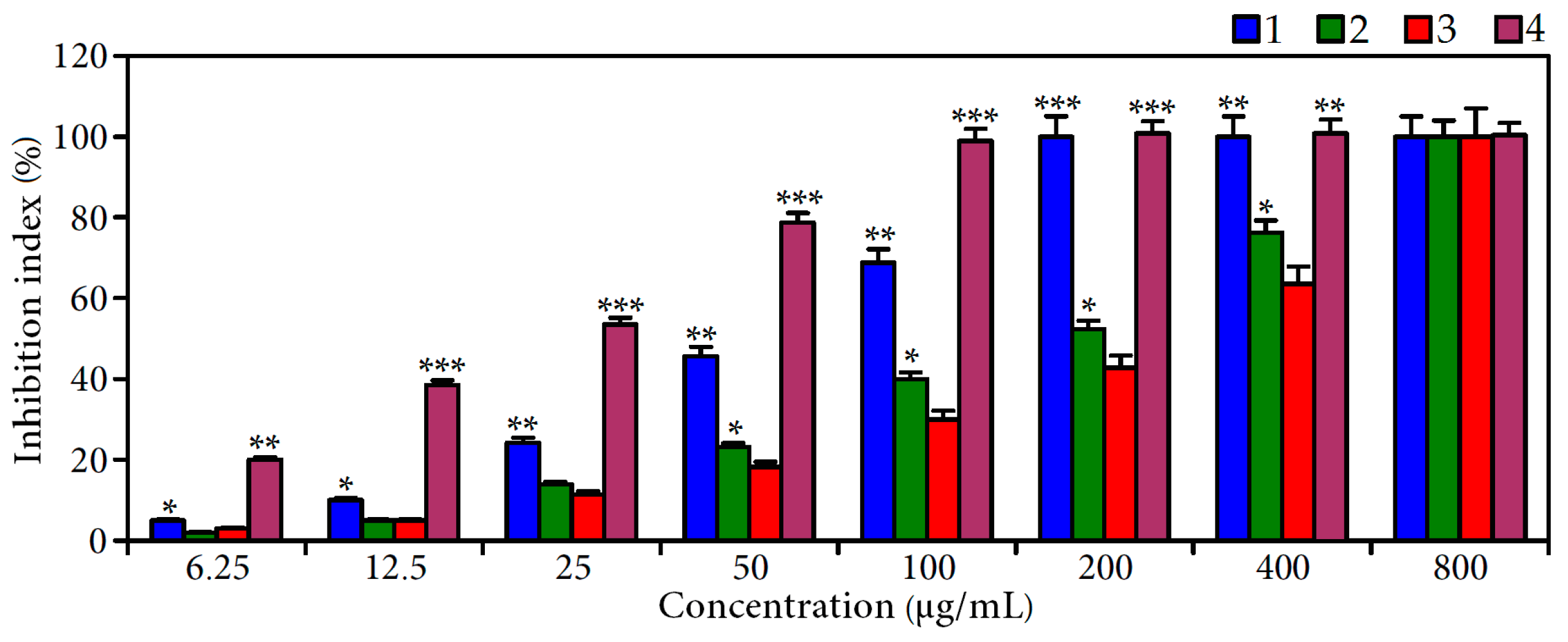
2.3. Effect of Ellagitannin Remove from C. palustre Extract on α-Glucosidase Inhibition
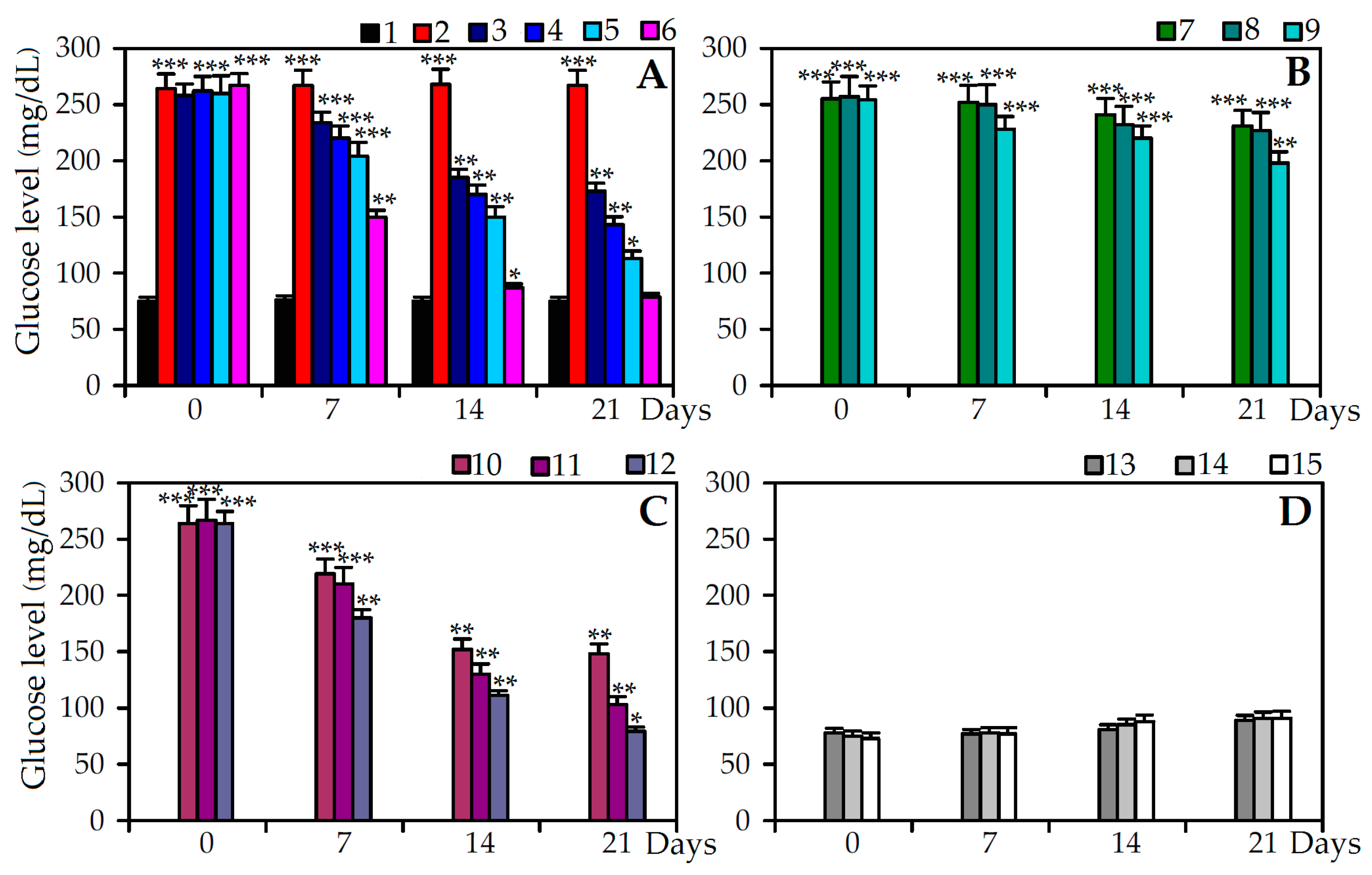
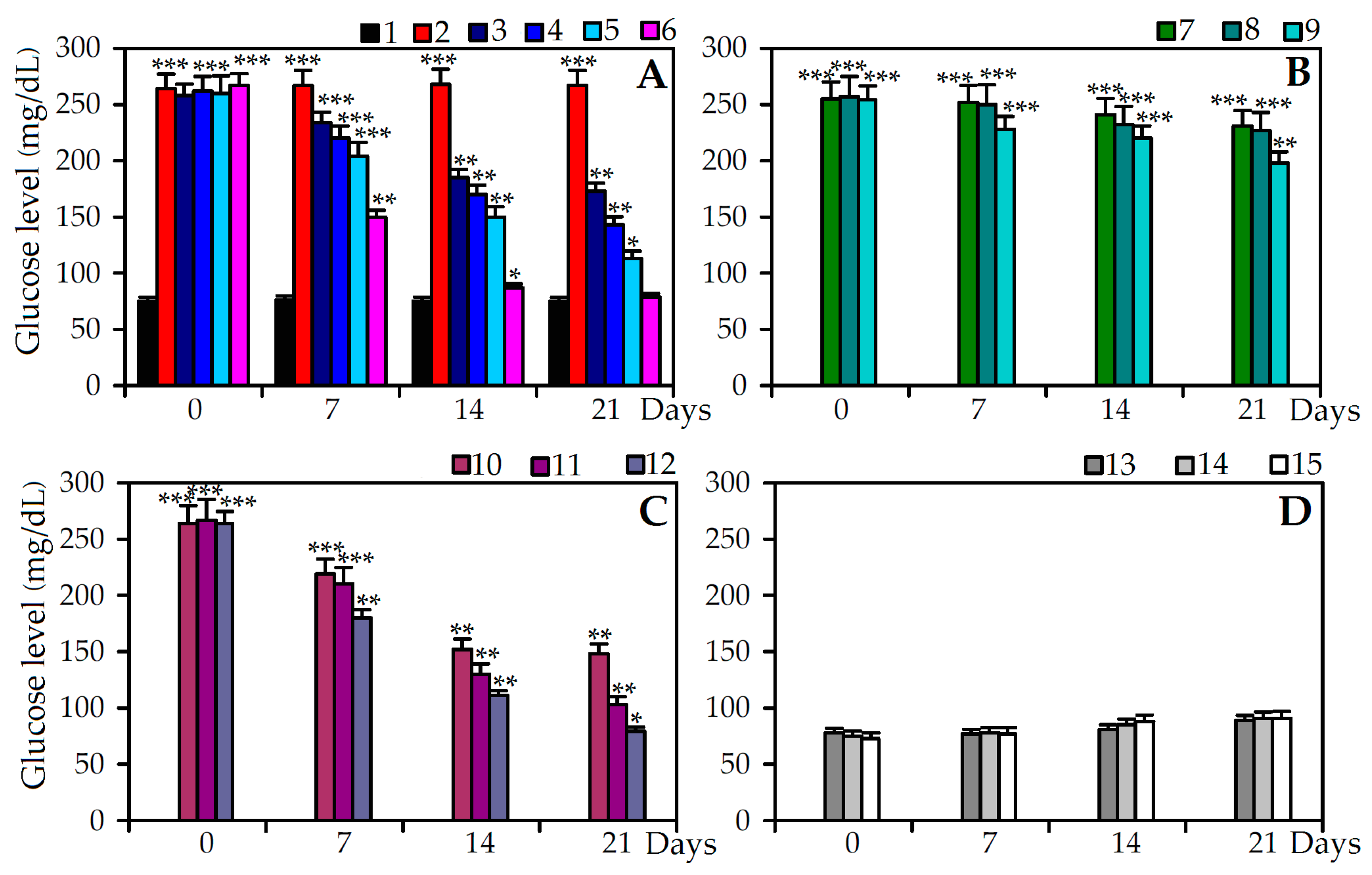
2.4. Antidiabetic Effect of C. palustre Extracts and Agrimoniin in Stretpozotocin Induced Diabetic Rats
3. Materials and Methods
3.1. Plant Materials and Chemicals
3.2. Sample Preparation
3.3. MC-RP-HPLC-UV-MS and MC-HPLC-UV Conditions
3.4. HPLC Activity-Based Profiling
3.5. Ellagitannin Remove SPE-Procedure
3.6. α-Glucosidase Inhibiting Assay
3.7. Hypoglycemic Activity
3.7.1. Experimental Animals
3.7.2. Hyperglycemia Induction and Experimental Design
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Health Organization. Global Report on Diabetes; World Health Organization: Geneva, Switzerland, 2016.
- Ríos, J.L.; Francini, F.; Schinella, G.R. Natural products for the treatment of type 2 diabetes mellitus. Planta Med. 2015, 81, 975–994. [Google Scholar] [CrossRef] [PubMed]
- Scheen, A.J. Is there a role for α-glucosidase inhibitors in the prevention of type 2 diabetes mellitus? Drugs 2003, 63, 933–951. [Google Scholar] [CrossRef] [PubMed]
- Kihara, Y.; Ogami, Y.; Tabaru, A.; Unoki, H.; Otsuki, M. Safe and effective treatment of diabetes mellitus associated with chronic liver diseases with an alpha-glucosidase inhibitor, acarbose. J. Gastroenterol. 1997, 32, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Güemes, M.; Melikyan, M.; Senniappan, S.; Hussain, K. Idiopathic postprandial hyperinsulinic hypoglycaemia. J. Pediatr. Endocrinol. Metab. 2016, 29, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Gray, A.M.; Flatt, P.R. Nature’s own pharmacy: The diabetes perspective. Proc. Nutr. Soc. 1997, 56, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Kai, G.; Yamamoto, K.; Chen, X. Advance in dietary polyphenols as α-glucosidases inhibitors: A review on structure-activity relationship aspect. Crit. Rev. Food Sci. Nutr. 2013, 53, 818–836. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, S.; Yin, P.; Yan, L.; Han, J.; Shi, L.; Zhou, X.; Liu, Y.; Ma, C. α-Glucosidase inhibitory activity of polyphenols from the burs of Castanea mollissima Blume. Molecules 2014, 19, 8373–8386. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.; Wan, C.; Ma, H.; Seeram, N. New phenolics from the flowers of Punica granatum and their in vitro α-glucosidase inhibitory activities. Planta Med. 2013, 79, 1674–1679. [Google Scholar] [CrossRef] [PubMed]
- Flores-Bocanegra, L.; Pérez-Vásquez, A.; Torres-Piedra, M.; Bye, R.; Linares, E.; Mata, R. α-Glucosidase inhibitors from Vauquelinia corymbosa. Molecules 2015, 20. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, K.; Itrat, F.; Azhar, M.; Rehana, A.; Abdul, M.; Sumaira, T.; Muhammad, I.C. Purunusides A-C, α-glucosidase inhibitory homoisoflavone glucosides from Prunus domestica. Arch. Pharm. Res. 2009, 32, 1705–1710. [Google Scholar]
- Šaponjak, V.T.; Gironés-Vilaplana, A.; Djilas, S.; Mena, P.; Ćetković, G.; Moreno, D.A.; Ćanadanović-Brunet, J.; Vulić, J.; Stajćić, S.; Krunić, M. Anthocyanin profiles and biological properties of caneberry (Rubus spp.) press residues. J. Sci. Food Agric. 2014, 94, 2393–2400. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.H.; Wang, J.J.; Gu, X.Z.; Gul, H.P.; Kang, W.Y. Antioxidant and a-glucosidase inhibitory activity of red raspberry (Harrywaters) fruits in vitro. Afr. J. Pharm. Pharmacol. 2012, 6, 3118–3123. [Google Scholar] [CrossRef]
- Traditions and Innovations in Life and Culture of the Siberian Peoples; Rusakova, L.M. (Ed.) SD RAS: Novosibirsk, Russia, 1983; p. 175.
- Makarov, A.A. Plant Remedies of the Traditional Yakutian Medicine; Yakutian Scientific Center: Yakutsk, Russia, 1974; p. 31. [Google Scholar]
- Popov, S.V.; Popova, G.Y.; Ovodova, R.G.; Ovodov, Y.S. Antiinflammatory activity of the pectic polysaccharide from Comarum palustre. Fitoterapia 2005, 76, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Popov, S.V.; Ovodova, R.G.; Markov, P.A.; Nikitina, I.R.; Ovodov, Y.S. Protective effects of comaruman, a pectin of cinquefoil Comarum palustre L. on acetic acid-induced colitis in mice. Dig. Dis. Sci. 2006, 51, 1532–1537. [Google Scholar] [CrossRef] [PubMed]
- Lemus, I.; García, R.; Delvillar, E.; Knop, G. Hypoglycaemic activity of four plants used in Chilean popular medicine. Phytother. Res. 1999, 13, 91–94. [Google Scholar] [CrossRef]
- Elya, B.; Basah, K.; Mun’im, A.; Yuliastuti, W.; Bangun, A.; Septiana, E.K. Screening of α-Glucosidase inhibitory activity from some plants of Apocynaceae, Clusiaceae, Euphorbiaceae, and Rubiaceae. J. Biomed. Biotechnol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.D.; Ranilla, L.G.; Apostolidis, E.; Lajolo, F.M.; Genovese, M.I.; Shetty, K. Evaluation of antihyperglycemia and antihypertension potential of native Peruvian fruits using in vitro models. J. Med. Food 2009, 12, 278–291. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N. Ellagitannins and other phenolic compounds from Comarum palustre. Chem. Nat. Comp. 2016, 52, 721–723. [Google Scholar] [CrossRef]
- Julianti, T.; Mieri, M.D.; Zimmermann, S.; Ebrahimi, S.N.; Kaiser, M.; Neuburger, M.; Raith, M.; Brun, R.; Hamburger, M. HPLC-based activity profiling for antiplasmodial compounds in the traditional Indonesian medicinal plant Carica papaya L. J. Ethnopharmacol. 2014, 155, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Baburin, I.; Plitzko, I.; Hering, S.; Hamburger, M. HPLC-based activity profiling for GABAa receptor modulators from the traditional Chinese herbal drug Kushen (Sophora flavescens root). Mol. Divers. 2011, 15, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, K.; Riese, U.; Hamburger, M. HPLC-based bioactivity profiling of plant extracts: A kinetic assay for the identification of monoamine oxidase-A inhibitors using human recombinant monoamine oxidase-A. Phytochemistry 2004, 65, 2885–2891. [Google Scholar] [CrossRef] [PubMed]
- Potterat, O.; Hamburger, M. Natural products in drug discovery—Concepts and approaches for tracking bioactivity. Curr. Org. Chem. 2006, 10, 899–920. [Google Scholar] [CrossRef]
- Yuan, T.; Ding, Y.; Wan, C.; Li, L.; Xu, J.; Liu, K.; Slitt, A.; Ferreira, D.; Khan, I.A.; Seeram, N.P. Antidiabetic ellagitannins from pomegranate flowers: Inhibition of α-glucosidase and lipogenic gene expression. Org. Lett. 2012, 20, 5358–5361. [Google Scholar] [CrossRef] [PubMed]
- Santos-Buelga, C.; Scalbert, A. Proanthocyanidins and tannin-like compounds—Nature, occurrence, dietary intake and effects on nutrition and health. J. Sci. Food Agric. 2000, 80, 1094–1117. [Google Scholar] [CrossRef]
- Ochir, S.; Nishizawa, M.; Park, J.B.; Yamagashi, T. Inhibitory effects of Rosa gallica on the digestive enzymes. J. Nat. Med. 2010, 64, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Bellesia, A.; Verzelloni, E.; Taqliazucchi, D. Pomegranate ellagitannins inhibit α-glucosidase activity in vitro and reduce starch digestibility under simulated gastro-intestinal conditions. Int. J. Food Sci. Nutr. 2015, 66, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Zhang, W.; Feng, F.; Zhang, Y.; Kang, W. α-Glucosidase inhibitors isolated from medicinal plants. Food Sci. Hum. Wellness 2014, 3, 136–174. [Google Scholar] [CrossRef]
- McDougall, G.J.; Shpiro, F.; Dobson, P.; Smith, P.; Blake, A.; Stewart, D. Different polyphenolic components of soft fruits inhibit alpha-amylase and alpha-glucosidase. J. Agric. Food Chem. 2005, 53, 2760–2766. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Ni, X.; Kai, G.; Chen, X. A review on structure-activity relationship of dietary polyphenols inhibiting α-amylase. Crit. Rev. Food Sci. Nutr. 2013, 53, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Du, Y.-J.; Song, H.-C. α-Glucosidase and α-amylase inhibitory activities of guava leaves. Food Chem. 2010, 123, 6–13. [Google Scholar] [CrossRef]
- Li, Y.Q.; Zhou, F.C.; Gao, F.; Bian, J.S.; Shan, F. Comparative evaluation of quercetin, isoquercetin and rutin as inhibitors of α-glucosidase. J. Agric. Food Chem. 2009, 57, 11463–11468. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kwon, C.; Son, K. Inhibition of alpha-glucosidase and amylase by luteolin, a flavonoid. Biosci. Biotehnol. Biochem. 2000, 64, 2458–2461. [Google Scholar]
- Zhou, H.; Xing, J.; Liu, S.; Song, F.; Cai, Z.; Pi, Z.; Liua, Z.; Liua, S. Screening and determination for potential α-glucosidase inhibitors from leaves of Acanthopanax senticosus Harms by using UF-LC/MS and ESI-MSn. Phytochem. Anal. 2012, 24, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Toda, M.; Kawabata, J.; Kasai, T. Inhibitory effects of ellagi- and gallotannins on rat intestinal α-Glucosidase complexes. Biosci. Biotech. Biochem. 2001, 65, 542–547. [Google Scholar] [CrossRef]
- Spencer, C.M.; Cai, Y.; Martin, R.; Gaffney, S.H.; Goulding, P.N.; Mangnolato, D.; Lilley, Y.; Haslam, E. Polyphenol complexation—Some thoughts and observatios. Phytochemistry 1988, 27, 2397–2409. [Google Scholar] [CrossRef]
- Bai, N.; He, K.; Roller, M.; Zheng, B.; Chen, X.; Shao, Z.; Peng, T.; Zheng, Q. Active compounds from Lagerstroemia speciosa, insulin-like glucose uptake-stimulatory/inhibitory and adipocyte differentiation-inhibitory activities in 2T3-L1 cells. J. Agric. Food Chem. 2008, 56, 11668–11674. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Kim, J.K.; Li, Y.; Li, J.; Liu, F.; Chen, X. Tannic acid stimulates glucose transport and inhibits adipocyte differentiation in 3T3-L1 cells. J. Nutr. 2005, 135, 165–171. [Google Scholar] [PubMed]
- Broadhurst, C.L.; Polansky, M.M.; Anderson, R.A. Insulin-like biological activity of culinary and medicinal plant aqueous extracts in vitro. J. Agric. Food Chem. 2000, 48, 849–852. [Google Scholar] [CrossRef] [PubMed]
- Juśkiewicz, J.; Jurgonski, A.; Ko󠅲łodziejczyk, K.; Kosmala, M.; Milana, J.; Zduńczyk, Z.; Fotschki, B.; Zary-Sikorska, E. Blood glucose lowering efficacy of strawberry extracts rich in ellagitannins with different degree of polymerization in rats. Pol. J. Food Nutr. Sci. 2016, 66, 109–117. [Google Scholar] [CrossRef]
- Gomes, J.R.; Vedasiromoni, M.D.; Sharma, R.M.; Ganguly, D.K. Antihyperglycemic effect of black tea (Camellia sinensis) in rat. J. Ethnopharmacol. 2001, 27, 243–275. [Google Scholar]
- Mohamed, A.K.; Bierhaus, A.; Schiekofer, S.; Tritschler, H.; Ziegler, R.; Nawroth, P.P. The role of oxidative stress and NF-κB activation in late diabetic complications. BioFactors 1999, 10, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Weir, G.C.; Clore, E.T.; Zmachiroski, C.J.; Bonner-Weir, S. Islet secretion in a new experiment model for non-insulin dependent diabetes. Diabetes 1981, 5, 30–590. [Google Scholar]
- Saravanan, R.; Ramachandran, V. Effect of rebaudioside A, a diterpenoid on glucose homeostasis in STZ-induced diabetic rats. J. Physiol. Biochem. 2012, 68, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Lenzen, S. The mechanisms of alloxan- and streptozotocin-induced diabetes. Diabetologia 2008, 51, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Pareek, H.; Sharma, S.; Khajja, B.S.; Jain, K.; Jain, G.C. Evaluation of hypoglycemic and anti-hyperglycemic potential of Tridax procumbens (Linn.). BMC Complement. Altern. Med. 2009, 48. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, D.; Sharma, M.; Kumar, V.; Bajaj, H.K.; Verma, A. 2β-hydroxybetulinic acid 3β-caprylate: An active principle from Euryale ferox Salisb. seeds with antidiabetic, antioxidant, pancreas & hepatoprotective potential in streptozotocin induced diabetic rats. J. Food Sci. Technol. 2015, 52, 5427–5441. [Google Scholar] [PubMed]
- Gray, A.M.; Flatt, P.R. Actions of the traditional anti-diabetic plant, Agrimony eupatoria (agrimony): Effects on hyperglycaemia, cellular glucose metabolism and insulin secretion. Br. J. Nutr. 1998, 80, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Granica, S.; Krupa, K.; Kłębowska, A.; Kiss, A.K. Development and validation of HPLC-DAD-CAD-MS3 method for qualitative and quantitative standardization of polyphenols in Agrimoniae eupatoriae herba (Ph. Eur). J. Pharm. Biomed. Anal. 2013, 86, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N.; Kashchenko, N.I. Compotential profile and amylase inhibiting activity of phenolic compounds from Calendula officinalis L. leaves. Sci. World J. 2014. [Google Scholar] [CrossRef] [PubMed]
- Courteix, C.; Bardin, M.; Chantelauze, C.; Lavarenne, J.; Eschalier, A. Study of the sensitivity of the diabetes-induced pain model in rats to a range of analgesics. Pain 1994, 57, 153–160. [Google Scholar] [CrossRef]
- Stanley, A.; Mainzen, P.; Venugopal, M.P. Anti-oxidant action of Tinospora cordifolia root extract in alloxan diabetic rats. Phytother. Res. 2001, 15, 213–218. [Google Scholar]
- Gupta, S.; Kataria, M.; Gupta, P.K.; Murganandan, S.; Yashroy, R.C. Protective role of extracts of neem seeds in diabetes caused by streptozotocin in rats. J. Ethnopharmacol. 2004, 90, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Serra-Barcellona, C.; Coll Aráoz, M.V.; Cabrera, W.M.; Habib, N.C.; Honoré, S.M.; Catalán, C.A.N.; Grau, A.; Genta, S.B.; Sánchez, S.S. Smallanthus macroscyphus: A new source of antidiabetic compounds. Chem. Biol. Interact. 2014, 209, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the Comarum palustre plant material and extracts are available from the authors.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Part | Decoction | Infusion | Tincture | 30% Ethanol Extract | 60% Ethanol Extract | 96% Ethanol Extract |
|---|---|---|---|---|---|---|
| Herb | 127.4 ± 5.3 iii | 142.9 ± 5.8 iii | 98.4 ± 3.7 ii | 89.4 ± 3.3 i,ii | 52.0 ± 1.7 i | 154.7 ± 6.0 iii,iv |
| Roots | >300 | >300 | >300 | >300 | 272.7 ± 10.6 v | >300 |
| Flowers | >300 | >300 | 254.9 ± 9.9 iv,v | 183.3 ± 7.1 iv | 104.2 ± 4.3 iii | >300 |
| Seeds | >300 | >300 | >300 | >300 | >300 | >300 |
| Peak No. | Compound | tR,min | λmax,nm | ESI-MS, m/z | Method a |
|---|---|---|---|---|---|
| 1 | 2-Pyrone-4,6-dicarboxylic acid | 0.75 | 212, 316 | 138 [M − H]− | i, ii, iii |
| 2 | Gallic acid | 0.92 | 220, 270 | 169 [M − H]− | i*, ii*, iii* |
| 3 | α-Pedunculagin | 2.53 | 235 | 783 [M − H]− | i, ii, iii |
| 4 | β-Pedunculagin | 3.31 | 235 | 783 [M − H]− | i, ii, iii |
| 5 | Procyanidin B3 | 3.80 | 240, 278 | 577 [M − H]− | i*, ii*, iii* |
| 6 | (+)-Catechin | 4.15 | 240, 278 | 289 [M − H]− | i*, ii*, iii* |
| 7 | Potentillin | 6.21 | 220, 256 | 935 [M − H]− | i, ii, iii |
| 8 | Agrimoniin | 6.98 | 228, 270 | 1869 [M − H]−, 934 [M − 2H]2− | i*, ii*, iii* |
| 9 | Ellagic acid | 7.20 | 250, 367 | 301 [M − H]−, 603 [2M − H]− | i*, ii*, iii* |
| 10 | Rutin | 7.33 | 258, 356 | 609 [M − H]−, 301 [M − H − Rut]− | i*, ii*, iii* |
| 11 | Miquelianin | 7.50 | 254, 355 | 477 [M − H]−, 301 [M − H − GlcA]− | i*, ii*, iii* |
| 12 | Isoquercitrin | 7.72 | 254, 356 | 463 [M − H]−, 301 [M − H − Glc]− | i*, ii*, iii* |
| 13 | Nicotiflorin | 8.12 | 364, 350 | 593 [M − H]−, 285 [M − H − Rut]− | i*, ii*, iii* |
| 14 | Astragalin | 8.55 | 265, 350 | 447 [M − H]−, 285 [M − H − Glc]− | i*, ii*, iii* |
| 15 | Afzelin | 9.24 | 265, 351 | 431 [M − H]−, 285 [M − H − Rha]− | i*, ii*, iii* |
| HPLC Peak No. | Compound | Extract A, mg·g−1 a | Extract B, mg·g−1 a |
|---|---|---|---|
| 1 | 2-Pyrone-4,6-dicarboxylic acid | 3.25 ± 0.06 | 4.52 ± 0.11 |
| 2 | Gallic acid | 0.50 ± 0.01 | n.d. |
| 3 | α-Pedunculagin | 9.80 ± 0.25 | n.d. |
| 4 | β-Pedunculagin | 10.79 ± 0.33 | n.d. |
| 5 | Procyanidin B3 | 30.02 ± 0.78 | 0.52 ± 0.01 |
| 6 | (+)-Catechin | 28.02 ± 0.67 | n.d. |
| 7 | Potentillin | 22.82 ± 0.66 | n.d. |
| 8 | Agrimoniin | 240.94 ± 6.74 | 1.03 ± 0.02 |
| 9 | Ellagic acid | 6.67 ± 0.18 | n.d. |
| 10 | Rutin | 8.07 ± 0.21 | 12.20 ± 0.32 |
| 11 | Miquelianin | 80.81 ± 2.10 | 120.27 ± 3.72 |
| 12 | Isoquercitrin | 8.67 ± 0.25 | 12.95 ± 0.33 |
| 13 | Nicotiflorin | 6.45 ± 0.18 | 9.82 ± 0.26 |
| 14 | Astragalin | 25.40 ± 0.71 | 32.11 ± 0.99 |
| 15 | Afzelin | 6.35 ± 0.17 | 9.64 ± 0.26 |
| Total ellagitannins (Σ3,4,7,8) | 284.35 | 1.03 | |
| Total flavonoids (Σ10–15) | 135.75 | 196.99 | |
| Total catechines (Σ5,6) | 58.04 | 0.52 | |
| Other classes (Σ1,2,9) | 10.42 | 4.52 | |
| Total phenolics (Σ1–15) | 488.56 | 203.06 | |
| Experimental group | Body Weight, g | Insulin, U/L | Hb, mg/dL | HbA1c, %Hb | |
|---|---|---|---|---|---|
| 0 Day | 21 Day | ||||
| Control | 195 ± 6 | 232 ± 7 | 15.90 ± 1.23 | 14.02 ± 0.84 | 4.82 ± 0.28 |
| STZ + saline | 198 ± 5 | 121 ± 6 a | 6.97 ± 0.41 a | 7.11 ± 0.43 a | 12.93 ± 0.77 a |
| STZ + Extract A (100 mg/kg) | 192 ± 4 | 149 ± 5 a,c | 7.35 ± 0.44 a | 8.39 ± 0.41 a,c | 12.52 ± 0.62 a |
| STZ + Extract A (200 mg/kg) | 192 ± 4 | 151 ± 5 a,c | 9.14 ± 0.59 a,d | 9.63 ± 0.48 a,c | 10.83 ± 0.54 a,c |
| STZ + Extract A (400 mg/kg) | 199 ± 6 | 197 ± 7 b,d | 14.05 ± 0.91 d | 12.92 ± 0.64 d | 7.22 ± 0.36 b,d |
| STZ + Extract B (100 mg/kg) | 190 ± 3 | 120 ± 3 a | 6.90 ± 0.40 a | 7.32 ± 0.36 a | 12.70 ± 0.60 a |
| STZ + Extract B (200 mg/kg) | 192 ± 4 | 127 ± 5 a | 7.04 ± 0.47 a | 7.55 ± 0.39 a | 12.34 ± 0.58 a |
| STZ + Extract B (400 mg/kg) | 195 ± 4 | 138 ± 4 a,c | 7.37 ± 0.42 a | 8.39 ± 0.41 a | 11.25 ± 0.56 a |
| STZ + Agrimoniin (25 mg/kg) | 193 ± 5 | 183 ± 3 b,d | 10.35 ± 0.70 ad | 8.56 ± 0.40 a,d | 11.38 ± 0.56 a |
| STZ + Agrimoniin (50 mg/kg) | 198 ± 7 | 202 ± 5 b,d | 14.10 ± 0.91 d | 10.81 ± 0.52 b,d | 9.06 ± 0.42 b,c |
| STZ + Agrimoniin (100 mg/kg) | 195 ± 3 | 219 ± 7 d | 15.22 ± 1.06 d | 13.26 ± 0.66 d | 6.14 ± 0.30 b |
| STZ + Insulin | 192 ± 4 | 228 ± 8 d | 15.63 ± 0.92 d | 13.73 ± 0.68 d | 5.11 ± 0.25 d |
| Extract A (400 mg/kg) | 195 ± 5 | 236 ± 7 d | 15.97 ± 1.11 d | 14.19 ± 0.73 d | 4.93 ± 0.23 d |
| Extract B (400 mg/kg) | 192 ± 5 | 243 ± 9 d | 15.86 ± 1.02 d | 14.11 ± 0.70 d | 4.87 ± 0.24 d |
| Agrimoniin (100 mg/kg) | 194 ± 4 | 230 ± 8 d | 15.90 ± 1.09 d | 14.06 ± 0.71 d | 4.80 ± 0.20 d |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kashchenko, N.I.; Chirikova, N.K.; Olennikov, D.N. Agrimoniin, an Active Ellagitannin from Comarum palustre Herb with Anti-α-Glucosidase and Antidiabetic Potential in Streptozotocin-Induced Diabetic Rats. Molecules 2017, 22, 73. https://doi.org/10.3390/molecules22010073
Kashchenko NI, Chirikova NK, Olennikov DN. Agrimoniin, an Active Ellagitannin from Comarum palustre Herb with Anti-α-Glucosidase and Antidiabetic Potential in Streptozotocin-Induced Diabetic Rats. Molecules. 2017; 22(1):73. https://doi.org/10.3390/molecules22010073
Chicago/Turabian StyleKashchenko, Nina I., Nadezhda K. Chirikova, and Daniil N. Olennikov. 2017. "Agrimoniin, an Active Ellagitannin from Comarum palustre Herb with Anti-α-Glucosidase and Antidiabetic Potential in Streptozotocin-Induced Diabetic Rats" Molecules 22, no. 1: 73. https://doi.org/10.3390/molecules22010073
APA StyleKashchenko, N. I., Chirikova, N. K., & Olennikov, D. N. (2017). Agrimoniin, an Active Ellagitannin from Comarum palustre Herb with Anti-α-Glucosidase and Antidiabetic Potential in Streptozotocin-Induced Diabetic Rats. Molecules, 22(1), 73. https://doi.org/10.3390/molecules22010073