
Protein-Carbohydrate Interaction between Sperm and the Egg-Coating Envelope and Its Regulation by Dicalcin, a Xenopus laevis Zona Pellucida Protein-Associated Protein

Abstract
:1. Introduction
2. Brief Introduction to Fertilization
3. Sperm Storage in the Oviduct
Spermadhesin and Oligosaccharides of the Oviductal Epithelial Cells
4. Sperm Recognition of the ZP
4.1. Structure of the ZP
4.2. ZP Proteins of the Extracellular Egg-Coating Envelope
4.3. Carbohydrates of the ZP

{kind=link}
{kind=link}
{kind=link}
| N-Linked Glycan | O-Linked Glycan | Major Terminal Residues | Lectin-Staining Intensity | |
|---|---|---|---|---|
| Human | NeuAcα2-3Galβ1-4(Fucα1-3)GlcNAc | NeuAcα2-3Galβ1-4(Fucα1-3)GlcNAc | NeuAc from sialyl-LewisX | WGA > ConA, MPA |
| Mouse | High-mannose type Complex type | Core type-1 (Minor) Core type-2 (Major) | GlcNAc Gal from LacNAc NeuAc from Sd1 antigen | GS-I, DBA > WGA, RCA-I, PNA |
| Frog | High-mannose type Complex type | Trisaccharides (GalNAc-βGal-βGal) | GlcNAc | WGA > ConA = MPA |
4.4. Carbohydrate Structures Determining Gamete Recognition
4.5. Interactive Proteins with Carbohydrates in the ZP
4.5.1. FUT5 and N-Acetyllactosamines
4.5.2. Galactosyltransferase (GalT) and N-Acetylgulucosamine on ZPC
4.5.3. ZP3 Receptor (ZP3R, Formerly Called sp56) and O-Linked Oligosaccharide on ZPC
4.5.4. Pro/Acrosin and Polysaccharide of the ZP
5. Dicalcin as An Inherent Suppressive Factor for Fertilization in Xenopus laevis
5.1. Structure and Distribution of Dicalcin
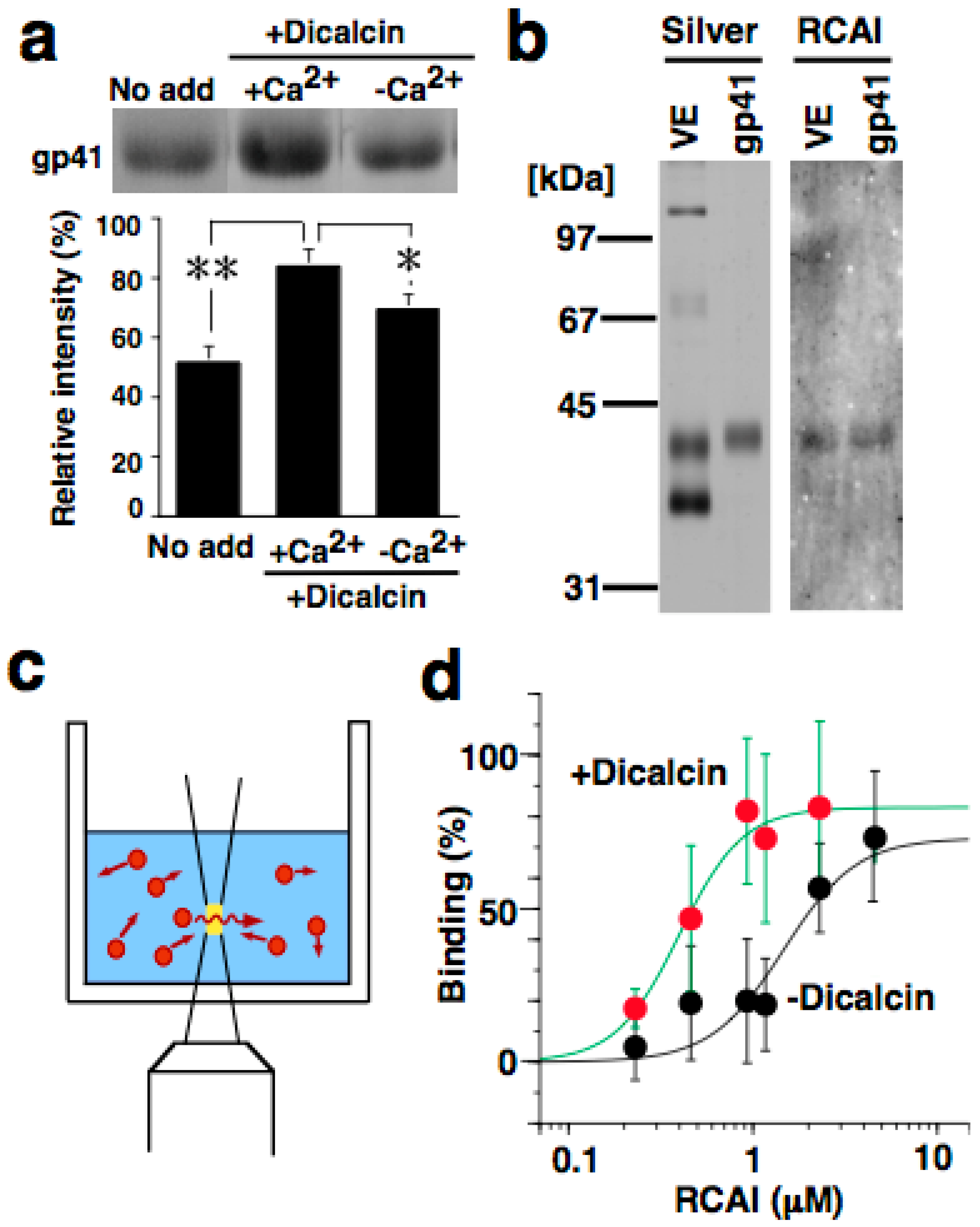
5.2. Binding of Dicalcin to Target Proteins
5.3. Effects of Dicalcin on Fertilization Success
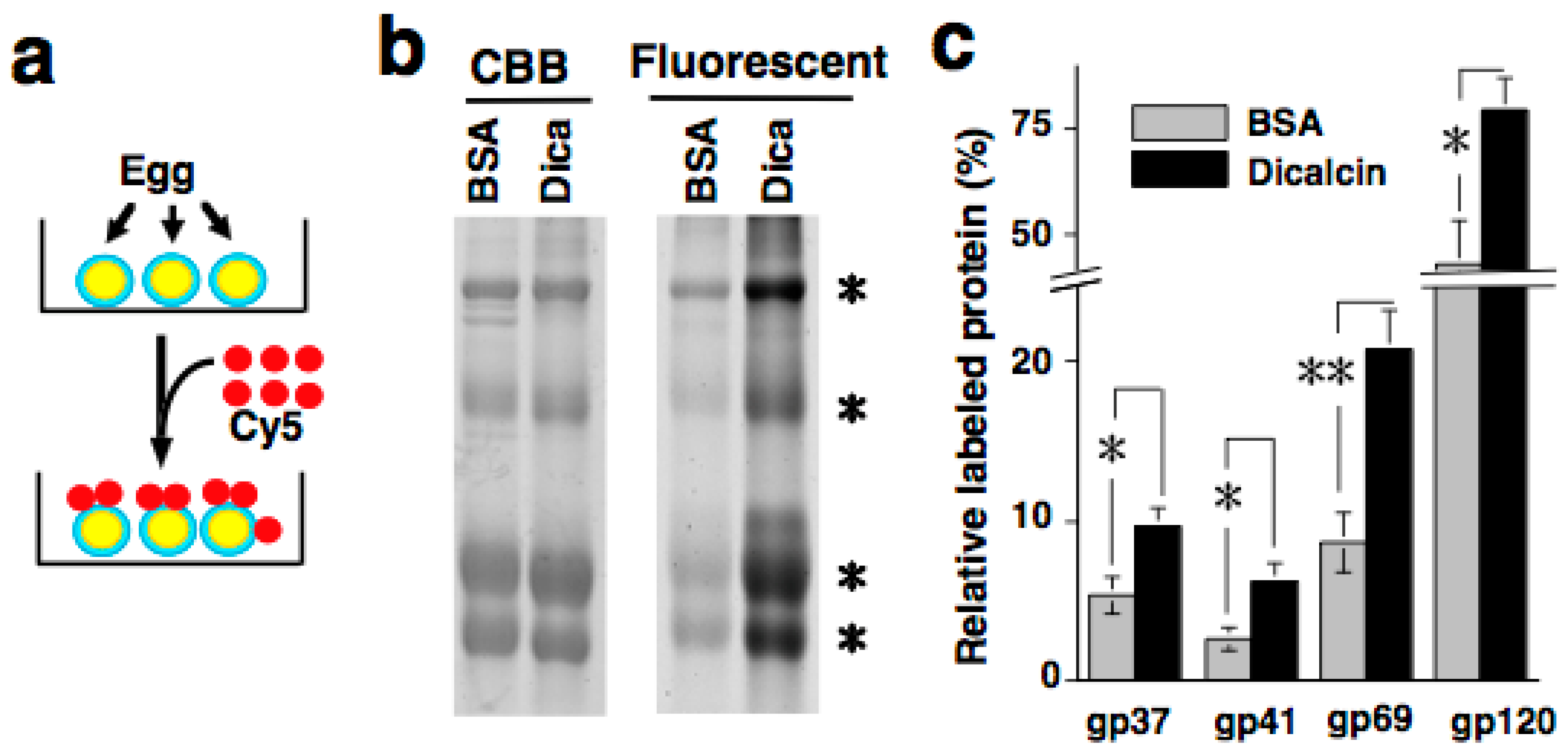
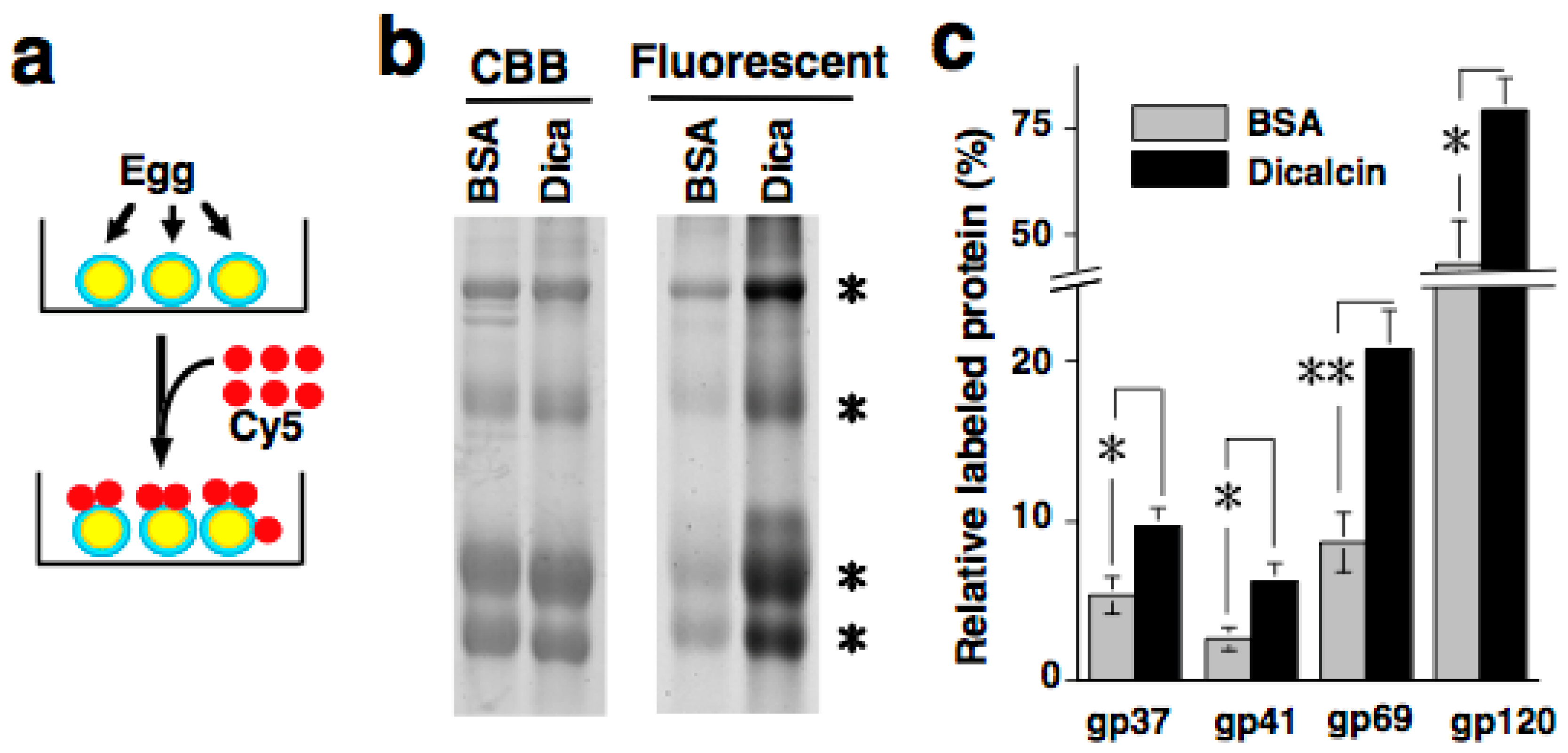
6. Regulation of Oligosaccharide-Lectin Interaction by Dicalcin through Its Binding to ZPC
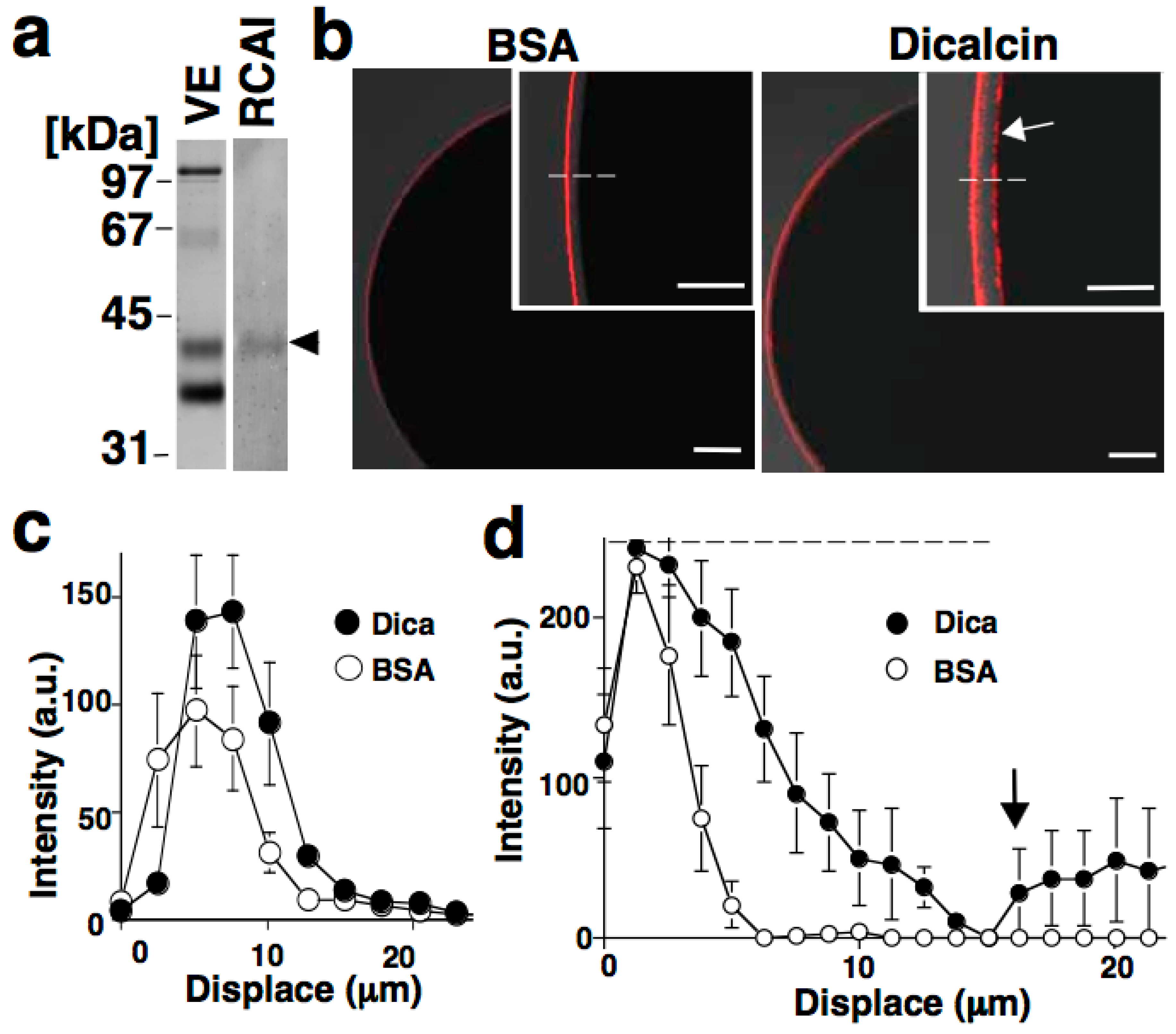
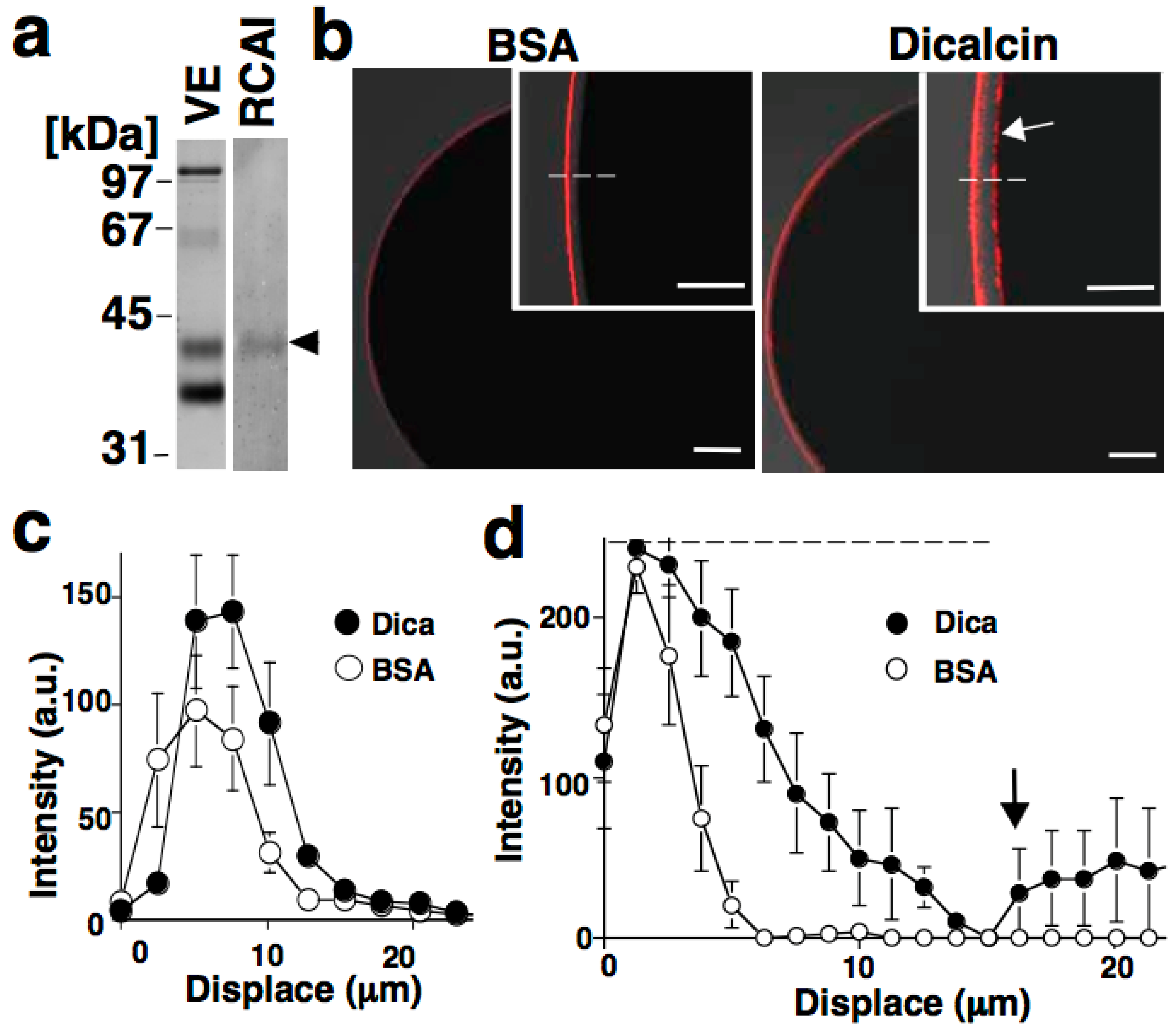
6.1. Sperm Binding to Xenopus Laevis ZP
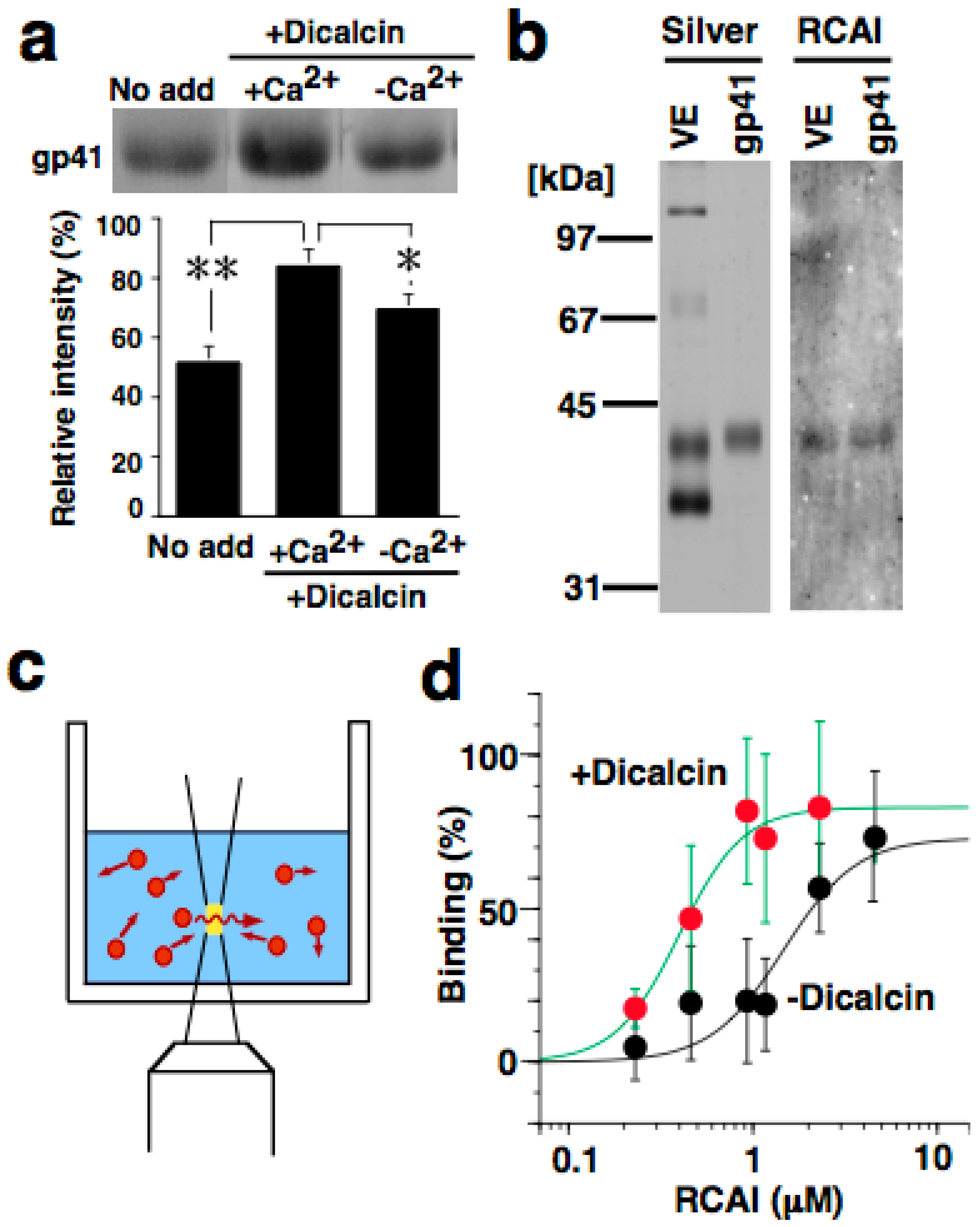
6.2. Regulation of Oligosaccharide-Lectin Interaction by Dicalcin
7. Biological Significance of Dicalcin
8. Future Perspectives
Acknowledgments
Abbreviations
| αGal | α-galactose |
| βGal | β-galactose |
| GlcNAc | N-acetylglucosamine |
| GalNAc | N-acetylgalactosamine |
| NeuAc | N-acetylneuraminic acid |
Conflicts of Interest
References
- Soupart, P.; Clewe, T.H. Sperm penetration of rabbit zona pellucida inhibited by treatment of ova with neuraminidase. Fertil. Steril. 1965, 16, 677–689. [Google Scholar] [PubMed]
- Tsuzuki, H.; Aketa, K. A study on the possible significance of carbohydrate moiety in the sperm-binding protein from sea urchin egg. Exp. Cell Res. 1969, 55, 43–45. [Google Scholar] [CrossRef]
- Clark, G.F. Molecular models for mouse sperm-oocyte binding. Glycobiology 2011, 21, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Vacquier, V.D. The quest for the sea urchin egg receptor for sperm. Biochem. Biophys. Res. Commun. 2012, 425, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Visconti, P.E.; Florman, H.M. Mechanisms of sperm-egg interactions: Between sugars and broken bonds. Sci. Signal. 2010, 3, e35. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, J.L. Anuran and pig egg zona pellucida glycoproteins in fertilization and early development. Int. J. Dev. Biol. 2008, 52, 683–701. [Google Scholar] [CrossRef] [PubMed]
- Töpfer-Petersen, E. Carbohydrate-based interactions on the route of a spermatozoon to fertilization. Hum. Reprod. Update 1999, 5, 314–329. [Google Scholar] [CrossRef] [PubMed]
- Macek, M.B.; Shur, B.D. Protein-carbohydrate complementarity in mammalian gamete recognition. Gamete Res. 1988, 20, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Tulsiani, D.R.; Yoshida-Komiya, H.; Araki, Y. Mammalian fertilization: A carbohydrate-mediated event. Biol. Reprod. 1997, 57, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Garbers, D.L. Molecular basis of fertilization. Annu. Rev. Biochem. 1989, 58, 719–742. [Google Scholar] [CrossRef] [PubMed]
- Bleil, J.D.; Wassarman, P.M. Sperm-egg interactions in the mouse: sequence of events and induction of the acrosome reaction by a zona pellucida glycoprotein. Dev. Biol. 1983, 95, 317–324. [Google Scholar] [CrossRef]
- Miwa, N.; Ogawa, M.; Shinmyo, Y.; Hiraoka, Y.; Takamatsu, K.; Kawamura, S. Dicalcin inhibits fertilization through its binding to a glycoprotein in the egg envelope in Xenopus laevis. J. Biol. Chem. 2010, 285, 15627–15636. [Google Scholar] [CrossRef] [PubMed]
- Gahlay, G.; Gauthier, L.; Baibakov, B.; Epifano, O.; Dean, J. Gamete recognition in mice depends on the cleavage status of an egg’s zona pellucida protein. Science 2010, 329, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Yanagimachi, R. Mammalian fertilization. In The Physiology of Reproduction; Knobil, E., Neill, J.D., Eds.; Raven Press Ltd: New York, NY, USA, 1994; pp. 189–317. [Google Scholar]
- Töpfer-Petersen, E.; Romero, A.; Varela, P.F.; Ekhlasi-Hundrieser, M.; Dostàlovà, Z.; Sanz, L.; Calvete, J.J. Spermadhesins: A new protein family. Facts, hypotheses and perspectives. Andrologia 1998, 30, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Busso, D.; Goldweic, N.M.; Hayashi, M.; Kasahara, M.; Cuasnicú, P.S. Evidence for the involvement of testicular protein CRISP2 in mouse sperm-egg fusion. Biol. Reprod. 2007, 76, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Frenette, G.; Girouard, J.; D’Amours, O.; Allard, N.; Tessier, L.; Sullivan, R. Characterization of two distinct populations of epididymosomes collected in the intraluminal compartment of the bovine cauda epididymis. Biol. Reprod. 2010, 83, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Visconti, P.E.; Krapf, D.; de la Vega-Beltrán, J.L.; Acevedo, J.J.; Darszon, A. Ion channels, phosphorylation and mammalian sperm capacitation. Asian J. Androl. 2011, 13, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.L. Factors regulating sperm capacitation. Syst. Biol. Reprod. Med. 2010, 56, 334–348. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Navarro, B.; Perez, G.; Jackson, A.C.; Hsu, S.; Shi, Q.; Tilly, J.L.; Clapham, D.E. A sperm ion channel required for sperm motility and male fertility. Nature 2001, 413, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.T. The modulation of sperm function by the oviductal epithelium. Biol. Reprod. 1998, 58, 1102–1104. [Google Scholar] [CrossRef] [PubMed]
- Suarez, S.S. The oviductal sperm reservoir in mammals: Mechanisms of formation. Biol. Reprod. 1998, 58, 1105–1107. [Google Scholar] [CrossRef] [PubMed]
- Ekhlasi-Hundrieser, M.; Schäfer, B.; Philipp, U.; Kuiper, H.; Leeb, T.; Mehta, M.; Kirchhoff, C.; Töpfer-Petersen, E. Sperm-binding fibronectin type II-module proteins are genetically linked and functionally related. Gene 2007, 392, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Ekhlasi-Hundrieser, M.; Gohr, K.; Wagner, A.; Tsolova, M.; Petrunkina, A.; Töpfer-Petersen, E. Spermadhesin AQN1 is a candidate receptor molecule involved in the formation of the oviductal sperm reservoir in the pig. Biol. Reprod. 2005, 73, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Dostálová, Z.; Calvete, J.J.; Sanz, L.; Töpfer-Petersen, E. Boar spermadhesin AWN-1. Oligosaccharide and zona pellucida binding characteristics. Eur. J. Biochem. 1995, 230, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Carrera, E.; Sanz, L.; Töpfer-Petersen, E. Boar spermadhesins AQN-1 and AQN-3: oligosaccharide and zona pellucida binding characteristics. Biol. Chem. 1996, 377, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Wassarman, P.M.; Litscher, E.S. Mammalian fertilization: The egg’s multifunctional zona pellucida. Int. J. Dev. Biol. 2008, 52, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Yamagami, K.; Hamazaki, T.S.; Yasumasu, S.; Masuda, K.; Iuchi, I. Molecular and cellular basis of formation, hardening, and breakdown of the egg envelope in fish. Int. Rev. Cytol. 1992, 136, 51–92. [Google Scholar] [PubMed]
- Wassarman, P.M.; Jovine, L.; Litscher, E.S. A profile of fertilization in mammals. Nat. Cell Biol. 2001, 3, E59–E64. [Google Scholar] [CrossRef] [PubMed]
- Larabell, C.A.; Chandler, D.E. Quick-freeze, deep-etch, rotary-shadow views of the extracellular matrix and cortical cytoskeleton of Xenopus laevis eggs. J. Electron Microsc. Tech. 1989, 13, 228–243. [Google Scholar] [CrossRef] [PubMed]
- Larabell, C.A.; Chandler, D.E. The extracellular matrix of Xenopus laevis eggs: A quick-freeze, deep-etch analysis of its modification at fertilization. J. Cell Biol. 1988, 107, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Litscher, E.S.; Wassarman, P.M. Egg extracellular coat proteins: from fish to mammals. Histol. Histopathol. 2007, 22, 337–347. [Google Scholar] [PubMed]
- Repin, V.S.; Akimova, I.M. A study of the protein composition of the zona pellucida of mammalian oocytes and zygotes by a method of microelectrophoresis in polyacrylamide gel. Biokhimiia 1976, 41, 50–57. [Google Scholar] [PubMed]
- Bleil, J.D.; Wassarman, P.M. Synthesis of zona pellucida proteins by denuded and follicle-enclosed mouse oocytes during culture in vitro. Proc. Natl. Acad. Sci. USA 1980, 77, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, J.L. Comparative structural and antigenic properties of zona pellucida glycoproteins. J. Reprod. Fertil. Suppl. 1996, 50, 9–17. [Google Scholar] [PubMed]
- Epifano, O.; Liang, L.F.; Familari, M.; Moos, M.C., Jr.; Dean, J. Coordinate expression of the three zona pellucida genes during mouse oogenesis. Development 1995, 121, 1947–1956. [Google Scholar] [PubMed]
- Harris, J.D.; Hibler, D.W.; Fontenot, G.K.; Hsu, K.T.; Yurewicz, E.C.; Sacco, A.G. Cloning and characterization of zona pellucida genes and cDNAs from a variety of mammalian species: The ZPA, ZPB and ZPC gene families. DNA Seq. 1994, 4, 361–393. [Google Scholar] [CrossRef] [PubMed]
- Plaza, S.; Chanut-Delalande, H.; Fernandes, I.; Wassarman, P.M.; Payre, F. From A to Z: Apical structures and zona pellucida-domain proteins. Trends Cell Biol. 2010, 20, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Spargo, S.C.; Hope, R.M. Evolution and nomenclature of the zona pellucida gene family. Biol. Reprod. 2003, 68, 358–362. [Google Scholar] [CrossRef] [PubMed]
- Boja, E.S.; Hoodbhoy, T.; Fales, H.M.; Dean, J. Structural characterization of native mouse zona pellucida proteins using mass spectrometry. J. Biol. Chem. 2003, 278, 34189–34202. [Google Scholar] [CrossRef] [PubMed]
- Chalabi, S.; Panico, M.; Sutton-Smith, M.; Haslam, S.M.; Patankar, M.S.; Lattanzio, F.A.; Morris, H.R.; Clark, G.F.; Dell, A. Differential O-glycosylation of a conserved domain expressed in murine and human ZP3. Biochemistry 2006, 45, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Bork, P.; Sander, C. A large domain common to sperm receptors (Zp2 and Zp3) and TGF-beta type III receptor. FEBS Lett. 1992, 300, 237–240. [Google Scholar] [PubMed]
- Jovine, L.; Darie, C.C.; Litscher, E.S.; Wassarman, P.M. Zona pellucida domain proteins. Annu. Rev. Biochem. 2005, 74, 83–114. [Google Scholar] [CrossRef] [PubMed]
- Monne, M.; Han, L.; Schwend, T.; Burendahi, S.; Jovine, L. Crystal structure of the ZP-N domain of ZP3 reveals the core fold of animal egg coats. Nature 2008, 456, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Monné, M.; Okumura, H.; Schwend, T.; Cherry, A.L.; Flot, D.; Matsuda, T.; Jovine, L. Insights into egg coat assembly and egg-sperm interaction from the X-ray structure of full-length ZP3. Cell 2010, 143, 404–415. [Google Scholar] [CrossRef] [PubMed]
- Williams, Z.; Litscher, E.S.; Jovine, L.; Wassarman, P.M. Polypeptide encoded by mouse ZP3 exon-7 is necessary and sufficient for binding of mouse sperm in vitro. J. Cell Physiol. 2006, 207, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Millar, S.E.; Chamow, S.M.; Baur, A.W.; Oliver, C.; Robey, F.; Dean, J. Vaccination with a synthetic zona pellucida peptide produces long-term contraception in female mice. Science 1989, 246, 935–938. [Google Scholar] [CrossRef] [PubMed]
- Skutelsky, E.; Ranen, E.; Shalgi, R. Variations in the distribution of sugar residues in the zona pellucida as possible species-specific determinants of mammalian oocytes. J. Reprod. Fertil. 1994, 100, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Talevi, R.; Gualtieri, R.; Tartaglione, G.; Fortunato, A. Heterogeneity of the zona pellucida carbohydrate distribution in human oocytes failing to fertilize in vitro. Hum. Reprod. 1997, 12, 2773–2780. [Google Scholar] [CrossRef] [PubMed]
- Pang, P.C.; Chiu, P.C.; Lee, C.L.; Chang, L.Y.; Panico, M.; Morris, H.R.; Haslam, S.M.; Khoo, K.H.; Clark, G.F.; Yeung, W.S.; et al. Human sperm binding is mediated by the sialyl-Lewis(x) oligosaccharide on the zona pellucida. Science 2011, 333, 1761–1764. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.F. A role for carbohydrate recognition in mammalian sperm-egg binding. Biochem. Biophys. Res. Commun. 2014, 450, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Katsumata, T.; Noguchi, S.; Yonezawa, N.; Tanokura, M.; Nakano, M. Structural characterization of the N-linked carbohydrate chains of the zona pellucida glycoproteins from bovine ovarian and fertilized eggs. Eur. J. Biochem. 1996, 240, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Easton, R.L.; Patankar, M.S.; Lattanzio, F.A.; Leaven, T.H.; Morris, H.R.; Clark, G.F.; Dell, A. Structural analysis of murine zona pellucida glycans. J. Biol. Chem. 2000, 275, 7731–7742. [Google Scholar] [CrossRef] [PubMed]
- Vo, L.H.; Hedrick, J.L. Independent and hetero-oligomeric-dependent sperm binding to egg envelope glycoprotein ZPC in Xenopus laevis. Biol. Reprod. 2000, 62, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.F.; Patankar, M.S.; Hinsch, K.D.; Oehninger, S. New concepts in human sperm-zona pellucida interaction. Hum. Reprod. 1995, 10, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Vo, L.H.; Yen, T.Y.; Macher, B.A.; Hedrick, J.L. Identification of the ZPC oligosaccharide ligand involved in sperm binding and the glycan structures of Xenopus laevis vitelline envelope glycoproteins. Biol. Reprod. 2003, 69, 1822–1830. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Gong, H.; Lennarz, W. Xenopus laevis sperm receptor gp69/64 glycoprotein is a homolog of the mammalian sperm receptor ZP2. Proc. Natl. Acad. Sci. USA 1999, 96, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Costache, M.; Apoil, P.A.; Cailleau, A.; Elmgren, A.; Larson, G.; Henry, S.; Blancher, A.; Iordachescu, D.; Oriol, R.; Mollicone, R. Evolution of fucosyltransferase genes in vertebrates. J. Biol. Chem. 1997, 272, 29721–29728. [Google Scholar] [CrossRef] [PubMed]
- Pykäri, M.; Toivonen, S.; Natunen, J.; Niemela, R.; Salminen, H.; Aitio, O.; Ekström, M.; Parmanne, P.; Välimäki, M.; Alais, J.; et al. The acceptor and site specificity of alpha 3-fucosyltransferase V. High reactivity of the proximal and low of the distal galbeta 1–4GlcNAc unit in i-type polylactosamines. J. Biol. Chem. 2000, 275, 40057–40063. [Google Scholar] [CrossRef] [PubMed]
- De Vries, T.; Knegtel, R.M.; Holmes, E.H.; Macher, B.A. Fucosyltransferases: Structure/function studies. Glycobiology 2001, 11, 119R–128R. [Google Scholar] [CrossRef] [PubMed]
- Chiu, P.C.; Chung, M.K.; Koistinen, R.; Koistinen, H.; Seppala, M.; Ho, P.C.; Ng, E.H.; Lee, K.F.; Yeung, W.S. Glycodelin-A interacts with fucosyltransferase on human sperm plasma membrane to inhibit spermatozoa-zona pellucida binding. J. Cell Sci. 2007, 120, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Shur, B.D.; Hall, N.G. A role for mouse sperm surface galactosyltransferase in sperm binding to the egg zona pellucida. J. Cell Biol. 1982, 95, 574–579. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Shur, B.D. Sperm from beta 1,4-galactosyltransferase-null mice are refractory to ZP3-induced acrosome reactions and penetrate the zona pellucida poorly. Development 1997, 124, 4121–4131. [Google Scholar] [PubMed]
- Gong, X.; Dubois, D.H.; Miller, D.J.; Shur, B.D. Activation of a G protein complex by aggregation of beta-1,4-galactosyltransferase on the surface of sperm. Science 1995, 269, 1718–1721. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.J.; Macek, M.B.; Shur, B.D. Complementarity between sperm surface beta-1,4-galactosyltransferase and egg-coat ZP3 mediates sperm-egg binding. Nature 1992, 357, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.; Le, T.; Palacios, M.; Bookbinder, L.H.; Wassarman, P.M.; Suzuki, F.; Bleil, J.D. Sperm-egg recognition in the mouse: Characterization of sp56, a sperm protein having specific affinity for ZP3. J. Cell Biol. 1994, 125, 867–878. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.A.; Friday, B.B.; Maulit, M.T.; Blobel, C.; Winfrey, V.P.; Olson, G.E.; Kim, K.S.; Gerton, G.L. AM67, a secretory component of the guinea pig sperm acrosomal matrix, is related to mouse sperm protein sp56 and the complement component 4-binding proteins. J. Biol. Chem. 1997, 272, 12714–12722. [Google Scholar] [CrossRef] [PubMed]
- He, X.B.; Yan, Y.C.; Li, Y.P.; Koide, S.S. Cloning of rat sp56, the homologue of mouse sperm ZP3 receptor-sp56. Cell Res. 2003, 13, 121–129. [Google Scholar] [PubMed]
- Kim, K.S.; Foster, J.A.; Gerton, G.L. Differential release of guinea pig sperm acrosomal components during exocytosis. Biol. Reprod. 2001, 64, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Muro, Y.; Buffone, M.G.; Okabe, M.; Gerton, G.L. Function of the acrosomal matrix: Zona pellucida 3 receptor (ZP3R/sp56) is not essential for mouse fertilization. Biol. Reprod. 2012, 86, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Nayernia, K.; Adham, I.; Kremling, H.; Reim, K.; Schlicker, M.; Schlüter, G.; Engel, W. Stage and developmental specific gene expression during mammalian spermatogenesis. Int. J. Dev. Biol. 1996, 40, 379–383. [Google Scholar] [PubMed]
- Klemm, U.; Flake, A.; Engel, W. Rat sperm acrosin: cDNA sequence, derived primary structure and phylogenetic origin. Biochim. Biophys. Acta. 1991, 1090, 270–272. [Google Scholar] [CrossRef]
- Jones, R. Interaction of zona pellucida glycoproteins, sulphated carbohydrates and synthetic polymers with proacrosin, the putative egg-binding protein from mammalian spermatozoa. Development 1991, 111, 1155–1163. [Google Scholar] [PubMed]
- Töpfer-Petersen, E.; Henschen, A. Acrosin shows zona and fucose binding, novel properties for a serine proteinase. FEBS Lett. 1987, 226, 38–42. [Google Scholar] [CrossRef]
- Baba, T.; Azuma, S.; Kashiwabara, S.; Toyoda, Y. Sperm from mice carrying a targeted mutation of the acrosin gene can penetrate the oocyte zona pellucida and effect fertilization. J. Biol. Chem. 1994, 269, 31845–31849. [Google Scholar] [PubMed]
- Kawano, N.; Kang, W.; Yamashita, M.; Koga, Y.; Yamazaki, T.; Hata, T.; Miyado, K.; Baba, T. Mice lacking two sperm serine proteases, ACR and PRSS21, are subfertile, but the mutant sperm are infertile in vitro. Biol. Reprod. 2010, 83, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Miwa, N.; Shinmyo, Y.; Kawamura, S. Cloning and characterization of Xenopus dicalcin, a novel S100-like calcium-binding protein in Xenopus eggs. DNA Seq. 2007, 18, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Donato, R.; Cannon, B.R.; Sorci, G.; Riuzzi, F.; Hsu, K.; Weber, D.J.; Geczy, C.L. Functions of S100 proteins. Curr. Mol. Med. 2013, 13, 24–57. [Google Scholar] [CrossRef] [PubMed]
- Heizmann, C.W.; Fritz, G.; Schäfer, B.W. S100 proteins: Structure, functions and pathology. Front. Biosci. 2002, 7, d1356–d1368. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Miwa, N.; Kawamura, S.; Sohma, H.; Nitta, K.; Matsushima, N. Molecular modeling of single polypeptide chain of calcium-binding protein p26olf from dimeric S100B(betabeta). Protein Eng. 1999, 12, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Miwa, N.; Shinmyo, Y.; Kawamura, S. Calcium-binding by p26olf, an S100-like protein in the frog olfactory epithelium. Eur. J. Biochem. 2001, 268, 6029–6036. [Google Scholar] [CrossRef] [PubMed]
- Miwa, N.; Kobayashi, M.; Takamatsu, K.; Kawamura, S. Purification and molecular cloning of a novel calcium-binding protein, p26olf, in the frog olfactory epithelium. Biochem. Biophys. Res. Commun. 1998, 251, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Miwa, N.; Kawamura, S. Frog p26olf, a molecule with two S100-like regions in a single peptide. Microsc. Res. Tech. 2003, 15, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Nickel, W. The unconventional secretory machinery of fibroblast growth factor 2. Traffic 2011, 12, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Miwa, N.; Uebi, T.; Kawamura, S. Characterization of p26olf, a novel calcium-binding protein in the frog olfactory epithelium. J. Biol. Chem. 2000, 275, 27245–27249. [Google Scholar] [CrossRef] [PubMed]
- Uebi, T.; Miwa, N.; Kawamura, S. Comprehensive interaction of dicalcin with annexins in frog olfactory and respiratory cilia. FEBS J. 2007, 274, 4863–4876. [Google Scholar] [CrossRef] [PubMed]
- Miwa, N.; Uebi, T.; Kawamura, S. S100-annexin complexes—Biology of conditional association. FEBS J. 2008, 275, 4945–4955. [Google Scholar] [CrossRef] [PubMed]
- Kubo, H.; Matsushita, M.; Kotani, M.; Kawasaki, H.; Saido, T.C.; Kawashima, S.; Katagiri, C.; Suzuki, A. Molecular basis for oviductin-mediated processing from gp43 to gp41, the predominant glycoproteins of Xenopus laevis egg envelopes. Dev. Gen. 1999, 25, 123–129. [Google Scholar] [CrossRef]
- Tian, J.; Gong, H.; Thomsen, G.H.; Lennarz, W.J. Xenopus laevis sperm-egg adhesion is regulated by modifications in the sperm receptor and the egg vitelline envelope. Dev. Biol. 1997, 187, 143–153. [Google Scholar] [PubMed]
- Kimura, H.; Matsuda, J.; Ogura, A.; Asano, T.; Naiki, M. Affinity binding of hamster oviductin to spermatozoa and its influence on in vitro fertilization. Mol. Reprod. Dev. 1994, 39, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Mathialagan, N.; Walters, E.; Mao, J.; Lai, L.; Becker, D.; Li, W.; Critser, J.; Prather, R.S. Osteopontin reduces polyspermy during in vitro fertilization of porcine oocytes. Biol. Reprod. 2006, 75, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Suarez, S.S.; Pacey, A.A. Sperm transport in the female reproductive tract. Hum. Reprod. Update 2006, 12, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Töpfer-Petersen, E.; Ekhlasi-Hundrieser, M.; Tsolova, M. Glycobiology of fertilization in the pig. Int. J. Dev. Biol. 2008, 52, 717–736. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.F. The mammalian zona pellucida: A matrix that mediates both gamete binding and immune recognition? Syst. Biol. Reprod. Med. 2010, 56, 349–364. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miwa, N. Protein-Carbohydrate Interaction between Sperm and the Egg-Coating Envelope and Its Regulation by Dicalcin, a Xenopus laevis Zona Pellucida Protein-Associated Protein. Molecules 2015, 20, 9468-9486. https://doi.org/10.3390/molecules20059468
Miwa N. Protein-Carbohydrate Interaction between Sperm and the Egg-Coating Envelope and Its Regulation by Dicalcin, a Xenopus laevis Zona Pellucida Protein-Associated Protein. Molecules. 2015; 20(5):9468-9486. https://doi.org/10.3390/molecules20059468
Chicago/Turabian StyleMiwa, Naofumi. 2015. "Protein-Carbohydrate Interaction between Sperm and the Egg-Coating Envelope and Its Regulation by Dicalcin, a Xenopus laevis Zona Pellucida Protein-Associated Protein" Molecules 20, no. 5: 9468-9486. https://doi.org/10.3390/molecules20059468
APA StyleMiwa, N. (2015). Protein-Carbohydrate Interaction between Sperm and the Egg-Coating Envelope and Its Regulation by Dicalcin, a Xenopus laevis Zona Pellucida Protein-Associated Protein. Molecules, 20(5), 9468-9486. https://doi.org/10.3390/molecules20059468