
Unfolding/Refolding Study on Collagen from Sea Cucumber Based on 2D Fourier Transform Infrared Spectroscopy


Abstract
:1. Introduction
2. Results and Discussion
2.1. Amino Acid Compositions of ICFs and PSCs
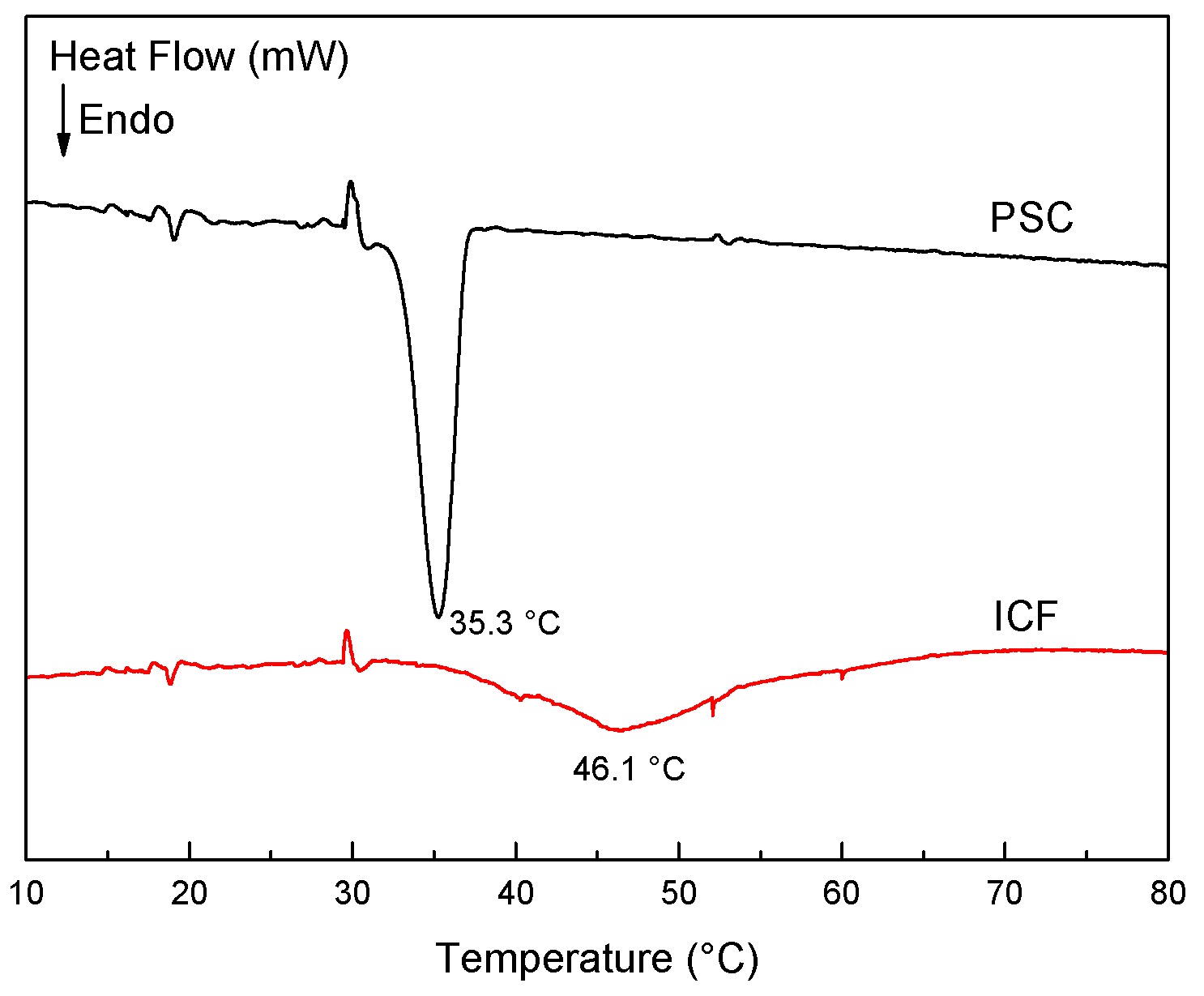
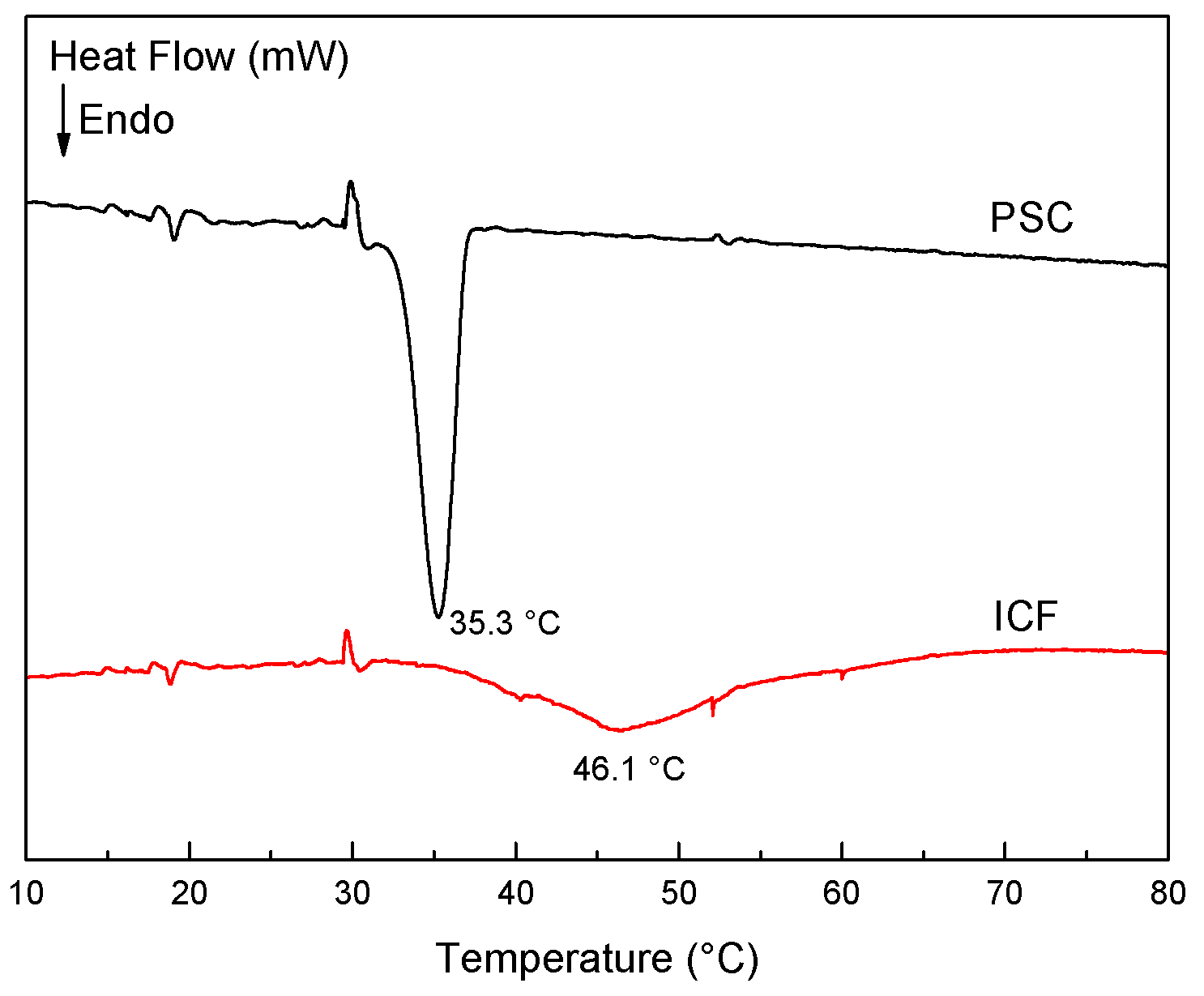
2.2. Denaturation Temperature of Sea Cucumber Collagen
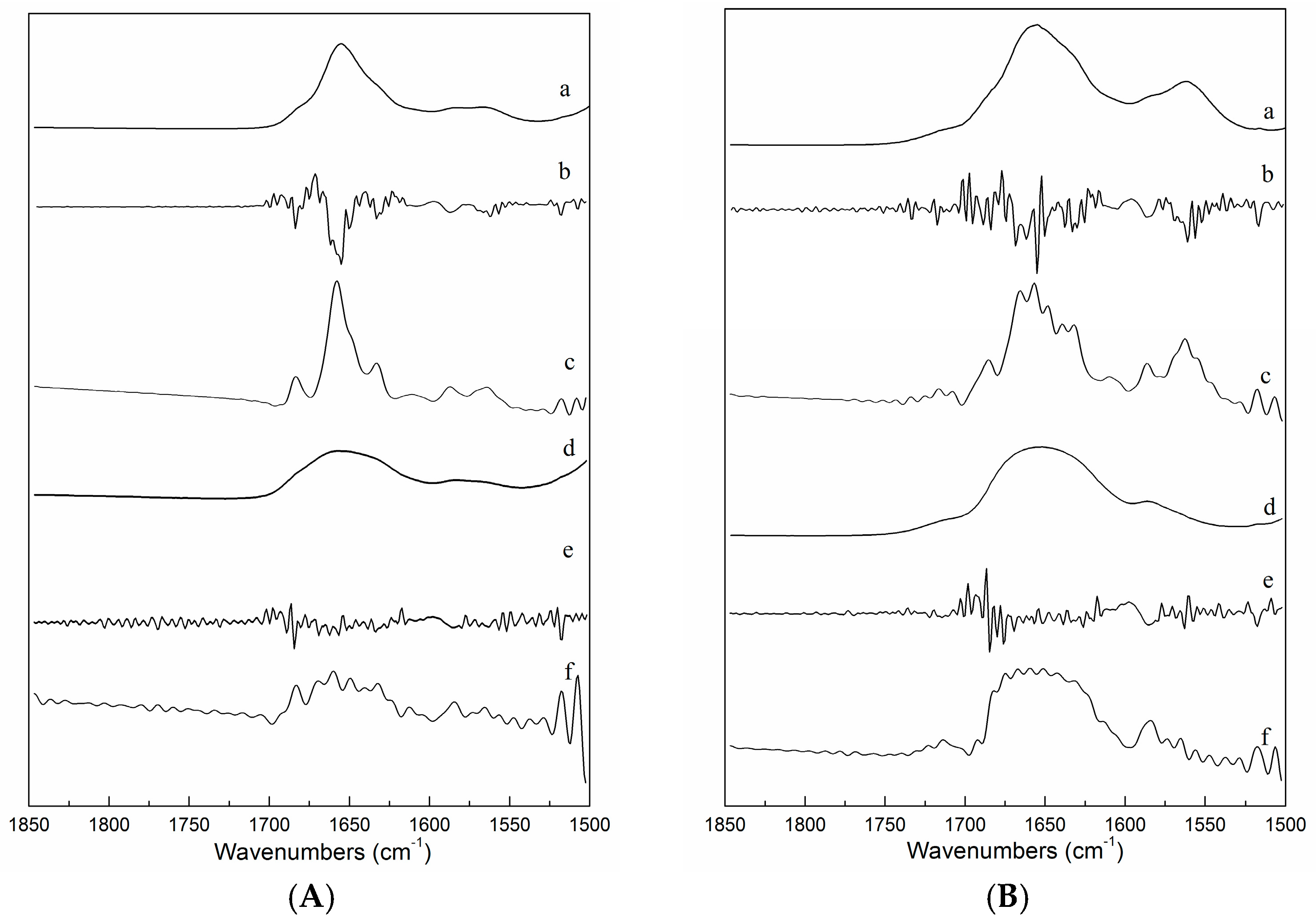
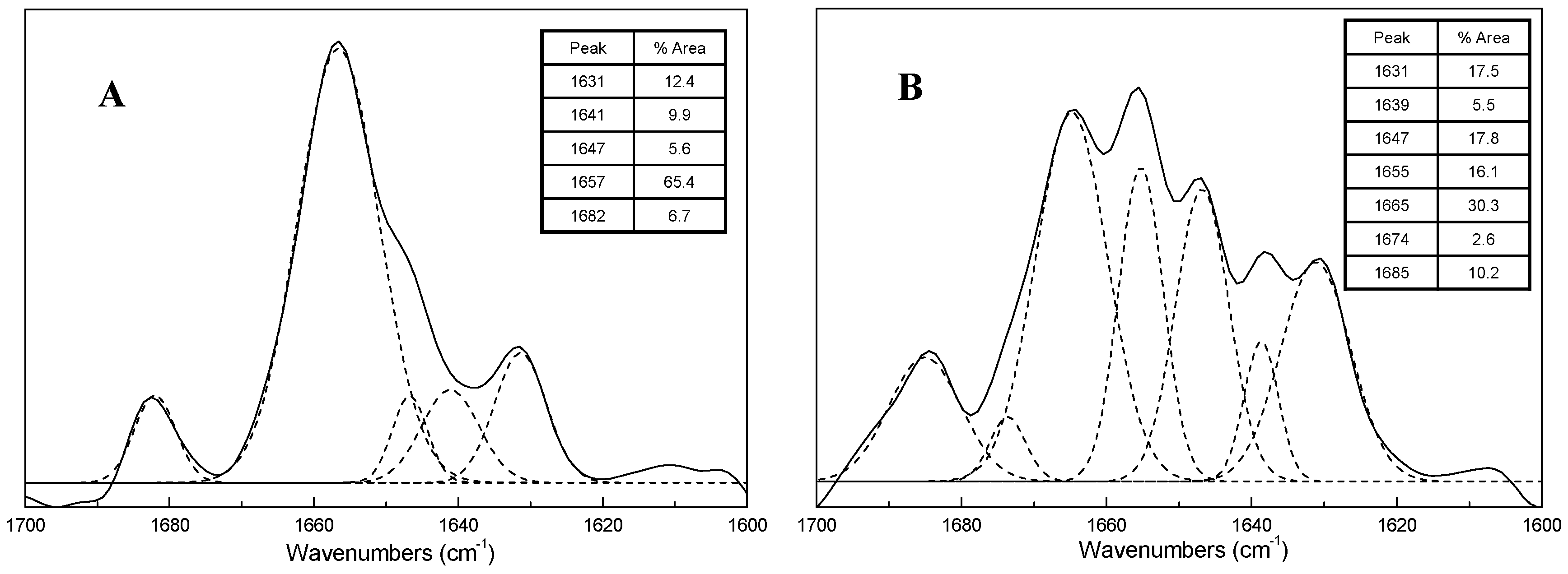
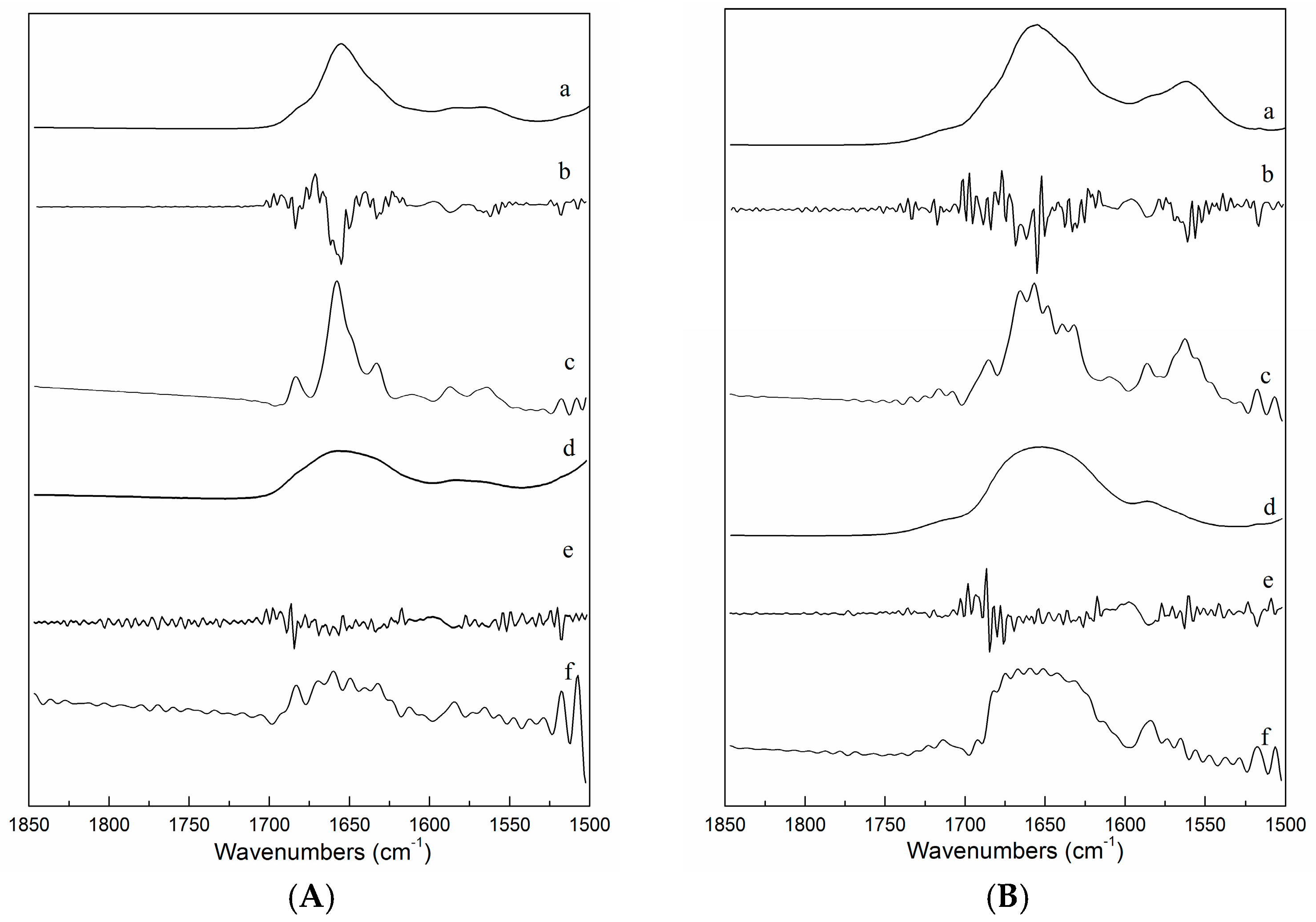
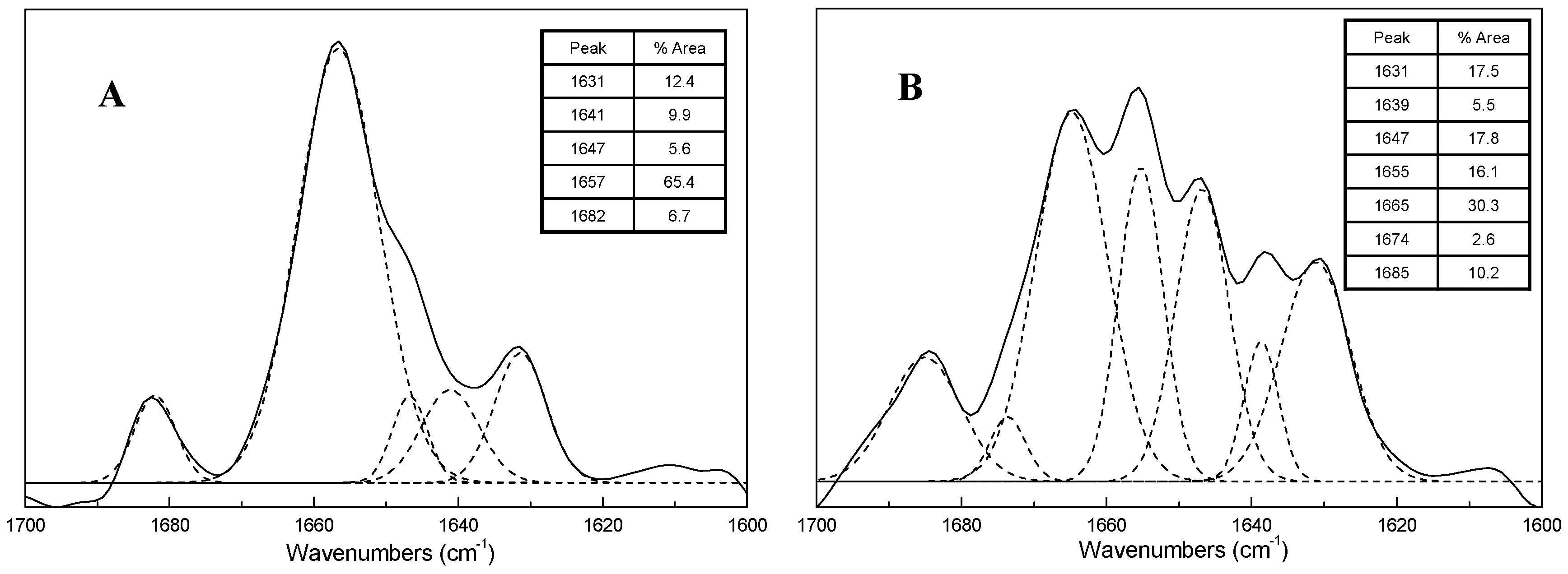
2.3. FTIR Spectra of ICFs and PSCs
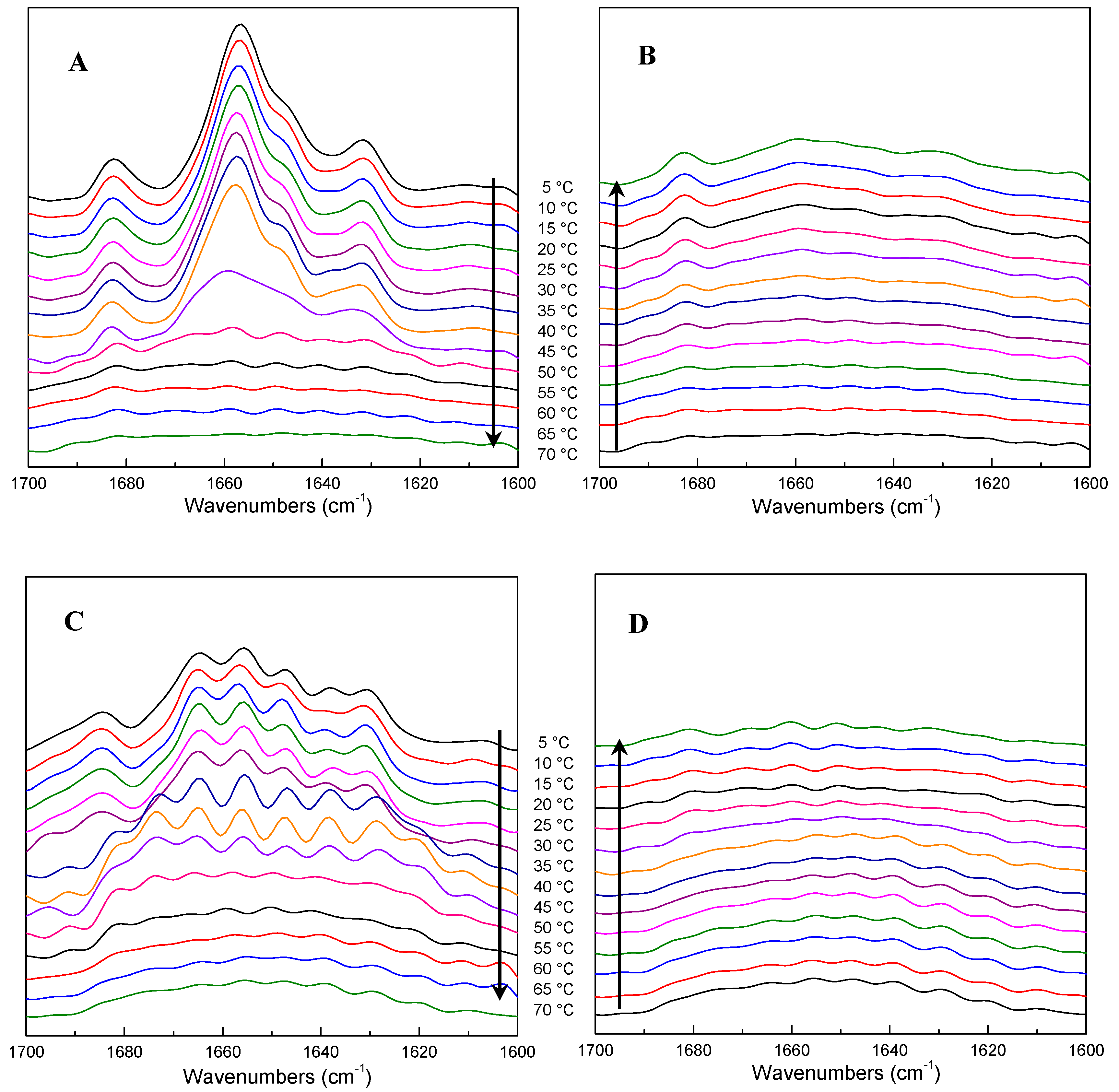
2.4. FTIR Spectra as a Function of Temperature during Heating and Cooling of ICFs or PSCs
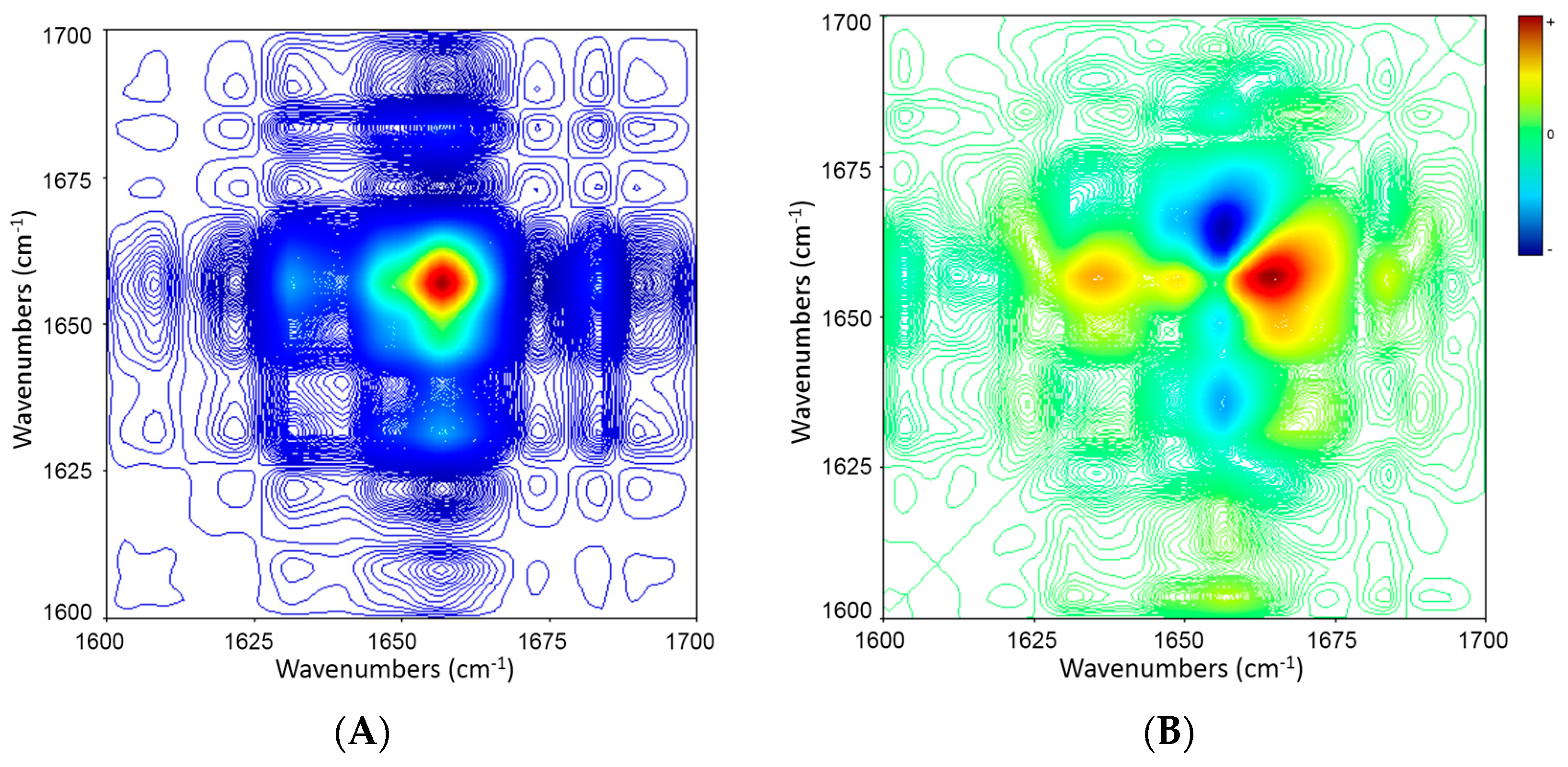
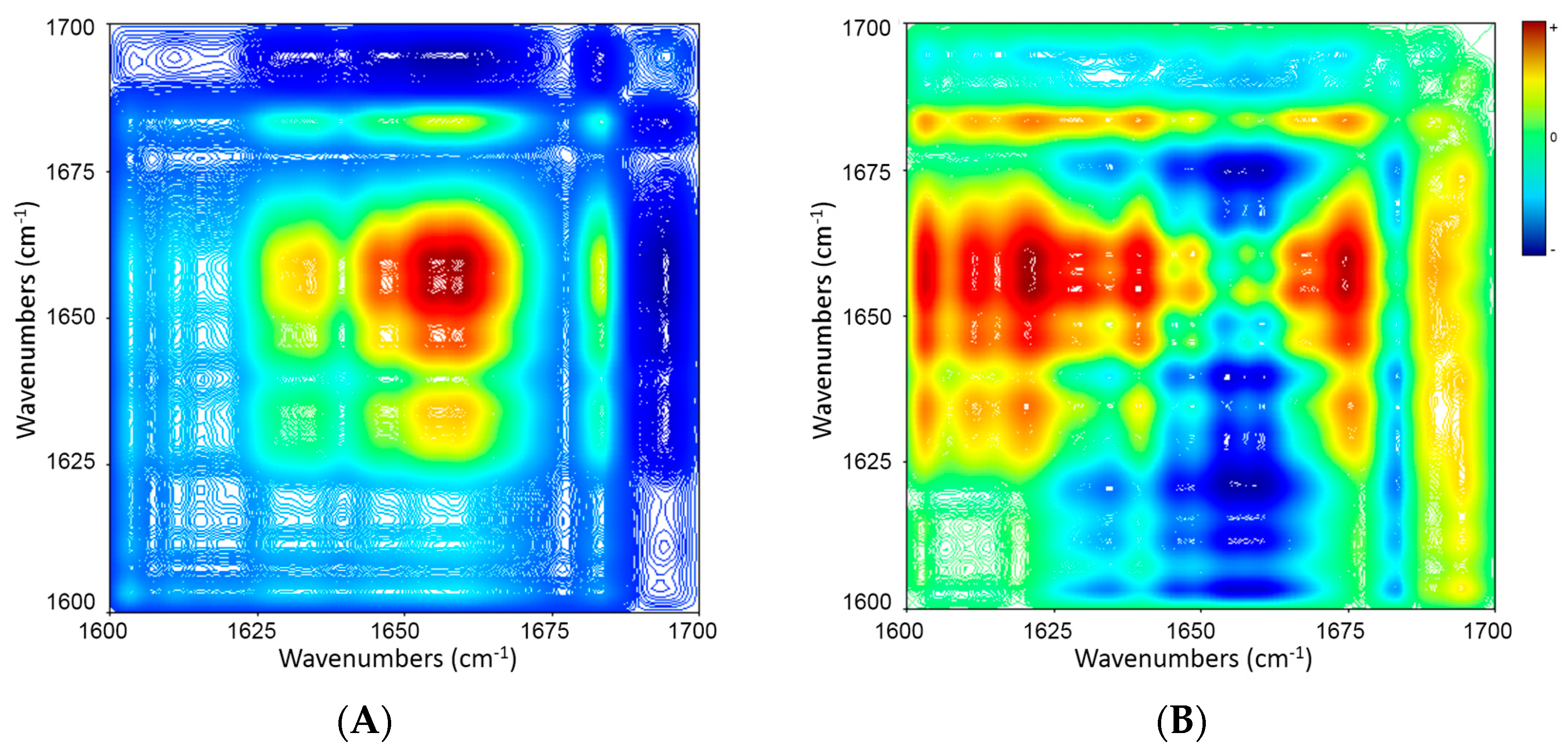
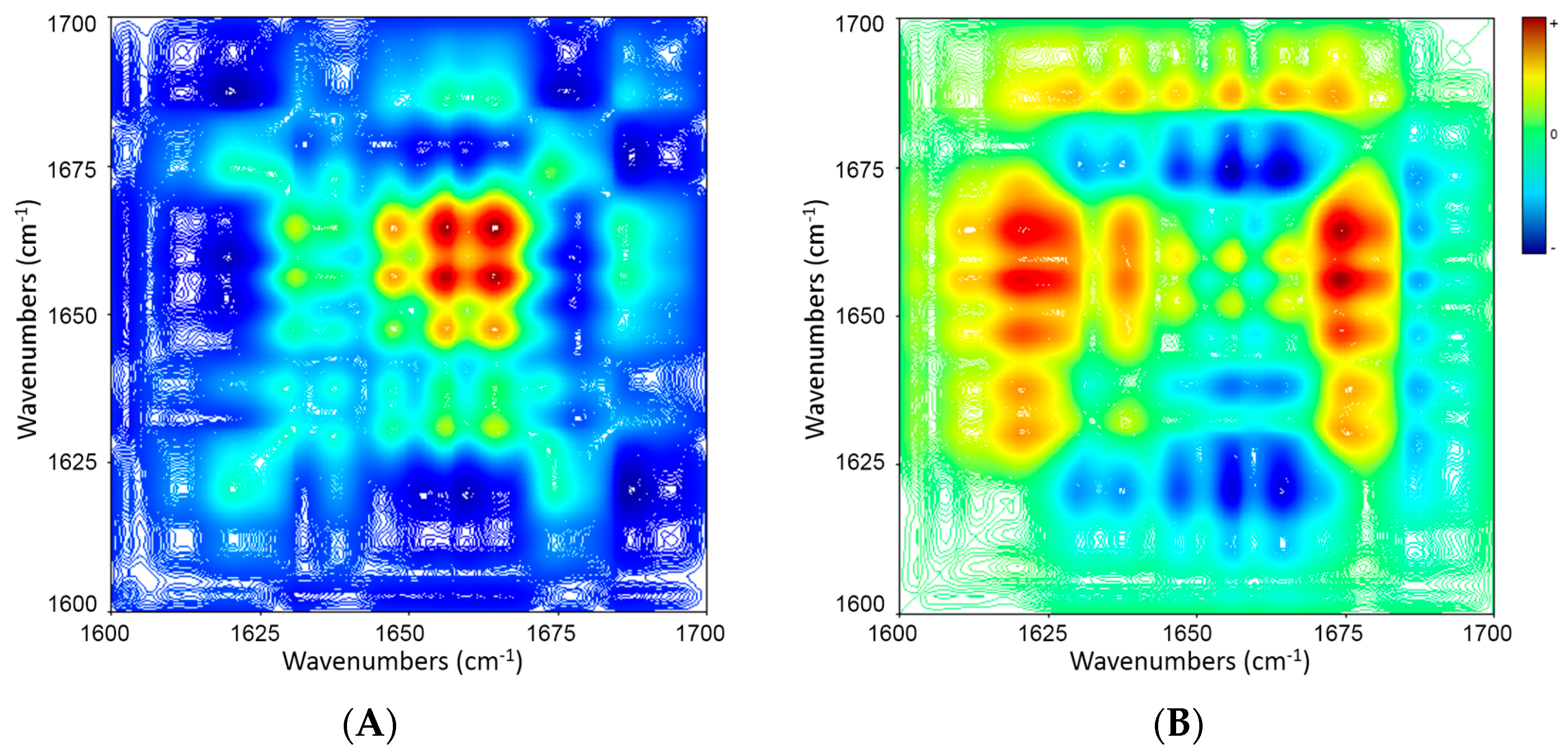
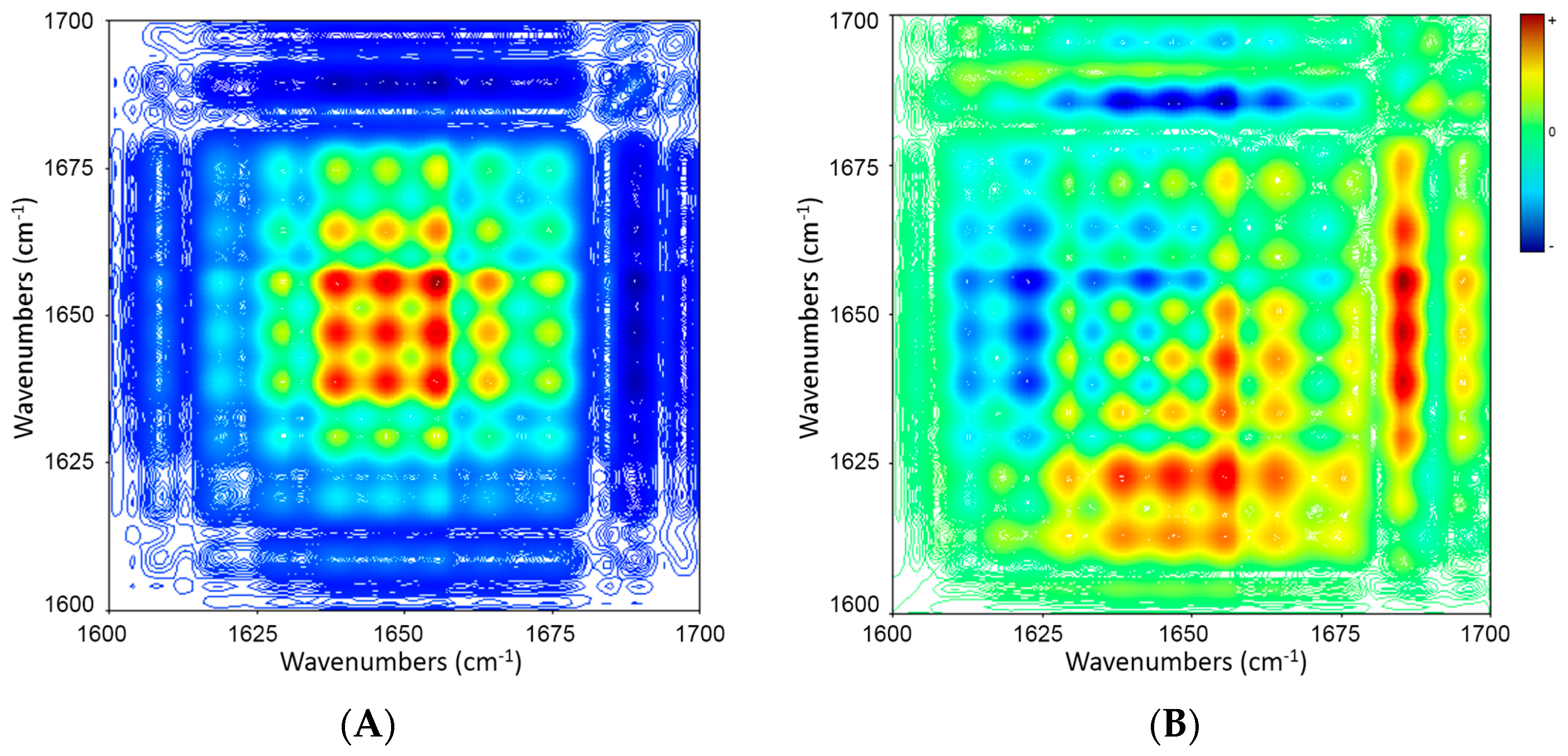
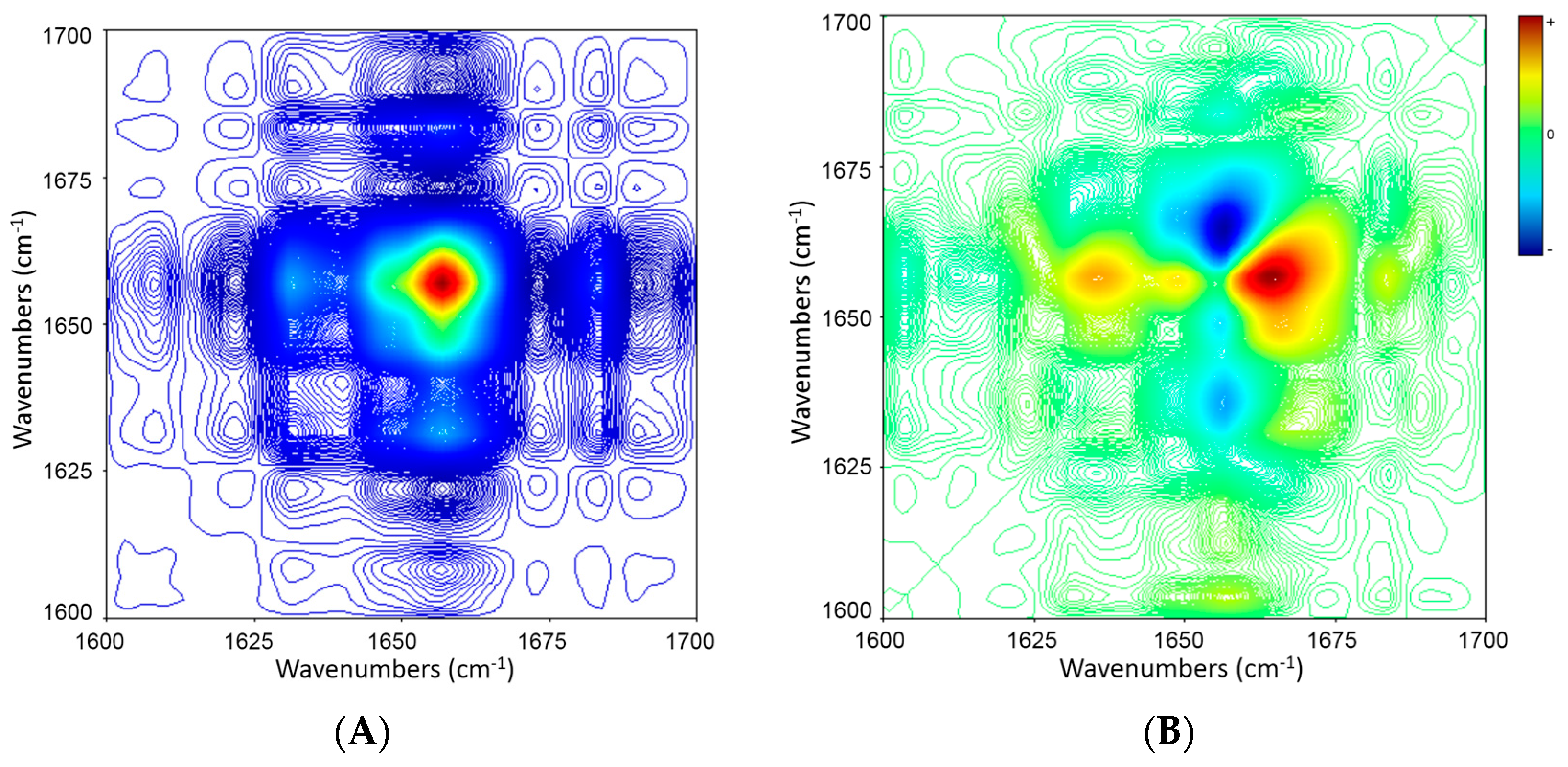
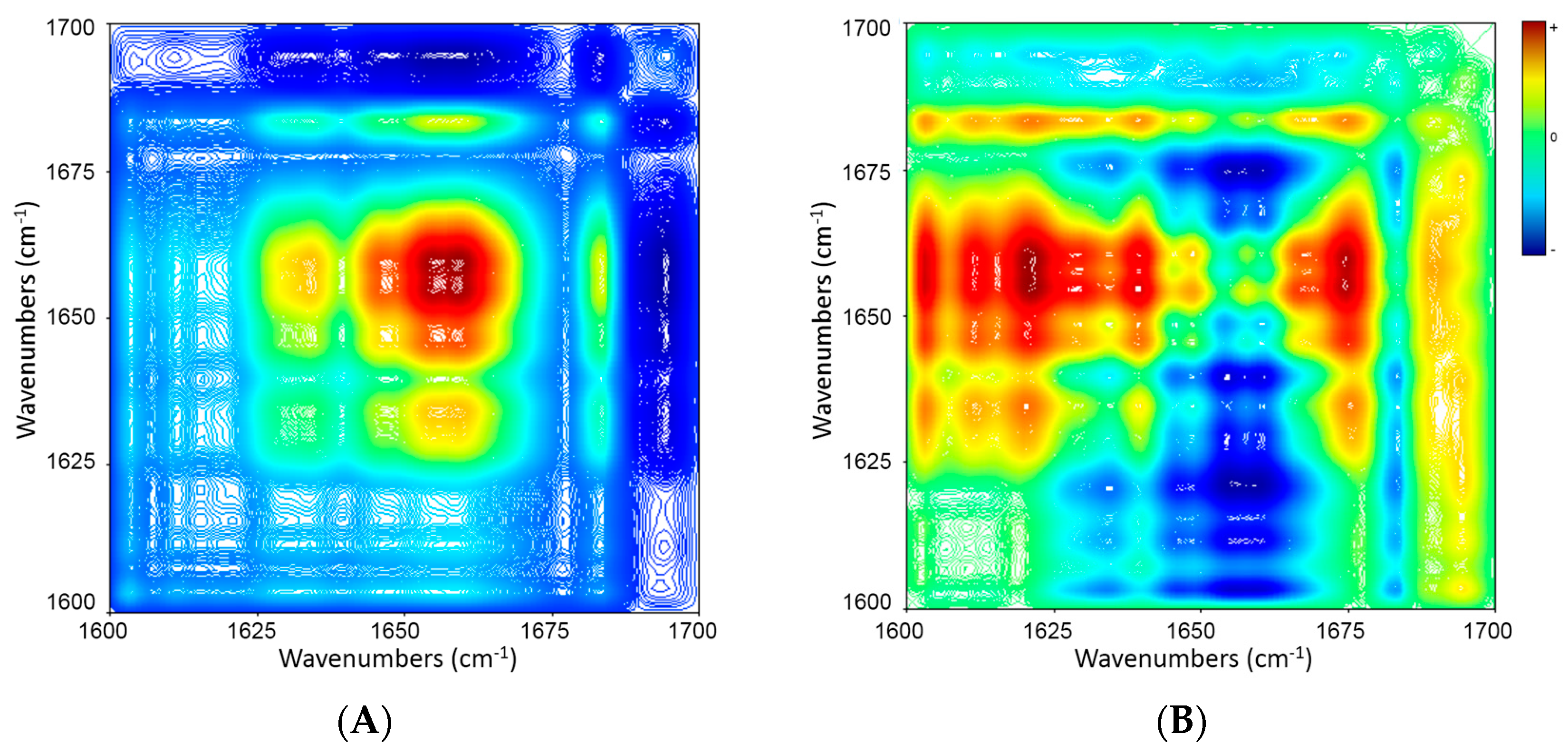
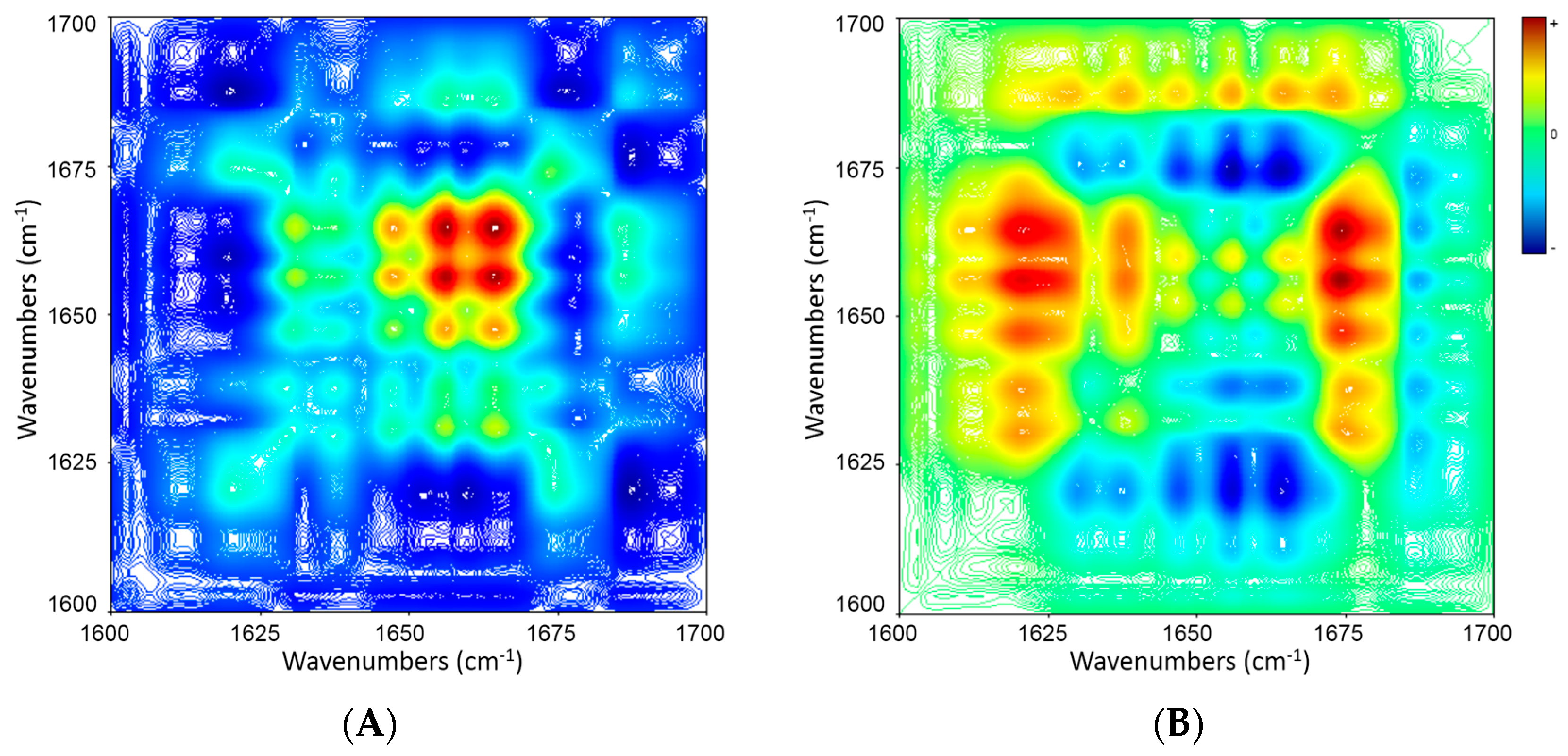
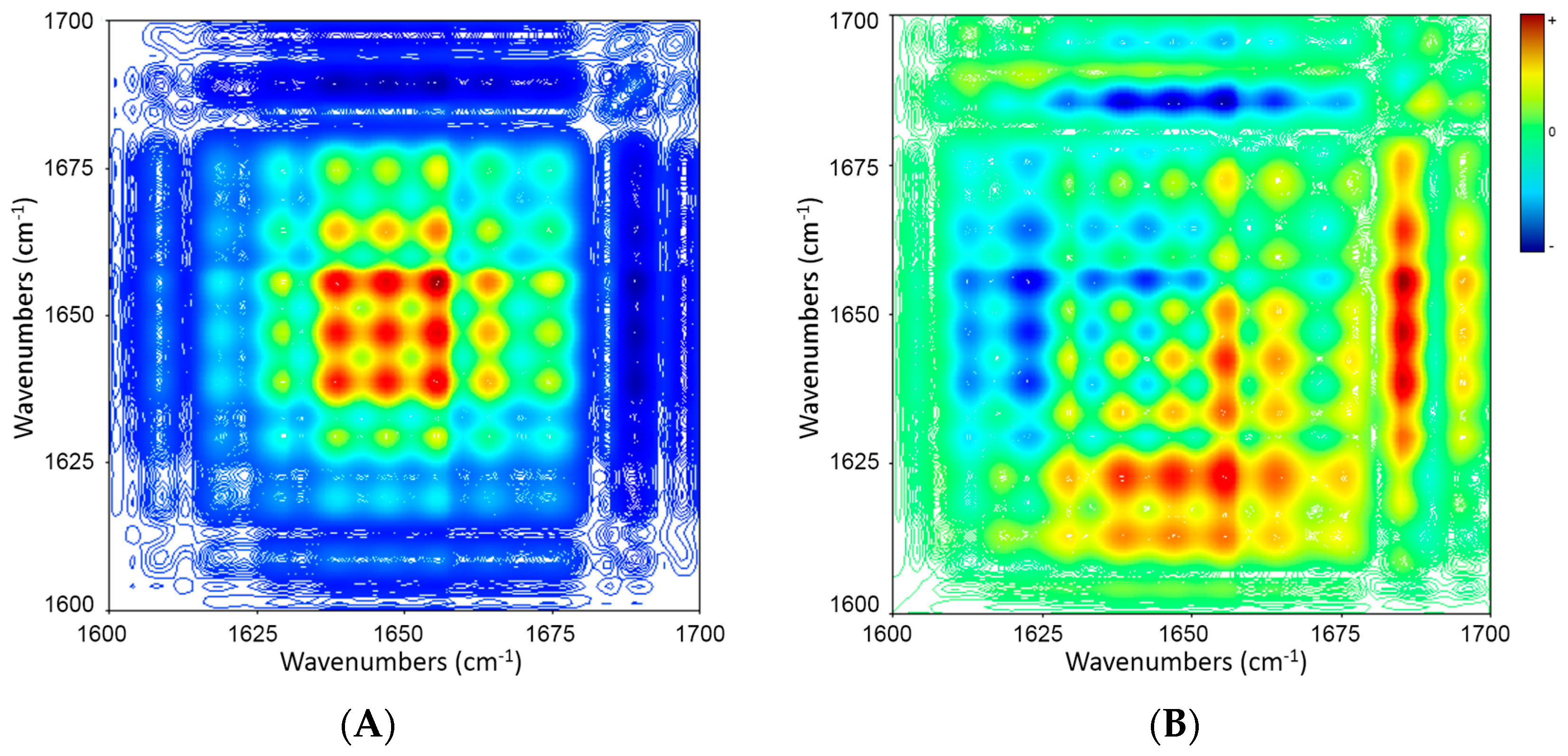
2.5. 2D Correlation Analysis of ICFs and PSCs
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Extraction of ICFs and PSCs from Sea Cucumber Body Walls
3.3. Amino Acid Composition Analysis
3.4. Thermal Analysis of ICFs and PSCs
3.5. FTIR Measurements
3.6. Curve-Fitting
3.7. 2D Correlation Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Saito, M.; Kunisaki, N.; Urano, N.; Kimura, S. Collagen as the major edible component of sea cucumber (Stichopus japonicus). J. Food Sci. 2002, 67, 1319–1322. [Google Scholar] [CrossRef]
- Motokawa, T.; Tsuchi, A. Dynamic mechanical properties of body-wall dermis in various mechanical states and their implications for the behavior of sea cucumbers. Biol. Bull. 2003, 205, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.W.; Dong, X.P.; Zhou, D.Y.; Gao, Y.; Yang, J.F.; Li, D.M.; Zhao, X.K.; Ren, T.T.; Ye, W.X.; Tan, H.; et al. Physicochemical properties and radical scavenging capacities of pepsin-solubilized collagen from sea cucumber Stichopus japonicus. Food Hydrocoll. 2012, 28, 182–188. [Google Scholar] [CrossRef]
- Thurmond, F.A.; Koob, T.J.; Bowness, J.M.; Trotter, J.A. Partial biochemical and immunologic characterization of fibrillin microfibrils from sea cucumber dermis. Connect. Tissue Res. 1997, 36, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Xue, D.; Zhang, Z.; Xu, J.; Xue, C. Rheological and structural properties of sea cucumber Stichopus japonicus during heat treatment. J. Ocean Univ. Chin. 2005, 4, 244–247. [Google Scholar] [CrossRef]
- Matsumura, T.; Shinmei, M.; Nagai, Y. Disaggregation of connective tissue: Preparation of fibrous components from sea cucumber body wall and calf skin. J. Biochem. 1973, 73, 155–162. [Google Scholar] [PubMed]
- Makareeva, E.; Leikin, S. Collagen structure, folding and function. In Osteogenesis Imperfecta; Shapiro, J.R., Byers, P.H., Glorieux, F.H., Sponseller, P.D., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 71–84. [Google Scholar]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.X.; Xue, C.H.; Li, Z.J.; Zhang, Y.Q.; Dong, P.; Fu, X.Y.; Gao, X. Characterization and subunit composition of collagen from the body wall of sea cucumber Stichopus japonicus. Food Chem. 2007, 100, 1120–1125. [Google Scholar] [CrossRef]
- Park, S.-Y.; Lim, H.K.; Lee, S.; Hwang, H.C.; Cho, S.K.; Cho, M. Pepsin-solubilised collagen (psc) from red sea cucumber (Stichopus japonicus) regulates cell cycle and the fibronectin synthesis in hacat cell migration. Food Chem. 2012, 132, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.W.; Yu, J.W.; Zhang, Z.S.; Zhou, D.Y.; Yang, J.F.; Li, D.M.; Murata, Y. Purification and partial characterization of an acid phosphatase from the body wall of sea cucumber Stichopus japonicus. Process Biochem. 2009, 44, 875–879. [Google Scholar] [CrossRef]
- Saguer, E.; Alvarez, P.; Ismail, A.A. Heat-induced denaturation/aggregation of porcine plasma and its fractions studied by ftir spectroscopy. Food Hydrocoll. 2012, 27, 208–219. [Google Scholar] [CrossRef]
- Saguer, E.; Alvarez, P.; Sedman, J.; Ramaswamy, H.S.; Ismail, A.A. Heat-induced gel formation of plasma proteins: New insights by ftir 2d correlation spectroscopy. Food Hydrocoll. 2009, 23, 874–879. [Google Scholar] [CrossRef]
- Saguer, E.; Alvarez, P.A.; Sedman, J.; Ismail, A.A. Study of the denaturation/aggregation behaviour of whole porcine plasma and its protein fractions during heating under acidic ph by variable-temperature ftir spectroscopy. Food Hydrocoll. 2013, 33, 402–414. [Google Scholar] [CrossRef]
- Murayama, K.; Tomida, M. Heat-induced secondary structure and conformation change of bovine serum albumin investigated by fourier transform infrared spectroscopy. Biochemistry 2004, 43, 11526–11532. [Google Scholar] [CrossRef] [PubMed]
- Prystupa, D.A.; Donald, A.M. Infrared study of gelatin conformations in the gel and sol states. Polym. Gels Netw. 1996, 4, 87–110. [Google Scholar] [CrossRef]
- Lazarev, Y.A.; Grishkovsky, B.A.; Khromova, T.B. Amide i band of ir spectrum and structure of collagen and related polypeptides. Biopolymers 1985, 24, 1449–1478. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.R.; Wang, B.; Chi, C.F.; Zhang, Q.H.; Gong, Y.D.; Tang, J.J.; Luo, H.Y.; Ding, G.F. Isolation and characterization of acid soluble collagens and pepsin soluble collagens from the skin and bone of spanish mackerel (scomberomorous niphonius). Food Hydrocoll. 2013, 31, 103–113. [Google Scholar] [CrossRef]
- Liu, D.; Liang, L.; Regenstein, J.M.; Zhou, P. Extraction and characterisation of pepsin-solubilised collagen from fins, scales, skins, bones and swim bladders of bighead carp (hypophthalmichthys nobilis). Food Chem. 2012, 133, 1441–1448. [Google Scholar] [CrossRef]
- Singh, P.; Benjakul, S.; Maqsood, S.; Kishimura, H. Isolation and characterisation of collagen extracted from the skin of striped catfish (pangasianodon hypophthalmus). Food Chem. 2011, 124, 97–105. [Google Scholar] [CrossRef]
- Joseph, W.; Brauner, R.M. A quantitative reconstruction of the amide i contour in the ir spectra of peptides and proteins: From structure to spectrum. Biomed. Eng. 2012, 17–52. [Google Scholar]
- Bryan, M.A.; Brauner, J.W.; Anderle, G.; Flach, C.R.; Brodsky, B.; Mendelsohn, R. Ftir studies of collagen model peptides: Complementary experimental and simulation approaches to conformation and unfolding. J. Am. Chem. Soc. 2007, 129, 7877–7884. [Google Scholar] [CrossRef] [PubMed]
- Cruzeiro, L.; Freedman, H. The temperature dependent amide i band of crystalline acetanilide. Phys. Lett. A 2013, 377, 1593–1596. [Google Scholar] [CrossRef]
- Gough, K.M.; Tzadu, L.; Kastyak, M.Z.; Kuzyk, A.C.; Julian, R.L. Theoretical and experimental considerations for interpretation of amide i bands in tissue. Vib. Spectrosc. 2010, 53, 71–76. [Google Scholar] [CrossRef]
- Noda, I. Generalized two-dimensional correlation spectroscopy. In Frontiers of Molecular Spectroscopy; Laane, J., Ed.; Elsevier: Amsterdam, The Netherlands, 2009; pp. 367–381. [Google Scholar]
- Zimm, J.K.; Bragg, J.K. Theory of the phase transition between helix and random coil in polypeptide chains. J. Chem. Phys. 1959, 31, 526–535. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; Macrae, T.P.; Rogers, G.E. Keratins, Their Composition Structure and Biosynthesis; Thomas Publishing: New York, NY, USA, 1972. [Google Scholar]
- Chys, P.; Gielens, C.; Meersman, F. Ftir 2d correlation spectroscopy of α1 and α2 fractions of an alkali-pretreated gelatin. Biochim. Biophys. Acta 2011, 1814, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Meersman, F.; Smeller, L.; Heremans, K. Comparative fourier transform infrared spectroscopy study of cold-, pressure-, and heat-induced unfolding and aggregation of myoglobin. Biophys. J. 2002, 82, 2635–2644. [Google Scholar] [CrossRef]
- Paquet, M.J.; Laviolette, M.; Pézolet, M.; Auger, M. Two-dimensional infrared correlation spectroscopy study of the aggregation of cytochrome c, in the presence of dimyristoylphosphatidylglycerol. Biophys. J. 2001, 81, 305–312. [Google Scholar] [CrossRef]
- Sun, W.Y.; Fang, J.L.; Cheng, M.; Xia, P.Y.; Tang, W.X. Secondary structure dependent on metal ions of copper, zinc superoxide dismutase investigated by fourier transform ir spectroscopy. Biopolymers 1997, 42, 297–303. [Google Scholar] [CrossRef]
- Popescu, M.V.C.; Craciunescu, O.; Popescu, M.C. Structural analysis of some soluble elastins by means of ft-ir and 2d ir correlation spectroscopy. Biopolymers 2010, 93, 1072–1084. [Google Scholar] [CrossRef] [PubMed]
- Petibois, C.; Déléris, G. Chemical mapping of tumor progression by FT-IR imaging: Towards molecular histopathology. Trends Biotechnol. 2006, 24, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Petibois, C.; Gouspillou, G.; Wehbe, K.; Delage, J.P.; Déléris, G. Analysis of type i and iv collagens by FT-IR spectroscopy and imaging for a molecular investigation of skeletal muscle connective tissue. Anal. Bioanal. Chem. 2006, 386, 1961–1966. [Google Scholar] [CrossRef] [PubMed]
- Susi, H.; Byler, D.M. Fourier deconvolution of the amide i raman band of proteins as related to conformation. Appl. Spectrosc. 1988, 42, 819–826. [Google Scholar] [CrossRef]
- Choong, Y.K.; Sun, S.Q.; Zhou, Q.; Ismail, Z.; Rashid, B.A.A.; Tao, J.X. Determination of storage stability of the crude extracts of Ganoderma lucidum using FTIR and 2D-IR spectroscopy. Vib. Spectrosc. 2011, 57, 87–96. [Google Scholar] [CrossRef]
- Sample Availability: Not available.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | ICF | PSC |
|---|---|---|
| Aspartic acid | 6.57 ± 0.08 | 6.66 ± 0.03 |
| Glutamic acid | 10.71 ± 0.05 | 10.66 ± 0.08 |
| Hydroxyproline | 6.06 ± 0.10 | 6.13 ± 0.03 |
| Serine | 4.61 ± 0.03 | 4.60 ± 0.04 |
| Arginine | 5.71 ± 1.61 | 3.69 ± 1.81 |
| Glycine | 34.05 ± 0.95 | 36.44 ± 1.72 |
| Threonine | 3.87 ± 0.04 | 3.88 ± 0.02 |
| Proline | 9.02 ± 0.02 | 8.98 ± 0.03 |
| Alanine | 7.07 ± 0.02 | 7.03 ± 0.02 |
| Valine | 2.24 ± 0.05 | 2.15 ± 0.06 |
| Methionine | 0.70 ± 0.14 | 0.51 ± 0.47 |
| Isoleucine | 1.74 ± 0.02 | 1.72 ± 0.03 |
| Leucine | 1.92 ± 0.03 | 1.88 ± 0.03 |
| Phenylalanine | 0.76 ± 0.11 | 1.89 ± 0.60 |
| Histidine | 3.02 ± 0.40 | 2.08 ± 0.80 |
| Lysine | 0.85 ± 0.01 | 0.77 ± 0.06 |
| Tyrosine | 1.11 ± 0.18 | 0.91 ± 0.11 |
| Wavenumber (cm−1) | Assignment |
|---|---|
| 1624 ± 4 | β-sheet |
| 1631 ± 3 | β-sheet |
| 1637 ± 3 | β-sheet |
| 1641 ± 2 | 310-helix |
| 1645 ± 4 | random coil |
| 1653 ± 4 | α-helix |
| 1663 ± 4 | β-turn |
| 1671 ± 3 | β-turn |
| 1675 ± 5 | β-sheet |
| 1683 ± 2 | β-turn |
| 1689 ± 2 | β-turn |
| 1694 ± 2 | β-turn |
| Wavenumber (cm−1) | 1683 ↓ | 1656 ↓ |
|---|---|---|
| 1631 ↓ | ≈ | + − → |
| 1656 ↓ | + + ← |
| Wavenumber (cm−1) | 1695 ↓ | 1683 ↑ | 1657 ↑ | 1646 ↑ | 1634 ↑ |
|---|---|---|---|---|---|
| 1611 ↑ | − + → | + − → | + − → | + − → | + − → |
| 1634 ↑ | − + → | + − → | + − → | + − → | |
| 1646 ↑ | − + → | + − → | + − → | ||
| 1657 ↑ | − + → | + − → | |||
| 1683 ↑ | − + → |
| Wavenumber (cm−1) | 1686 ↓ | 1673 ↓ | 1664 ↓ | 1656 ↓ | 1648 ↓ | 1637 ↓ |
|---|---|---|---|---|---|---|
| 1630 ↓ | + − → | + + ← | + − → | + − → | + − → | ≈ |
| 1637 ↓ | + − → | + + ← | + − → | + − → | + − → | |
| 1648 ↓ | + − → | + + ← | ≈ | ≈ | ||
| 1656 ↓ | + − → | + + ← | + − → | |||
| 1664 ↓ | + − → | + + ← | ||||
| 1673 ↓ | + − → |
| Wavenumber (cm−1) | 1689 ↓ | 1675 ↑ | 1664 ↑ | 1656 ↑ | 1647 ↑ | 1639 ↑ | 1629 ↑ |
|---|---|---|---|---|---|---|---|
| 1618 ↑ | − + → | + + ← | + + ← | + + ← | + + ← | + + ← | + + ← |
| 1629 ↑ | − + → | + + ← | + + ← | + + ← | + + ← | ≈ | |
| 1639 ↑ | − + → | ≈ | + + ← | + + ← | ≈ | ||
| 1647 ↑ | − + → | + − → | + + ← | + + ← | |||
| 1656 ↑ | − + → | + − → | + + ← | ||||
| 1664 ↑ | − + → | + − → | |||||
| 1675 ↑ | − + → |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, L.; Bi, J.-R.; Li, D.-M.; Dong, M.; Zhao, Z.-Y.; Dong, X.-P.; Zhou, D.-Y.; Zhu, B.-W. Unfolding/Refolding Study on Collagen from Sea Cucumber Based on 2D Fourier Transform Infrared Spectroscopy. Molecules 2016, 21, 1546. https://doi.org/10.3390/molecules21111546
Qin L, Bi J-R, Li D-M, Dong M, Zhao Z-Y, Dong X-P, Zhou D-Y, Zhu B-W. Unfolding/Refolding Study on Collagen from Sea Cucumber Based on 2D Fourier Transform Infrared Spectroscopy. Molecules. 2016; 21(11):1546. https://doi.org/10.3390/molecules21111546
Chicago/Turabian StyleQin, Lei, Jing-Ran Bi, Dong-Mei Li, Meng Dong, Zi-Yuan Zhao, Xiu-Ping Dong, Da-Yong Zhou, and Bei-Wei Zhu. 2016. "Unfolding/Refolding Study on Collagen from Sea Cucumber Based on 2D Fourier Transform Infrared Spectroscopy" Molecules 21, no. 11: 1546. https://doi.org/10.3390/molecules21111546
APA StyleQin, L., Bi, J.-R., Li, D.-M., Dong, M., Zhao, Z.-Y., Dong, X.-P., Zhou, D.-Y., & Zhu, B.-W. (2016). Unfolding/Refolding Study on Collagen from Sea Cucumber Based on 2D Fourier Transform Infrared Spectroscopy. Molecules, 21(11), 1546. https://doi.org/10.3390/molecules21111546