
An Aqueous Extract of Tuberaria lignosa Inhibits Cell Growth, Alters the Cell Cycle Profile, and Induces Apoptosis of NCI-H460 Tumor Cells


 ,
, 
 and
and
Abstract
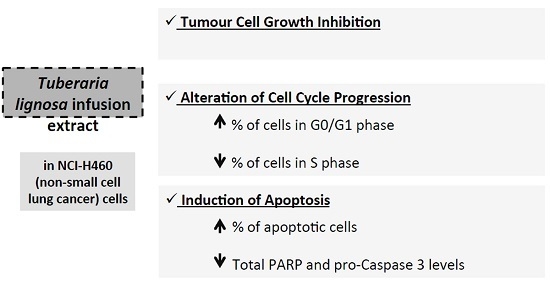
:
1. Introduction
2. Results and Discussion
2.1. T. lignosa Extracts Inhibited Tumor Cell Growth
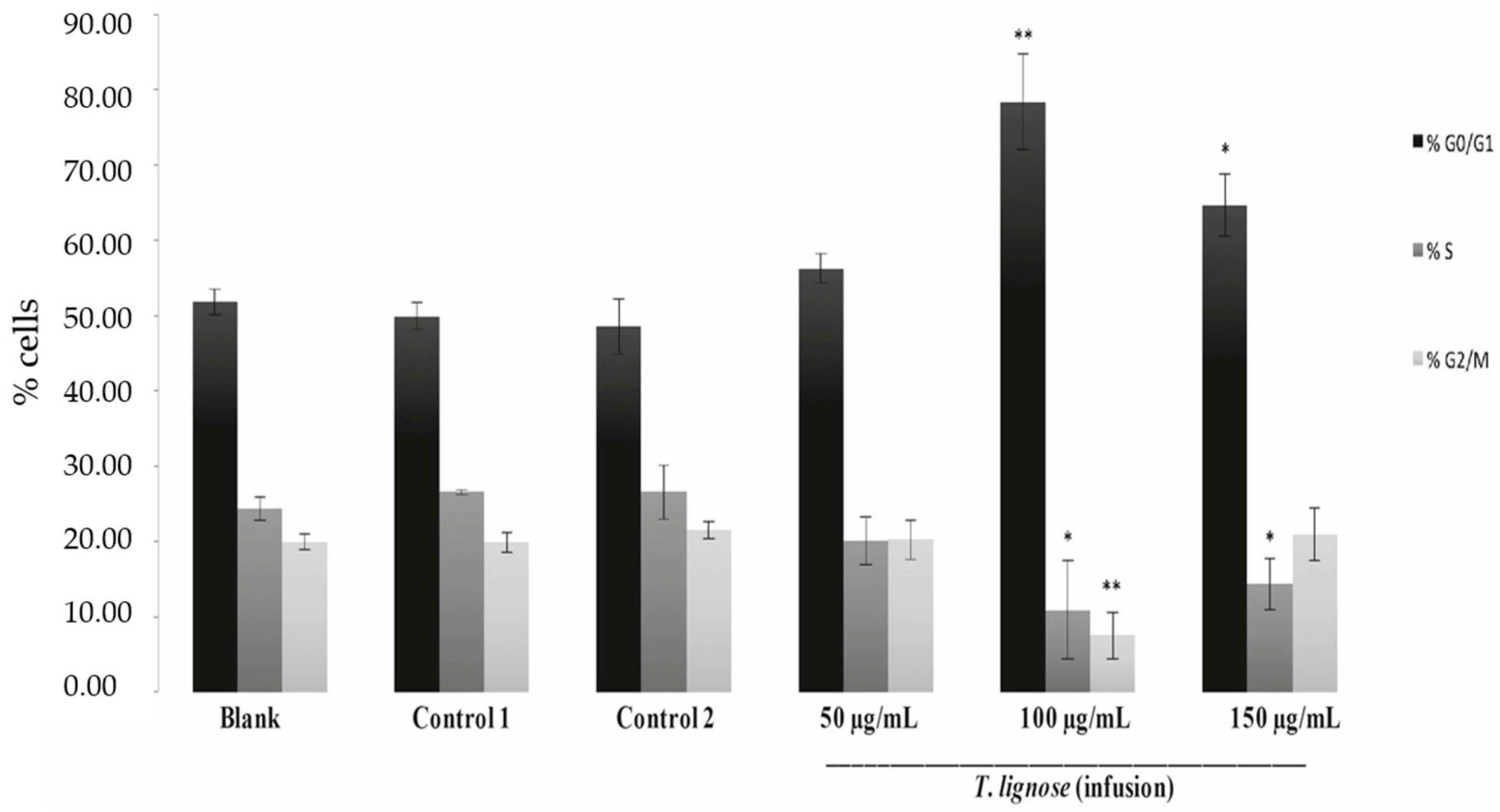
2.2. T. lignosa Infusion Extract Blocked the Cell Cycle Progression of NCI-H460 Cells
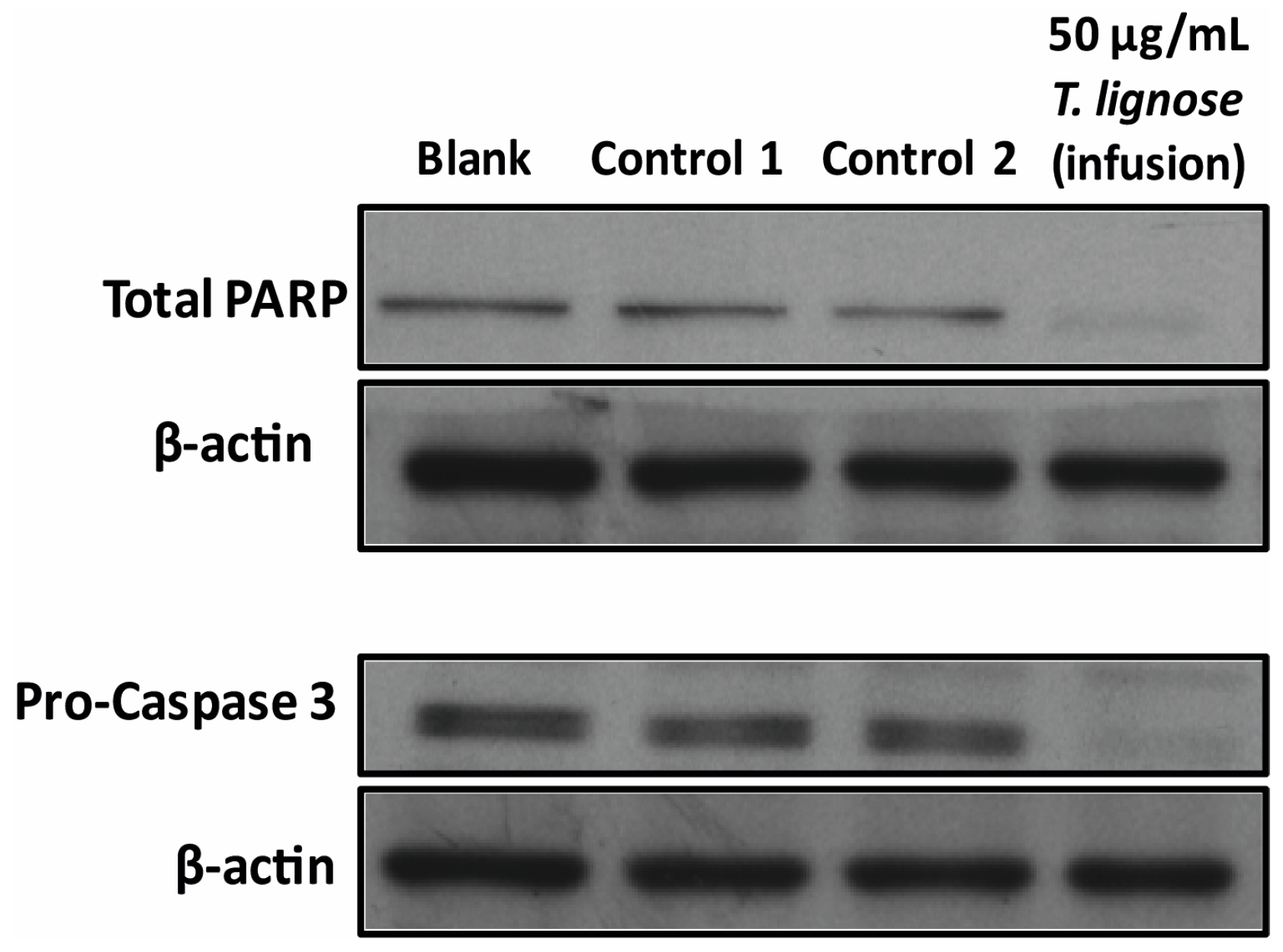
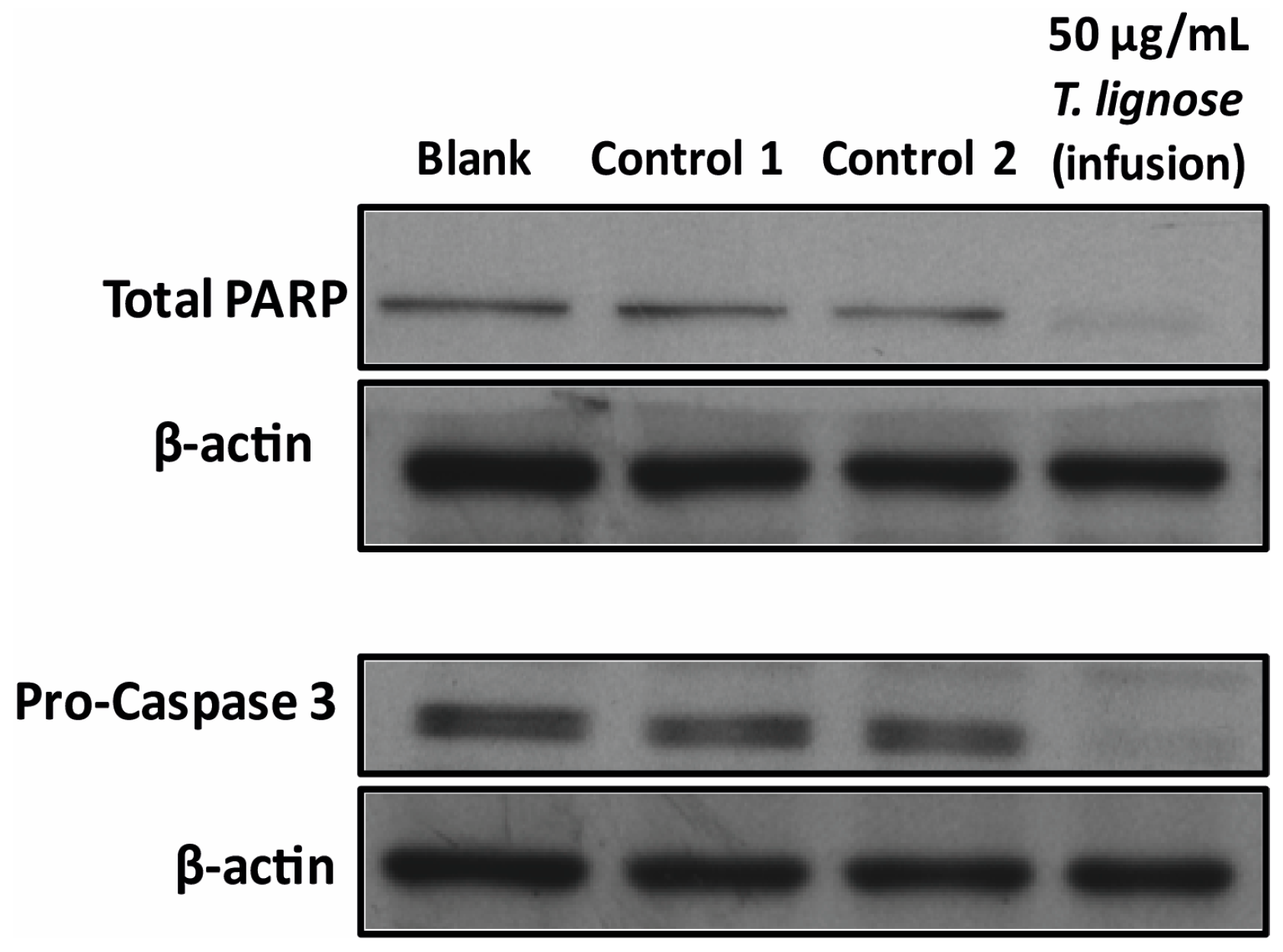
2.3. T. lignosa Infusion Extract Induced Apoptosis in NCI-H460 Cells
3. Material and Methods
3.1. Samples and Preparation of the T. lignosa Extracts
3.2. Cell Culture
3.3. Cell Growth Inhibition Assay
3.4. Preparation of Cells for Other Analyses
3.5. Analysis of Cell Cycle Profile
3.6. Analysis of Apoptosis
3.7. Analysis of Protein Expression
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cragg, G.M.; Newman, D.J. Plants as a source of anti-cancer agents. J. Ethnopharmacol. 2005, 100, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Newman, D.J. Natural products: A continuing source of novel drug leads. Biochim. Biophys. Acta 2014, 1830, 3670–3695. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.Y.; Wei, W.C.; Jian, F.Y.; Yang, N.S. Therapeutic applications of herbal medicines for cancer patients. Evid. Based Complement. Altern. Med. 2013, 2013, 302426. [Google Scholar] [CrossRef] [PubMed]
- Desai, A.G.; Qazi, G.N.; Ganju, R.K.; El-Tamer, M.; Singh, J.; Saxena, A.K.; Bedi, Y.S.; Taneja, S.C.; Bhat, H.K. Medicinal plants and cancer chemoprevention. Curr. Drug Metab. 2008, 9, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Sultana, S.; Asif, H.M.; Nazar, H.M.; Akhtar, N.; Rehman, J.U.; Rehman, R.U. Medicinal plants combating against cancer—A green anticancer approach. Asian Pac. J. Cancer Prev. 2014, 15, 4385–4394. [Google Scholar] [CrossRef] [PubMed]
- Xavier, C.P.; Pereira-Wilson, C. Medicinal plants of the genuses Salvia and Hypericum are sources of anticolon cancer compounds: Effects on PI3K/Akt and MAP kinases pathways. PharmaNutrition 2015. [Google Scholar] [CrossRef]
- Mondal, S.; Bandyopadhyay, S.; Ghosh, M.K.; Mukhopadhyay, S.; Roy, S.; Mandal, C. Natural products: Promising resources for cancer drug discovery. Anticancer Agents Med. Chem. 2012, 12, 49–75. [Google Scholar] [CrossRef] [PubMed]
- Prakash, O.; Kumar, A.; Kumar, P.; Ajeet. Anticancer potential of plants and natural products: A review. Am. J. Pharmacol. Sci. 2013, 1, 104–115. [Google Scholar]
- Sakarkar, D.M.; Deshmukh, V.N. Ethnopharmacological review of traditional medicinal plants for anticancer activity. Int. J. PharmTech. Res. 2011, 3, 298–308. [Google Scholar]
- Wirger, A.; Perabo, F.G.; Burgemeister, S.; Haase, L.; Schmidt, D.H.; Doehn, C.; Mueller, S.C.; Jocham, D. Flavopiridol, an inhibitor of cyclin-dependent kinases, induces growth inhibition and apoptosis in bladder cancer cells in vitro and in vivo. Anticancer Res. 2005, 25, 4341–4347. [Google Scholar] [PubMed]
- Hosseini, A.; Ghorbani, A. Cancer therapy with phytochemicals: Evidence from clinical studies. Avicenna J. Phytomed. 2015, 5, 84–97. [Google Scholar] [PubMed]
- Pinela, J.; Barros, L.; Duenas, M.; Carvalho, A.M.; Santos-Buelga, C.; Ferreira, I.C. Antioxidant activity, ascorbic acid, phenolic compounds and sugars of wild and commercial tuberaria lignosa samples: Effects of drying and oral preparation methods. Food Chem. 2012, 135, 1028–1035. [Google Scholar] [CrossRef] [PubMed]
- Pinela, J.; Antonio, A.L.; Barros, L.; Barreira, J.C.M.; Carvalho, A.M.; Oliveira, M.B.P.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Combined effects of gamma-irradiation and preparation method on antioxidant activity and phenolic composition of tuberaria lignosa. RSC Adv. 2015, 5, 14756–14767. [Google Scholar] [CrossRef]
- Huang, W.Y.; Cai, Y.Z.; Zhang, Y. Natural phenolic compounds from medicinal herbs and dietary plants: Potential use for cancer prevention. Nutr. Cancer 2010, 62, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Cullen, J.J.; Buettner, G.R. Ascorbic acid: Chemistry, biology and the treatment of cancer. Biochim. Biophys. Acta 2012, 1826, 443–457. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2015. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Safarzadeh, E.; Sandoghchian Shotorbani, S.; Baradaran, B. Herbal medicine as inducers of apoptosis in cancer treatment. Adv. Pharm. Bull. 2014, 4, 421–427. [Google Scholar] [PubMed]
- Xavier, C.P.; Lima, C.F.; Fernandes-Ferreira, M.; Pereira-Wilson, C. Salvia fruticosa, salvia officinalis, and rosmarinic acid induce apoptosis and inhibit proliferation of human colorectal cell lines: The role in mapk/erk pathway. Nutr. Cancer 2009, 61, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Xavier, C.P.; Lima, C.F.; Fernandes-Ferreira, M.; Pereira-Wilson, C. Hypericum androsaemum water extract inhibits proliferation in human colorectal cancer cells through effects on map kinases and PI3K/Akt pathway. Food Funct. 2012, 3, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, F.H.; Li, Y.; Wang, Z.; Kong, D. Cellular signaling perturbation by natural products. Cell. Signal. 2009, 21, 1541–1547. [Google Scholar] [CrossRef] [PubMed]
- Neergheen, V.S.; Bahorun, T.; Taylor, E.W.; Jen, L.S.; Aruoma, O.I. Targeting specific cell signaling transduction pathways by dietary and medicinal phytochemicals in cancer chemoprevention. Toxicology 2010, 278, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.R.; Kim, H.M.; Kang, J.S.; Lee, K.; Lee, S.D.; Hyun, D.H.; In, M.J.; Park, S.K.; Kim, D.C. Lipid-soluble ginseng extract induces apoptosis and G0/G1 cell cycle arrest in NCI-H460 human lung cancer cells. Plant. Foods Hum. Nutr. 2011, 66, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Kaur, G. Tinospora cordifolia induces differentiation and senescence pathways in neuroblastoma cells. Mol. Neurobiol. 2015, 52, 719–733. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Sun, L.; Zhou, J.Q.; Zhang, C.; Qin, S.; Tang, Y.; Liu, Y.; Lin, S.S.; Yuan, S.T. Marsdenia tenacissima extract induces G0/G1 cell cycle arrest in human esophageal carcinoma cells by inhibiting mitogen-activated protein kinase (MAPK) signaling pathway. Chin. J. Nat. Med. 2015, 13, 428–437. [Google Scholar] [CrossRef]
- Park, S.R.; Chen, A. Poly(adenosine diphosphate-ribose) polymerase inhibitors in cancer treatment. Hematol. Oncol. Clin. North. Am. 2012, 26, 649–670. [Google Scholar] [CrossRef] [PubMed]
- Soldatenkov, V.A.; Potaman, V.N. DNA-binding properties of poly(ADP-ribose) polymerase: A target for anticancer therapy. Curr Drug Targets 2004, 5, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Green, D.R.; Llambi, F. Cell death signaling. Cold Spring Harb. Perspect. Biol. 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Goldar, S.; Khaniani, M.S.; Derakhshan, S.M.; Baradaran, B. Molecular mechanisms of apoptosis and roles in cancer development and treatment. Asian Pac. J. Cancer Prev. 2015, 16, 2129–2144. [Google Scholar] [CrossRef] [PubMed]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef] [PubMed]
- Neves, M.P.; Cidade, H.; Pinto, M.; Silva, A.M.; Gales, L.; Damas, A.M.; Lima, R.T.; Vasconcelos, M.H.; de Sao Jose Nascimento, M. Prenylated derivatives of baicalein and 3,7-dihydroxyflavone: Synthesis and study of their effects on tumor cell lines growth, cell cycle and apoptosis. Eur. J. Med. Chem. 2011, 46, 2562–2574. [Google Scholar] [CrossRef] [PubMed]
- Vaz, J.A.; Almeida, G.M.; Ferreira, I.C.; Martins, A.; Vasconcelos, M.H. Clitocybe alexandri extract induces cell cycle arrest and apoptosis in a lung cancer cell line: Identification of phenolic acids with cytotoxic potential. Food Chem. 2012, 132, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Nunez, R. DNA measurement and cell cycle analysis by flow cytometry. Curr. Issues Mol. Biol. 2001, 3, 67–70. [Google Scholar] [PubMed]
- Lecoeur, H.; Prevost, M.C.; Gougeon, M.L. Oncosis is associated with exposure of phosphatidylserine residues on the outside layer of the plasma membrane: A reconsideration of the specificity of the annexin v/propidium iodide assay. Cytometry 2001, 44, 65–72. [Google Scholar] [CrossRef]
- Neves, M.P.; Cravo, S.; Lima, R.T.; Vasconcelos, M.H.; Nascimento, M.S.; Silva, A.M.; Pinto, M.; Cidade, H.; Correa, A.G. Solid-phase synthesis of 2′-hydroxychalcones. Effects on cell growth inhibition, cell cycle and apoptosis of human tumor cell lines. Bioorg. Med. Chem. 2012, 20, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Lima, J.M.; Sousa, D.; Lima, R.T.; Carvalho, A.M.; Ferreirad, I.C.F.R.; Vasconcelosa, M.H. Flower extracts of Filipendula ulmaria (L.) maxim inhibit the proliferation of the NCI-H460 tumour cell line. Ind. Crop. Prod. 2014, 59, 149–153. [Google Scholar] [CrossRef]
- Preto, A.; Goncalves, J.; Rebocho, A.P.; Figueiredo, J.; Meireles, A.M.; Rocha, A.S.; Vasconcelos, H.M.; Seca, H.; Seruca, R.; Soares, P.; et al. Proliferation and survival molecules implicated in the inhibition of BRAF pathway in thyroid cancer cells harbouring different genetic mutations. BMC Cancer 2009, 9, 387. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Plant voucher specimens are available from the authors.

{kind=link}
{kind=link}
{kind=link}
| Extracts | GI50 Concentrations (µg/mL) in Different Cell Lines | ||
|---|---|---|---|
| MCF-7 | HCT-15 | NCI-H460 | |
| Decoction | 135.1 ± 14.6 | 47.0 ± 2.2 | 103.7 ± 19.0 |
| Infusion | 72.9 ± 10.2 | 57.5 ± 8.2 | 43.4 ± 7.8 |
| Conditions | % Apoptosis | |
|---|---|---|
| Blank | 9.51 ± 0.9 | |
| Control (H2O) | Control 1 | 9.45 ± 0.3 |
| Control 2 | 12.7 ± 1.9 | |
| T. lignosa (infusion) | 50 µg/mL | 15.8 ± 7.1 |
| 100 µg/mL | 22.0 ± 6.1 | |
| 150 µg/mL | 34.7 ± 7.9 * | |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, J.M.; Lopes-Rodrigues, V.; Xavier, C.P.R.; Lima, M.J.; Lima, R.T.; Ferreira, I.C.F.R.; Vasconcelos, M.H. An Aqueous Extract of Tuberaria lignosa Inhibits Cell Growth, Alters the Cell Cycle Profile, and Induces Apoptosis of NCI-H460 Tumor Cells. Molecules 2016, 21, 595. https://doi.org/10.3390/molecules21050595
Pereira JM, Lopes-Rodrigues V, Xavier CPR, Lima MJ, Lima RT, Ferreira ICFR, Vasconcelos MH. An Aqueous Extract of Tuberaria lignosa Inhibits Cell Growth, Alters the Cell Cycle Profile, and Induces Apoptosis of NCI-H460 Tumor Cells. Molecules. 2016; 21(5):595. https://doi.org/10.3390/molecules21050595
Chicago/Turabian StylePereira, Joana M., Vanessa Lopes-Rodrigues, Cristina P. R. Xavier, M. João Lima, Raquel T. Lima, Isabel C. F. R. Ferreira, and M. Helena Vasconcelos. 2016. "An Aqueous Extract of Tuberaria lignosa Inhibits Cell Growth, Alters the Cell Cycle Profile, and Induces Apoptosis of NCI-H460 Tumor Cells" Molecules 21, no. 5: 595. https://doi.org/10.3390/molecules21050595
APA StylePereira, J. M., Lopes-Rodrigues, V., Xavier, C. P. R., Lima, M. J., Lima, R. T., Ferreira, I. C. F. R., & Vasconcelos, M. H. (2016). An Aqueous Extract of Tuberaria lignosa Inhibits Cell Growth, Alters the Cell Cycle Profile, and Induces Apoptosis of NCI-H460 Tumor Cells. Molecules, 21(5), 595. https://doi.org/10.3390/molecules21050595