
The Complete Chloroplast Genome of Capsicum annuum var. glabriusculum Using Illumina Sequencing

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chloroplast Genome Assembly
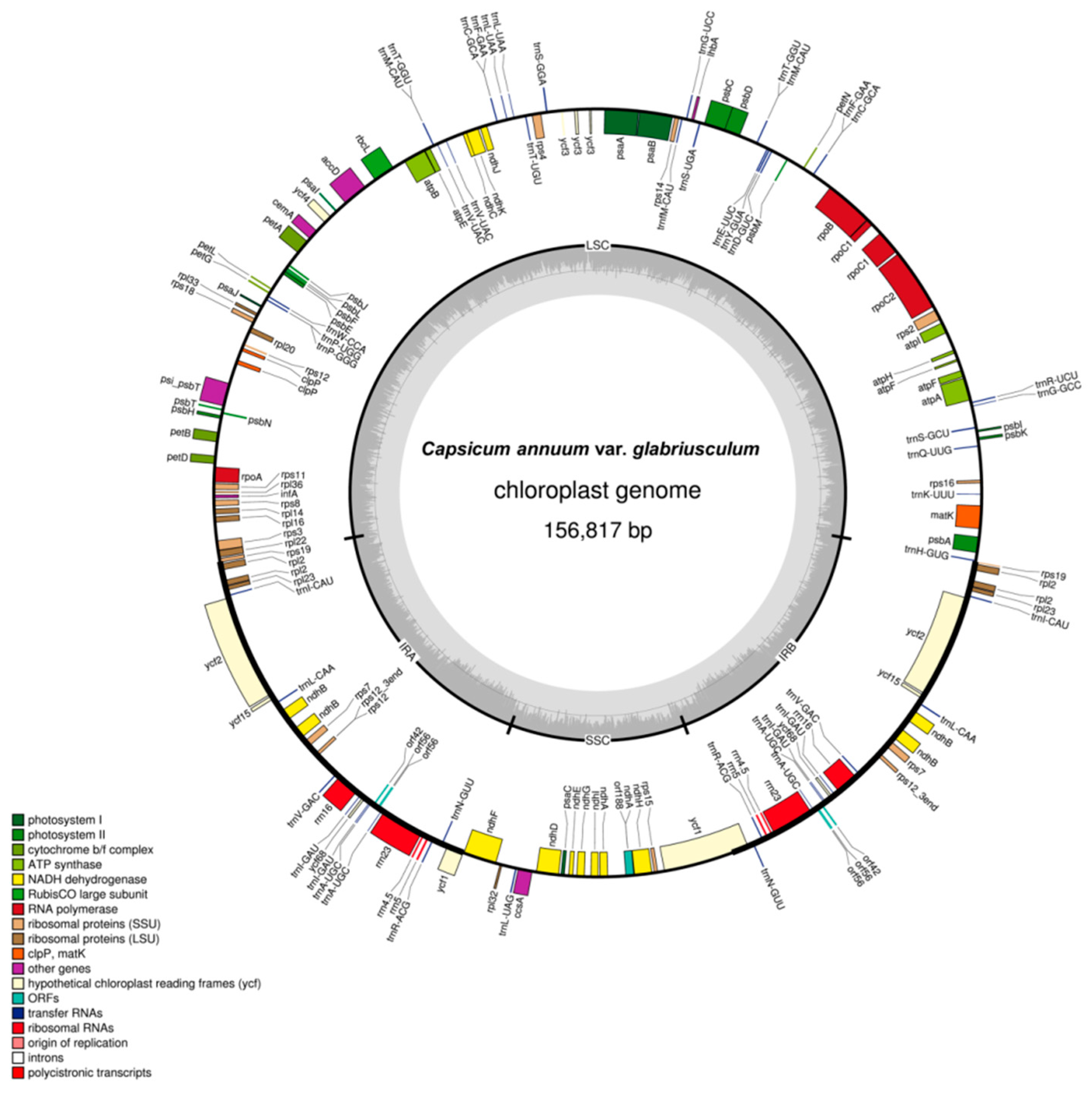
2.2. Features of the C. annuum var. glabriusculum Chloroplast Genome

{kind=link}
| Features | Chloroplast |
|---|---|
| Genome size (bp) | 156,817 |
| GC content (%) | 37.7 |
| Total number of genes | 132 |
| Protein coding genes | 87 |
| No. of rRNA genes | 8 |
| No. of tRNA genes | 37 |
| No. of gene duplicated in IR regions | 7 |
| Total introns | 12 |
| Single intron (gene) | 9 |
| Double introns (gene) | 3 |
| Single intron (tRNA) | 6 |
2.3. Discovery of SSRs and SNPs
2.4. Discussion
| Gene Products of Capsicum annuum var. glabriusculum | |
|---|---|
| Photosystem I | psaA, B, C, I, J, ycf3 2, ycf4 |
| Photosystem II | psbA, B, C, D, E, F, H, I, J, K, L, M, N, T, Z |
| Cytochrome b6/f | petA, B 1, D 1, G, L, N |
| ATP synthase | atpA, B, E, F 1, H, I |
| Rubisco | rbcL |
| NADH oxidoreductase | ndhA 1, B 1,3, C, D, E, F, G, H, I, J, K |
| Large subunit ribosomal proteins | rpl2 1,3, 14, 16 1, 20, 22, 23 3, 32, 33, 36 |
| Small subunit ribosomal proteins | rps2, 3, 4, 7 3, 8, 11, 12 2,3,4, 14, 15, 16 1, 18, 19 |
| RNA polymerase | rpoA, B, C1 1, C2 |
| Unknown function protein coding gene | ycf1 3, 2 3, 15 3 |
| Other genes | accD, ccsA, cemA, clpP 2, matK |
| rRNAs | rrn16 3, 23 3, 4.5 3, 5 3 |
| tRNAs | trnA-UGC 1,3, trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnG-UCC 1, trnG-GCC, trnH-GUG, trnI-CAU 3, trnI-GAU 1,3, trnK-UUU 1, trnL-UAA 1, trnL-UAG, trnL-CAA 3, trnfM-CAU, trnM-CAU, trnN-GUU 3, trnP-UGG, trnQ-UUG, trnR-ACG 3, trnR-UCU, trnS-GCU, trnS-GGA, trnS-UGA, trnT-GGU, trnT-UGU, trnV-UAC 1, trnV-GAC 3, trnW-CCA, trnY-GUA |
3. Experimental Section
3.1. Sampling and DNA Extraction
3.2. Library Preparation and Sequencing
3.3. Chloroplast Genome Assembly
3.4. Gene Annotation
3.5. Discovery of SNPs and SSRs
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rodriguez-Ezpeleta, N.; Brinkmann, H.; Burey, S.C.; Roure, B.; Burger, G.; Loffelhardt, W.; Bohnert, H.J.; Philippe, H.; Lang, B.F. Monophyly of primary photosynthetic eukaryotes: Green plants, red algae, and glaucophytes. Curr. Biol. 2005, 15, 1325–1330. [Google Scholar] [CrossRef] [PubMed]
- Tetlow, I.J.; Rawsthorne, S.; Raines, C.; Emes, M.J. Plastid metabolic pathways. In Annual Plant Reviews, Plastids; Moller, S.G., Ed.; Blackwell Publishing: Hoboken, NJ, USA, 2009; Volume 13, pp. 60–125. [Google Scholar]
- Ohyama, K.; Fukuzawa, H.; Kohchi, T.; Shirai, H.; Sano, T.; Sano, S.; Umesono, K.; Shiki, Y.; Takeuchi, M.; Chang, Z.; et al. Chloroplast gene organization deduced from complete sequence of liverwort marchantia polymorpha chloroplast DNA. Nature 1986, 322, 572–574. [Google Scholar] [CrossRef]
- Shinozaki, K.; Ohme, M.; Tanaka, M.; Wakasugi, T.; Hayashida, N.; Matsubayashi, T.; Zaita, N.; Chunwongse, J.; Obokata, J.; Yamaguchi-Shinozaki, K.; et al. The complete nucleotide sequence of the tobacco chloroplast genome: Its gene organization and expression. EMBO J. 1986, 5, 2043–2049. [Google Scholar] [PubMed]
- Available online: http://chloroplast.ocean.washington.edu/cpbase (accessed on 6 July 2015).
- Sugiura, M.; Hirose, T.; Sugita, M. Evolution and mechanism of translation in chloroplasts. Ann. Rev. Genet. 1998, 32, 437–459. [Google Scholar] [CrossRef] [PubMed]
- Daniell, H.; Lee, S.B.; Grevich, J.; Saski, C.; Quesada-Vargas, T.; Guda, C.; Tomkins, J.; Jansen, R.K. Complete chloroplast genome sequences of solanum bulbocastanum, solanum lycopersicum and comparative analyses with other solanaceae genomes. Theor. Appl. Genet. 2006, 112, 1503–1518. [Google Scholar] [CrossRef] [PubMed]
- Leseberg, C.H.; Duvall, M.R. The complete chloroplast genome of coix lacryma-jobi and a comparative molecular evolutionary analysis of plastomes in cereals. J. Mol. Evol. 2009, 69, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Moscone, E.A.; Scaldaferro, M.A.; Grabiele, M.; Cecchini, N.M.; Sanchez Garcia, Y.; Jarret, R.; Davina, J.R.; Ducasse, D.A.; Barboza, G.E.; Ehrendorfer, F. The evolution of chili peppers (Capsicum-Solanaceae): A cytogenetic perspective. Acta Hortic. 2007, 745, 137–170. [Google Scholar]
- Eshbaugh, W.H. History and exploitation of a serendipitous new crop discovery. In New Crops; Janick, J., Simon, J.E., Eds.; Wiley: New York, NY, USA, 1993. [Google Scholar]
- Pozzobon, M.T.; Schifino-Wittmann, M.T.; Bianchetti, L.D.B. Chromosome numbers in wild and semidomesticated Brazilian Capsicum L. (Solanaceae) species: Do x = 12 and x = 13 represent two evolutionary lines? Bot. J. Linn. Soc. 2005, 151, 259–269. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, R.; Singh, J. Cayenne/American pepper (Capsicum species). In Handbook of Herbs and Spices; Peter, K.V., Ed.; Woodhead: Cambridge, UK, 2006; pp. 299–312. [Google Scholar]
- Dong, W.; Liu, J.; Yu, J.; Wang, L.; Zhou, S. Highly variable chloroplast markers for evaluating plant phylogeny at low taxonomic levels and for DNA barcoding. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, P.M.; Graham, S.W.; Little, D.P. Choosing and using a plant DNA barcode. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Group, C.P.W.; Hollingsworth, P.M.; Forrest, L.L.; Spouge, J.L.; Hajibabaei, M.; Ratnasingham, S.; van der Bank, M.; Chase, M.W.; Cowan, R.S.; Erickson, D.L.; et al. A DNA barcode for land plants. Pro. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, D.; Sarma, R.; Shanmughanandhan, D.; Srinivasan, R.; Ramalingam, S. Evaluation of DNA barcode candidates for the discrimination of the large plant family Apocynaceae. Plant Syst. Evol. 2015, 301, 1263–1273. [Google Scholar] [CrossRef]
- Spouge, J.L.; Mariño-Ramírez, L. The practical evaluation of DNA barcode efficacy. Methods Mol. Biol. 2012, 858, 365–377. [Google Scholar] [PubMed]
- Zeng, F.C.; Gao, C.W.; Gao, L.Z. The complete chloroplast genome sequence of American bird pepper (Capsicum annuum var. glabriusculum). Mitochondrial DNA 2014. [Google Scholar] [CrossRef]
- Sebastin, R.; Young-Ah, J.; Jung-Ro, L.; Gi-An, L.; Kyung Jun, L.; Gyu-Taek, C.; Kyung-Ho, M.; Sok-Young, L.; Jong-Wook, C. The complete chloroplast genome sequence of Korean landrace “subicho” pepper (Capsicum annuum var. annuum). Plant Breed. Biotechnol. 2015, 3, 88–94. [Google Scholar]
- Jo, Y.D.; Park, J.; Kim, J.; Song, W.; Hur, C.G.; Lee, Y.H.; Kang, B.C. Complete sequencing and comparative analyses of the pepper (Capsicum annuum L.) plastome revealed high frequency of tandem repeats and large insertion/deletions on pepper plastome. Plant Cell Rep. 2011, 30, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.H.; Gui, S.T.; Quan, Z.W.; Pan, L.; Wang, S.Z.; Ke, W.D.; Liang, D.Q.; Ding, Y. A precise chloroplast genome of Nelumbo nucifera (Nelumbonaceae) evaluated with Sanger, Illumina Miseq, and Pacbio RS II sequencing platforms: Insight into the plastid evolution of basal eudicots. BMC Plant Biol. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, M. The chloroplast genome. Plant Mol. Biol. 1992, 19, 149–168. [Google Scholar] [CrossRef] [PubMed]
- Cardle, L.; Ramsay, L.; Milbourne, D.; Macaulay, M.; Marshall, D.; Waugh, R. Computational and experimental characterization of physically clustered simple sequence repeats in plants. Genetics 2000, 156, 847–854. [Google Scholar] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with burrows-wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Proc, G.P.D. The sequence alignment/map format and samtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Exploring single-sample SNP and INDEL calling with whole-genome de novo assembly. Bioinformatics 2012, 28, 1838–1844. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples are not available.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raveendar, S.; Na, Y.-W.; Lee, J.-R.; Shim, D.; Ma, K.-H.; Lee, S.-Y.; Chung, J.-W. The Complete Chloroplast Genome of Capsicum annuum var. glabriusculum Using Illumina Sequencing. Molecules 2015, 20, 13080-13088. https://doi.org/10.3390/molecules200713080
Raveendar S, Na Y-W, Lee J-R, Shim D, Ma K-H, Lee S-Y, Chung J-W. The Complete Chloroplast Genome of Capsicum annuum var. glabriusculum Using Illumina Sequencing. Molecules. 2015; 20(7):13080-13088. https://doi.org/10.3390/molecules200713080
Chicago/Turabian StyleRaveendar, Sebastin, Young-Wang Na, Jung-Ro Lee, Donghwan Shim, Kyung-Ho Ma, Sok-Young Lee, and Jong-Wook Chung. 2015. "The Complete Chloroplast Genome of Capsicum annuum var. glabriusculum Using Illumina Sequencing" Molecules 20, no. 7: 13080-13088. https://doi.org/10.3390/molecules200713080
APA StyleRaveendar, S., Na, Y.-W., Lee, J.-R., Shim, D., Ma, K.-H., Lee, S.-Y., & Chung, J.-W. (2015). The Complete Chloroplast Genome of Capsicum annuum var. glabriusculum Using Illumina Sequencing. Molecules, 20(7), 13080-13088. https://doi.org/10.3390/molecules200713080