
Gallic Acid Content in Taiwanese Teas at Different Degrees of Fermentation and Its Antioxidant Activity by Inhibiting PKCδ Activation: In Vitro and in Silico Studies



Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of the Fermentation Process on Catechins in Teas
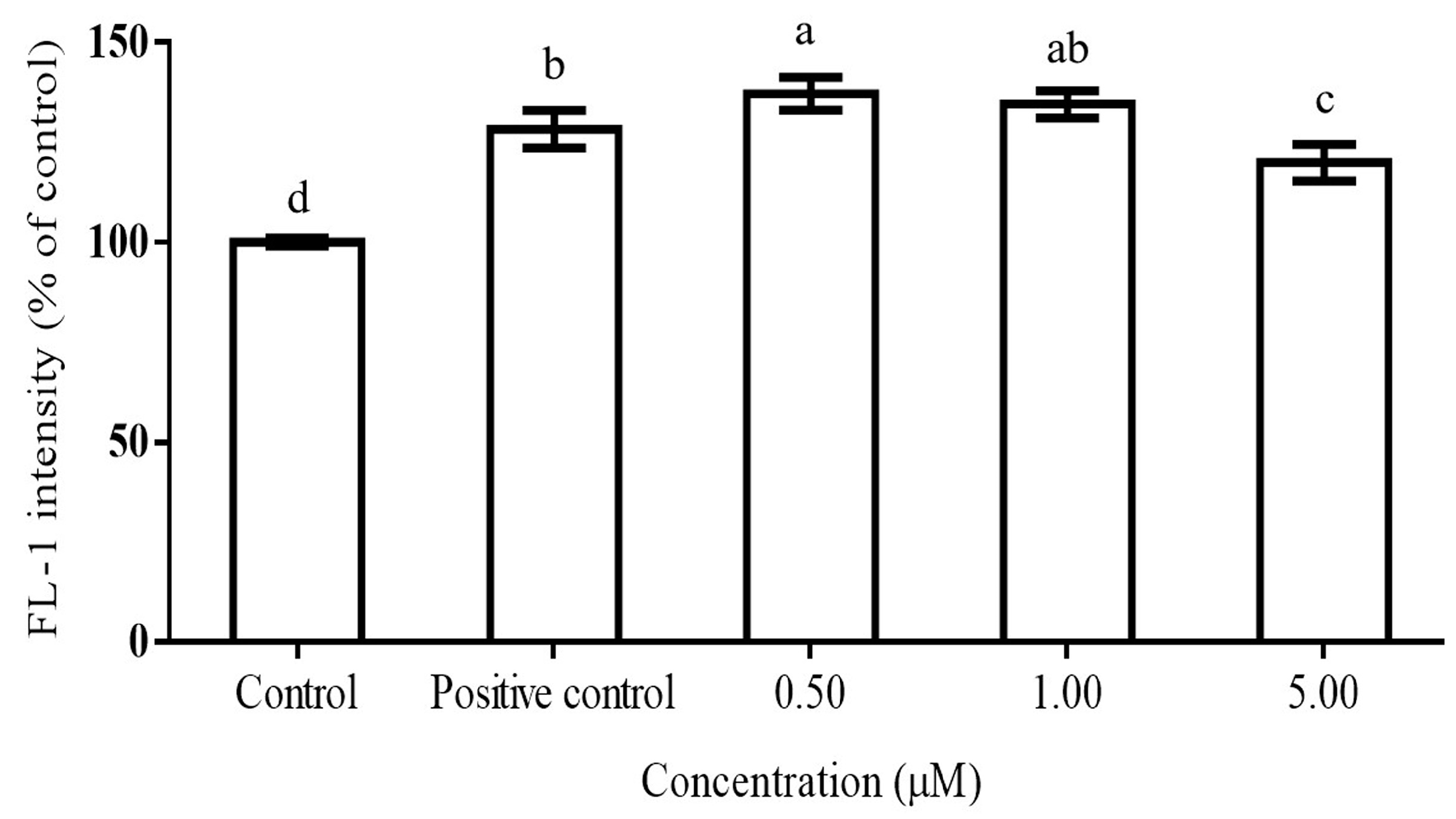
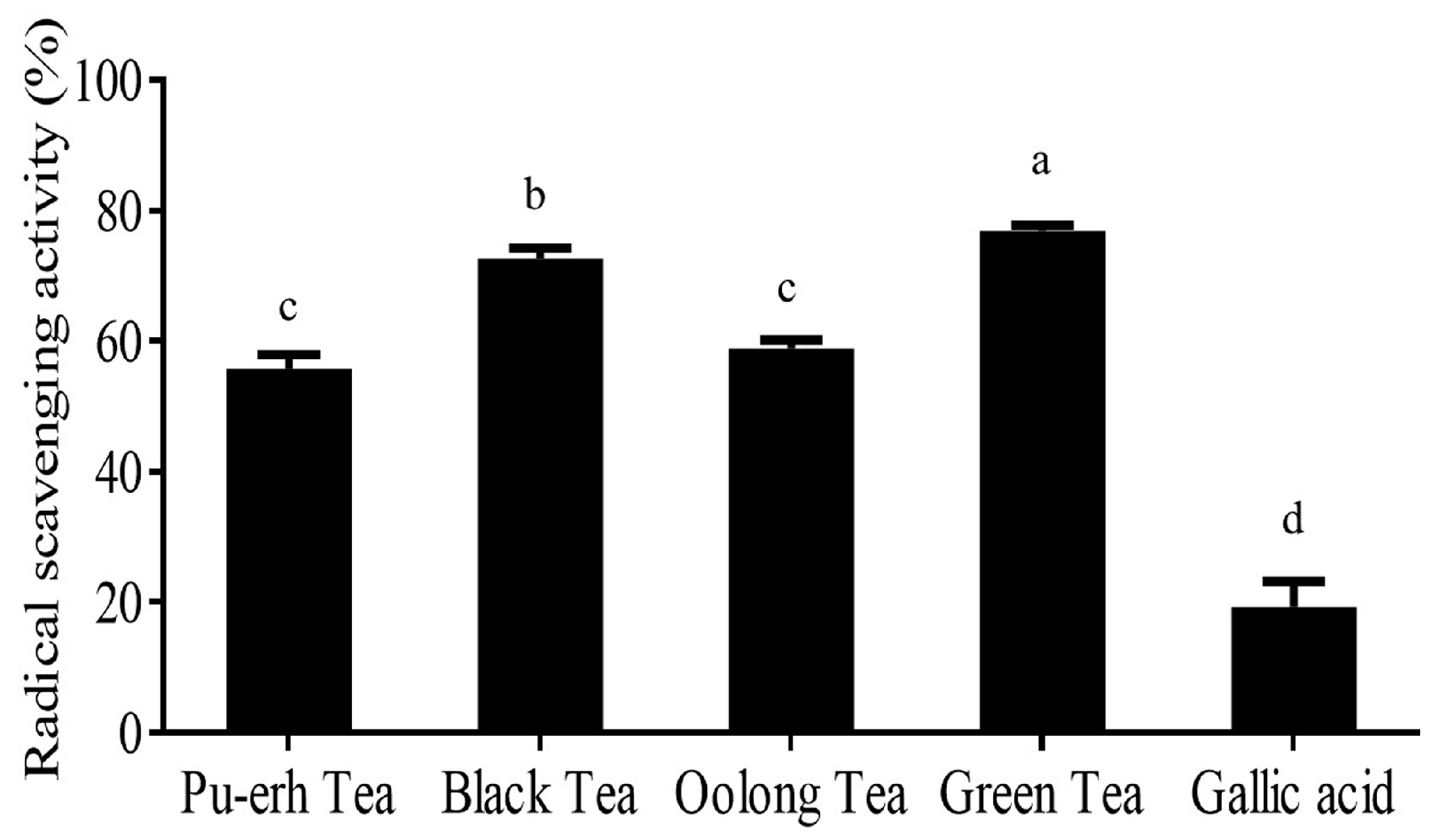
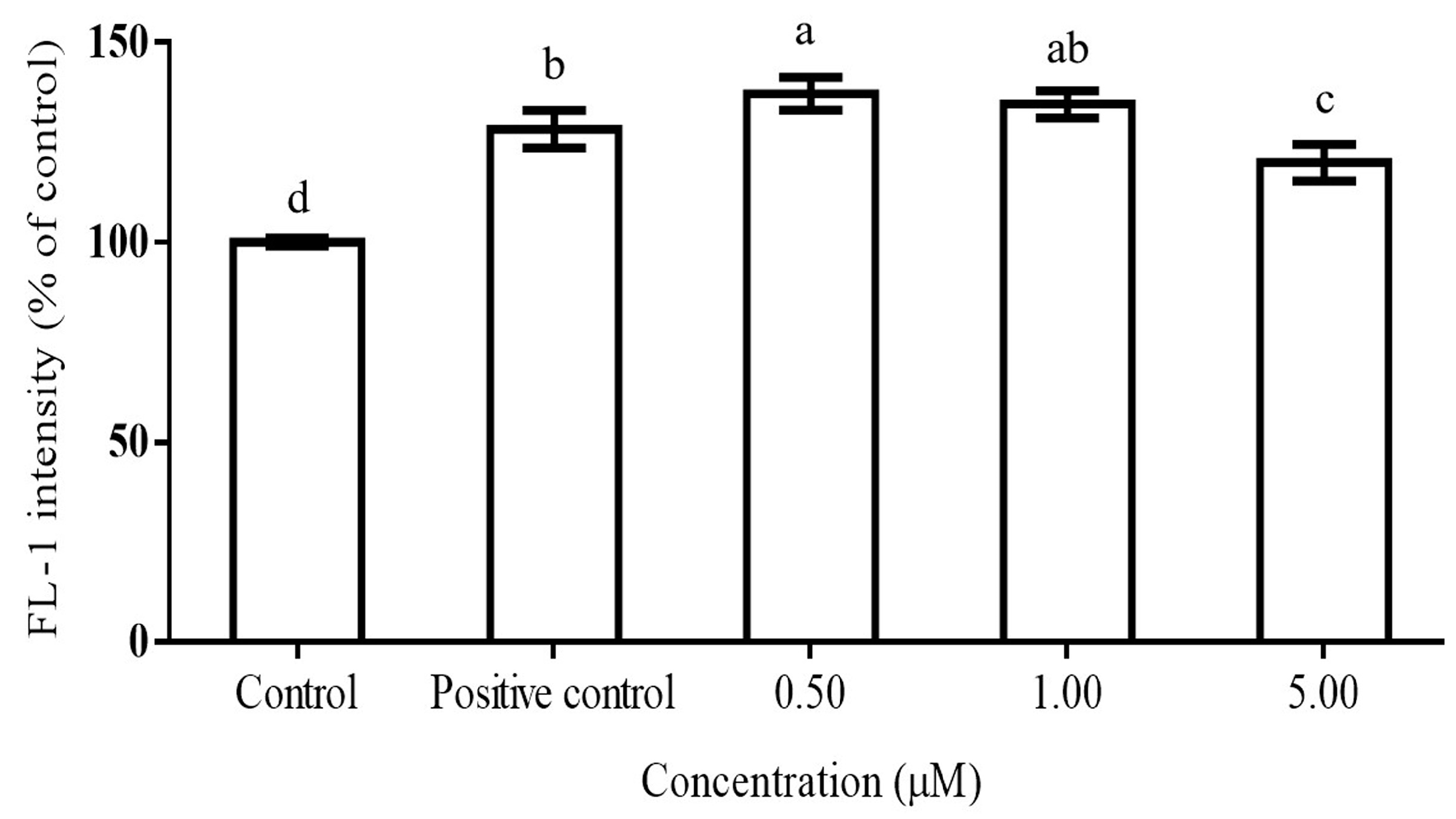
2.2. DPPH Scavenging Activity of Teas and Inhibition of ROS Production by Gallic Acid in PMA-Activated RAW264.7 Cells
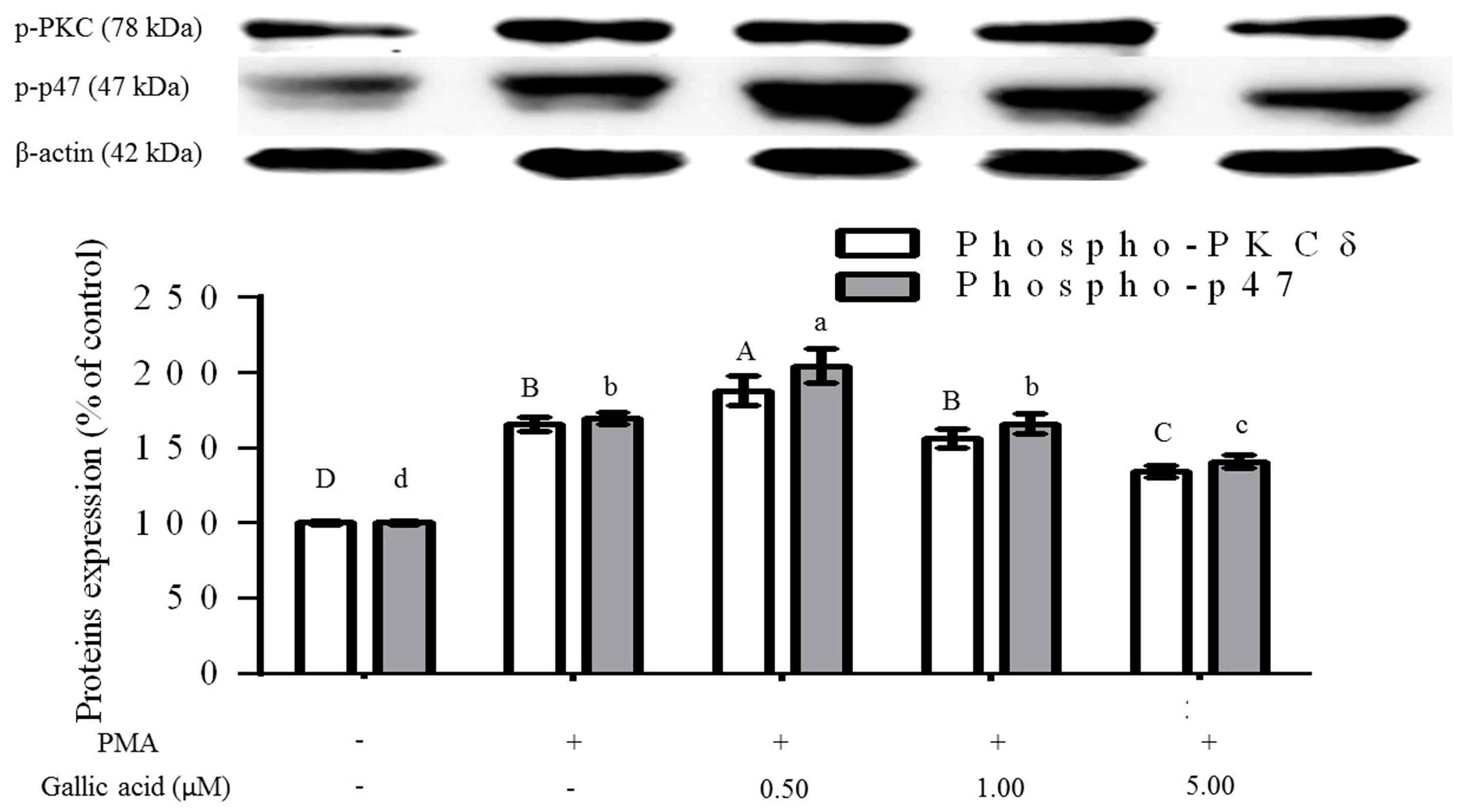
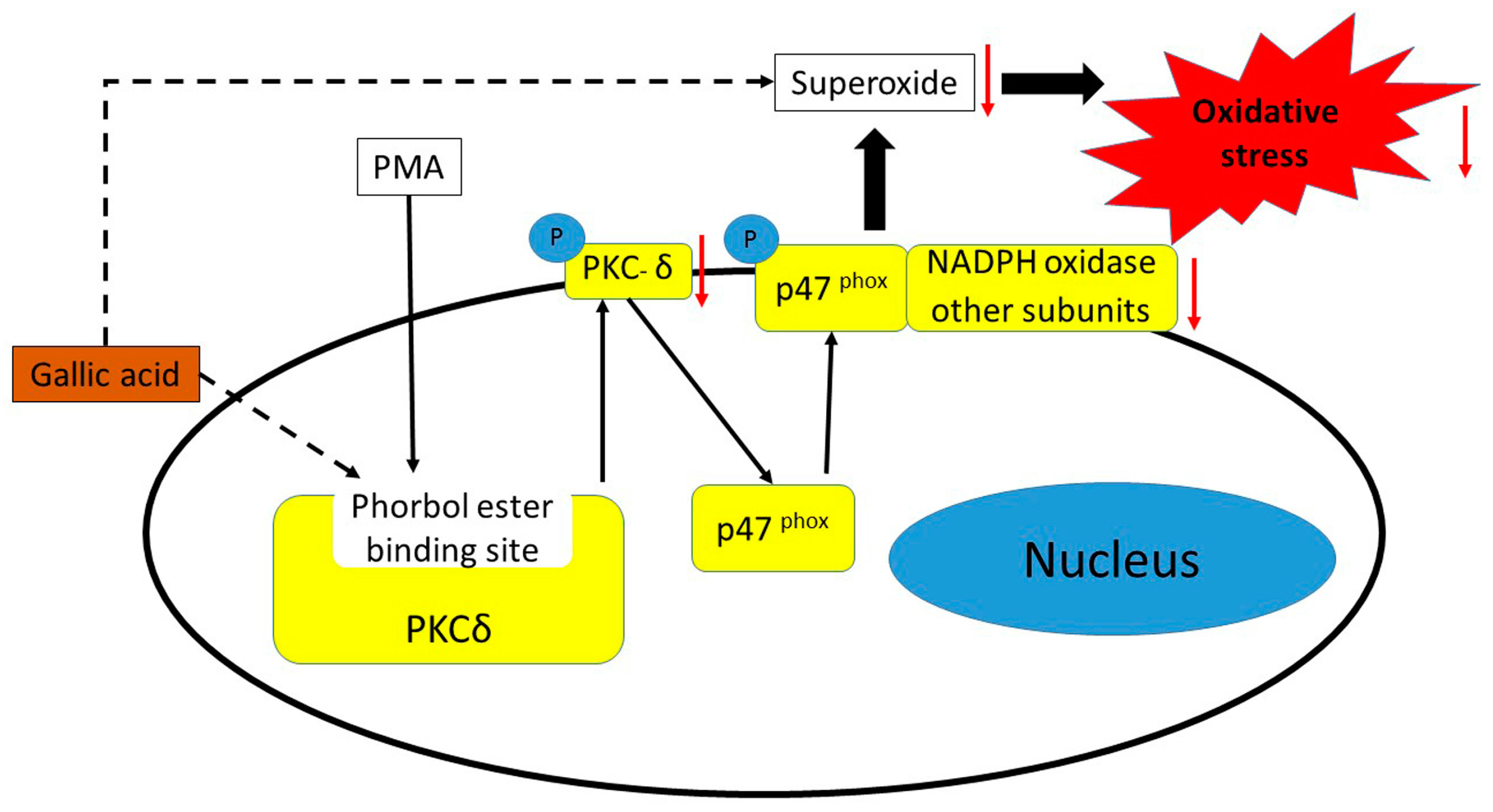
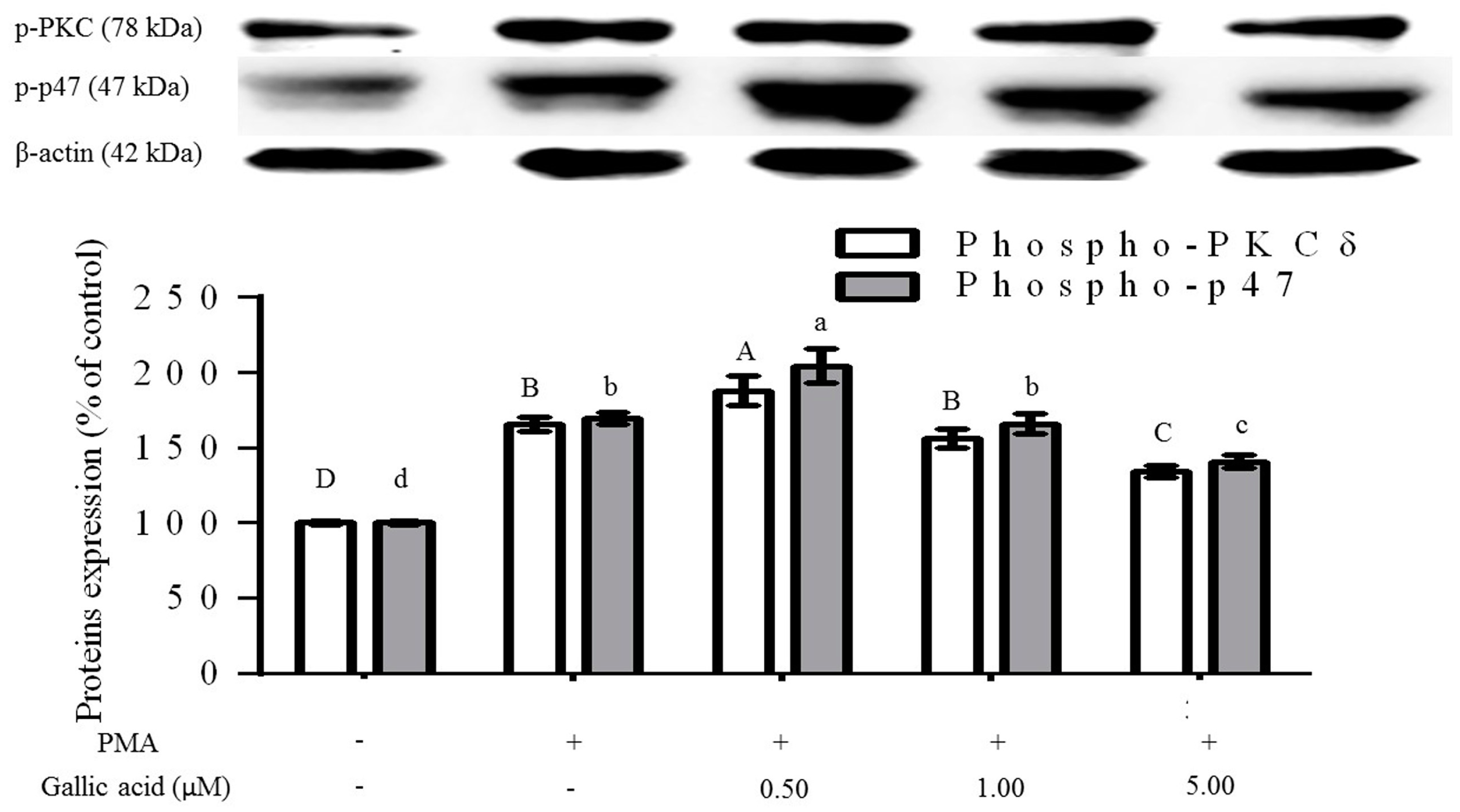
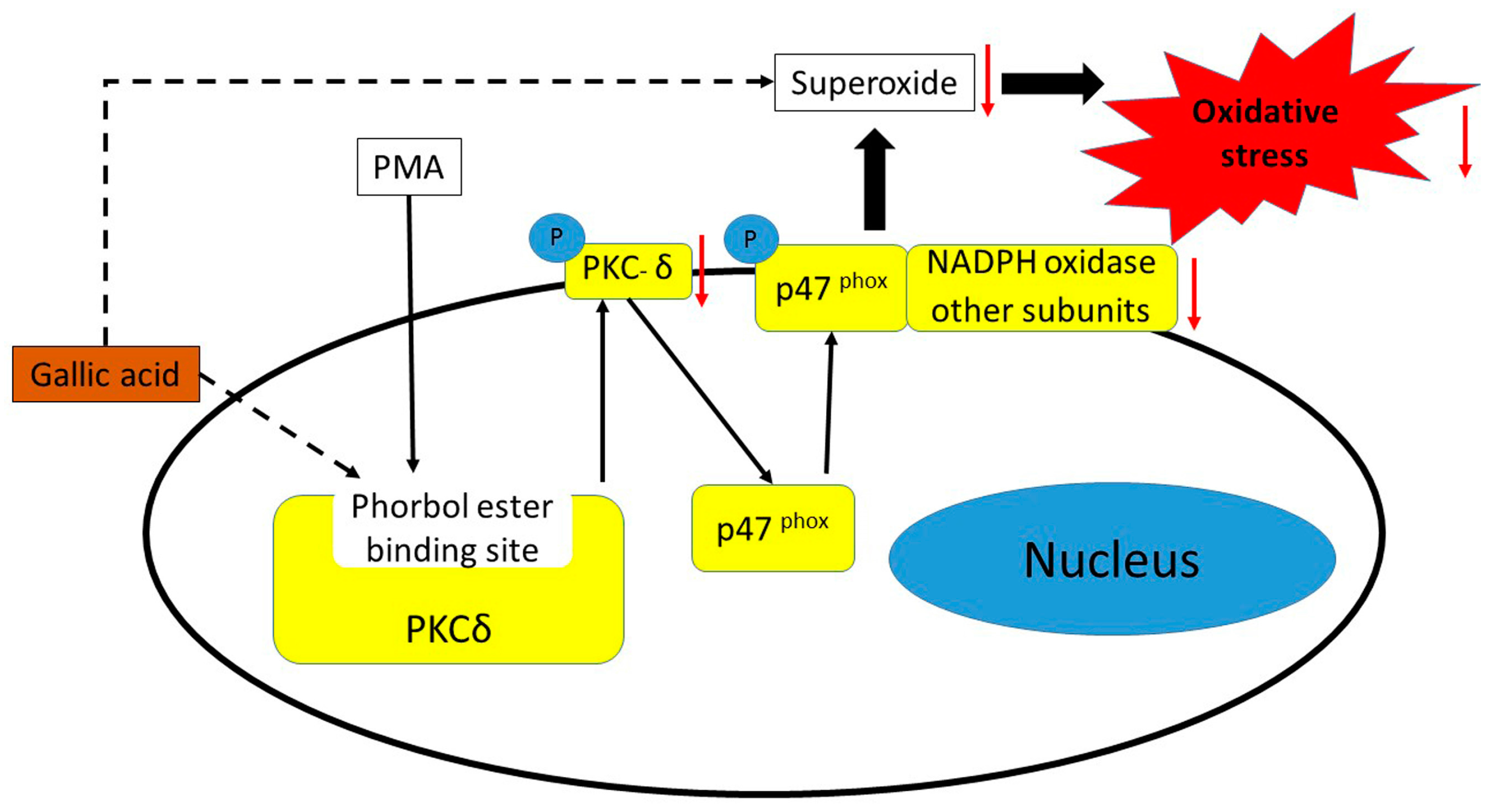
2.3. Inhibition of Phosphorylation of PKCδ and p47phox Proteins
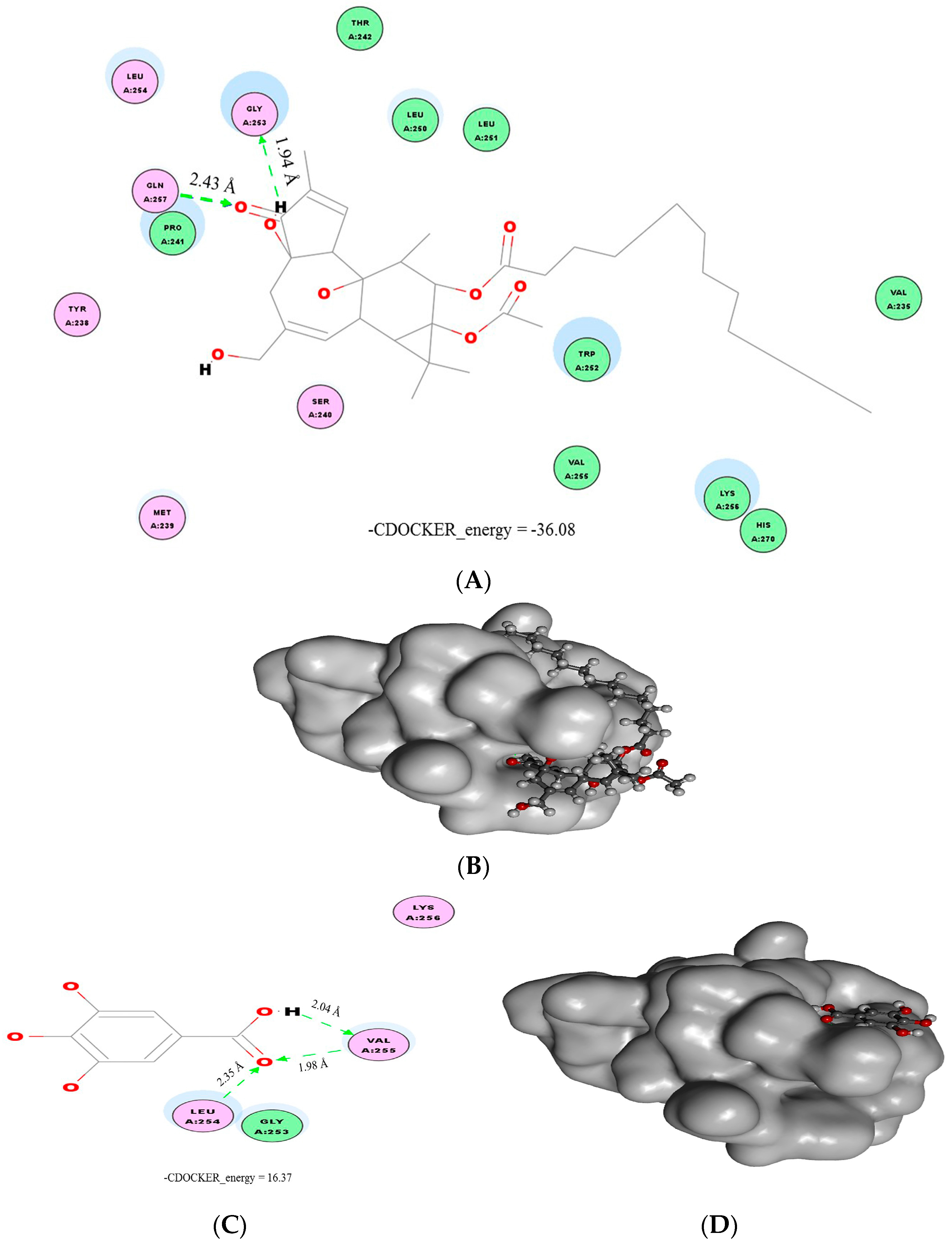
2.4. Molecular Docking
3. Materials and Methods
3.1. Chemical and Raw Materials
3.2. Tea Extraction
3.3. HPLC Analysis
3.4. DPPH Radical Scavenging Assay
3.5. Cell Culture
3.6. Cell Viability Test
3.7. ROS Inhibition and IC50 Determination
3.8. Protein Immunoblotting Assay
3.9. Molecular Docking
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Conflicts of Interest
References
- Tseng, H.C.; Wang, C.J.; Cheng, S.H.; Sun, Z.J.; Chen, P.S.; Lee, C.T.; Lin, S.H.; Yang, Y.K.; Yang, Y.C. Tea-drinking habit among new university students: Associated factors. Kaohsiung J. Med. Sci. 2014, 30, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.M.; Lan, S.J.; Lee, Y.H.; Huang, J.K.; Huang, C.H.; Hsieh, C.C. Tea consumption: Fluid intake and bladder cancer risk in southern Taiwan. Urology 1999, 54, 823–828. [Google Scholar] [CrossRef]
- Huang, Y.; Xiao, X.; Cong, L.; Wu, M.; Huang, Y.; Yao, Y. A fermented tea with high levels of gallic acid processed by anaerobic solid-state fermentation. LWT-Food Sci. Technol. 2016, 71, 260–267. [Google Scholar] [CrossRef]
- Yang, C.S.; Lambert, J.D. Research on tea and health. Pharmacol. Res. 2011, 64, 85–86. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Chiu, A.; Barone, M.K.; Avino, D.; Wang, F.; Coleman, C.I.; Phung, O.J. Green tea catechins decrease total and low-density lipoprotein cholesterol: A systematic review and meta-analysis. J. Am. Diet. Assoc. 2011, 111, 1720–1729. [Google Scholar] [CrossRef] [PubMed]
- Huxley, R.; Lee, C.M.Y.; Barzi, F.; Timmermeister, L.; Czernichow, S.; Perkovic, V.; Grobbee, D.E.; Batty, D.; Woodward, M. Coffee, decaffeinated coffee, and tea consumption in relation to incident type 2 diabetes mellitus: A systematic review with meta-analysis. Arch. Intern. Med. 2009, 169, 2053–2063. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Han, G.; Hu, Y.; Bi, Y.; Li, L.; Zhu, D. Tea consumption and risk of type 2 diabetes: A meta-analysis of cohort studies. J. Gen. Intern. Med. 2009, 24, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.M.; Zhou, B.; Wang, Y.S.; Gong, Q.Y.; Wang, Q.M.; Yan, J.J.; Gao, W.; Wang, L.S. Black and green tea consumption and the risk of coronary artery disease: A meta-analysis. Am. J. Clin. Nutr. 2011, 93, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Goodner, K.L.; Park, J.D.; Choi, J.; Talcott, S.T. Changes in antioxidant phytochemicals and volatile composition of Camellia sinensis by oxidation during tea fermentation. Food Chem. 2011, 129, 1331–1342. [Google Scholar] [CrossRef]
- Duh, P.D.; Yen, G.C.; Yen, W.J.; Wang, B.S.; Chang, L.W. Effects of Pu-erh tea on oxidative damage and nitric oxide scavenging. J. Agric. Food Chem. 2004, 52, 8169–8176. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.Y.; Huangfu, L.T.; Dong, L.L.; Liu, S.L. Functional groups and antioxidant activities of polysaccharides from five categories of tea. Ind. Crop. Prod. 2014, 58, 31–35. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Q.; Xing, H.; Lu, X.; Zhao, L.; Qu, K.; Bi, K. Evaluation of antioxidant activity of ten compounds in different tea samples by means of an on-line HPLC-DPPH assay. Food Res. Int. 2013, 53, 847–856. [Google Scholar] [CrossRef]
- Lin, J.K.; Lin, C.L.; Liang, Y.C.; Shiau, S.Y.L.; Juan, I.M. Survey of catechins, gallic acid, and methylxanthines in green, Oolong, Pu-erh, and black Teas. J. Agric. Food Chem. 1998, 46, 3635–3642. [Google Scholar] [CrossRef]
- Surya, S.; Geethanandan, K.; Sadasivan, C. Gallic acid binding to Spatholobus parviflorus lectin provides insight to its quaternary structure forming. Int. J. Biol. Macromol. 2016, 91, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Pal, T.K. Docking analysis of gallic acid derivatives as HIV-1 protease inhibitors. Int. J. Bioinform. Res. Appl. 2015, 11, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Kongpichitchoke, T.; Hsu, J.L.; Huang, T.C. Number of hydroxyl groups on the B‑Ring of flavonoids affects their antioxidant activity and interaction with phorbol ester binding site of PKCδ C1B Domain: In vitro and in silico studies. J. Agric. Food Chem. 2015, 63, 4580–4586. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, N.; Ma, Z.Z.; Tu, P.F. Comparison of the chemical constituents of aged Pu-erh tea, ripened Pu-erh Tea, and other teas using HPLC-DAD-ESI-MSn. J. Agric. Food Chem. 2011, 59, 8754–8760. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Chen, H.; Deng, Y. Simultaneous determination of catechins, caffeine and gallic acids in green, Oolong, black and pu-erh teas using HPLC with a photodiode array detector. Talanta 2002, 57, 307–316. [Google Scholar] [CrossRef]
- You, B.R.; Park, W.H. Gallic acid-induced lung cancer cell death is related to glutathione depletion as well as reactive oxygen species increase. Toxicol. In Vitro 2010, 24, 1356–1362. [Google Scholar] [CrossRef] [PubMed]
- Weng, C.J.; Yen, G.C. Chemopreventive effects of dietary phytochemicals against cancer invasion and metastasis: Phenolic acids, monophenol, polyphenol, and their derivatives. Cancer Treat. Rev. 2012, 38, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Singh, A.; Mishra, A. Gallic acid: Molecular rival of cancer. Environ. Toxicol. Pharmacol. 2013, 35, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Nie, G.; Belton, P.S.; Tang, H.; Zhao, B. Structure-activity relationship analysis of antioxidant ability and neuroprotective effect of gallic acid derivatives. Neurochem. Int. 2006, 48, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Boly, R.; Franck, T.; Kohnen, S.; Lompo, M.; Guissou, I.P.; Dubois, J.; Serteyn, D.; Mickalad, A.M. Evaluation of antiradical and anti-inflammatory activities of ethyl acetate and butanolic subfractions of Agelanthus dodoneifolius (DC.) Polhill & Wiens (Loranthaceae) using equine myeloperoxidase and both PMA-activated neutrophils and HL-60 cells. Evid. Based Complement. Altern. Med. 2015. [Google Scholar] [CrossRef]
- Haute, G.V.; Caberlon, E.; Squizani, E.; Mesquita, F.C.; Pedrazza, L.; Martha, B.A.; Melo, D.A.S.; Cassel, E.; Czepielewski, R.S.; Bitencourt, S.; et al. Gallic acid reduces the effect of LPS on apoptosis and inhibits the formation of neutrophil extracellular traps. Toxicol. In Vitro 2015, 30, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.K.; Chen, P.C.; Ho, C.T.; Shian, S.Y.L. Inhibition of xanthine oxidase and NADPH oxidase by tea polyphenols. In Free Radicals in Food; ACS Publications: Washington, DC, USA, 2002; pp. 264–284. [Google Scholar]
- Zhang, G.; Kazanietz, M.G.; Blumberg, P.M.; Hurley, J.H. Crystal structure of the Cys2 activator-binding domain of protein kinase Cδ in complex with phorbol ester. Cell 1995, 81, 917–924. [Google Scholar] [CrossRef]
- Majhi, A.; Rahman, G.M.; Panchal, S.; Das, J. Binding of curcumin and its long chain derivatives to the activator binding domain of novel protein kinase C. Bioorg. Med. Chem. 2010, 18, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Mizukami, Y.; Sawai, Y.; Yamaguchi, Y. Simultaneous analysis of catechins, gallic Acid, strictinin, and purine alkaloids in green tea by using catechol as an internal standard. J. Agric. Food Chem. 2007, 55, 4957–4964. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.J.; Sterling, T.; Mysinger, M.M.; Bolstad, E.S.; Coleman, R.G. ZINC: A Free Tool to Discover Chemistry for Biology. J. Chem. Inf. Model. 2012, 52, 1757–1768. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are not available from the authors.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Bioactive Compound Content (mg/g) | ||||||
|---|---|---|---|---|---|---|---|
| GA | EGC | C | EGCG | EC | GCG | ECG | |
| Pu-erh Tea | 21.98 ± 1.03 | 0.02 ± 0.01 | 0.50 ± 0.09 | 0 | 1.45 ± 0.33 | 0.49 ± 0.12 | 0.22 ± 0.02 |
| Black Tea | 6.09 ± 0.06 | 0.07 ± 0.01 | 0.55 ± 0.03 | 0 | 0.95 ± 0.35 | 1.00 ± 0.05 | 0.45 ± 0.10 |
| Oolong Tea | 1.92 ± 0.56 | 2.33 ± 0.50 | 1.09 ± 0.47 | 13.34 ± 3.87 | 4.54 ± 0.14 | 1.04 ± 0.34 | 2.42 ± 0.71 |
| Green Tea | 1.67 ± 0.06 | 3.46 ± 0.06 | 2.30 ± 0.08 | 14.03 ± 0.89 | 8.86 ± 0.09 | 1.21 ± 0.04 | 1.89 ± 0.07 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kongpichitchoke, T.; Chiu, M.-T.; Huang, T.-C.; Hsu, J.-L. Gallic Acid Content in Taiwanese Teas at Different Degrees of Fermentation and Its Antioxidant Activity by Inhibiting PKCδ Activation: In Vitro and in Silico Studies. Molecules 2016, 21, 1346. https://doi.org/10.3390/molecules21101346
Kongpichitchoke T, Chiu M-T, Huang T-C, Hsu J-L. Gallic Acid Content in Taiwanese Teas at Different Degrees of Fermentation and Its Antioxidant Activity by Inhibiting PKCδ Activation: In Vitro and in Silico Studies. Molecules. 2016; 21(10):1346. https://doi.org/10.3390/molecules21101346
Chicago/Turabian StyleKongpichitchoke, Teeradate, Ming-Tzu Chiu, Tzou-Chi Huang, and Jue-Liang Hsu. 2016. "Gallic Acid Content in Taiwanese Teas at Different Degrees of Fermentation and Its Antioxidant Activity by Inhibiting PKCδ Activation: In Vitro and in Silico Studies" Molecules 21, no. 10: 1346. https://doi.org/10.3390/molecules21101346
APA StyleKongpichitchoke, T., Chiu, M.-T., Huang, T.-C., & Hsu, J.-L. (2016). Gallic Acid Content in Taiwanese Teas at Different Degrees of Fermentation and Its Antioxidant Activity by Inhibiting PKCδ Activation: In Vitro and in Silico Studies. Molecules, 21(10), 1346. https://doi.org/10.3390/molecules21101346