
Differential Effects of Viscum album Preparations on the Maturation and Activation of Human Dendritic Cells and CD4+ T Cell Responses


 and
and
Abstract
:1. Introduction
2. Results
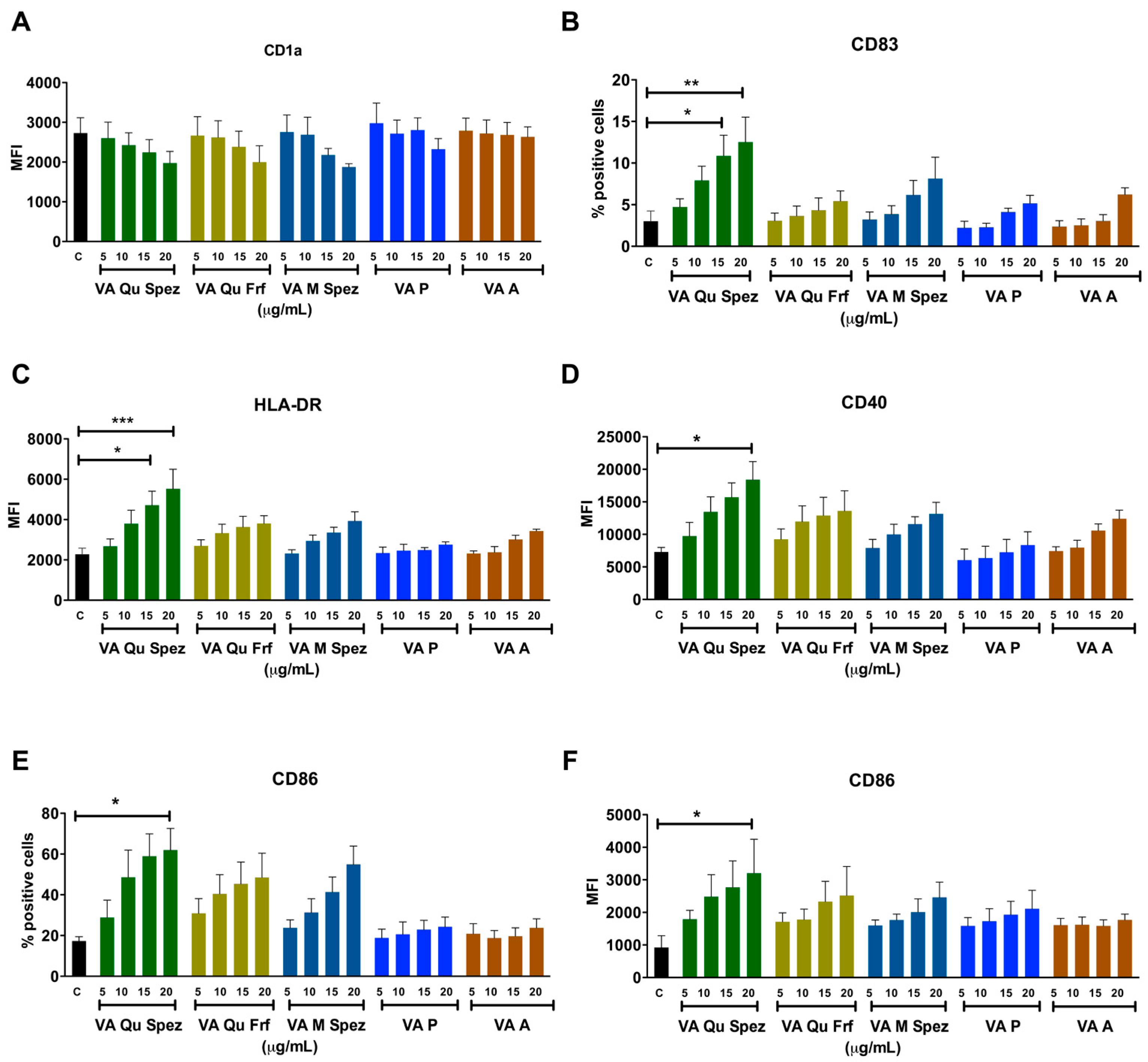
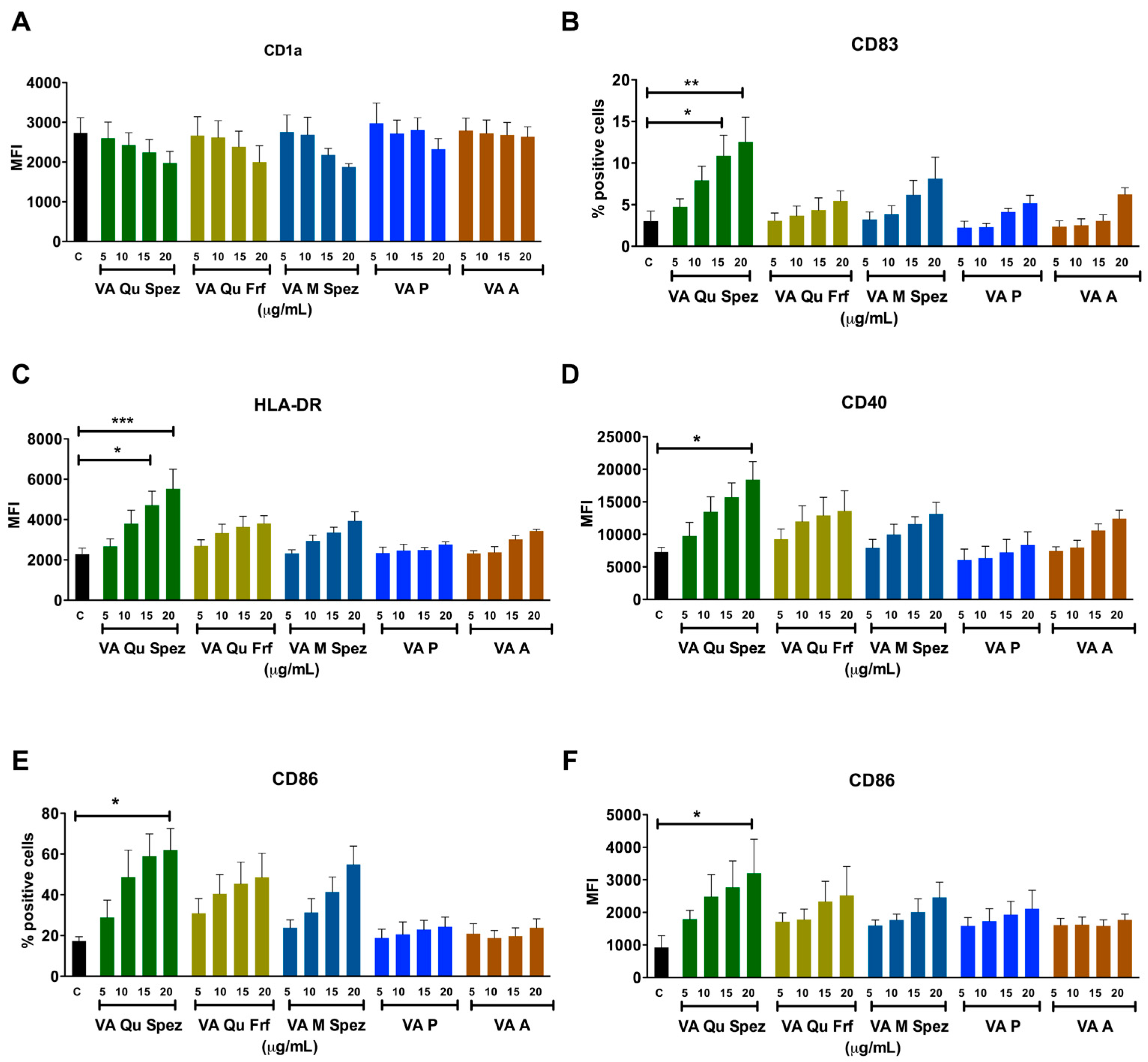
2.1. Effect of Different VA Preparations on the Maturation of DCs
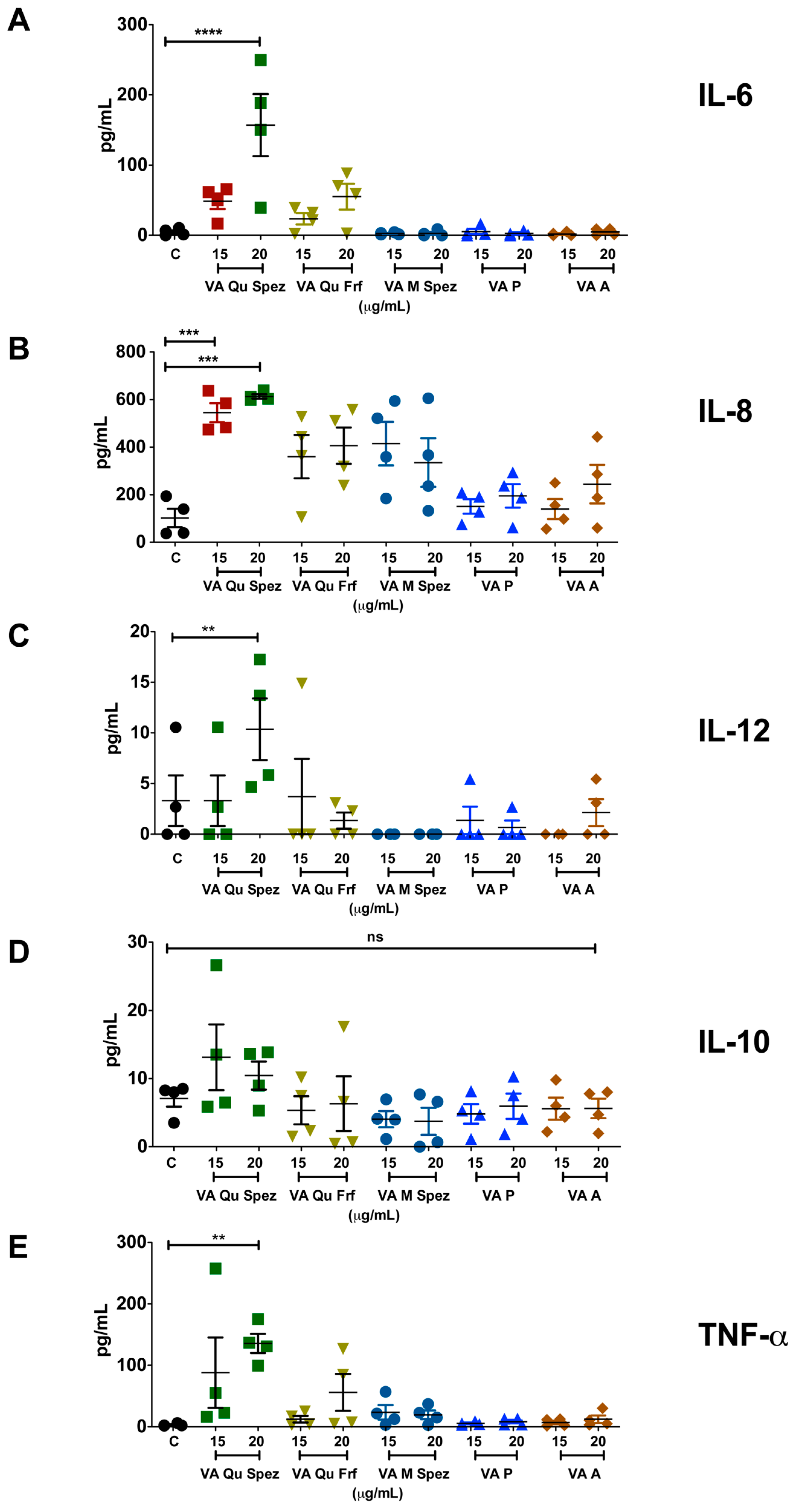
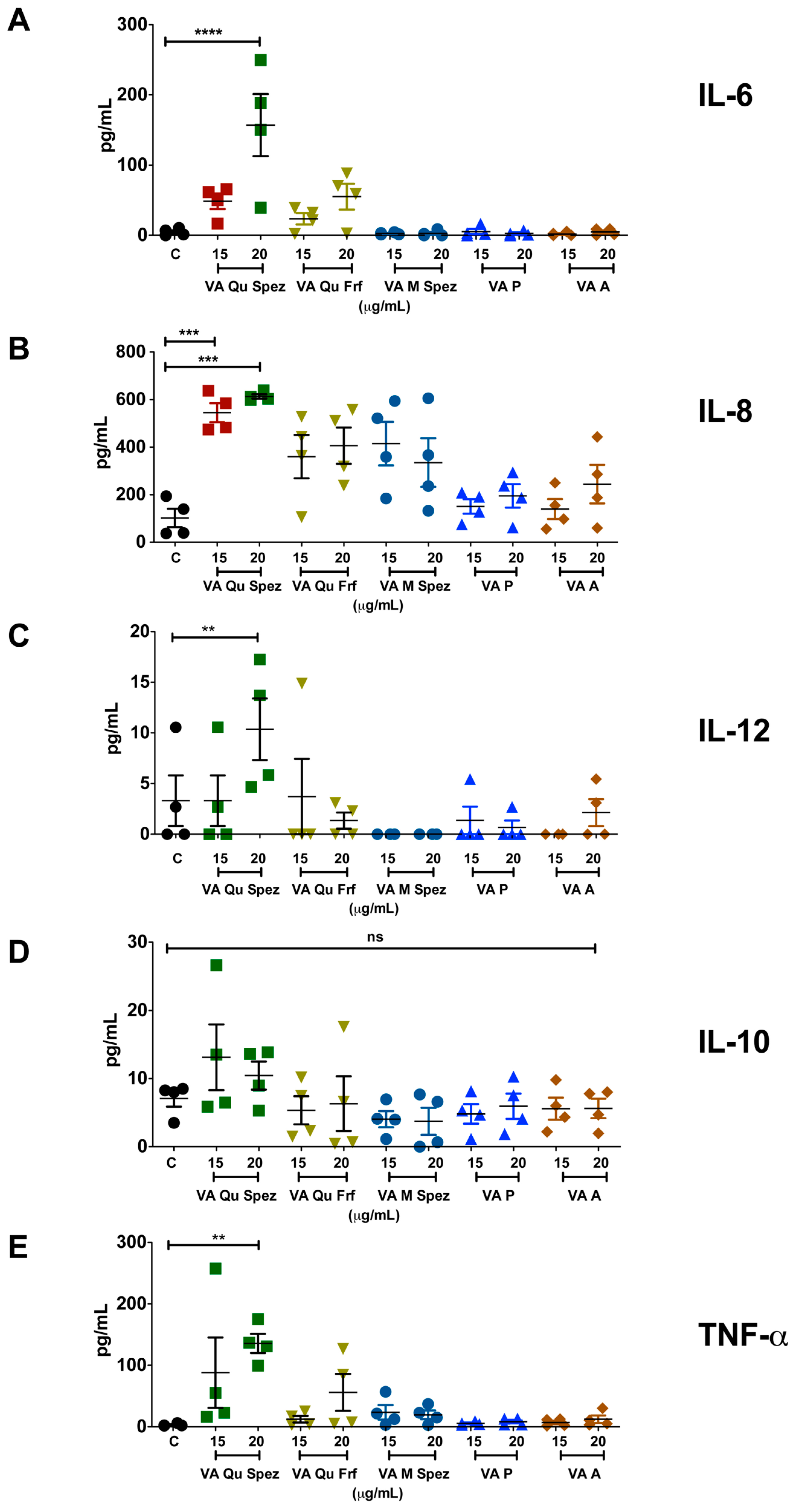
2.2. VA Qu Spez but Not Other VA Preparations Stimulate the Secretion of DC Cytokines
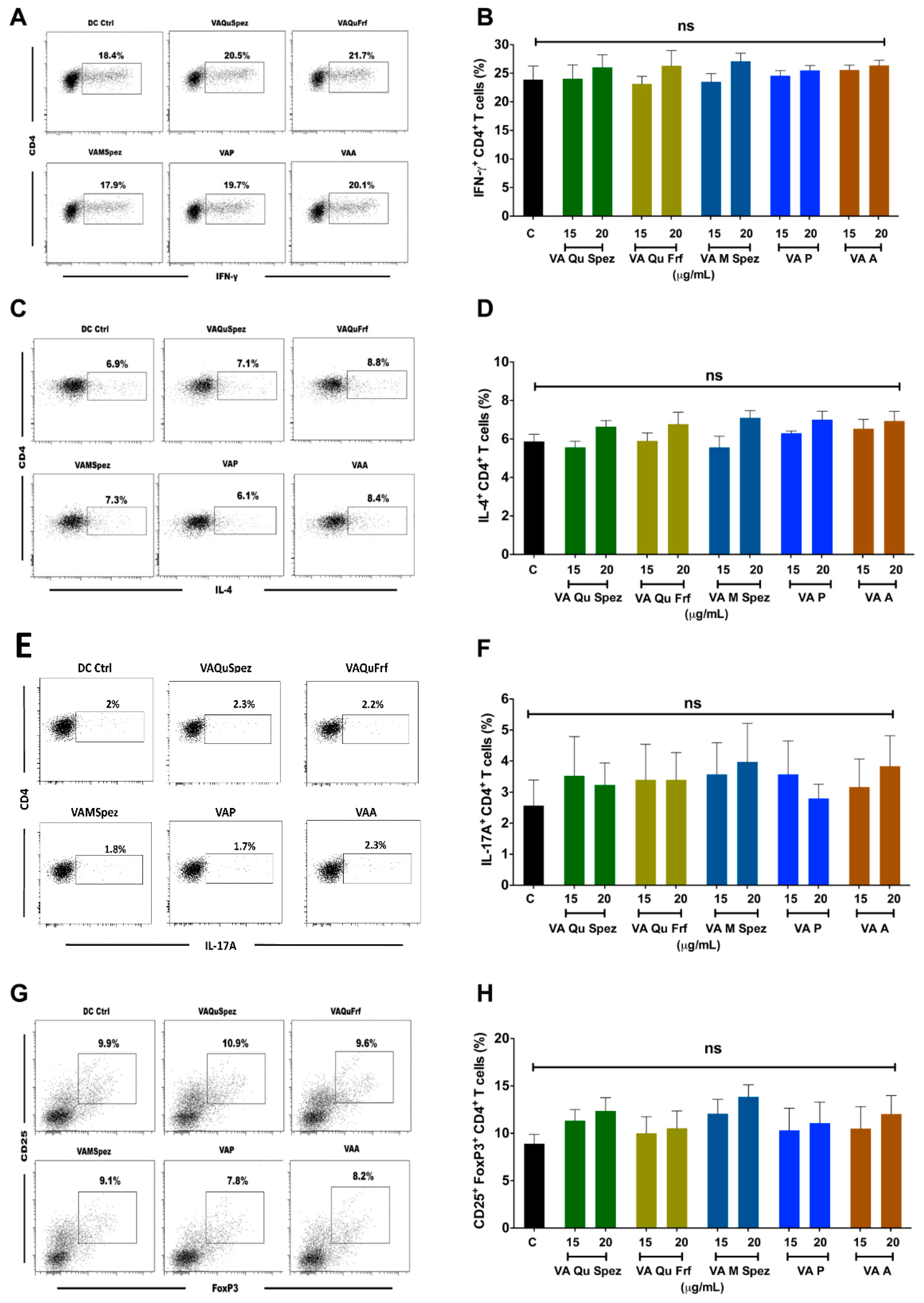
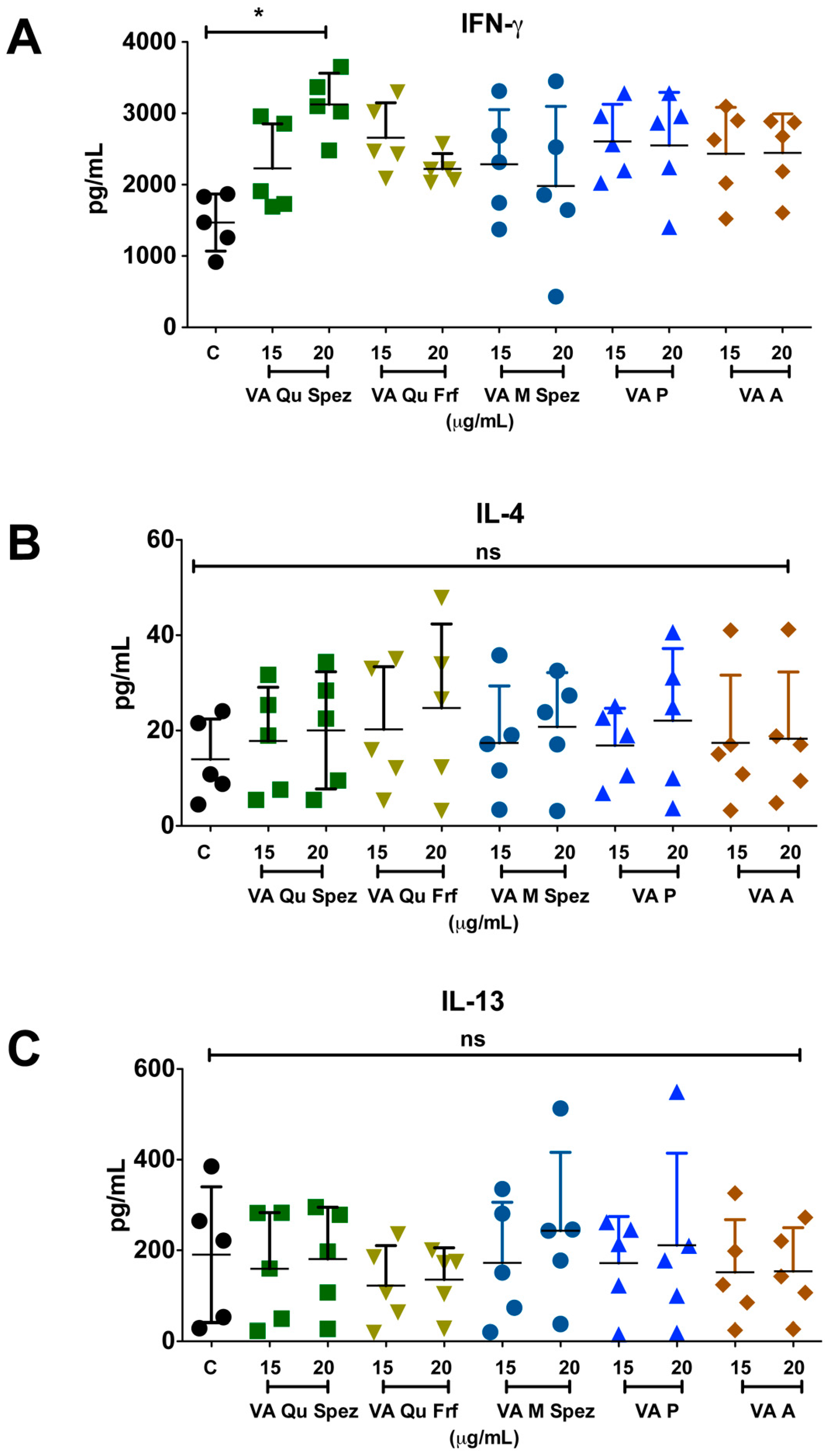
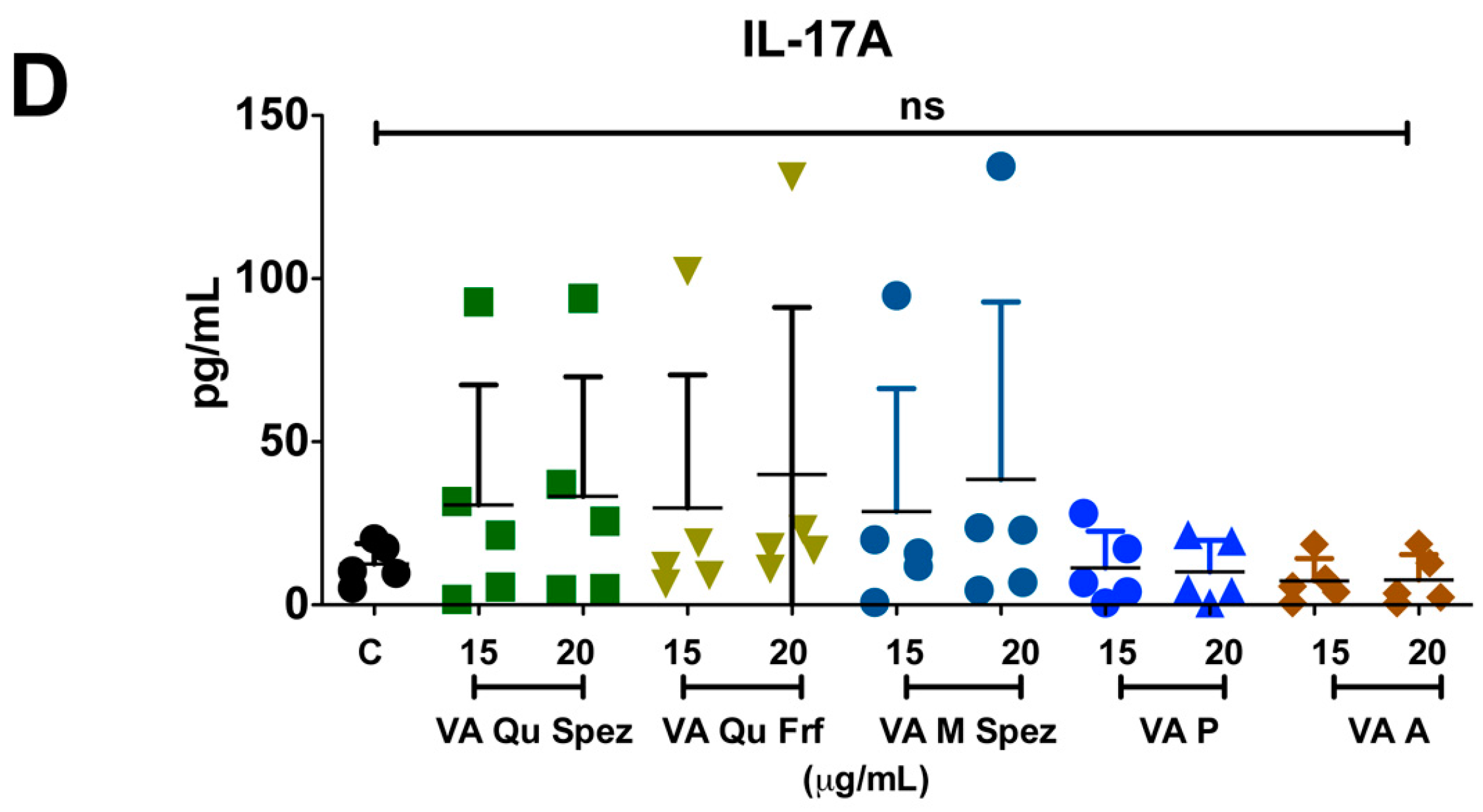
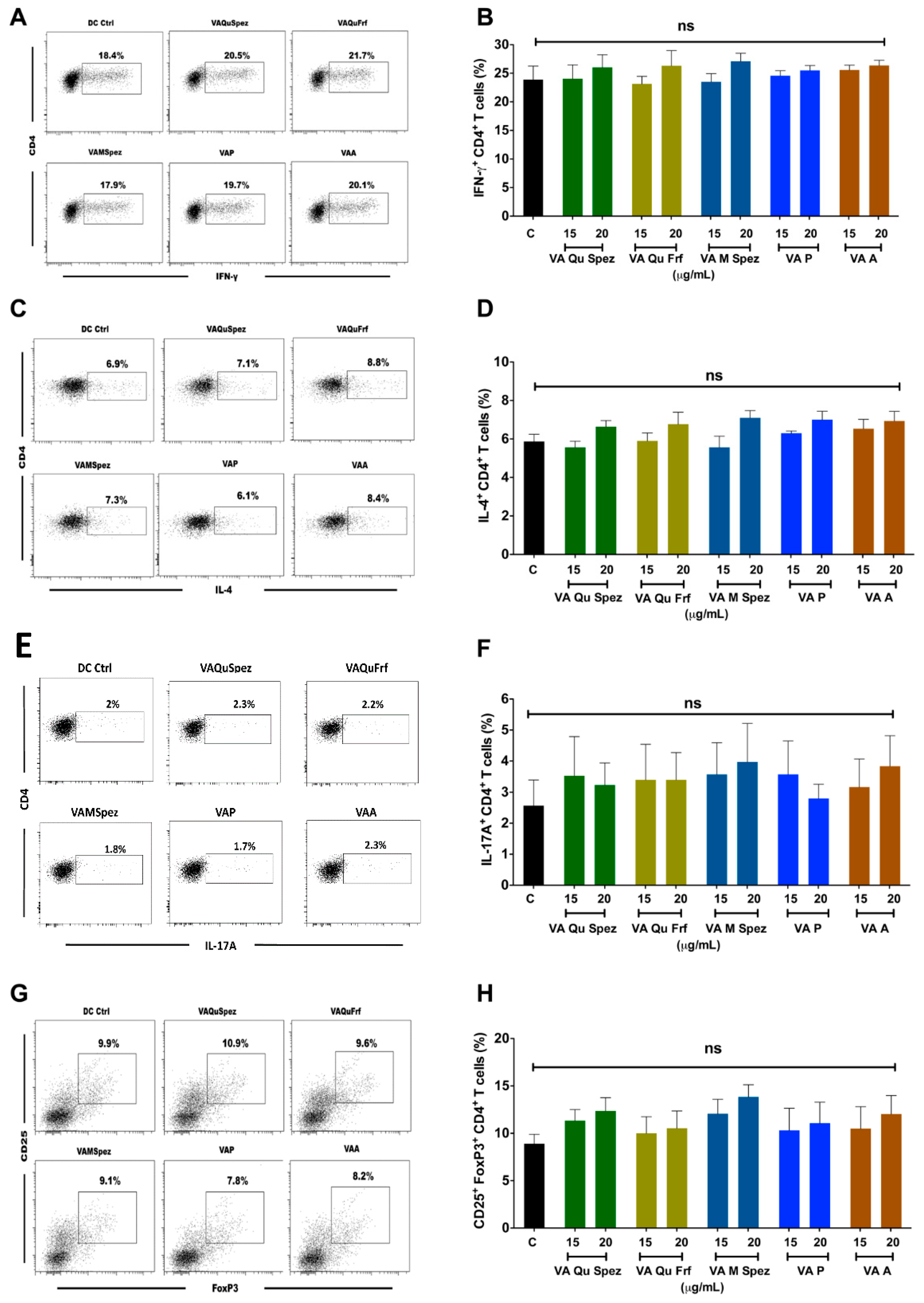
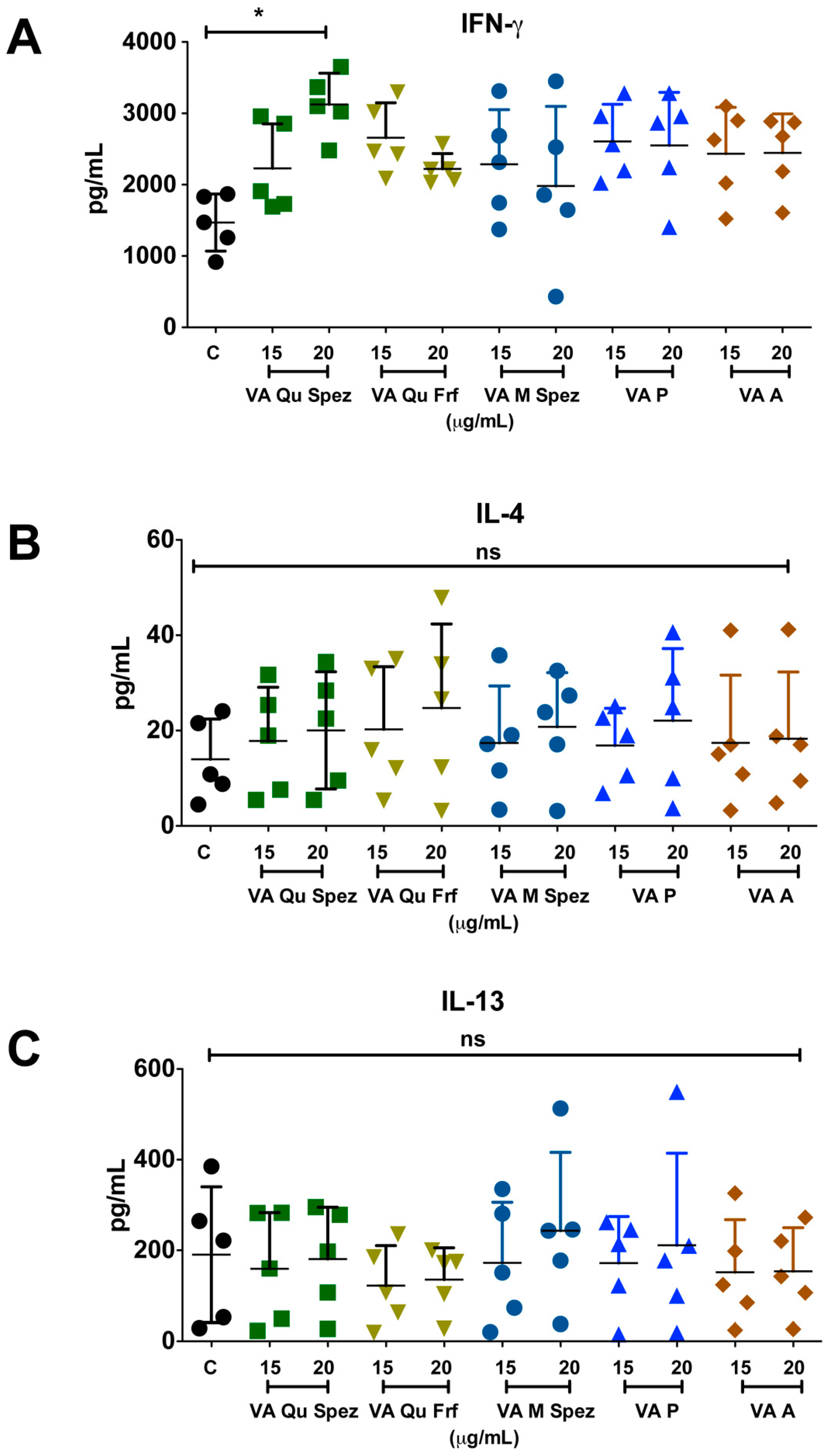
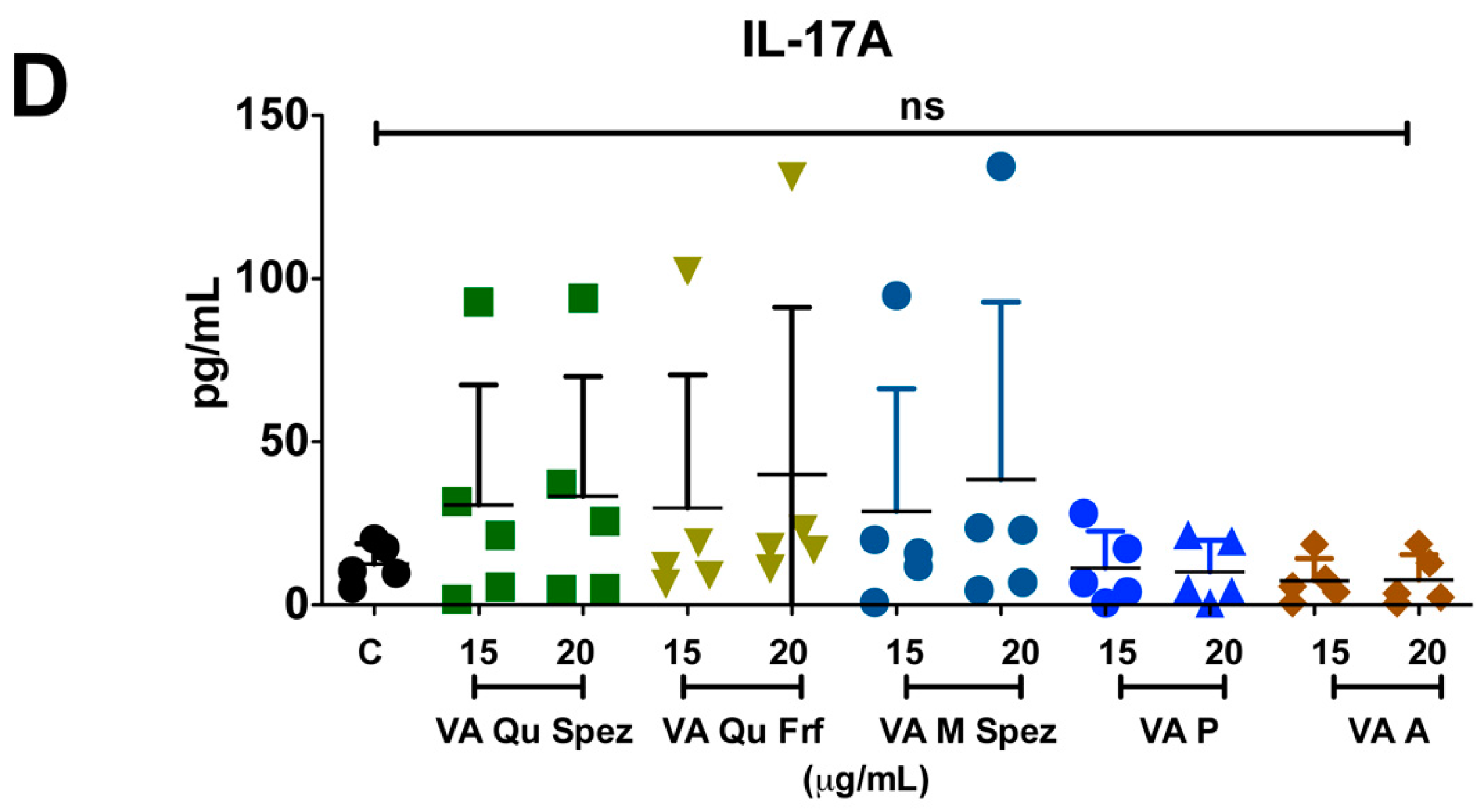
2.3. Differential Effects of VA Preparations on the CD4+ T Cell Response
3. Discussion
4. Materials and Methods
4.1. VA Preparations
4.2. Human DCs
4.3. Viscum Album Treatment of DCs
4.4. DC: CD4+ T Cell Co-Cultures
4.5. Flow Cytometry
4.6. Cytokine Assay
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bock, P.R.; Hanisch, J.; Matthes, H.; Zanker, K.S. Targeting inflammation in cancer-related-fatigue: A rationale for mistletoe therapy as supportive care in colorectal cancer patients. Inflamm. Allergy Drug Targets 2014, 13, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Podlech, O.; Harter, P.N.; Mittelbronn, M.; Poschel, S.; Naumann, U. Fermented mistletoe extract as a multimodal antitumoral agent in gliomas. Evid. Based Complement. Altern. Med. 2012, 2012, 501796. [Google Scholar] [CrossRef] [PubMed]
- Kienle, G.S.; Berrino, F.; Bussing, A.; Portalupi, E.; Rosenzweig, S.; Kiene, H. Mistletoe in cancer—A systematic review on controlled clinical trials. Eur. J. Med. Res. 2003, 8, 109–119. [Google Scholar] [PubMed]
- Singh, B.N.; Saha, C.; Galun, D.; Upreti, D.K.; Bayry, J.; Kaveri, S.V. European Viscum album: A potent phytotherapeutic agent with multifarious phytochemicals, pharmacological properties and clinical evidence. RSC Adv. 2016, 6, 23837–23857. [Google Scholar] [CrossRef]
- Augustin, M.; Bock, P.R.; Hanisch, J.; Karasmann, M.; Schneider, B. Safety and efficacy of the long-term adjuvant treatment of primary intermediate- to high-risk malignant melanoma (UICC/AJCC stage II and III) with a standardized fermented European mistletoe (Viscum album L.) extract. Results from a multicenter, comparative, epidemiological cohort study in Germany and Switzerland. Arzneimittelforschung 2005, 55, 38–49. [Google Scholar] [PubMed]
- Melzer, J.; Iten, F.; Hostanska, K.; Saller, R. Efficacy and safety of mistletoe preparations (Viscum album) for patients with cancer diseases. A systematic review. Forsch. Komplementmed. 2009, 16, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Endo, Y.; Tsurugi, K.; Franz, H. The site of action of the A-chain of mistletoe lectin I on eukaryotic ribosomes. The RNA N-glycosidase activity of the protein. FEBS Lett. 1988, 231, 378–380. [Google Scholar] [CrossRef]
- Stirpe, F.; Barbieri, L.; Battelli, M.G.; Soria, M.; Lappi, D.A. Ribosome-inactivating proteins from plants: Present status and future prospects. Biotechnology 1992, 10, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Steinman, R.M.; Banchereau, J. Taking dendritic cells into medicine. Nature 2007, 449, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Nakano, H.; Lin, K.L.; Yanagita, M.; Charbonneau, C.; Cook, D.N.; Kakiuchi, T.; Gunn, M.D. Blood-derived inflammatory dendritic cells in lymph nodes stimulate acute T helper type 1 immune responses. Nat. Immunol. 2009, 10, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B.; Tang, H.; Manicassamy, S. Programming dendritic cells to induce T(H)2 and tolerogenic responses. Nat. Immunol. 2010, 11, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; Plantinga, M.; Deswarte, K.; Pouliot, P.; Willart, M.A.; Kool, M.; Muskens, F.; Lambrecht, B.N. Inflammatory dendritic cells—Not basophils—Are necessary and sufficient for induction of Th2 immunity to inhaled house dust mite allergen. J. Exp. Med. 2010, 207, 2097–2111. [Google Scholar] [CrossRef] [PubMed]
- Segura, E.; Touzot, M.; Bohineust, A.; Cappuccio, A.; Chiocchia, G.; Hosmalin, A.; Dalod, M.; Soumelis, V.; Amigorena, S. Human inflammatory dendritic cells induce Th17 cell differentiation. Immunity 2013, 38, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Maddur, M.S.; Sharma, M.; Hegde, P.; Stephen-Victor, E.; Pulendran, B.; Kaveri, S.V.; Bayry, J. Human B cells induce dendritic cell maturation and favour Th2 polarization by inducing OX-40 ligand. Nat. Commun. 2014, 5, 4092. [Google Scholar]
- Lewis, K.L.; Reizis, B. Dendritic cells: Arbiters of immunity and immunological tolerance. Cold Spring Harb. Perspect. Biol. 2012, 4, a007401. [Google Scholar] [CrossRef] [PubMed]
- Vitali, C.; Mingozzi, F.; Broggi, A.; Barresi, S.; Zolezzi, F.; Bayry, J.; Raimondi, G.; Zanoni, I.; Granucci, F. Migratory, and not lymphoid-resident, dendritic cells maintain peripheral self-tolerance and prevent autoimmunity via induction of iTreg cells. Blood 2012, 120, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, D.; Tsunn, A.; Li, B. Foxp3+ regulatory T cells and their functional regulation. Cell. Mol. Immunol. 2015, 12, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Bayry, J. Repressing immunity in autoimmune disease. N. Engl. J. Med. 2016, 374, 2090–2092. [Google Scholar] [CrossRef] [PubMed]
- Almand, B.; Resser, J.R.; Lindman, B.; Nadaf, S.; Clark, J.I.; Kwon, E.D.; Carbone, D.P.; Gabrilovich, D.I. Clinical significance of defective dendritic cell differentiation n cancer. Clin. Cancer Res. 2000, 6, 1755–1766. [Google Scholar] [PubMed]
- Vicari, A.P.; Caux, C.; Trinchieri, G. Tumour escape from immune surveillance through dendritic cell inactivation. Sem. Cancer Biol. 2002, 12, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Gabrilovich, D.I.; Corak, J.; Ciernik, I.F.; Kavanaugh, D.; Carbone, D.P. Decreased antigen presentation by dendritic cells in patients with breast cancer. Clin. Cancer Res. 1997, 3, 483–490. [Google Scholar] [PubMed]
- Saito, H.; Tsujitani, S.; Ikeguchi, M.; Maeta, M.; Kaibara, N. Relationship between the expression of vascular endothelial growth factor and the density of dendritic cells in gastric adenocarcinoma tissue. Br. J. Cancer 1998, 78, 1573–1577. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Bullock, T.N. Metabolic influences that regulate dendritic cell function in tumors. Front. Immunol. 2014, 5, 24. [Google Scholar] [CrossRef] [PubMed]
- Hargadon, K.M.; Bullock, T.N. The role of tumor/dendritic cell interactions in the regulation of anti-tumor immunity: The good, the bad, and the ugly. Front. Immunol. 2014, 5, 178. [Google Scholar] [CrossRef] [PubMed]
- Elluru, S.R.; Duong van Huyen, J.P.; Delignat, S.; Kazatchkine, M.D.; Friboulet, A.; Kaveri, S.V.; Bayry, J. Induction of maturation and activation of human dendritic cells: A mechanism underlying the beneficial effect of Viscum album as complimentary therapy in cancer. BMC Cancer 2008, 8, 161. [Google Scholar] [CrossRef] [PubMed]
- Knopfl-Sidler, F.; Viviani, A.; Rist, L.; Hensel, A. Human cancer cells exhibit in vitro individual receptiveness towards different mistletoe extracts. Pharmazie 2005, 60, 448–454. [Google Scholar] [PubMed]
- Hulsen, H.; Doser, C.; Mechelke, F. Differences in the in vitro effectiveness of preparations produced from mistletoes of various host trees. Arzneimittelforschung 1986, 36, 433–436. [Google Scholar] [PubMed]
- Kleijnen, J.; Knipschild, P. Mistletoe treatment for cancer review of controlled trials in humans. Phytomedicine 1994, 1, 255–260. [Google Scholar] [CrossRef]
- Ribereau-Gayon, G.; Jung, M.L.; di Scala, D.; Beck, J.P. Comparison of the effects of fermented and unfermented mistletoe preparations on cultured tumor cells. Oncology 1986, 43 (Suppl. S1), 35–41. [Google Scholar] [CrossRef] [PubMed]
- Eggenschwiler, J.; von Balthazar, L.; Stritt, B.; Pruntsch, D.; Ramos, M.; Urech, K.; Rist, L.; Simões-Wüst, A.P.; Viviani, A. Mistletoe lectin is not the only cytotoxic component in fermented preparations of Viscum album from white fir (Abies pectinata). BMC Complement. Altern. Med. 2007, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Li, X.; Zhong, Y. Inflammatory cytokines: Potential biomarkers of immunologic dysfunction in autism spectrum disorders. Mediators Inflamm. 2015, 2015, 531518. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Shankaran, V.; Ikeda, H.; Bruce, A.T.; White, J.M.; Swanson, P.E.; Old, L.J.; Schreiber, R.D. IFNγ and lymphocytes prevent primary tumour development and shape tumour immunogenicity. Nature 2001, 410, 1107–1111. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Qian, J.; Yang, J.; Li, H.; Kwak, L.W.; Yi, Q. Roles of idiotype-specific t cells in myeloma cell growth and survival: Th1 and CTL cells are tumoricidal while Th2 cells promote tumor growth. Cancer Res. 2008, 68, 8456–8464. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, D.H.; Shankaran, V.; Dighe, A.S.; Stockert, E.; Aguet, M.; Old, L.J.; Schreiber, R.D. Demonstration of an interferon gamma-dependent tumor surveillance system in immunocompetent mice. Proc. Natl. Acad. Sci. USA 1998, 95, 7556–7561. [Google Scholar] [CrossRef] [PubMed]
- Palucka, K.; Banchereau, J. Cancer immunotherapy via dendritic cells. Nat. Rev. Cancer 2012, 12, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Trinath, J.; Hegde, P.; Sharma, M.; Maddur, M.S.; Rabin, M.; Vallat, J.M.; Magy, L.; Balaji, K.N.; Kaveri, S.V.; Bayry, J. Intravenous immunoglobulin expands regulatory T cells via induction of cyclooxygenase-2-dependent prostaglandin E2 in human dendritic cells. Blood 2013, 122, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Maddur, M.S.; Trinath, J.; Rabin, M.; Bolgert, F.; Guy, M.; Vallat, J.M.; Magy, L.; Balaji, K.N.; Kaveri, S.V.; Bayry, J. Intravenous immunoglobulin-mediated expansion of regulatory T cells in autoimmune patients is associated with increased prostaglandin E2 levels in the circulation. Cell. Mol. Immunol. 2015, 12, 650–652. [Google Scholar] [CrossRef] [PubMed]
- Holla, S.; Stephen-Victor, E.; Prakhar, P.; Sharma, M.; Saha, C.; Udupa, V.; Kaveri, S.V.; Bayry, J.; Balaji, K.N. Mycobacteria-responsive sonic hedgehog signaling mediates programmed death-ligand 1- and prostaglandin E2-induced regulatory T cell expansion. Sci. Rep. 2016, 6, 24193. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, D.K.; Dhodapkar, M.V.; Matayeva, E.; Steinman, R.M.; Dhodapkar, K.M. Expansion of FOXP3 high regulatory T cells by human dendritic cells (DCs) in vitro and after injection of cytokine-matured DCs in myeloma patients. Blood 2006, 108, 2655–2661. [Google Scholar] [CrossRef] [PubMed]
- Hegde, P.; Maddur, M.S.; Friboulet, A.; Bayry, J.; Kaveri, S.V. Viscum album exerts anti-inflammatory effect by selectively inhibiting cytokine-induced expression of cyclooxygenase-2. PLoS ONE 2011, 6, e26312. [Google Scholar] [CrossRef] [PubMed]
- Saha, C.; Hegde, P.; Friboulet, A.; Bayry, J.; Kaveri, S.V. Viscum album-mediated COX-2 inhibition implicates destabilization of COX-2 mRNA. PLoS ONE 2015, 10, e0114965. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Lee, J.; So, H.S.; Lee, K.M.; Jung, B.H.; Chung, S.Y.; Moon, S.R.; Kim, N.S.; Ko, C.B.; Kim, H.J.; et al. Gamma-interferon (IFN-gamma) augments apoptotic response to mistletoe lectin-II via upregulation of Fas/Fas L expression and caspase activation in human myeloid U937 cells. Immunopharmacol. Immunotoxicol. 2011, 23, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Bantel, H.; Engels, I.H.; Voelter, W.; Schulze-Osthoff, K.; Wesselborg, S. Mistletoe lectin activates caspase-8/FLICE independently of death receptor signaling and enhances anticancer drug-induced apoptosis. Cancer Res. 1999, 59, 2083–2090. [Google Scholar] [PubMed]
- Stettin, A.; Schultze, J.L.; Stechemesser, E.; Berg, P.A. Anti-mistletoe lectin antibodies are produced in patients during therapy with an aqueous mistletoe extract derived from Viscum album L. and neutralize lectin-induced cytotoxicity in vitro. Klin. Wochenschr. 1990, 68, 896–900. [Google Scholar] [CrossRef] [PubMed]
- Lavastre, V.; Pelletier, M.; Saller, R.; Hostanska, K.; Girard, D. Mechanisms involved in spontaneous and Viscum album agglutinin-I-induced human neutrophil apoptosis: Viscum album agglutinin-I accelerates the loss of antiapoptotic MCL-1 expression and the degradation of cytoskeletal paxillin and vimentin proteins via caspases. J. Immunol. 2002, 168, 1419–1427. [Google Scholar] [PubMed]
- Walzel, H.; Blach, M.; Neels, P.; Schulz, U.; Wollenhaupt, K.; Brock, J. The B-chain of mistletoe lectin I efficiently stimulates calcium signaling in human Jurkat T-cells. Immunol. Lett. 2001, 78, 57–66. [Google Scholar] [CrossRef]
- Kayser, K.; Gabius, H.J.; Gabius, S. Biotinylated ligands for receptor localization: An alternative for immunohistochemistry. Zentralbl. Pathol. 1991, 137, 473–478. [Google Scholar] [PubMed]
- Maletzki, C.; Linnebacher, M.; Savai, R.; Hobohm, U. Mistletoe lectin has a shiga toxin-like structure and should be combined with other Toll-like receptor ligands in cancer therapy. Cancer Immunol. Immunother. 2013, 62, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Hong, J.H.; Kwon, H.J.; Kim, Y.; Lee, K.H.; Kim, J.B.; Song, S.K. TLR4-mediated activation of mouse macrophages by Korean mistletoe lectin-C (KML-C). Biochem. Biophys. Res. Commun. 2010, 396, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Abagyan, R.A.; Batalov, S. Do aligned sequences share the same fold? J. Mol. Biol. 1997, 273, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Horneber, M.A.; Bueschel, G.; Huber, R.; Linde, K.; Rostock, M. Mistletoe therapy in oncology. Cochrane Database Syst. Rev. 2008, 2, CD003297. [Google Scholar] [PubMed]
- Duong Van Huyen, J.P.; Bayry, J.; Delignat, S.; Gaston, A.T.; Michel, O.; Bruneval, P.; Kazatchkine, M.D.; Nicoletti, A.; Kaveri, S.V. Induction of apoptosis of endothelial cells by Viscum album: A role for anti-tumoral properties of mistletoe lectins. Mol. Med. 2002, 8, 600–606. [Google Scholar] [PubMed]
- Bussing, A.; Suzart, K.; Schweizer, K. Differences in the apoptosis-inducing properties of Viscum album L. extracts. Anticancer Drugs 1997, 8 (Suppl. S1), S9–S14. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, E. Serum levels of IL-12 and the production of IFN-gamma, IL-2 and IL-4 by peripheral blood mononuclear cells (PBMC) in cancer patients treated with Viscum album extract. Biomed. Pharmacother. 2000, 54, 305–310. [Google Scholar] [CrossRef]
- Stein, G.M.; Edlund, U.; Pfuller, U.; Bussing, A.; Schietzel, M. Influence of polysaccharides from Viscum album L. on human lymphocytes, monocytes and granulocytes in vitro. Anticancer Res. 1999, 19, 3907–3914. [Google Scholar] [PubMed]
- Braun, J.M.; Ko, H.L.; Schierholz, J.M.; Beuth, J. Standardized mistletoe extract augments immune response and down-regulates local and metastatic tumor growth in murine models. Anticancer Res. 2002, 22, 4187–4190. [Google Scholar] [PubMed]
- Duong Van Huyen, J.P.; Delignat, S.; Bayry, J.; Kazatchkine, M.D.; Bruneval, P.; Nicoletti, A.; Kaveri, S.V. Interleukin-12 is associated with the in vivo anti-tumor effect of mistletoe extracts in B16 mouse melanoma. Cancer Lett. 2006, 243, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Estko, M.; Baumgartner, S.; Urech, K.; Kunz, M.; Regueiro, U.; Heusser, P.; Weissenstein, U. Tumour cell derived effects on monocyte/macrophage polarization and function and modulatory potential of Viscum album lipophilic extract in vitro. BMC Complement. Altern. Med. 2015, 15, 130. [Google Scholar] [CrossRef] [PubMed]
- Delebinski, C.I.; Twardziok, M.; Kleinsimon, S.; Hoff, F.; Mulsow, K.; Rolff, J.; Jäger, S.; Eggert, A.; Seifert, G. A Natural Combination Extract of Viscum album L. Containing Both Triterpene Acids and Lectins Is Highly Effective against AML in Vivo. PLoS ONE 2015, 10, e0133892. [Google Scholar] [CrossRef] [PubMed]
- Holla, S.; Sharma, M.; Vani, J.; Kaveri, S.V.; Balaji, K.N.; Bayry, J. GM-CSF along with IL-4 but not alone is indispensable for the differentiation of human dendritic cells from monocytes. J. Allergy Clin. Immunol. 2014, 133, 1500–1502. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Preparation Concentration | Host Trees | Lectin Content (ng/mL) | Viscotoxin Content (µg/mL) | Method of Preparation |
|---|---|---|---|---|
| VA Qu Spez 10 mg | Quercus (Oak) | 785 ± 10% | 5 ± 5% | Fermented |
| VA Qu Frf 10 mg | Quercus (Oak) | 2391 ± 10% | 19 ± 5% | Unfermented |
| VA M Spez 10 mg | Malus (Apple) | 548 ± 10% | 4 ± 5% | Fermented |
| VA P 10 mg | Pinus (Pine) | 28 ± 10% | 6 ± 5% | Fermented |
| VA A 10 mg | Abies (Fir) | 23 ± 10% | 19 ± 5% | Fermented |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saha, C.; Das, M.; Stephen-Victor, E.; Friboulet, A.; Bayry, J.; Kaveri, S.V. Differential Effects of Viscum album Preparations on the Maturation and Activation of Human Dendritic Cells and CD4+ T Cell Responses. Molecules 2016, 21, 912. https://doi.org/10.3390/molecules21070912
Saha C, Das M, Stephen-Victor E, Friboulet A, Bayry J, Kaveri SV. Differential Effects of Viscum album Preparations on the Maturation and Activation of Human Dendritic Cells and CD4+ T Cell Responses. Molecules. 2016; 21(7):912. https://doi.org/10.3390/molecules21070912
Chicago/Turabian StyleSaha, Chaitrali, Mrinmoy Das, Emmanuel Stephen-Victor, Alain Friboulet, Jagadeesh Bayry, and Srini V. Kaveri. 2016. "Differential Effects of Viscum album Preparations on the Maturation and Activation of Human Dendritic Cells and CD4+ T Cell Responses" Molecules 21, no. 7: 912. https://doi.org/10.3390/molecules21070912
APA StyleSaha, C., Das, M., Stephen-Victor, E., Friboulet, A., Bayry, J., & Kaveri, S. V. (2016). Differential Effects of Viscum album Preparations on the Maturation and Activation of Human Dendritic Cells and CD4+ T Cell Responses. Molecules, 21(7), 912. https://doi.org/10.3390/molecules21070912