
Chemical Basis of the Fungicidal Activity of Tobacco Extracts against Valsa mali

Abstract
:1. Introduction
2. Results and Discussion
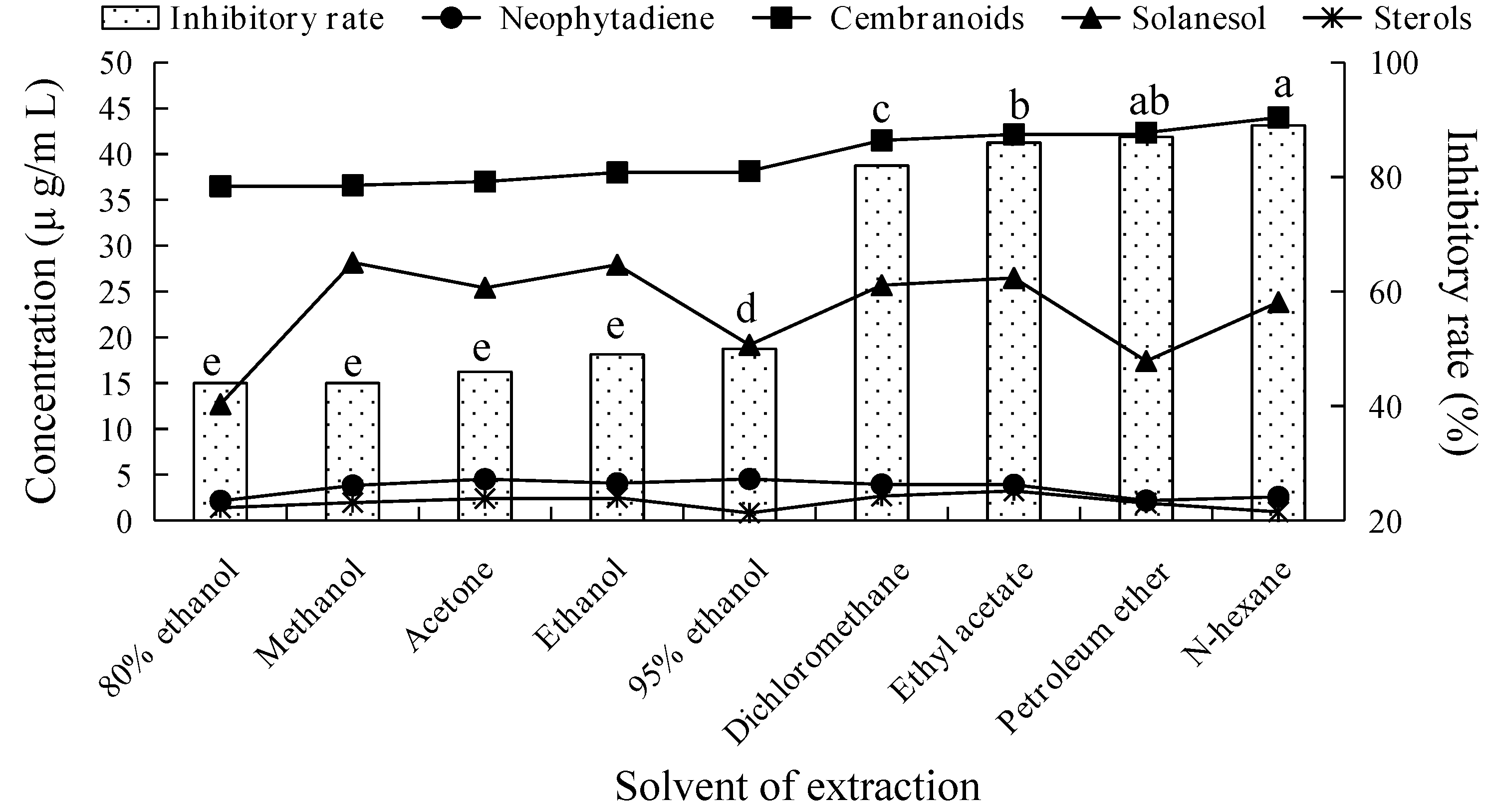
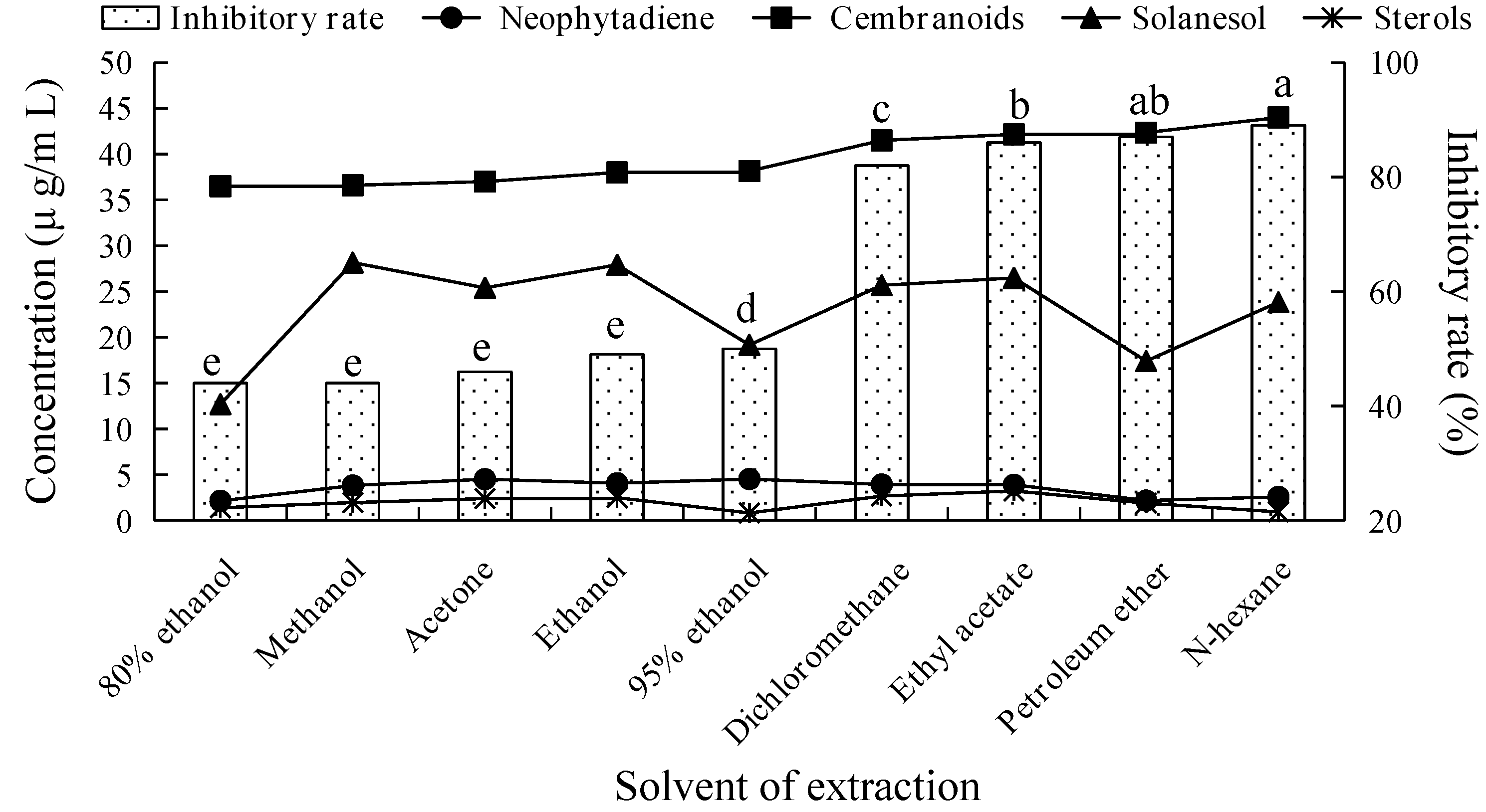
2.1. Fungicidal Activity of Tobacco Leaf Extracts Using Different Solvents and Investigation of Their Main Fungicidal Components
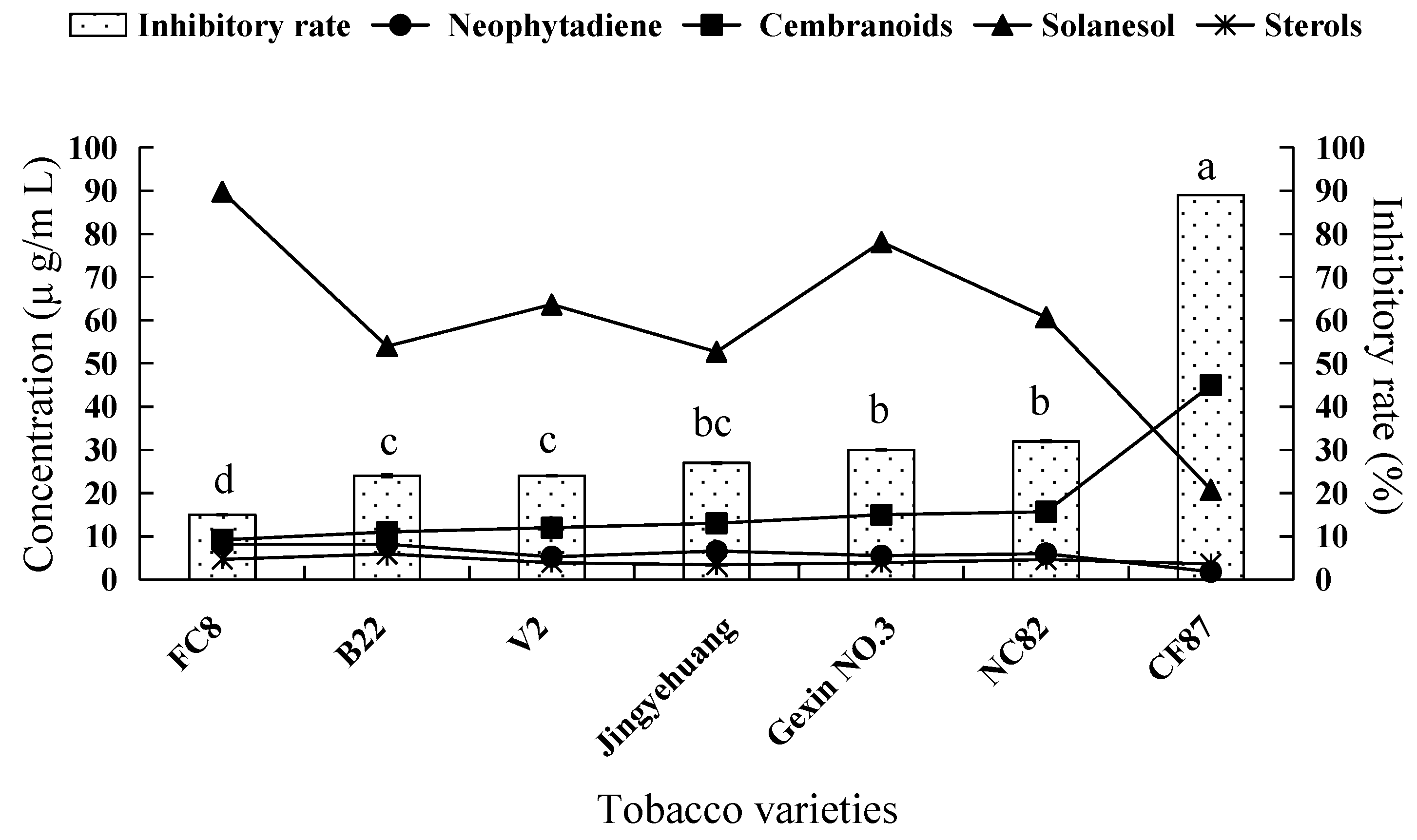
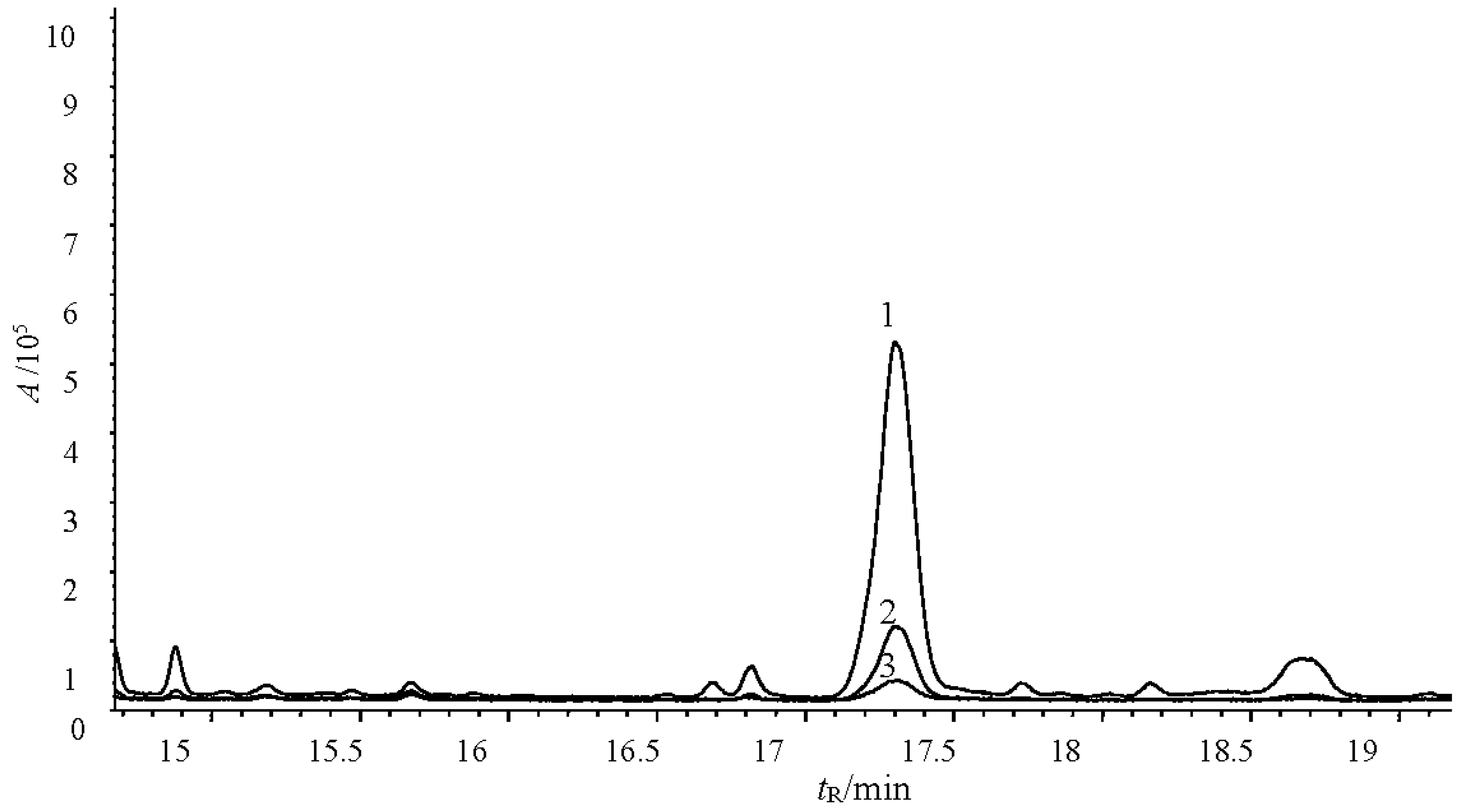
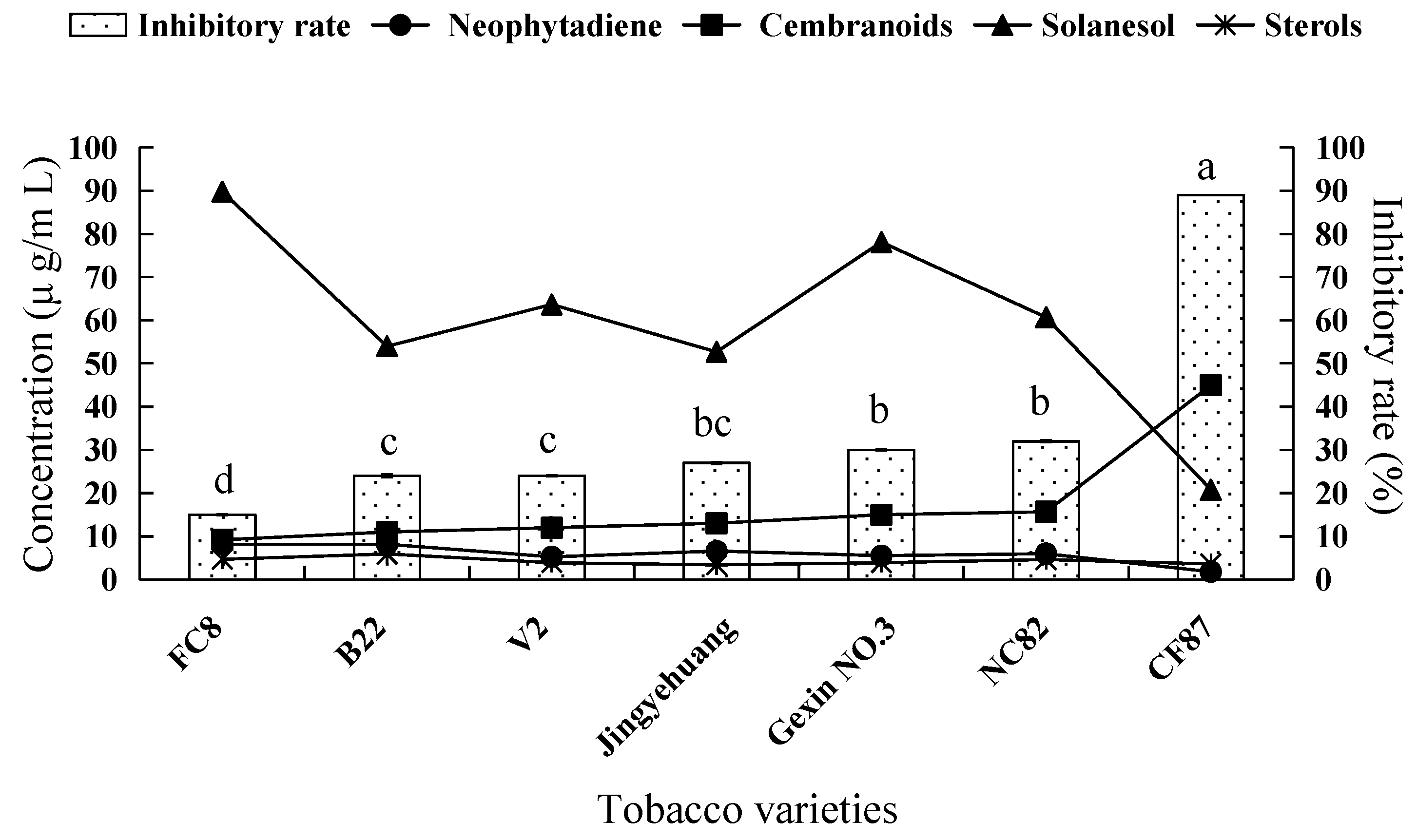
2.2. Fungicidal Activity of Tobacco Leaf Extracts from Different Varieties and the Investigation of the Main Antifungal Components
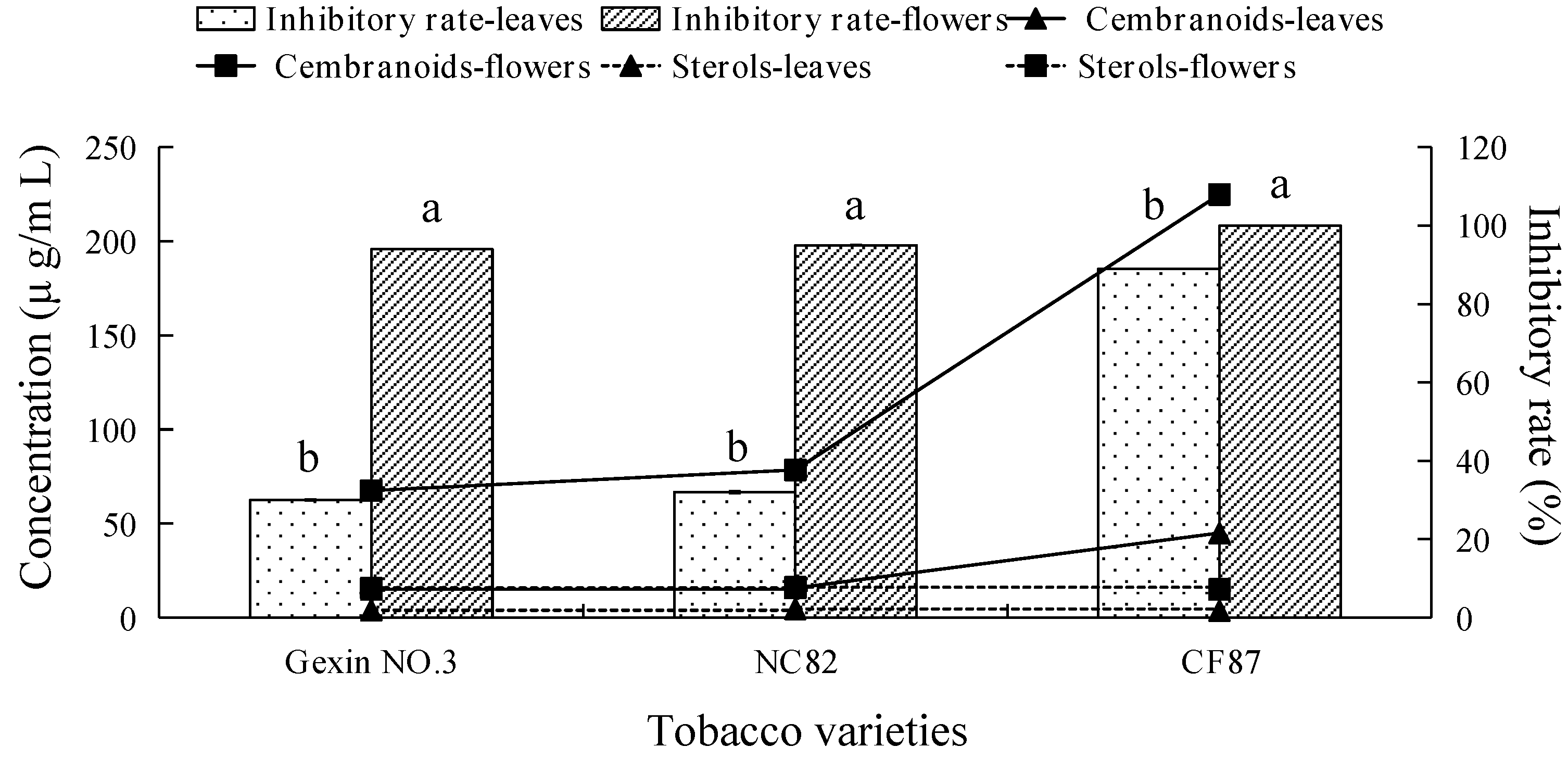
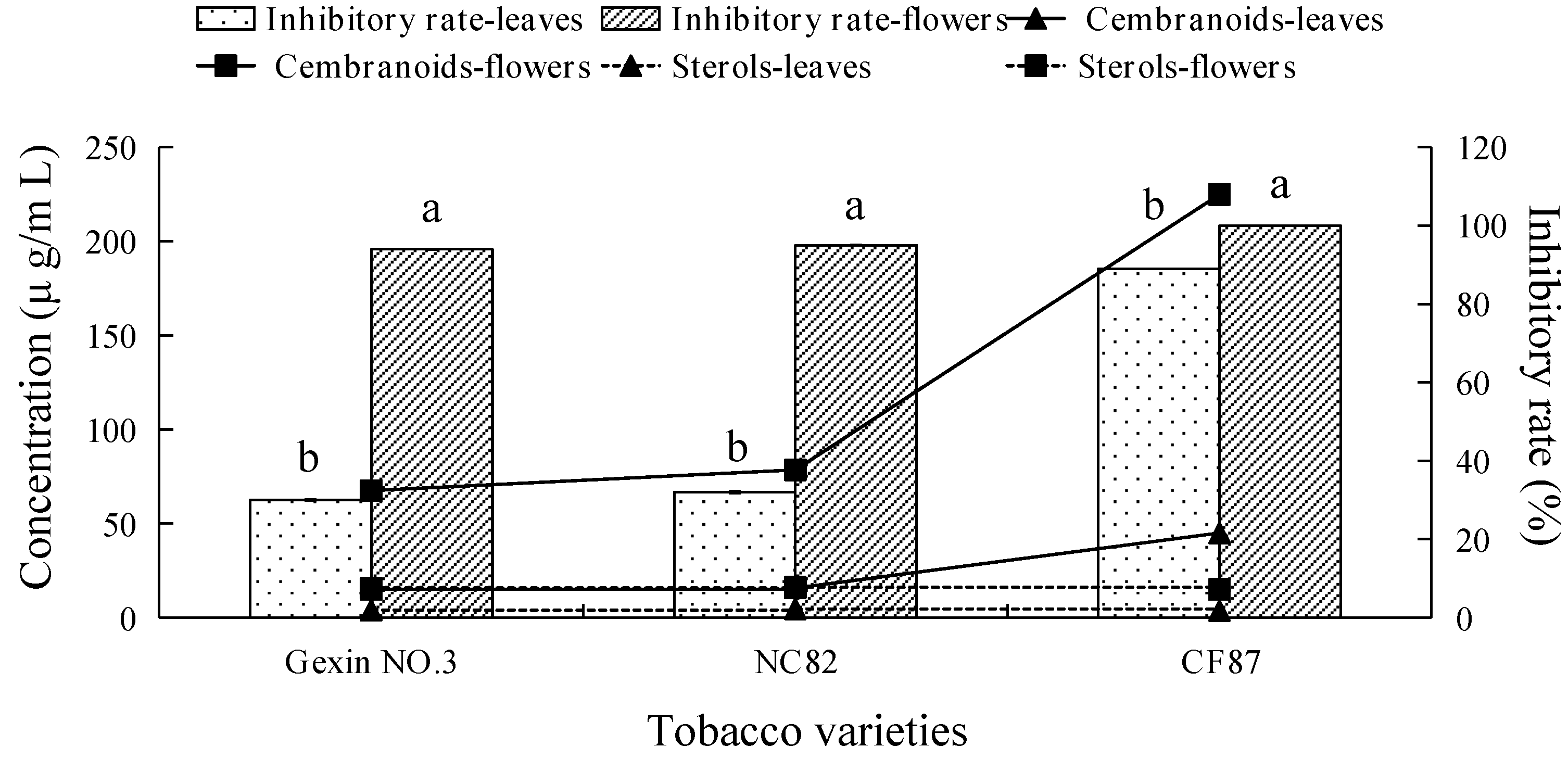
2.3. Fungicidal Activity and Main Antifungal Components of Tobacco Leaf and Flower Extracts
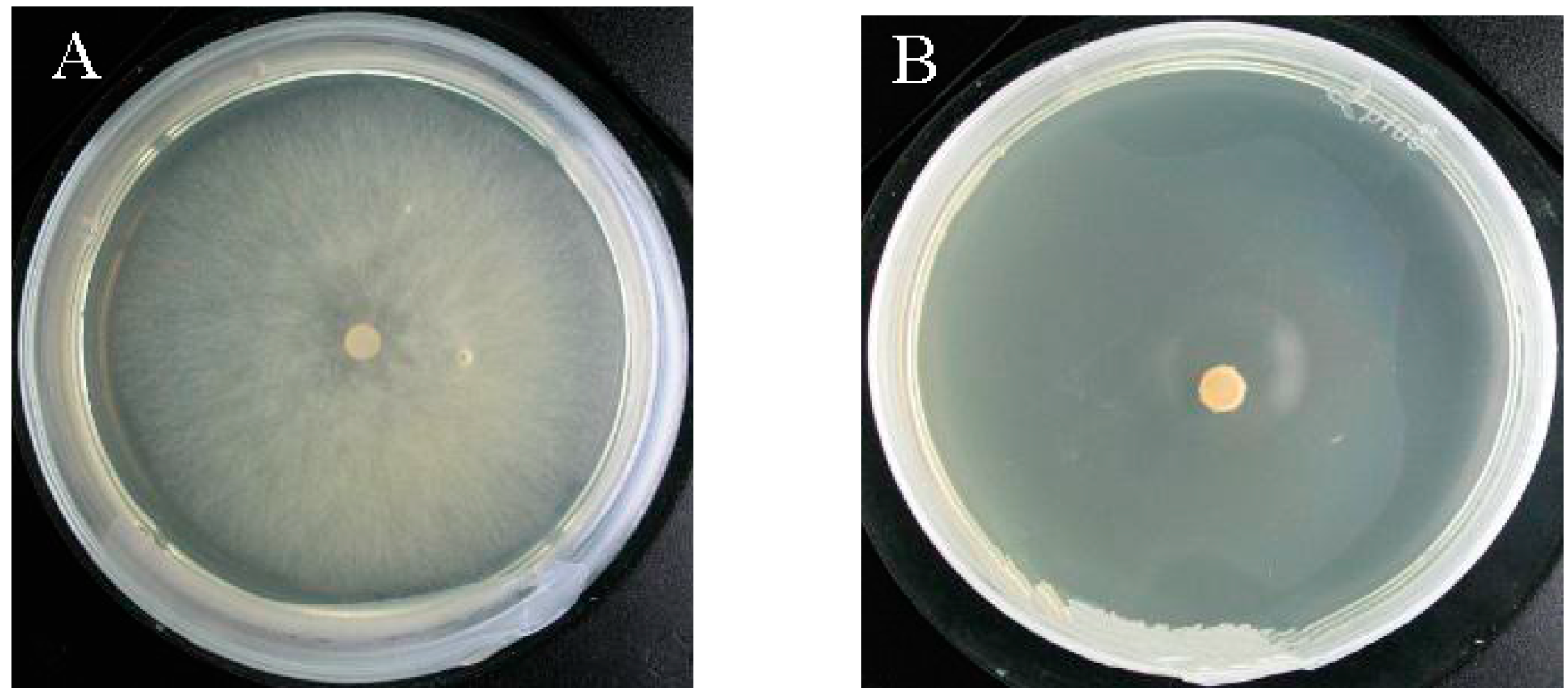
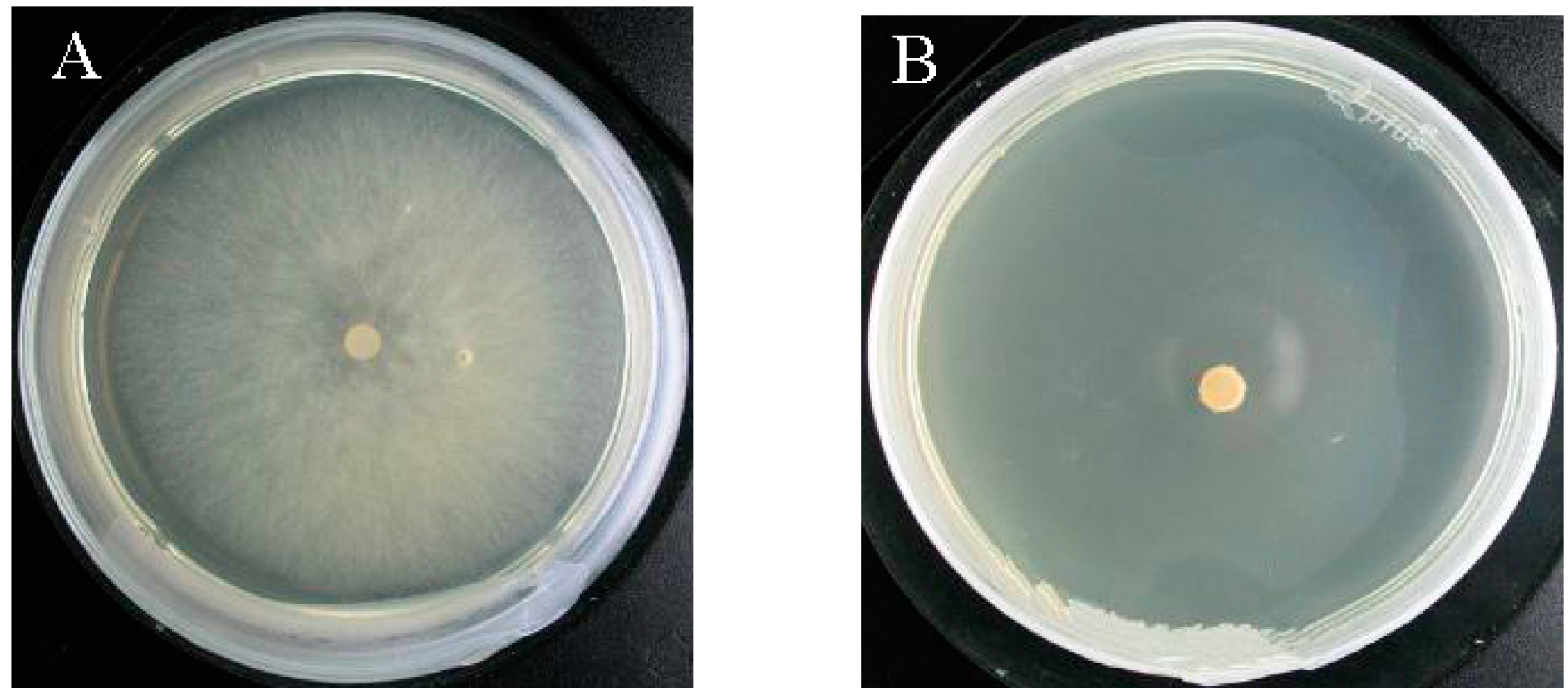
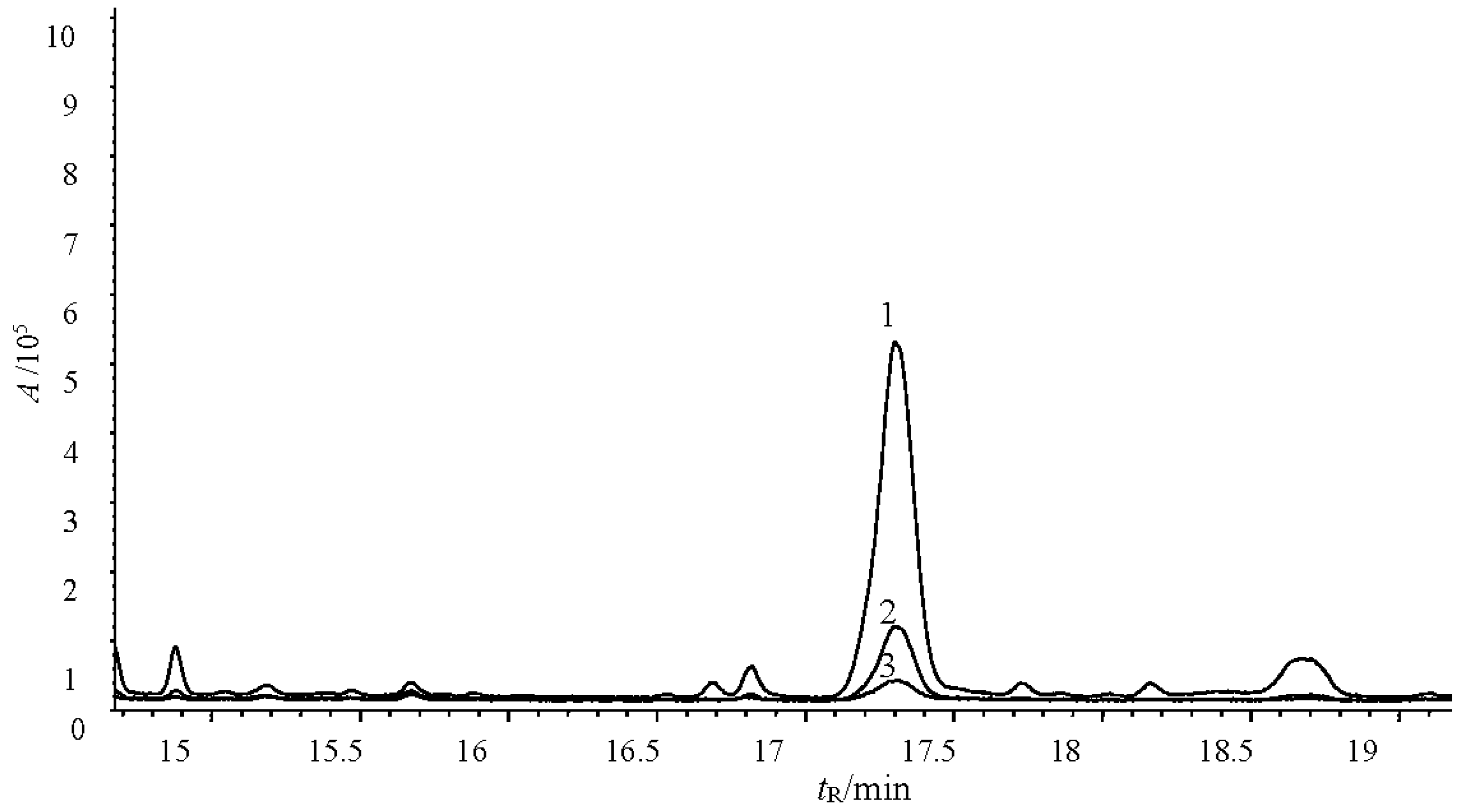
2.4. Verification of the Fungicidal Activity of Tobacco Cembranoids Extract on V. mali
2.5. Preliminary Mechanic Exploration of Tobacco Cembranoids Antifungal Activity
2.5.1. Effect of Tobacco Cembranoids on the Content of Ergosterol in the V. mali Cell Membrane
2.5.2. Ultrastructural Observation of V. mali by Scanning Electron Microscopy
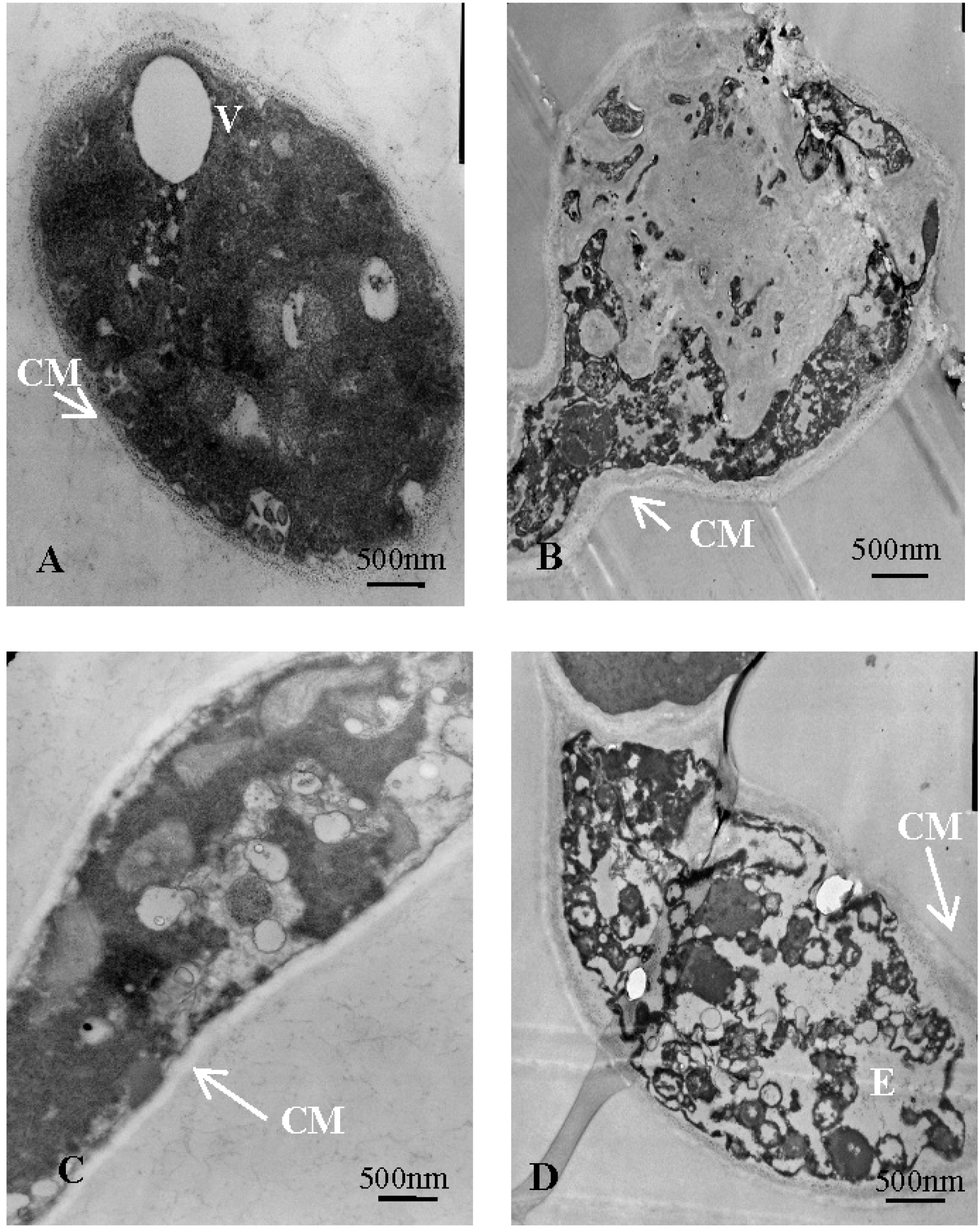
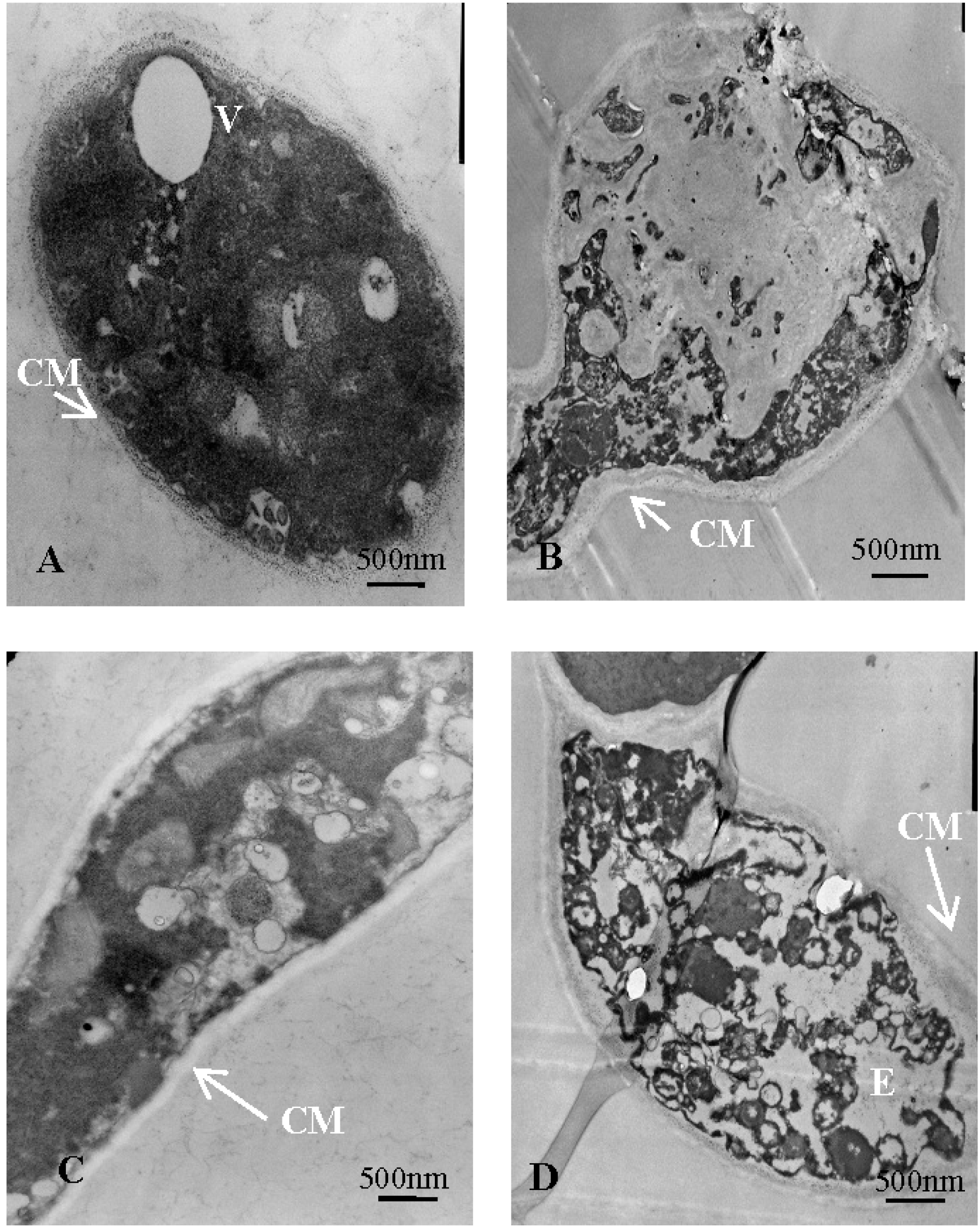
2.5.3. Ultrastructural Observation of V. mali with Transmission Electron Microscopy
3. Materials and Methods
3.1. Materials
3.2. Tobacco Field Cultivation
3.3. Preparation of Tobacco Samples
3.4. Chemical Compounds Extraction and Isolation
3.5. Preparation of Tobacco Cembranoids and Content Determination
3.6. Assessment of Fungicidal Activity
3.7. Determination of the Fungicide Activity of Tobacco Cembranoids Extracts
3.8. Observation of V. mali Ultrastructure
3.8.1. Observation of V. mali Ultrastructure by Scanning Electron Microscopy
3.8.2. Observation of V. mali Ultrastructure by Transmission Electron Microscopy
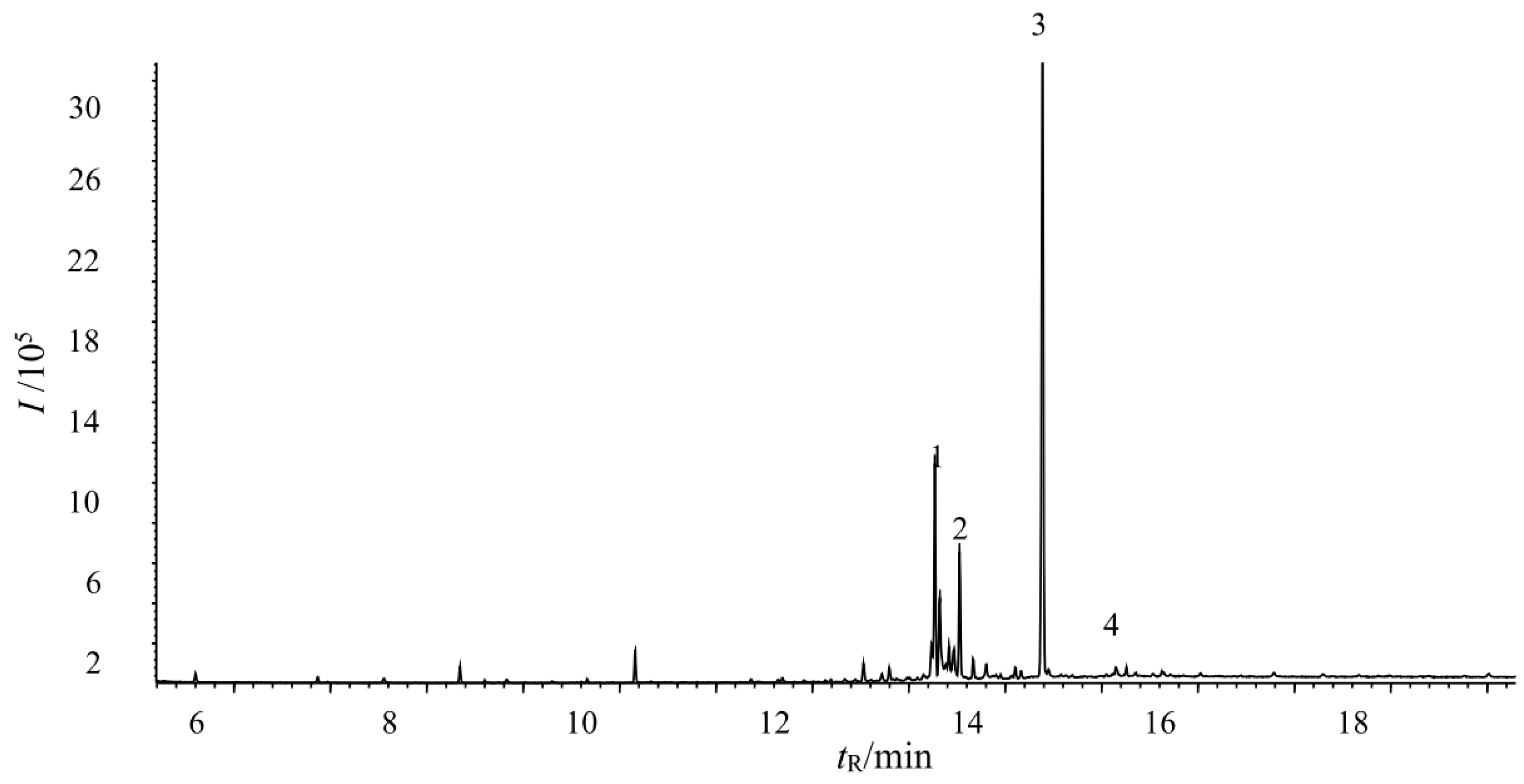
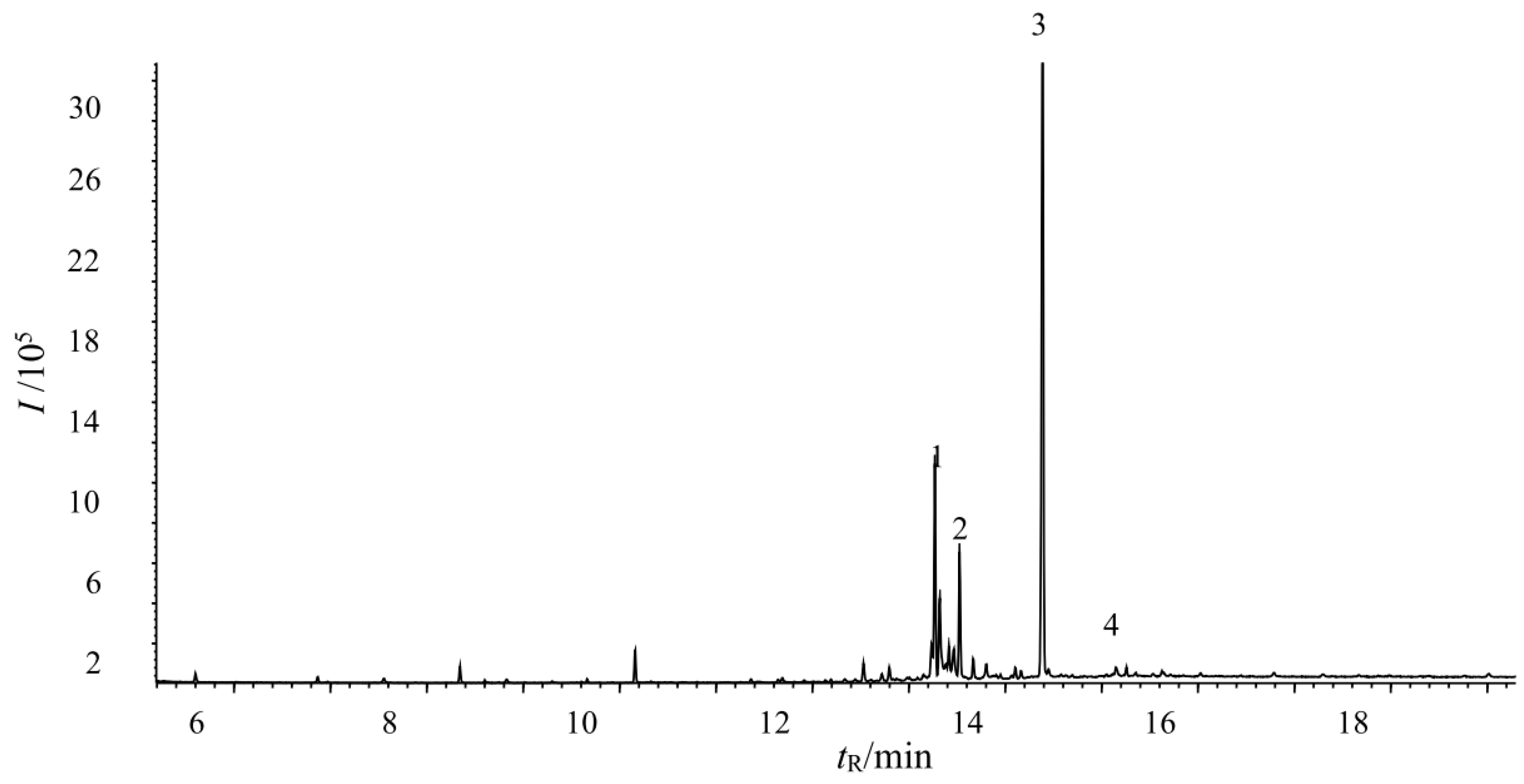
3.9. Determination of the Main Fungicide Components in the Tobacco Extracts
3.10. Determination of the Ergosterol Content in the Cell Membrane of V. mali
3.11. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| MIC | minimal inhibition concentration |
| MFC | minimal fungicidal concentration |
| EC50 | half-maximal effective concentration |
| GC/MS | gas chromatography/mass spectrometry |
| HPMS | high pressure mass spectrometry |
| HPLC | high performance liquid chromatography |
| CM | cytoplasmic membrane |
| V | vacuole |
| E | cavum |
References
- Tomizawa, M.; Casida, J.E. Selective toxicity of neonicotinoids attributable to specificity of insect and mammalian nicotinic receptors. Annu. Rev. Entomol. 2003, 48, 339–364. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Ni, J.; Huang, W. Comparison on bioactivities of solanesol extracted from tobacco leaves by different methods. J. Anhui Agric. Sci. 2008, 36, 6356–6359. [Google Scholar]
- Cen, X.; Ding, W.; Ding, J. Preliminary studies on the activity of crude extracts from Nicotiana tabacum stems against Tetranychus cinnabar inus. Plant Prot. 2009, 35, 156–158. [Google Scholar]
- Hu, S. Research of the nicotine to disinfect pathogenic bacteria affects. Biotechnology 2009, 19, 73–75. [Google Scholar]
- Li, J.; Zou, Y.; Ren, X. Humble opinion about modern apple industry in China. J. Fruit Sci. 2008, 25, 378–381. [Google Scholar]
- Cao, K.; Guo, L.; Li, B.; Sun, G.; Chen, H. Investigations on the occurrence and control of apple canker in China. Plant Prot. 2009, 35, 114–116. [Google Scholar]
- Wang, S.; Li, Y.; Zhang, H. Research advances on antibacterial plants. Biotechnol. Bull. 2011, 8, 26–29. [Google Scholar]
- Cui, H.; Zhang, S.T.; Yang, H.J.; Ji, H.; Wang, X.J. Gene expression profile analysis of tobacco leaf trichomes. BMC Plant Biol. 2011, 11, 76. [Google Scholar] [CrossRef] [PubMed]
- Severson, R.F.; Johnson, A.W.; Jackson, D.M. Cuticular constituents of tobacco: Factors affecting their production and their role in insect and disease resistance and smoke quality. Rec. Adv. Tob. Sci. 1985, 11, 105–173. [Google Scholar]
- Wagner, G.J. Secreting glandular trichomes: More than just hairs. Plant Physiol. 1991, 96, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, B.S.; Nielsen, M.T.; Severson, R.F. Biorationals from Nicotiana protect cucumbers against Colletotrichum lagenarium (Pass.) Ell. & Halst disease development. J. Chem. Ecol. 1995, 21, 221–231. [Google Scholar] [PubMed]
- Ferchmin, P.A.; Pagan, O.R.; Ulrich, H.; Szeto, A.C.; Hann, R.M.; Eterovic, V.A. Actions of octocoral and tobacco cembranoids on nicotinic receptors. Toxicon 2009, 54, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Aqil, F.; Zahin, M.; El Sayed, K.A.; Ahmad, I.; Orabi, K.Y.; Arif, J.M. Antimicrobial, antioxidant, and antimutagenic activities of selected marine natural products and tobacco cembranoids. Drug Chem. Toxicol. 2011, 34, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Du, Y.; Hou, X.; Li, D.; Ren, X.; Dong, W.; Zhao, W.; Zhang, Z. Inhibitory effects of tobacco extracts on eleven phytopathogenic fungi. Nat. Prod. Res. Dev. 2015, 27, 470–474. [Google Scholar]
- Yan, N.; Du, Y.; Liu, X.; Zhang, H.; Liu, Y.; Zhang, P.; Gong, D.; Zhang, Z. Chemical structures, biosynthesis, bioactivities, biocatalysis and semisynthesis of tobacco cembranoids: An overview. Ind. Crops Prod. 2016, 83, 66–80. [Google Scholar] [CrossRef]
- Yan, N.; Du, Y.; Liu, X.; Zhang, H.; Liu, Y.; Shi, J.; Xue, S.J.; Zhang, Z. Analyses of effects of α-cembratrien-diol on cell morphology and transcriptome of Valsa mali var. mali. Food Chem. 2017, 214, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.; Eterovic, V.A.; Ferchmin, P.A.; Rios-Olivares, E.; Wang, D.; Nath, A.; Rodriguez, J.W. Modulation of HIV-1 replication, inflammation, and neurotoxicity by a tobacco cembranoid 4R: Therapeutic implications for HIV-associated neurocognitive disorders. J. NeuroVirol. 2010, 16, 73. [Google Scholar]
- El Sayed, K.A.; Shah, G.; Sylvester, P.W. Anticancer Tobacco Cembranoids. Patent No. US7977384B1, 12 July 2011. [Google Scholar]
- El Sayed, K.A.; Sylvester, P.W. Biocatalytic and semisynthetic studies of the anticancer tobacco cembranoids. Expert Opin. Investig. Drugs 2007, 16, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Nacoulma, A.P.; Megalizzi, V.; Pottier, L.R.; de Lorenzi, M.; Thoret, S.; Dubois, J.; Vandeputte, O.M.; Duez, P.; Vereecke, D.; El Jaziri, M. Potent antiproliferative cembrenoids accumulate in tobacco upon infection with Rhodococcus fascians and trigger unusual microtubule dynamics in human glioblastoma cells. PLoS ONE 2013, 8, e77529. [Google Scholar] [CrossRef] [PubMed]
- Zubair, M.S.; Anam, S.; Al-Footy, K.O.; Abdel-Lateef, A.; Alarif, W.M. Cembranoid diterpenes as antitumour: Molecular docking study to several protein receptor targets. In Proceedings of the 3rd International Conference on Computation for Science and Technology, Bali, Indonesia, 23–25 September 2014; Tjahjono, D.H., Ed.; Atlantis Press: Paris, France, 2014; Volume 2, pp. 121–125. [Google Scholar]
- Wang, Y.; Ji, L.; Liu, Y.; Zhang, Y.; Yi, X. Mechanism of extracts obtained from Polygonum cuspidatum to suppress Valsa mali. Chin. J. Biol. Control 2015, 31, 148–156. [Google Scholar]
- Yang, Y.; Wei, H.; Zhao, Z.; Liu, P.; Wu, Y. Effect of polyhydroxy dinaphthaldehyde extracts from plant against Valsa mali in apple tree. Acta Phytopathol. Sin. 2011, 41, 421–427. [Google Scholar]
- Han, J.; Zhang, Z.; Liu, H.; Wang, X.; Liu, H.; Mao, G. Research advance in tobacco glandular trichomes and their secretion substance cembranoids. Acta Tab. Sin. 2013, 19, 118–124. [Google Scholar]
- Nielsen, M.T.; Severson, R.F. Variation of flavor components on leaf surfaces of tobacco genotypes differing in trichome density. J. Agric. Food Chem. 1990, 38, 467–471. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, Y.; Ai, Q. Inhibitory effect and antimicrobial mechanism of pyrolin on Monilinia fructicola in peach. Sci. Agric. Sin. 2009, 42, 2784–2792. [Google Scholar]
- Dos Santos, E.C.G.; Donnici, C.L.; da Silva Camargos, E.R.; de Rezende, A.A.; de Aguiar Andrade, E.H.; Soares, L.A.L.; de Macêdo Farias, L.; de Carvalho, M.A.R.; das Graças Almeida, M. Effects of Copaifera duckei Dwyer oleoresin on the cell wall and cell division of Bacillus cereus. J. Med. Microbiol. 2013, 62, 1032–1037. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; He, J.; Shen, H. The inhibition and antifungal mechanism of cocklebur extract to Botrytiscinerea. Acta Pratacult. Sin. 2008, 17, 99–104. [Google Scholar]
- Xie, J.; Dong, G.; Liu, Z. Preparation methods of microbiological sample by SEM. J. Chin. Electron Microsc. Soc. 2005, 24, 440. [Google Scholar]
- Wang, H.Y.; Zhao, M.M.; Yang, B.; Jiang, Y.M.; Rao, G.H. Identification of polyphenols in tobacco leaf and their antioxidant and antimicrobial activities. Food Chem. 2008, 107, 1399–1406. [Google Scholar] [CrossRef]
- Liu, C.C.; Zhang, H.B.; Du, Y.M.; Hou, X.D.; Li, D.D.; Yan, N. The simultaneous extraction and saponification of tobacco solanesol using ultra performance liquid chromatography. Chin. Tob. Sci. 2015, 36, 79–84. [Google Scholar]
- Severson, R.F.; Ellington, J.J.; Arrendale, R.F.; Snook, M.E. Quantitative gas chromatographic method for the analysis of aliphatic hydrocarbons, terpenes, fatty alcohols, fatty acids and sterols in tobacco. J. Chromatogr. 1978, 160, 155–168. [Google Scholar] [CrossRef]
- Gao, F.; Wu, Y.; Wang, M. Action of the metabolite produced by Streptomyces cacaoi strain 182–2 on Alternaria alternate. Sci. Agric. Sin. 2013, 46, 4933–4940. [Google Scholar]
- Sample Availability: Samples of the compounds are not available from the authors.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Regression Equation | R2 | p-Value | MIC (μg/mL) | MFC (μg/mL) | EC50 (μg/mL) |
|---|---|---|---|---|---|---|
| Tobacco cembranoids extract | y = 1.5923x + 3.2114 | 0.97 | 0.003 | 2.02 | 80 | 13.18 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, C.B.o.t.F.A.o.T.E.a.V.m.S.; Du, Y.; Hou, X.; Yan, N.; Dong, W.; Mao, X.; Zhang, Z. Chemical Basis of the Fungicidal Activity of Tobacco Extracts against Valsa mali. Molecules 2016, 21, 1743. https://doi.org/10.3390/molecules21121743
Duan CBotFAoTEaVmS, Du Y, Hou X, Yan N, Dong W, Mao X, Zhang Z. Chemical Basis of the Fungicidal Activity of Tobacco Extracts against Valsa mali. Molecules. 2016; 21(12):1743. https://doi.org/10.3390/molecules21121743
Chicago/Turabian StyleDuan, Chemical Basis of the Fungicidal Activity of Tobacco Extracts against Valsa mali Suzhen, Yongmei Du, Xiaodong Hou, Ning Yan, Weijie Dong, Xinxin Mao, and Zhongfeng Zhang. 2016. "Chemical Basis of the Fungicidal Activity of Tobacco Extracts against Valsa mali" Molecules 21, no. 12: 1743. https://doi.org/10.3390/molecules21121743
APA StyleDuan, C. B. o. t. F. A. o. T. E. a. V. m. S., Du, Y., Hou, X., Yan, N., Dong, W., Mao, X., & Zhang, Z. (2016). Chemical Basis of the Fungicidal Activity of Tobacco Extracts against Valsa mali. Molecules, 21(12), 1743. https://doi.org/10.3390/molecules21121743