
Insoluble-Bound Phenolics in Food



Abstract
:1. Introduction
2. Results
2.1. Localization of Soluble and Insoluble-Bound Phenolics
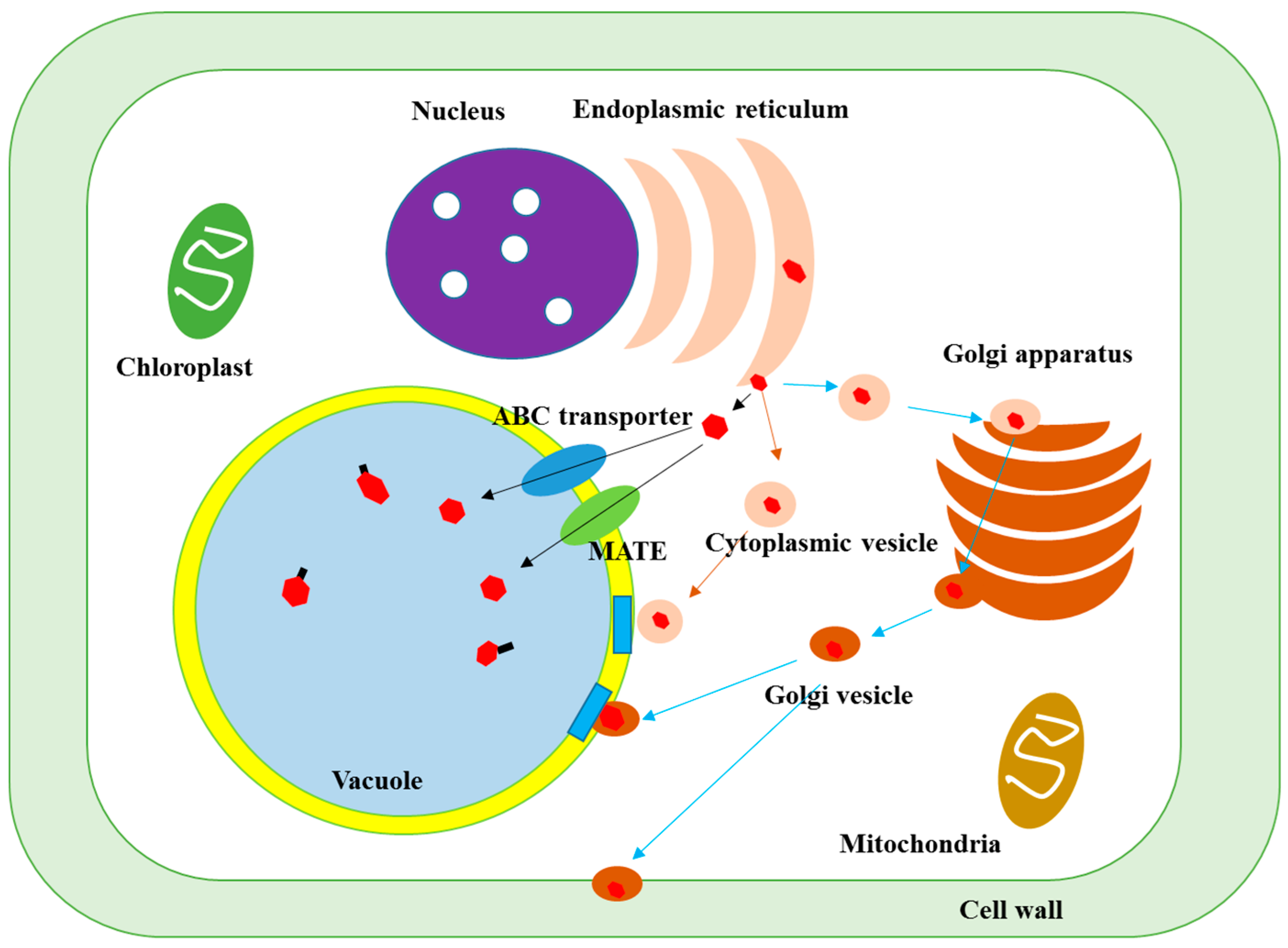
2.2. Synthesis, Transport and Formation of Insoluble-Bound Phenolics
2.2.1. Synthesis
2.2.2. Transport
2.2.3. Formation of Insoluble-Bound Phenolics in the Cell Walls
2.3. Phenolic Acids and Flavonoids Contents in Cereals, Legumes and Other Seeds
2.3.1. Phenolic Acids
2.3.2. Flavonoids
2.4. Metabolism of Insoluble-Bound Phenolics in Human Digestive Tract
2.5. Bioactivities of Insoluble-Bound Phenolics
2.5.1. Phenolic Acids
2.5.2. Flavonoids
2.6. Extraction of Insoluble-Bound Phenolics
2.6.1. Acid Hydrolysis
2.6.2. Alkaline Hydrolysis
2.6.3. Enzymatic Hydrolysis
2.7. Release of Insoluble-Bound Phenolics from Cell Wall Matrix
2.7.1. Non-Thermal Processing
Fermentation Process in Food
Germination
2.7.2. Thermal and Hydrothermal Processing
Roasting
Extrusion Cooking
Boiling
3. Conclusions
Acknowledgments
Conflicts of Interest
References
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Stafford, H.A. Flavonoid evolution: An enzymic approach. Plant Physiol. 1991, 96, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Mathesius, U. Flavonoids induced in cells undergoing nodule organogenesis in white clover are regulators of auxin breakdown by peroxidase. J. Exp. Bot. 2001, 52, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Nayak, B.; Liu, R.H.; Tang, J. Effect of processing on phenolic antioxidants of fruits, vegetables and grains: A review. Crit. Rev. Food Sci. Nutr. 2015, 55, 887–918. [Google Scholar] [CrossRef] [PubMed]
- Martinoia, E.; Ratajczak, R. Transport of organic molecules across the tonoplast. In The Plant Vacuole; Leigh, R.A., Sanders, D., Eds.; Academic Press: Cambridge, MA, USA, 1997; pp. 388–390. [Google Scholar]
- Meyer, D.; Pajonk, S.; Micali, C.; O’Connell, R.; Schulze-Lefert, P. Extracellular transport and integration of plant secretory proteins into pathogen-induced cell wall compartments. Plant J. 2009, 57, 986–999. [Google Scholar] [CrossRef] [PubMed]
- Bradford, K.J.; Bewley, J.D. Seeds: Biology, technology and role in agriculture. In Plants, Genes and Crop Biotechnology; Chrispeels, M.J., Sadava, D.E., Eds.; Jones and Bartlett Publishers: Burlington, MA, USA, 2003; pp. 214–217. [Google Scholar]
- Jung, H.J.G.; Samac, D.A.; Sarath, G. A review: Modifying crops to increase cell wall digestibility. Plant Sci. 2012, 185, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S. Transport of flavonoids: From cytosolic synthesis to vacuolar accumulation. In Science of Flavonoids; Grotewold, E., Ed.; Springer: Berlin, Germany, 2006; pp. 123–146. [Google Scholar]
- Grotewold, E. Subcellular trafficking of phytochemicals. Recent Res. Develop. Plant Physiol. 2001, 2, 31–48. [Google Scholar]
- Grotewold, E. The challenges of moving chemicals within and out of cells: Insights into the transport of plant natural products. Planta 2004, 219, 906–909. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Dixon, R.A. The ‘ins’ and ‘outs’ of flavonoid transport. Trends Plant Sci. 2009, 15, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance: A review. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Yazaki, K. Transporters of secondary metabolites. Curr. Opin. Plant Biol. 2005, 8, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Strack, D.; Heilemann, J.; Mömken, M.; Wray, V. Cell wall-conjugated phenolics from Coniferae leaves. Phytochemistry 1988, 27, 3517–3521. [Google Scholar] [CrossRef]
- Markham, K.R.; Ryan, K.G.; Gould, K.S.; Richards, G.K. Cell wall sited flavonoids in lisianthus flower petals. Phytochemistry 2000, 54, 681–687. [Google Scholar] [CrossRef]
- McLusky, S.R.; Bennet, M.H.; Beale, M.H.; Lewis, M.J.; Gaskin, P.; Mansfield, J.W. Cell wall alterations and localized accumulation of feruoyl-3’-methoxytyramine in onion epidermis at sites of attempted penetration of Botrytis allii are associated with actin polarization, peroxidase activity and suppression of flavonoid biosynthesis. Plant J. 1999, 17, 523–534. [Google Scholar] [CrossRef]
- Nicholson, R.L.; Hammerschmidt, R. Phenolic compounds and their role in disease resistance. Ann. Rev. Phytopathol. 1992, 30, 369–389. [Google Scholar] [CrossRef]
- Jansen, M.A.; van den Noort, R.E.; Tan, M.Y.; Prinsen, E.; Lagrimini, L.M.; Thorneley, R.N. Phenol-oxidizing peroxidases contribute to the protection of plants from ultraviolet radiation stress. Plant Physiol. 2001, 126, 1012–1023. [Google Scholar] [CrossRef] [PubMed]
- Alves, G.H.; Ferreira, C.D.; Vivian, P.G.; Monks, J.L.F.; Elias, M.C.; Vanier, N.L.; De Oliveira, M. The revisited levels of free and bound phenolics in rice: Effects of the extraction procedure. Food Chem. 2016, 208, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Das, A.K.; Singh, V. Antioxidative free and bound phenolic constituents in botanical fractions of Indian specialty maize (Zea mays L.) genotypes. Food Chem. 2016, 201, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Irakli, M.N.; Samanidou, V.F.; Biliaderis, C.G.; Papadoyannis, I.N. Development and validation of an HPLC-method for determination of free and bound phenolic acids in cereals after solid-phase extraction. Food Chem. 2012, 134, 1624–1632. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekara, A.; Shahidi, F. Content of insoluble bound phenolics in millets and their contribution to antioxidant capacity. J. Agric. Food Chem. 2010, 58, 6706–6714. [Google Scholar] [CrossRef] [PubMed]
- Alshikh, N.; de Camargo, A.C.; Shahidi, F. Phenolics of selected lentil cultivars: Antioxidant activities and inhibition of low-density lipoprotein and DNA damage. J. Funct. Foods. 2015, 18, 1022–1038. [Google Scholar] [CrossRef]
- Chen, P.X.; Tang, Y.; Marcone, M.F.; Pauls, P.K.; Zhang, B.; Liu, R.; Tsao, R. Characterization of free, conjugated and bound phenolics and lipophilic antioxidants in regular- and non-darkening cranberry beans (Phaseolus vulgaris L.). Food Chem. 2015, 185, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Verardo, V.; Riciputi, Y.; Garrido-Frenich, A.; Caboni, M.F. Short communication: Determination of free and bound phenolic compounds in soy isoflavone concentrate using a PFP fused core column. Food Chem. 2015, 185, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Pajak, P.; Socha, R.; Gałkowska, D.; Roznowski, J.; Fortuna, T. Phenolic profile and antioxidant activity in selected seeds and sprouts. Food Chem. 2014, 143, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Ross, K.A.; Beta, T.; Arntfield, S.D. A comparative study on the phenolic acids identified and quantified in dry beans using HPLC as affected by different extraction and hydrolysis methods. Food Chem. 2009, 113, 336–344. [Google Scholar] [CrossRef]
- Gutiérrez-Uribe, J.A.; Rojas-García, C.; García-Lara, S.; Serna-Saldivar, S.O. Phytochemical analysis of wastewater (nejayote) obtained after lime-cooking of different types of maize kernels processed into masa for tortillas. J. Cereal Sci. 2010, 52, 410–416. [Google Scholar] [CrossRef]
- Wang, Y.K.; Zhang, X.; Chen, G.L.; Yu, J.; Yang, L.Q.; Gao, Y.Q. Antioxidant property and their free, soluble conjugate and insoluble-bound phenolic contents in selected beans. J. Funct. Food. 2016, 24, 359–372. [Google Scholar] [CrossRef]
- Naczk, M.; Shahidi, F. The effect of methanol-ammonia-water treatment on the content of phenolic acids of canola. Food Chem. 1989, 31, 159–164. [Google Scholar] [CrossRef]
- Beejmohun, V.; Fliniaux, O.; Grand, E.; Lamblin, F.; Bensaddek, L.; Christen, P. Microwave-assisted extraction of the main phenolic compounds in flaxseed. Phytochem. Anal. 2007, 18, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.S.G.; Negi, P.S.; Radha, C. Phenolic composition, antioxidant and antimicrobial activities of free and bound phenolic extracts of Moringa oleifera seed flour. J. Funct. Foods. 2013, 5, 1883–1891. [Google Scholar] [CrossRef]
- Min, B.; Gu, L.; McClung, A.M.; Bergman, C.J.; Chen, M.H. Free and bound total phenolic concentrations, antioxidant capacities and profiles of proanthocyanidins and anthocyanins in whole grain rice (Oryza sativa L.) of different bran colours. Food Chem. 2012, 133, 715–722. [Google Scholar] [CrossRef]
- Ayoub, M.; de Camargo, A.C.; Shahidi, F. Antioxidants and bioactivities of free, esterified and insoluble-bound phenolics from berry seed meals. Food Chem. 2016, 197, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhang, B.; Li, X.; Chen, P.X.; Zhang, H.; Liu, R.; Tsao, R. Bound phenolics of quinoa seeds released by acid, alkaline and enzymatic treatments and their antioxidant and α-glucosidase and pancreatic lipase inhibitory effects. J. Agric. Food Chem. 2016, 64, 1712–1719. [Google Scholar] [CrossRef] [PubMed]
- Das, A.K.; Singh, V. Antioxidative free and bound phenolic constituents in pericarp, germ and endosperm of Indian dent (Zea mays var. indentata) and flint (Zea mays var. indurata) maize. J. Funct. Foods 2015, 13, 363–374. [Google Scholar] [CrossRef]
- Podsedek, A.; Redzynia, M.; Klewicka, E.; Koziołkiewicz, M. Matrix effects on the stability and antioxidant activity of red cabbage anthocyanins under simulated gastrointestinal digestion. Biomed. Res. Int. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073–2085. [Google Scholar]
- Rodríguez-Roque, M.J.; Rojas-Graü, M.A.; Elez-Martínez, P.; Martín-Belloso, O. In vitro bioaccessibility of health-related compounds as affected by the formulation of fruit juice- and milk-based beverages. Food Res. Int. 2014, 62, 771–778. [Google Scholar] [CrossRef]
- Adom, K.K.; Liu, R.H. Antioxidant activity of grains. J. Agric. Food Chem. 2002, 50, 6182–6187. [Google Scholar] [CrossRef] [PubMed]
- Andreasen, M.F.; Kroon, P.A.; Williamson, G.; Garcia-Conesa, M.T. Intestinal release and uptake of phenolic antioxidant diferulic acids. Free Rad. Bio. Med. 2001, 31, 304–314. [Google Scholar] [CrossRef]
- Gibson, G.; Roberfroid, M. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [PubMed]
- Yi, W.; Fisher, J.; Krewer, G.; Akoh, C.C. Phenolic compounds from blueberries can inhibit colon cancer cell proliferation and induce apoptosis. J. Agric. Food Chem. 2005, 53, 7320–7329. [Google Scholar] [CrossRef] [PubMed]
- Yeh, R.D.; Chen, J.C.; Lai, T.Y.; Yang, J.S.; Yu, C.S.; Chiang, J.H.; Lu, C.C.; Yang, S.T.; Yu, C.C.; Chang, S.J.; et al. Gallic acid induces G(0)/G(1) phase arrest and apoptosis in humanleukemia HL-60 cells through inhibiting cyclin D and E and activating mitochondria-dependent pathway. Anticancer Res. 2011, 31, 2821–2832. [Google Scholar] [PubMed]
- Inoue, M.; Suzuki, R.; Sakaguchi, N.; Li, Z.; Takeda, T.; Ogihara, Y.; Jiang, B.Y.; Chen, Y. Selective induction of cell death in cancer cells by gallic acid. Biol. Pharm. Bull. 1995, 18, 1526–1530. [Google Scholar] [CrossRef] [PubMed]
- Kaur, M.; Velmurugan, B.; Rajamanickam, S.; Agarwal, R.; Agar-Wal, C. Gallic acid, an active constituent of grape seed extract, exhibits anti-proliferative, pro-apoptotic and anti-tumorigenic effects against prostate carcinoma xenograft growth in nude mice. Pharm. Res. 2009, 26, 2133–2140. [Google Scholar] [CrossRef] [PubMed]
- Veluri, R.; Singh, R.P.; Liu, Z.; Thompson, J.A.; Agarwal, R.; Agarwal, C. Fractionation of grape seed extract and identification of gallic acid as one of the major active constituents causing growth inhibition and apoptotic death of DU145 human prostate carcinoma cells. Carcinogenesis 2006, 27, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Kamatham, S.; Kumar, N.; Gudipalli, P. Isolation and characterization of gallic acid and methylgallate from the seed coats of Givotia rottleriformis Griff. and their anti-proliferative effect on human epidermoid carcinoma A431 cells. Toxicol. Rep. 2015, 2, 520–529. [Google Scholar] [CrossRef]
- Hsu, W.C.; Chang, S.P.; Lin, L.C.; Li, C.L.; Richardson, C.D.; Lin, C.C.; Lin, L.T. Limonium sinense and gallic acid suppress hepatitis C virus infection by blocking early viral entry. Antiviral Res. 2015, 118, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Morgan, R.L.; Baack, B.; Smith, B.D.; Yartel, A.; Pitasi, M.; Falck-Ytter, Y. Eradication of hepatitis C virus infection and the development of hepatocellular carcinoma: a meta-analysis of observational studies. Ann. Intern. Med. 2013, 158, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.G.; Kang, O.H.; Lee, Y.S.; Oh, Y.C.; Chae, H.S.; Jang, H.J.; Kim, J.H.; Sohn, D.H.; Shin, D.W.; Park, H.; et al. In vitro activity of methyl gallate isolated from Galla rhois alone and in combination with ciprofloxacin against clinical isolates of salmonella. J. Microbiol. Biotechnol. 2008, 18, 1848–1852. [Google Scholar] [PubMed]
- Kane, C.J.; Menna, J.H.; Sung, C.C.; Yeh, Y.C. Methyl gallate, methyl-3,4,5-trihydoxybenzoate, is a potent and highly specific inhibitor of herpes simplex virus in vitro. II. Antiviral activity of methyl gallate and its derivatives. Biosci. Rep. 1988, 8, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.S.; Oh, J.S.; Kang, I.C.; Hong, S.J.; Choi, C.H. Inhibitory effect of methyl gallate and gallic acid on oral bacteria. J. Microbiol. 2008, 46, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.Z.; Zhao, G.R.; Yang, J.; Yuan, Y.J.; Zhu, G.G.; Hiltunen, R. Protective effect of Ligusticum chuanxiong and Angelica sinensis on endothelial cell damage induced by hydrogen peroxide. Life Sci. 2004, 75, 1775–1786. [Google Scholar] [CrossRef] [PubMed]
- Fahrioğlu, U.; Dodurga, Y.; Elmas, L.; Seçme, M. Ferulic acid decreases cell viability and colony formation while inhibiting migration of MIA PaCa-2 human pancreatic cancer cells in vitro. Gene. 2016, 576, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, A.; Chinnaiyan, M.; Karundevi, B. Ferulic acid regulates hepatic GLUT2 gene expression in high fat and fructose-induced type-2 diabetic adult male rat. Eur. J. Pharmacol. 2015, 761, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Eitsuka, T.; Tatewaki, N.; Nishida, H.; Kurata, T.; Nakagawa, K.; Miyazawa, T. Synergistic inhibition of cancer cell proliferation with a combination of delta-tocotrienol and ferulic acid. Biochem. Biophys. Res. Commun. 2014, 453, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, C.; Santangelo, R. Ferulic acid: Pharmacological and toxicological aspects. Food Chem Toxicol. 2014, 65, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.E.; Park, E. Ferulic acid in combination with PARP inhibitor sensitizes breast cancer cells as chemotherapeutic strategy. Biochem. Biophys. Res. Commun. 2015, 458, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Janicke, B.; Hegardt, C.; Krogh, M.; Onning, G.; Akesson, B.; Cirenajwis, H.M.; Oredsson, S.M. The antiproliferative effect of dietary fiber phenolic compounds ferulic acid and p-coumaric acid on the cell cycle of Caco-2 cells. Nutr. Cancer. 2011, 63, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.Y.; Wang, H.M.; Chang, K.F.; Hu, H.T.; Hwang, L.J.; Fu, T.F.; Chen, B.H. Feruloyl-L-arabinose attenuates migration, invasion and production of reactive oxygen species in H1299 lung cancer cells. Food Chem. Toxicol. 2013, 58, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Bouzaiene, N.N.; Jaziri, S.K.; Kovacic, H.; Chekir-Ghedira, L.; Ghedira, K.; Luis, J. The effects of caffeic, coumaric and ferulic acids on proliferation, superoxide production, adhesion and migration of human tumor cells in vitro. Eur. J. Pharmacol. 2015, 766, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.C.; Kuo, Y.C.; Chou, C.J. Anti-herpes simplex virus type-1 flavonoids and a new flavanone from the root of Limonium sinense. Planta Med. 2000, 66, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Panat, N.A.; Singh, B.G.; Maurya, D.K.; Sandur, S.K.; Ghaskadbi, S.S. Troxerutin, a natural flavonoid binds to DNA minor groove and enhances cancer cell killing in response to radiation. Chem. Biol. Interact. 2016, 251, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Hirayama, R.; Kubota, N. The chemopreventive flavonoid apigenin confers radiosensitizing effect in human tumor cells grown as monolayers and spheroids. J. Radiat. Res. 2007, 48, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Kim, E.; Kim, W.; Seong, K.M.; Youn, H. Rhamnetin and cirsiliol induce radiosensitization and inhibition of epithelial-mesenchymal transition (EMT) by miR-34a-mediated suppression of Notch-1 expression in non-small cell lung cancer cell lines. J. Biol. Chem. 2013, 288, 27343–27357. [Google Scholar] [CrossRef] [PubMed]
- Chtourou, Y.; Aouey, B.; Aroui, S.; Kebieche, M.; Fetoui, H. Anti-apoptotic and anti-inflammatory effects of naringin on cisplatin-induced renal injury in the rat. Chem. Biol. Interact. 2016, 243, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ahn, T.G.; Kim, H.K.; Park, S.W.; Kim, S.A.; Lee, B.R.; Han, S.J. Protective effects of green tea polyphenol against cisplatin-induced nephrotoxicity in rats. Obstet. Gynecol. Sci. 2014, 57, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Hamad, R.; Jayakumar, C.; Ranganathan, P.; Mohamed, R.; El-Hamamy, M.M.; Dessouki, A.A.; Ibrahim, A.; Ramesh, G. Honey feeding protects kidney against cisplatin nephrotoxicity through suppression of inflammation. Clin. Exp. Pharmacol. Physiol. 2015, 42, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Kamel, K.M.; El-Raouf, A.; Ola, M.; Metwally, S.A.; El-Latif, A.; Hekma, A.; Elsayed, M.E. Hesperidin and rutin, antioxidant citrus flavonoids, attenuate cisplatin-induced nephrotoxicity in rats. J. Biochem. Mol. Toxic. 2014, 28, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Ju, S.M.; Kang, J.G.; Bae, J.S.; Pae, H.O.; Lyu, Y.S.; Jeon, B.H. The flavonoid apigenin ameliorates cisplatin-induced nephrotoxicity through reduction of p53 activation and promotion of PI3K/Akt pathway in human renal proximal tubular epithelial cells. Evid. Based Complement. Altern. Med. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Simkhada, D.; Kim, E.; Lee, H.C.; Sohng, J.K. Metabolic engineering of Escherichia coli for the biological synthesis of 7-O-xylosyl naringenin. Mol. Cells. 2009, 28, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Larson, A.J.; Symons, J.D.; Jalili, T. Therapeutic potential of quercetin to decrease blood pressure: review of efficacy and mechanisms. Adv. Nutr. 2012, 3, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Cherniack, E.P. A berry thought-provoking idea: the potential role of plant polyphenols in the treatment of age-related cognitive disorders. Br. J. Nutr. 2012, 108, 794–800. [Google Scholar] [CrossRef] [PubMed]
- Leung, H.W.C.; Lin, C.J.; Hour, M.J.; Yang, W.H.; Wang, M.Y.; Lee, H.Z. Kaempferol induces apoptosis in human lung non-small carcinoma cells accompanied by an induction of antioxidant enzymes. Food Chem. Toxicol. 2007, 45, 2005–2013. [Google Scholar] [CrossRef] [PubMed]
- Bestwick, C.S.; Milne, L.; Duthie, S.J. Kaempferol induced inhibition of HL-60 cell growth results from a heterogeneous response, dominated by cell cycle alterations. Chem. Biol. Interact. 2007, 170, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Chen, L.; Ma, X.; Jiao, R.; Li, X.; Wang, Y. Protective effects of kaempferol against reactive oxygen species induced hemolysis and its antiproliferative activity on human cancer cells. Eur. J. Med. Chem. 2016, 114, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Calderon-Montano, J.M.; Burgos-Moron, E.; Perez-Guerrero, C.; Lopez-Lazaro, M. A review on the dietary flavonoid kaempferol. Mini Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef] [PubMed]
- Hamalainen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. Anti-inflammatory effects of flavonoids: genistein, kaempferol, quercetin and daidzein inhibit STAT-1 and NF-kB activations, whereas flavone, isorhamnetin, naringenin and pelargonidin inhibit only NF-k B activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Mediat. Inflamm. 2007, 2007, 1–10. [Google Scholar]
- Toyoda, M.; Tanaka, K.; Hoshino, K.; Akiyama, H.; Tanimura, A.; Saito, Y. Profiles of potentially antiallergic flavonoids in 27 kinds of health tea and green tea infusions. J. Agric. Food Chem. 1997, 45, 2561–2564. [Google Scholar] [CrossRef]
- Trivedi, R.; Kumar, S.; Kumar, A.; Siddiqui, J.A.; Swarnkar, G.; Gupta, V.; Kendurker, A.; Dwivedi, A.K.; Romero, J.R.; Chattopadhyay, N. Kaempferol has osteogenic effect in ovariectomized adult Sprague Dawley rats. Mol. Cell. Endocrinol. 2008, 289, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Harnly, J.M.; Doherty, R.F.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Bhagwat, S. Flavonoid content of U.S. fruits, vegetables and nuts. J. Agric. Food Chem. 2006, 54, 9966–9977. [Google Scholar] [CrossRef] [PubMed]
- Nirmala, P.; Ramanathan, M. Effect of myricetin on 1,2 dimethylhydrazine induced rat colon carcinogenesis. J. Exp. Ther. Oncol. 2011, 9, 101–108. [Google Scholar] [PubMed]
- Orjala, J.; Wright, A.D.; Behrends, H.; Folkers, G.; Sticher, O.; Rüegger, H. Cytotoxic and antibacterial dihydrochalcones from Piper aduncum. J. Nat.Prod. 1994, 57, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Tamogami, S.; Kodama, O. Coronatine elicits phytoalexin production in rice leaves (Oryza sativa L.) in the same manner as jasmonic acid. Phytochemistry 2000, 54, 689–694. [Google Scholar] [CrossRef]
- Sani, I.M.; Iqbal, S.; Chan, K.W.; Ismail, M. Effect of acid and base catalyzed hydrolysis on the yield of phenolics and antioxidant activity of extracts from germinated brown rice (GBR). Molecules 2012, 17, 7584–7594. [Google Scholar] [CrossRef] [PubMed]
- Moussa-Ayoub, T.E.; El-Samahy, S.K.; Kroh, L.W.; Rohn, S. Identification and quantification of flavonol aglycons in cactus pear (Opuntia ficus indica) fruit using a commercial pectinase and cellulase preparation. Food Chem. 2011, 124, 1177–1184. [Google Scholar] [CrossRef]
- Fazary, A.E.; Ju, Y.H. Feruloyl esterases as biotechnological tools: Current and future perspectives. Acta Biochim. Biophys. Sinica 2007, 39, 811–828. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Krygier, K.; Sosulski, F.; Hogge, L. Free, esterified and insoluble-bound phenolic acids: Extraction and purification procedure. J. Agric. Food Chem. 1982, 30, 330–334. [Google Scholar] [CrossRef]
- Landbo, A.K.; Meyer, A.S. Enzyme-assisted extraction of antioxidative phenols from black currant juice press residues (Ribes nigrum). J. Agric. Food Chem. 2001, 49, 3169–3177. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Yagiz, Y.; Borejsza-Wysocki, W.; Lu, J.; Gu, L.; Ramírez-Rodrigues, M.M.; Marshall, M.R. Enzyme release of phenolics from muscadine grape (Vitis rotundifolia Michx.) skins and seeds. Food Chem. 2014, 157, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Hwang, I.W.; Chung, S.K. Enhancing polyphenol extraction from unripe apples by carbohydrate-hydrolyzing enzymes. J. Zhejiang Univer. Sci B. 2009, 10, 912–919. [Google Scholar] [CrossRef] [PubMed]
- Hur, S.J.; Lee, S.Y.; Kim, Y.C.; Choi, I.W.; Kim, G.B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cheng, J. Hydrolysis of lignocellulosic materials for ethanol production: A review. Bioresource Technol. 2002, 82, 1–11. [Google Scholar] [CrossRef]
- Tian, Y.; Jiang, Y.; Ou, S. Interaction of cellulase with three phenolic acids. Food Chem. 2013, 138, 1022–1027. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.; Kroon, P.A.; Faulds, C.B. Hairy plant polysaccharides: a close shave with microbial esterases. Microbiol. 1998, 144, 2011–2023. [Google Scholar] [CrossRef] [PubMed]
- Crepin, V.F.; Faulds, C.B.; Connerton, I.F. Functional classification of the microbial feruloyl esterases. Appl. Microbiol. Biotechnol. 2004, 63, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Tapin, S.; Sigoillot, J.C.; Asther, M.; Petit-Conil, M. Feruloyl esterase utilization for simultaneous processing of non-wood plants into phenolic compounds and pulp fibers. J. Agric. Food Chem. 2006, 54, 3697–3703. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Park, K.S.; Kim, M.J.; Kim, S.Y. Free phenolic contents and their antioxidant activities of fresh and fermented rice spent water. Food Sci. Biotechnol. 2010, 19, 1415–1420. [Google Scholar] [CrossRef]
- Dey, T.B.; Kuhad, R.C. Upgrading the antioxidant potential of cereals by their fungal fermentation under solid-state cultivation conditions. Lett. Appl. Microbiol. 2014, 59, 493–499. [Google Scholar]
- Nonogaki, H.; Basselb, G.W.; Bewley, J.D. Germination—still a mystery. Plant Sci. 2010, 179, 574–581. [Google Scholar] [CrossRef]
- Raven, P.H.; Evert, R.F.; Eichhorn, S.E. Biology of Plants, 7th ed.; Freeman and Company Publishers: New York, NY, USA, 2005; pp. 504–508. [Google Scholar]
- Ti, H.; Zhang, R.; Zhang, M.; Li, Q.; Wei, Z.; Zhang, Y.; Tang, X.; Deng, Y.; Liu, L.; Ma, Y. Dynamic changes in the free and bound phenolic compounds and antioxidant activity of brown rice at different germination stages. Food Chem. 2014, 161, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Yeo, J.D.; Shahidi, F. Critical evaluation of changes in the ratio of insoluble bound to soluble phenolics on antioxidant activity of lentils during germination. J. Agric. Food Chem. 2015, 63, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Basu, T.K.; Ooraikul, B. Studies on germination conditions and antioxidant contents of wheat grain. Int. J. Food Sci. Nutri. 2001, 52, 319–330. [Google Scholar] [CrossRef]
- Durmaz, G.; Gökmen, V. Changes in oxidative stability, antioxidant capacity and phytochemical composition of Pistacia terebinthus oil with roasting. Food Chem. 2011, 128, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Dewanto, V.; Wu, X.Z.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef] [PubMed]
- Nicoli, M.C.; Anese, M.; Parpinel, M. Influence of processing on the antioxidant properties of fruit and vegetables. Trends Food Sci. Technol. 1999, 10, 94–100. [Google Scholar] [CrossRef]
- Veldsink, J.W.; Muuse, B.G.; Meijer, M.M.T.; Cuperus, F.P.; Van de Sande, R.L.K.M.; Van Putte, K.P.A.M. Heat pretreatment of oilseeds: Effect on oil quality. Fett/Lipid 1999, 101, 244–248. [Google Scholar] [CrossRef]
- Chandrasekara, N.; Shahidi, F. Effect of roasting on phenolic content and antioxidant activities of whole cashew nuts, kernels and testa. J. Agric. Food Chem. 2011, 59, 5006–5014. [Google Scholar] [CrossRef] [PubMed]
- Vinson, J.A.; Cai, Y. Nuts, especially walnuts, have both antioxidant quantity and efficacy and exhibit significant potential health benefits. Food Funct. 2012, 3, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Nyembwe, P.; Minnaar, A.; Duodu, K.G.; Kock, H.L. Sensory and physicochemical analyses of roasted marama beans Tylosema esculentum (Burchell) A. Schreiber with specific focus on compounds that may contribute to bitterness. Food Chem. 2015, 178, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekara, N.; Shahidi, F. Antioxidative potential of cashew phenolics in food and biological model systems as affected by roasting. Food Chem. 2011, 129, 1388–1396. [Google Scholar] [CrossRef]
- Ee, K.Y.; Agboola, S.; Rehman, A.; Zhao, J. Characterisation of phenolic components present in raw and roasted wattle (Acacia victoriae Bentham) seeds. Food Chem. 2011, 129, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Ragaee, S.; Seetharaman, K.; Abdel-aal, E.M. A review: The impact of milling and thermal processing on phenolic compounds in cereal grains. Crit. Rev. Food Sci. Nutr. 2014, 54, 837–849. [Google Scholar] [CrossRef] [PubMed]
- Sarawong, C.; Schoenlechner, R.; Sekiguchi, K.; Berghofer, E.; Ng, P.K. Effect of extrusion cooking on the physicochemical properties, resistant starch, phenolic content and antioxidant capacities of green banana flour. Food Chem. 2014, 143, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Mora-Rochin, S.; Gutiérrez-Uribe, J.A.; Serna-Saldivar, S.O.; Sánchez-Peña, P.; Reyes-Moreno, C.; Milán-Carrillo, J. Phenolic content and antioxidant activity of tortillas produced from pigmented maize processed by conventional nixtamalization or extrusion cooking. J. Cereal Sci. 2010, 52, 502–508. [Google Scholar] [CrossRef]
- Gui, Y.; Ryu, G.H. Effects of extrusion cooking on physicochemical properties of white and red ginseng (powder). J. Ginseng Res. 2014, 38, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Ng, A.; Lecain, S.; Parker, M.L.; Smith, A.C.; Waldron, K.W. Modification of cell-wall polymers of onion waste III. Effect of extrusion-cooking on cell-wall material of outer fleshy tissues. Carbohydr. Polym. 1999, 39, 341–349. [Google Scholar] [CrossRef]
- Altan, A.; McCarthy, K.L.; Maskan, M. Effect of extrusion process on antioxidant activity, total phenolics and β-glucan content of extrudates developed from barley-fruit and vegetable by-products. Int. J. Food Sci. Technol. 2009, 44, 1263–1271. [Google Scholar] [CrossRef]
- Cardoso, L.M.; Pinheiro, S.S.; Carvalho, C.W.P.; Queiroz, V.A.V.; Menezes, C.B.; Moreira, A.V.B.; Barros, F.A.R.; Awika, J.M.; Martino, H.S.D.; Pinheiro-Sant’Ana, H.M. Phenolic compounds profile in sorghum processed by extrusion cooking and dry heat in a conventional oven. J. Cereal Sci. 2015, 65, 220–226. [Google Scholar] [CrossRef]
- Killeit, U. Vitamin retention in extrusion-cooking. Food Chem. 1994, 49, 149–155. [Google Scholar] [CrossRef]
- Yajnik, M.M.; Dolan, K.D.; Ng, P.K. Thermal and mechanical effects on retention of food-grade beta-carotene during extrusion processing. Int. J. Food Engineering. 2010, 6, 1556–3758. [Google Scholar] [CrossRef]
- Hansen, H.B.; Andreasen, M.F.; Nielsen, M.M.; Larsen, L.M.; Bach-Knudsen, K.E.; Meyer, A.S.; Christensen, L.P.; Hansen, Å. Changes in dietary fibre, phenolic acids and activity of endogenous enzymes during rye bread-making. Eur. Food Res. Technol. 2002, 214, 33–42. [Google Scholar] [CrossRef]
- Zdunczyk, Z.; Juskiewicz, J.; Wroblewska, M.; Flis, M. Effect of faba bean seeds with different content of proanthocyanidins on growth of rats, caecum enzyme activity and metabolism indices. Acta Aliment. 2003, 32, 161–168. [Google Scholar] [CrossRef]
- Cha´ vez-Reyes, Y.; Dorantes-Alvarez, L.; Arrieta-Baez, D.; Osorio-Esquivel, O.; Ortiz-Moreno, A. Polyphenol oxidase inactivation by microwave oven and its effect on phenolic profile of loquat (Eriobotrya japonica) fruit. Food Nutr. Sci. 2013, 4, 87–94. [Google Scholar] [CrossRef]
- Ebun, O.; Santosh, K. Effect of domestic cooking on the polyphenolic content and antioxidant capacity of plantain (Musa paradisiaca). World J. Dairy Food Sci. 2011, 6, 189–194. [Google Scholar]
- Chumyam, A.; Whangchai, K.; Jungklang, J.; Faiyue, B.; Saengnil, K. Effects of heat treatments on antioxidant capacity and total phenolic content of four cultivars of purple skin eggplants. Sci. Asia 2013, 39, 246–251. [Google Scholar] [CrossRef]
- Burgos, G.; Amoros, W.; Mun˜oa, L.; Sosa, P.; Cayhualla, E.; Sanchez, C.; Dı´az, C.; Bonierbale, M. Total phenolic, total anthocyanin and phenolic acid concentrations and antioxidant activity of purple-fleshed potatoes as affected by boiling. J. Food Comp. Anal. 2013, 30, 6–12. [Google Scholar] [CrossRef]
- Bellail, A.A.; Shaltout, O.E.; Youssef, M.M.; Gamal, A.M.E. Effect of home cooking methods on phenolic composition and antioxidant activity of sweet potato (Ipomoea batatas (L.) Lam.) cultivars grown in Egypt. Carbohydr. Polym. 2012, 39, 341–349. [Google Scholar]
- Yamaguchi, T.; Mizobuchi, T.; Kajikawa, R.; Kawashima, H.; Miyabe, F.; Terao, J.; Takamura, H.; Matoba, T. Radical-scavenging activity of vegetables and the effect of cooking on their activity. Food Sci. Technol. Res. 2001, 7, 250–257. [Google Scholar] [CrossRef]
- Miglio, C.; Chiavaro, E.; Visconti, A.; Fogliano, V.; Pellegrini, N. Effects of different cooking methods on nutritional and physicochemical characteristics of selected vegetables. J. Agric. Food Chem. 2008, 56, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Tsamo, C.V.P.; Herent, M.F.; Tomekpe, K.; Emaga, T.H.; Quetin-Leclercq, J.l.; Rogez, H.; Larondelle, Y.; Andre, C.M. Effect of boiling on phenolic profiles determined using HPLC/ESI-LTQ-Orbitrap-MS, physico-chemical parameters of six plantain banana cultivars (Musa sp). J. Food Comp. Anal. 2015, 44, 158–169. [Google Scholar] [CrossRef]
- Mazzeo, T.; N’Dri, D.; Chiavaro, E.; Visconti, A.; Fogliano, V.; Pellegrini, N. Effect of two cooking procedures on phytochemical compounds, total antioxidant capacity and colour of selected frozen vegetables. Food Chem. 2011, 128, 627–633. [Google Scholar] [CrossRef]
- Scaglioni, P.T.; de Souza, T.D.; Schmidt, C.G.; Badiale-Furlong, E. Availability of free and bound phenolic compounds in rice after hydrothermal treatment. J. Cereal Sci. 2014, 60, 526–532. [Google Scholar] [CrossRef]
- López, A.; El-Naggar, T.; Dueñas, M.; Ortega, T.; Estrella, I.; Hernández, T.; Gómez-Serranillos, M.P.; Palomino, O.M.; Carretero, M.E. Effect of cooking and germination on phenolic composition and biological properties of dark beans (Phaseolus vulgaris L.). Food Chem. 2013, 138, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Siah, S.; Wooda, J.A.; Agboola, S.; Konczak, I.; Blanchard, C.L. Effects of soaking, boiling and autoclaving on the phenolic contents and antioxidant activities of faba beans (Vicia faba L.) differing in seed coat colours. Food Chem. 2014, 142, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Xiaoyun, X.; Wende, L.; Zanhui, L.; Trust, B.; Arnie, W.H. Phenolic content, composition, antioxidant activity and their changes during domestic cooking of potatoes. J. Agric. Food Chem. 2009, 57, 10231–10238. [Google Scholar]
- Aguilera, Y.; Estrella, I.; Benitez, V.; Esteban, R.M.; Martin-Cabrejas, M.A. Bioactive phenolic compounds and functional properties of dehydrated beans flours. Food Res. Int. 2011, 44, 774–780. [Google Scholar] [CrossRef]
- Siddhuraju, P. The antioxidant activity and free radical-scavenging capacity of phenolics of raw and dry heated moth bean (Vigna aconitifolia) (Jacq.) Marechal seed extracts. Food Chem. 2006, 99, 149–157. [Google Scholar] [CrossRef]
- Turkmen, N.; Sari, F.; Velioglu, Y.S. The effect of cooking methods on total phenolics and antioxidant activity of selected green vegetables. Food Chem. 2005, 93, 713–718. [Google Scholar] [CrossRef]
- Hirawan, R.; Ser, W.Y.; Arntfield, S.D.; Beta, T. Antioxidant properties of commercial, regular- and whole-wheat spaghetti. Food Chem. 2010, 119, 258–264. [Google Scholar] [CrossRef]
- Ozdal, T.; Capanoglu, E.; Altay, F. A review on protein–phenolic interactions and associated changes. Food Res. Int. 2013, 51, 954–970. [Google Scholar] [CrossRef]
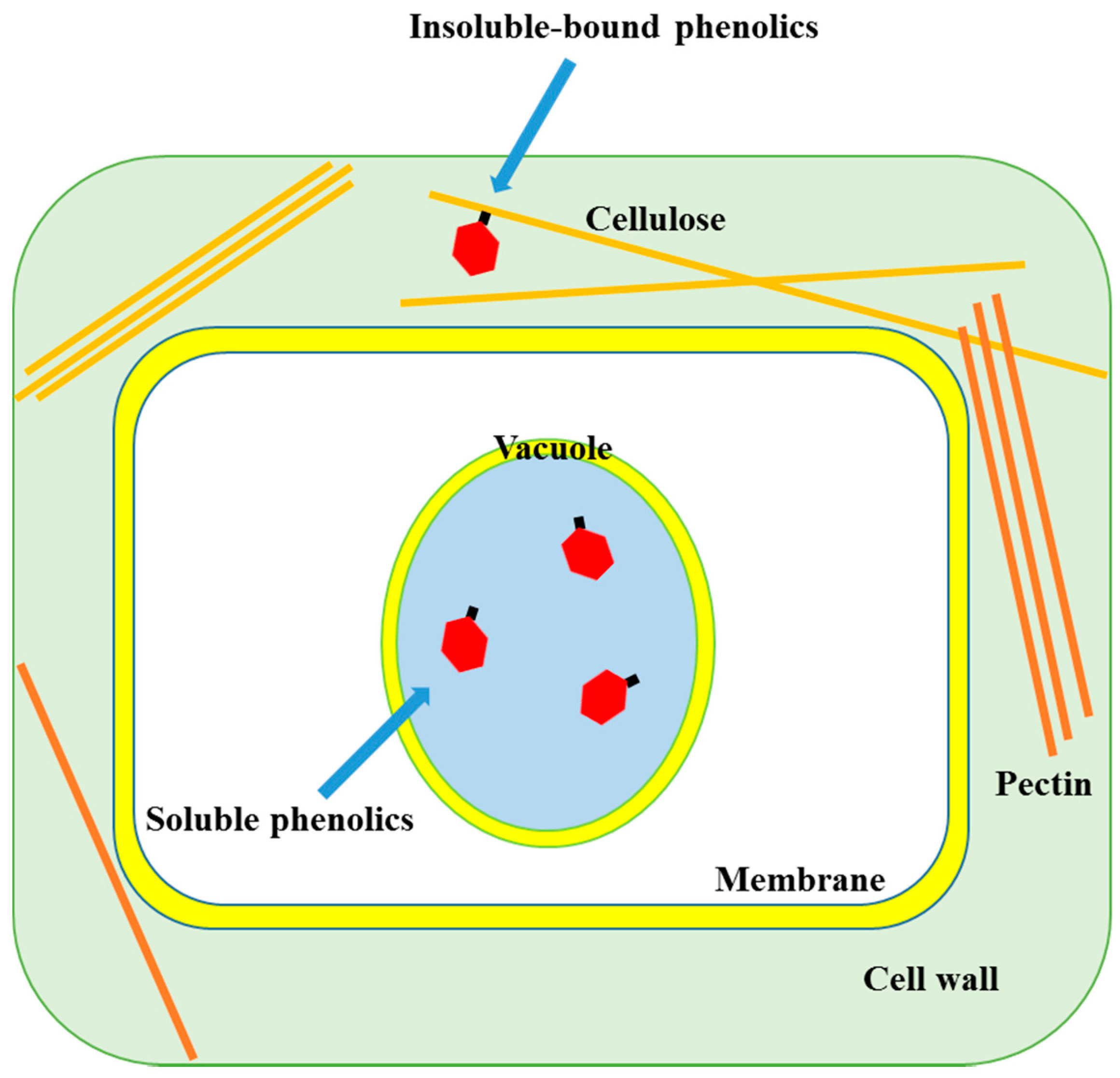
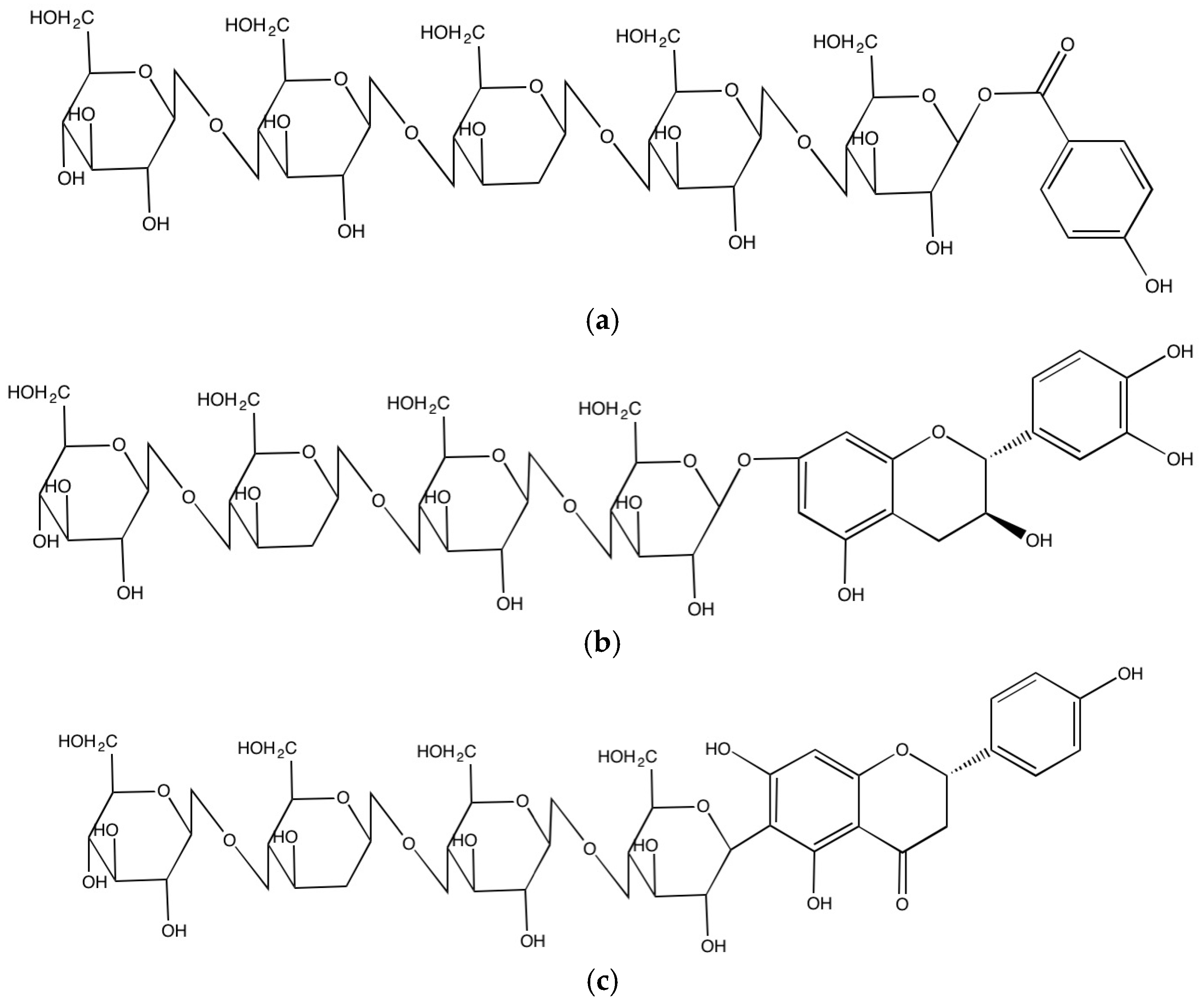


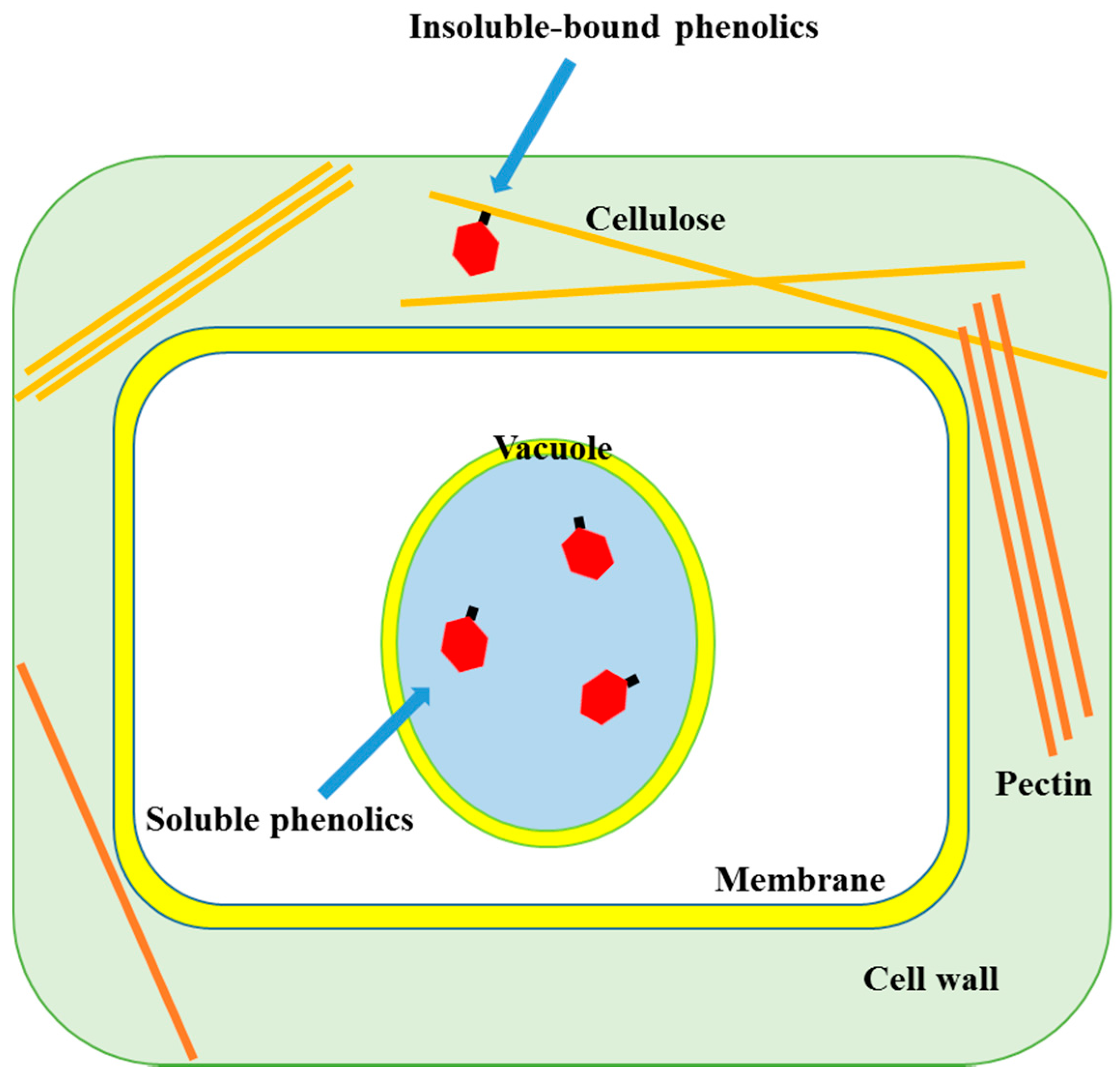{kind=link}
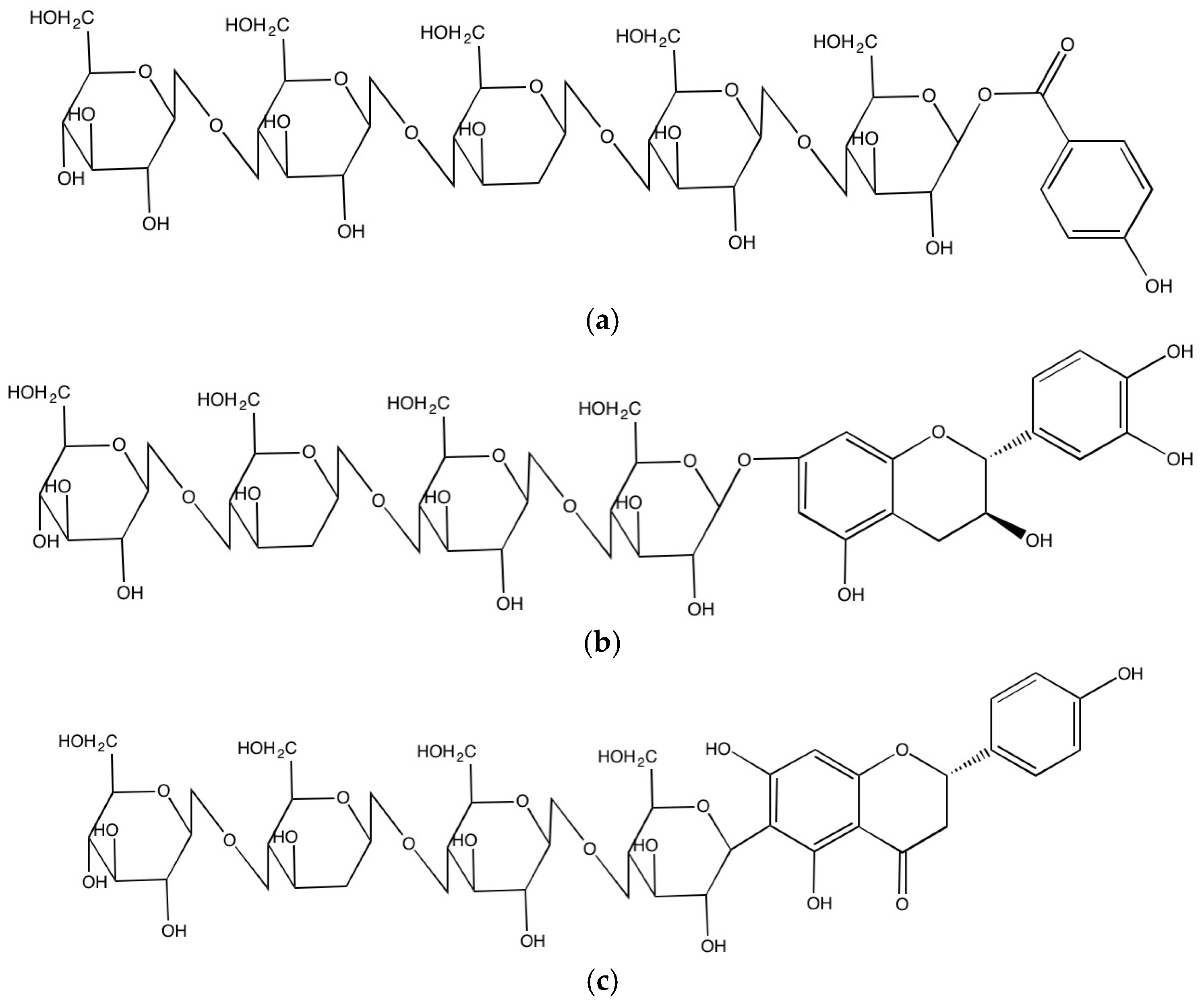{kind=link}
{kind=link}
| Sources | Bound Phenolic Acids (μg/g, DW) | Reference |
|---|---|---|
| Cereals | ||
| Black rice | Protocatechuic (162.1), 4-hydroxybenzoic (21.2), vanillic (27.7), p-coumaric (17.6) and ferulic (64.7) acids | [20] |
| Maize (Pericarp) | p-Hydroxybenzoic (499.0), vanillic (1788.0), syringic (83.0), p-coumaric (3.0), ferulic (3247.0) and isoferulic (842.0) acids | [21] |
| Maize (Germ) | p-Hydroxybenzoic (21.0), vanillic (522.0), syringic (897.0), caffeic (4.0), p-coumaric (12.0), ferulic (247.0) and isoferulic (58.0) acids | |
| Maize (endosperm) | p-Hydroxybenzoic (10.0), vanillic (6.0), caffeic (0.6), p-coumaric (0.3), ferulic (27.0) and isoferulic (1.0) acids | |
| Durum wheat | Gallic (6.0), 4-hydroxybenzoic (4.4), vanillic (2.9), syringic (2.9), p-coumaric (11.8), trans-ferulic (216.3), sinapic (41.2) and trans-cinnamic (5.4) acids | [22] |
| Bread wheat | Gallic (5.9), 4-hydroxybenzoic (4.7), vanillic (2.9), Syringic (3.7), p-coumaric (13.9), trans-ferulic (253.1), sinapic (45.5) and trans-cinnamic (5.2) acids | |
| Barley | Gallic (5.5), protocatechuic (4.9), 4-hydroxybenzoic (5.6), vanillic (5.4), caffeic (4.6), syringic (4.6), p-coumaric (75.9), trans-ferulic (320. 8), sinapic (41.9), salicylic (9.7) and trans-cinnamic (5.9) acids | |
| Oat | 4-Hydroxybenzoic (6.3), vanillic (12.9), caffeic (6.5), syringic (8.4), p-coumaric (603.0), trans-ferulic (1032.8), sinapic (100.6) and trans-cinnamic (5.4) acids | |
| Rye | Gallic (7.1), protocatechuic (4.2), 4-hydroxybenzoic (5.3), vanillic (4.1), caffeic (5.1), syringic (3.4), p-coumaric (20.7), trans-ferulic (216.7), sinapic (44.7) and trans-cinnamic (4.6) acids | |
| Rice | Gallic (5.1), 4-hydroxybenzoic (4.5), vanillic (1.7), syringic (2.8), p-coumaric (20.9), trans-ferulic (65.5), sinapic (22.8) and trans-cinnamic (4.9) acids | |
| Triticale | Gallic (4.8), protocatechuic (3.8), 4-hydroxybenzoic (5.4), vanillic (5.4), caffeic (4.3), syringic (4.1), p-coumaric (19.9), trans-ferulic (334.1), sinapic (58.4), salicylic (10.9) and trans-cinnamic (5.6) acids | |
| Corn | 4-Hydroxybenzoic (4.7), vanillic (4.9), caffeic (4.9), syringic (4.2), p-coumaric (95.4), trans-ferulic (954.4), sinapic (78.8) and trans-cinnamic (5.2) acids | |
| Millet (7 cultivars) | Ferulic (178.0-1685.0) and p-coumaric (20.0-1139.0) acids | [23] |
| Legumes | ||
| Lentils (6 cultivars) | Gallic acid (0.0-1.8), protocatechuic acid (0.0-4.4), p-coumaric acid (0.0-3.1) and ferulic acid (0.0-0.3) acids | [24] |
| Cranberry beans (7 cultivars) | p-Hydroxybenzoic (4.4-71.2), p-coumaric (4.9-18.4), ferulic (11.0-41.7) and sinapic (0.03-3.55) acids | [25] |
| Soy isoflavone concentrate | p-Hydroxybenzoic (12.0), vanillic (62.0), syringic (262.0), p-coumaric (61.0), ferulic (55.0) and sinapic (25.0) acids | [26] |
| Mung bean | Gallic (3.0), caffeic (0.3), p-coumaric (0.2), ferulic (2.2) and sinapic (1.5) acid | [27] |
| Pinto bean | Protocatechuic (25.6), p-hydroxybenzoic (42.7), p-coumaric (26.8), ferulic (34.7) and sinapic (89.9) acid | [28] |
| Black bean | Gallic (41.1), protocatechuic (14.1), p-hydroxybenzoic (5.6), caffeic (17.5), syringic (17.1), p-coumaric (22.1), ferulic (170.1) and sinapic (50.5) acid | |
| Kidney bean | Protocatechuic (64.4), p-hydroxybenzoic (21.3), p-coumaric (7.9), ferulic (138.0) and sinapic (73.4) acid | |
| Cowpea | Gallic (5.4) acid | [29] |
| Chickpea | Gallic (82.8) and protocatechuic (110.9) acid | [30] |
| Cow gram | Protocatechuic (172.6) acid | |
| Flower waist bean | Protocatechuic (128.4) acid | |
| Hyacinth bean | Protocatechuic (111.8) acid | |
| Pearl bean | Protocatechuic (121.9) acid | |
| Oilseeds | ||
| Sunflower | Gallic (11.2), protocatechuic (50.8), caffeic (25.5), p-coumaric (2.5), ferulic (16.9) and sinapic (15.6) acids | [27] |
| Rapeseed meal (seven cultivars) | 389-1050 μg sinapic acid equivalent/g DW (Main phenoic acids: sinapic, p-coumaric and trans-ferulic acid) | [31] |
| Flaxseed | p-Coumaric acid glucoside (3800.0) and ferulic acid glucoside (4400.0) | [32] |
| Moringa oleifera seed flour | Gallic (15.9) and p-coumaric (7.4) acids | [33] |
| Soy bean | Gallic (64.2), protocatechuic (238.8), vanillic (88.7), syringic (121.6), p-coumaric (101.7) and ferulic (87.1) acid | [34] |
| Fruit seeds | ||
| Blackberry seed meal | Protocatechuic (2.2), p-coumaric (0.2), gallic (3.4) and caffeic (2.1) acids | [35] |
| Black raspberry seed meal | Protocatechuic (0.9), p-coumaric (2.4), gallic (2.4) and caffeic (1.5) acids | |
| Blueberry seed meal | Protocatechuic (0.3), p-coumaric (0.2), gallic(3.5) and caffeic (0.2), syringic (0.9) acids; gallic hexoside (1.0) |
| Sources | Bound Flavonoids (μg/g, DW) | Reference |
|---|---|---|
| Cereals | ||
| Maize (pericarp) | Kaempferol (15.9) | [21] |
| Maize (germ) | Quercetin (28.8) | |
| Quinoa seed | Catechin (16.28), quercetin (47.2) and kaempferol (30.4) | [36] |
| Dent corn (pericarp portion) | Quercetin (35.9) | [37] |
| Flint corn (germ portion) | Quercetin (12.0) | |
| Rice | 240 μg catechin equivalent/g DW | [34] |
| Corn | 760 μg catechin equivalent/g DW | |
| Wheat | 430 μg catechin equivalent/g DW | |
| Barley | 370 μg catechin equivalent/g DW | |
| Legumes | ||
| Lentils (6 cultivars) | Catechin (15.0–78.4), epicatechin (0.5–7.9), (+)-catechin-3-glucoside (0.0–122.0) and luteolin 3′-7-diglucoside (0.0–49.3) | [24] |
| Cranberry beans (4 cultivars) | Kaempferol (0.2–0.3) | [25] |
| Mung bean | Quercetin (0.2) and kaempferol (0.1) | [27] |
| Black bean | (+)-Catechin (109.7), epicatechin (93.8), rutin (93.8), isoquercitrin (462.3) and quercetin (86.6) | [30] |
| Cow gram | Quercitrin (105.4) | |
| Hyacinth bean | Quercitrin (91.9) | |
| Pearl bean | (+)-Catechin (100.7) | |
| Red bean | Rutin (89.5) | |
| Red kidney bean | Catechin (88.2) and isoquercitrin (97.1) | |
| Spring bay bean | Catechin (158.8), rutin (124.2), isoquercitrin (85.7) and quercetin (86.8) | |
| Oilseeds | ||
| Moringa oleifera seed flour | Catechin (7490.0), epicatechin (810.0) and quercetin (18.7) | [33] |
| Sunflower | Quercetin (1.5), kaempferol (0.5) and apigenin (2.9) | [27] |
| Soy bean | Epicatechin (95.2), isoquercitrin (396.0) and quercetin (101.0) | [30] |
| Fruit seeds | ||
| Blackberry | (+)-Catechin (1.0), (−)-epicatechin (1.1), quercetin (2.0), quercetin pentose (3.3), epicatechin gallate (1.1) and quercetin-3-O-glucuronide (5.1) | [29] |
| Black raspberry | (+)-Catechin (0.4), (−)-epicatechin (3.1), quercetin (3.2), epigallocatechin (1.3), epicatechin gallate (0.9) and quercetin-3-O-glucuronide (11.4) | |
| Blueberry | (+)-Catechin (0.7), (−)-epicatechin (0.07), quercetin (0.2), epigallocatechin (0.4), myricetin (0.2), quercetin pentose (0.5) and kaempferol hexoside (0.6) |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahidi, F.; Yeo, J. Insoluble-Bound Phenolics in Food. Molecules 2016, 21, 1216. https://doi.org/10.3390/molecules21091216
Shahidi F, Yeo J. Insoluble-Bound Phenolics in Food. Molecules. 2016; 21(9):1216. https://doi.org/10.3390/molecules21091216
Chicago/Turabian StyleShahidi, Fereidoon, and JuDong Yeo. 2016. "Insoluble-Bound Phenolics in Food" Molecules 21, no. 9: 1216. https://doi.org/10.3390/molecules21091216
APA StyleShahidi, F., & Yeo, J. (2016). Insoluble-Bound Phenolics in Food. Molecules, 21(9), 1216. https://doi.org/10.3390/molecules21091216