
Almond Skin Inhibits HSV-2 Replication in Peripheral Blood Mononuclear Cells by Modulating the Cytokine Network

Abstract
:1. Introduction
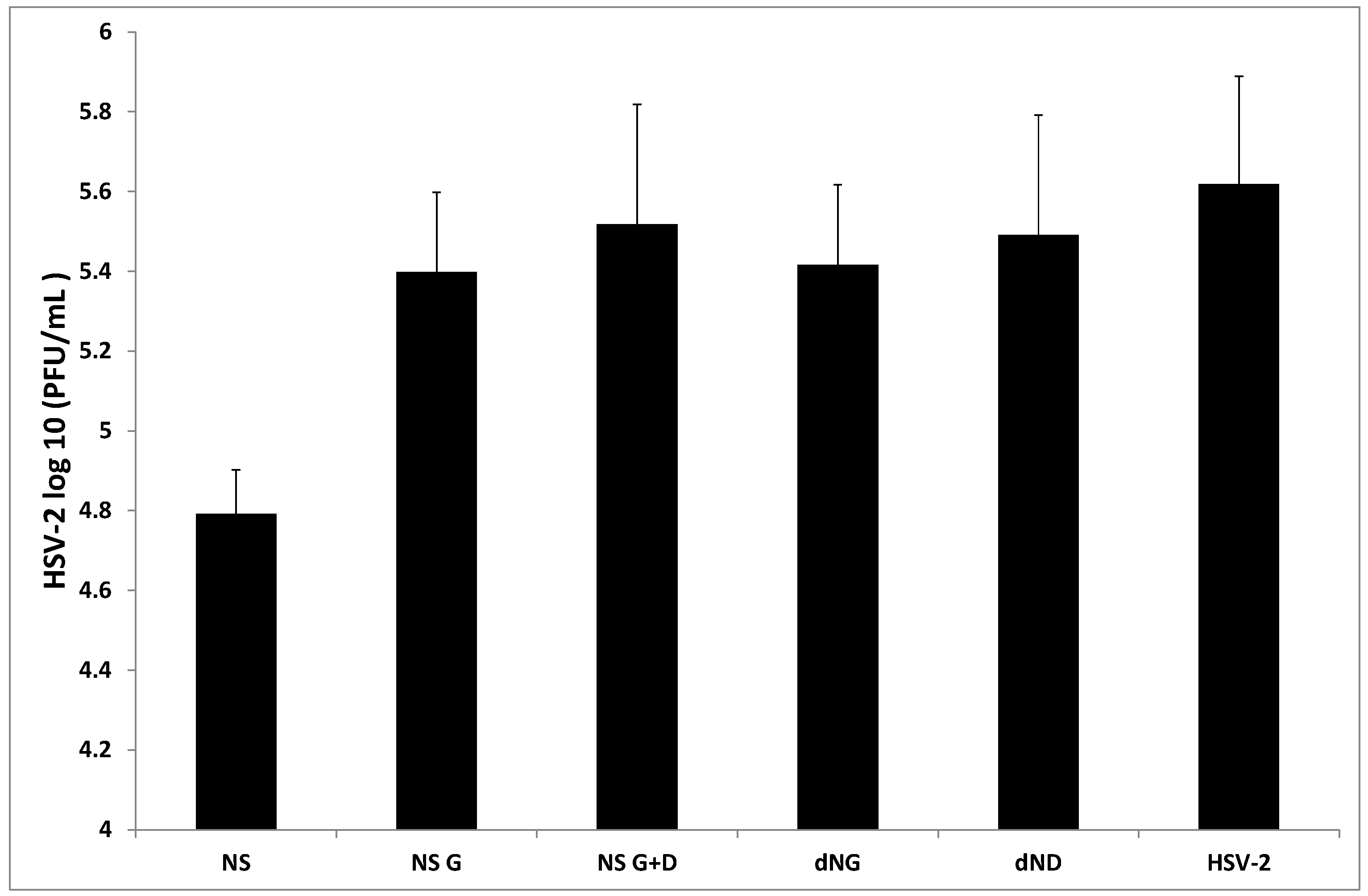
2. Results and Discussion

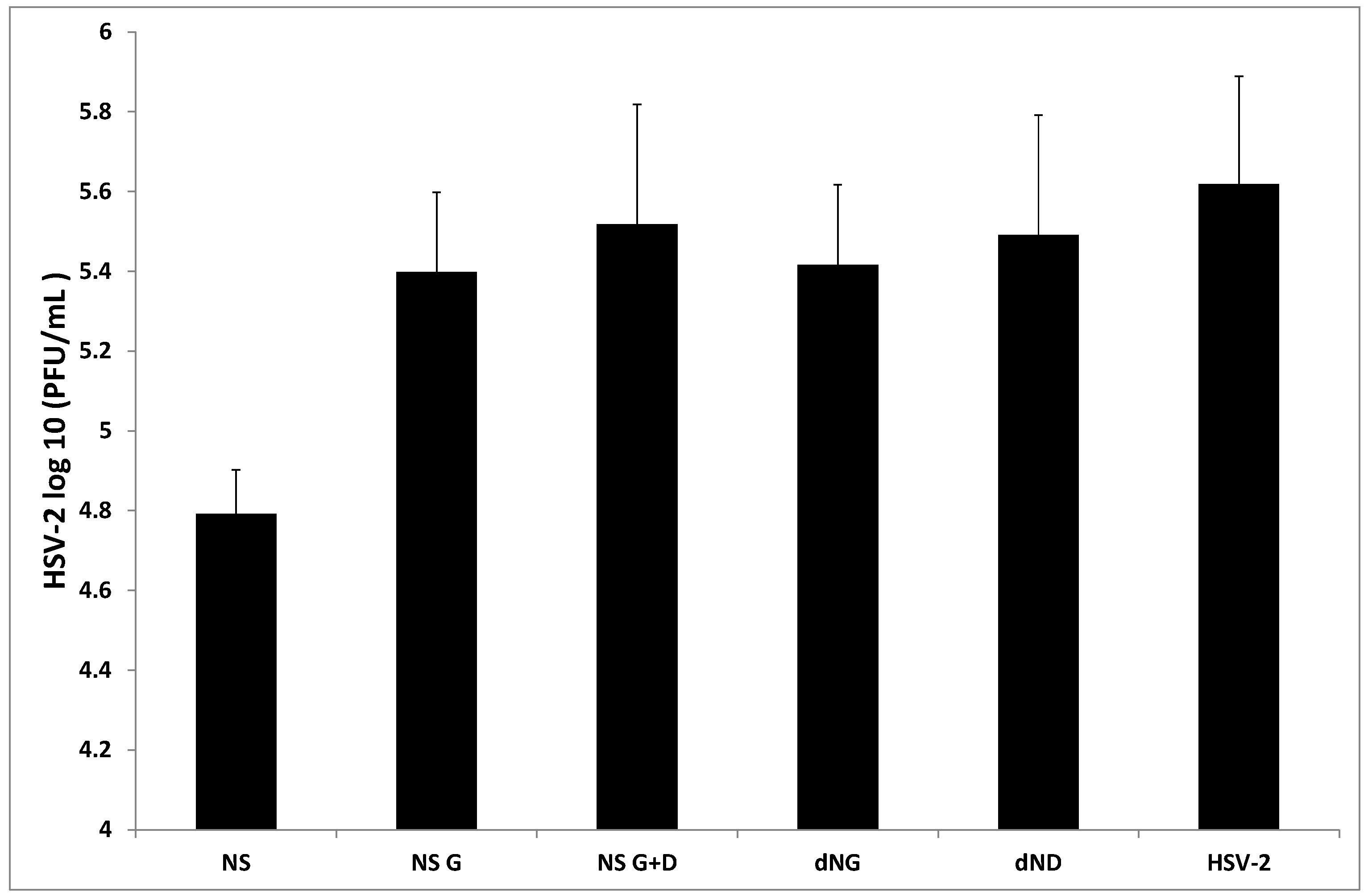{kind=link}
| Inducer | 500 μg/mL | 300 μg/mL | 100 μg/mL | 60 μg/mL |
| NS | 89 ± 9.1 | 40 ± 4.1 | 8 ± 0.6 | 0 |
| NS G | 91 ± 6.2 | 50 ± 4.8 | 10 ± 0.8 | 0 |
| NS G+D | 88 ± 7.8 | 90 ± 3.1 | 18 ± 1.1 | 0 |
| 100 μL/mL | 50 μL/mL | 20 μL/mL | 10 μL/mL | |
| d NG | 85 ± 5.9 | 50 ± 3.9 | 20 ± 0.7 | 0 |
| d ND | 60 ± 4.8 | 25 ± 1.8 | 8 ± 0.3 | 0 |
| IL-17 | IFN-γ | IFN-α | IL-4 | |||||
|---|---|---|---|---|---|---|---|---|
| PBMC | +HSV-2 | PBMC | +HSV-2 | PBMC | +HSV-2 | PBMC | +HSV-2 | |
| NS | <15 | <15 a | 1961 ± 257 | 2123 ± 380 | 98 ± 7.2 | 53 ± 3.8 a,b | 325 ± 69 | 298 ± 43 |
| NS G | <15 | 41 ± 3.8 b | <8 | <8 | <3.1 | <3.1 | 69 ± 31 | 42 ± 12 |
| NS G+D | <15 | 45 ± 4.3 b | <8 | <8 | <3.1 | <3.1 | 98 ± 9 | 52 ± 7 b |
| dNG | <15 | 29 ± 3.5 a,b | 163 ± 91 | 102 ± 96 a | <3.1 | <3.1 | 54 ± 26 | 83 ± 21 |
| dND | <15 | 39 ± 4.9 b | 189 ± 83 | <8 b | <3.1 | <3.1 | 41 ± 34 | 39 ± 8.4 |
| HSV-2 | 48 ± 4.1 | <8 | <3.1 | 181 ± 43 | ||||
| None | <15 | <15 | <8 | <8 | <3.1 | <3.1 | <10 | <10 |
| A | PBMC | + mAbvs IL-4 | + mAbvsIFN-γ | + mAbvsIFN-α | + mAbvsIFN-α and IL-4 |
| HSV-2 | 48 ± 4.1 | 86 ± 14.1 | nd | nd | nd |
| NS | <15 | 41 ± 3.6 | <15 | 116 ± 18.3 | 207 ± 25.9 |
| NS + HSV-2 | <15 | 54 ± 5.1 | <15 | 167 ± 20.9 | 309 ± 41.3 |
| B | PBMC | + rh IL-4 | + rh IFN-γ | + rh IFN-α | + rh IFN-α and rh IL-4 |
| HSV-2 | 48 ± 4.1 | 38 ± 5.5 | 51 ± 7.7 | 19 ± 2.1 | <15 |
| NS | <15 | <15 | <15 | <15 | <15 |
| NS + HSV-2 | <15 | <15 | <15 | <15 | <15 |
3. Experimental Section
3.1. General Information
3.2. Isolation of Human Peripheral Blood Mononuclear Cells (PBMC)
4. Conclusions
Acknowledgments
Author Contributions
Abbreviations
| IL | interleukin |
| IFN | interferon |
Conflicts of Interest
References
- Arena, A.; Bisignano, C.; Stassi, G.; Mandalari, G.; Wickham, M.S.; Bisignano, G. Immunomodulatory and antiviral activity of almond skins. Immunol. Lett. 2010, 132, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Tomaino, A.; Arcoraci, T.; Martorana, M.; Lo Turco, V.; Cacciola, F.; Rich, G.T.; Bisignano, C.; Saija, A.; Dugo, P.; et al. Characterization of polyphenols, lipids and dietary fibre from skins of almonds (Amygdalus communis L.). J. Food Comp. Anal. 2010, 23, 166–174. [Google Scholar] [CrossRef]
- Mandalari, G.; Tomaino, A.; Rich, G.T.; Lo Curto, R.B.; Arcoraci, T.; Martorana, M.; Bisignano, C.; Saija, A.; Parker, M.L.; Waldron, K.W.; et al. Polyphenol and nutrient release from skin of almonds during siulated human digestion. Food Chem. 2010, 122, 1083–1088. [Google Scholar] [CrossRef]
- Hirohata, S.; Shibuva, H.; Tejima, S. Suppressive influences of IFN-alpha on IL-17 expression in human CD4+ T cells. Clin. Immunol. 2010, 134, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhao, X.P.; Fan, Y.C.; Zhang, J.J.; Zhao, J.; Wang, K. IL-17A but not IL-22 suppresses the replication of hepatitis B virus mediated by over-expression of MxA and OAS mRNA in the HepG2.2.15 cell line. Antiviral Res. 2013, 97, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Kang, H.S.; Kim, B.S. Th17 cells enhance viral persistence and inhibit T cell cytotoxicity in a model of chronic virus infection. J. Exp. Med. 2009, 206, 313–328. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1989, 65, 55–63. [Google Scholar] [CrossRef]
- Boulanger, M.J.; Chow, D.C.; Breynova, E.; Martick, M.; Stanford, G.; Nocholas, J.; Garcia, K.C. Molecular mechanisms for viral mimicry of a human cytokine: Activation of gp 130 by HHV-8 interleukin-6. J. Mol. Biol. 2004, 335, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, T.H.; Melchjorsen, J.; Malmgaard, L.; Casola, A.; Paludan, S.R. Suppression of proinflammatory cytokine expression by herpes simplex virus type 1. J. Virol. 2004, 78, 5883–5890. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.B.; Sørensen, L.N.; Malmgaard, L.; Ank, N.; Baines, J.D.; Chen, Z.J.; Paludan, S.R. Type I interferon production during herpes simplex virus infection is controlled by cell-type-specific viral recognition through Toll-like receptor 9, the mitochondrial antiviral signaling protein pathway, and novel recognition systems. J. Virol. 2007, 81, 13315–13324. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: NS samples are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arena, A.; Bisignano, C.; Stassi, G.; Filocamo, A.; Mandalari, G. Almond Skin Inhibits HSV-2 Replication in Peripheral Blood Mononuclear Cells by Modulating the Cytokine Network. Molecules 2015, 20, 8816-8822. https://doi.org/10.3390/molecules20058816
Arena A, Bisignano C, Stassi G, Filocamo A, Mandalari G. Almond Skin Inhibits HSV-2 Replication in Peripheral Blood Mononuclear Cells by Modulating the Cytokine Network. Molecules. 2015; 20(5):8816-8822. https://doi.org/10.3390/molecules20058816
Chicago/Turabian StyleArena, Adriana, Carlo Bisignano, Giovanna Stassi, Angela Filocamo, and Giuseppina Mandalari. 2015. "Almond Skin Inhibits HSV-2 Replication in Peripheral Blood Mononuclear Cells by Modulating the Cytokine Network" Molecules 20, no. 5: 8816-8822. https://doi.org/10.3390/molecules20058816
APA StyleArena, A., Bisignano, C., Stassi, G., Filocamo, A., & Mandalari, G. (2015). Almond Skin Inhibits HSV-2 Replication in Peripheral Blood Mononuclear Cells by Modulating the Cytokine Network. Molecules, 20(5), 8816-8822. https://doi.org/10.3390/molecules20058816