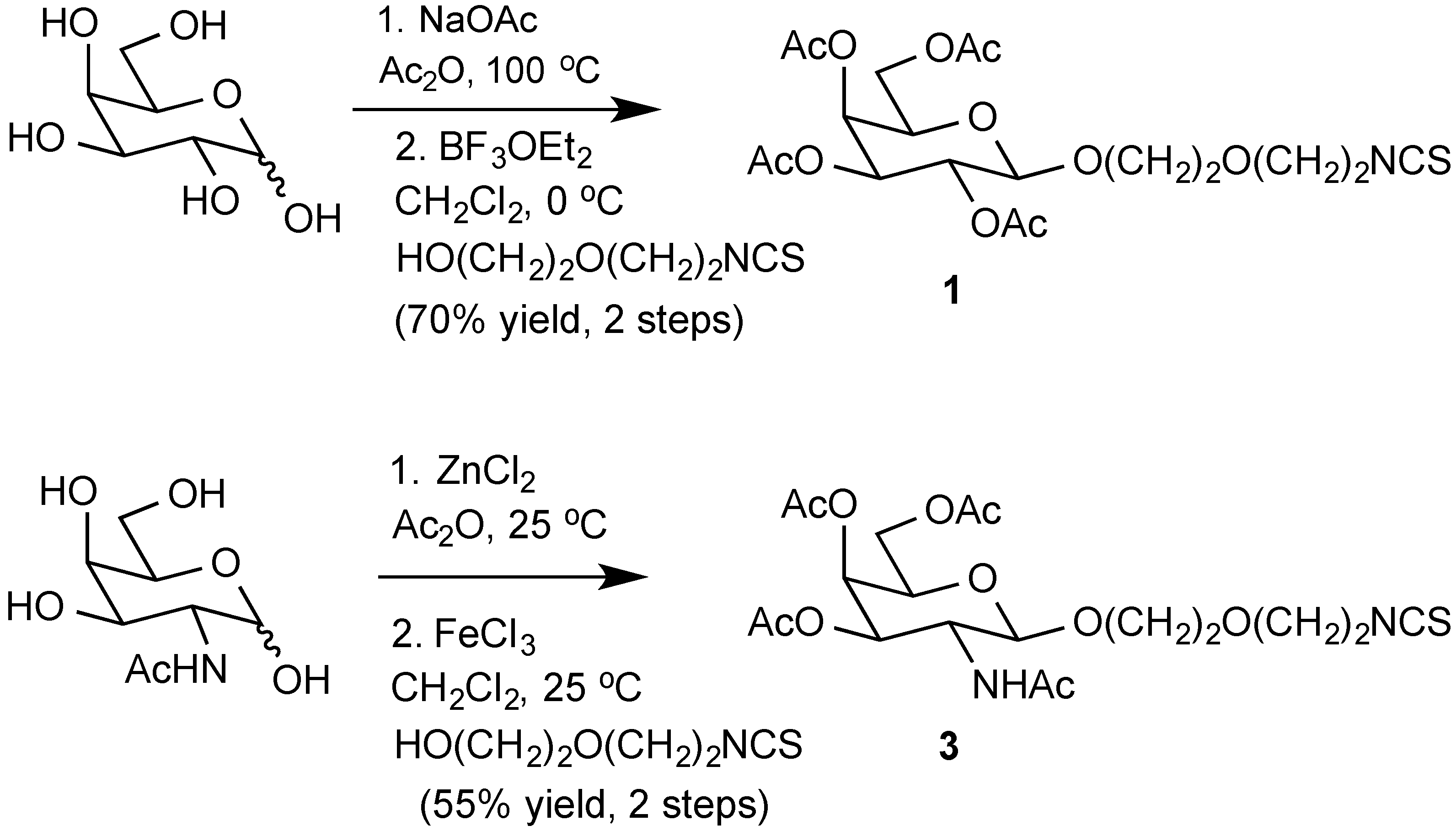
Acetylated
7a: 1H-NMR (500 MHz d6-DMSO) δ 7.95 (1H, bs, amide NH's), 7.74–7.79 (1.4H, m, amide NH's, NH'Ac), 7.46 (2H, bs, CH2NHC(S)NHCH2), 5.21 (1.2H, d, J = 3.2 Hz, H4), 5.17 (0.2H, s, H4'), 5.11 (1.2H, dd, J = 3.2, 10.3 Hz, H3), 4.92 (0.2H, d, J = 10.3 Hz, H3'), 4.89 (1.2H, t, J = 9.7 Hz, H2), 4.69 (1.2H, d, J = 7.6 Hz, H1), 4.51 (0.2H, d, J = 8.4 Hz, H1'), 4.12 (1.2H, m), 4.01 (3.1H, m), 3.83 (0.2H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.76 (2.0H, m), 3.27–3.59 (14.2H, m), 3.13 (3.1H, bs), 3.04 (2.4H, bs), 2.61 (3.1H, m), 2.16 (4.7H, bs), 2.08 (4H, s), 2.00 (2.8H, s), 1.96 (4.6H, s), 1.87 (3H, s) 1.72 (0.8H, s) ppm. MALDI-TOF (pos) m/z 19,100.
7b: 1H-NMR (500 MHz d6-DMSO) δ 7.95 (1H, bs, amide NH's), 7.79 (0.7H, d, J = 9.0 Hz, NH'Ac), 7.74 (0.9H, bs, amide NH's), 7.46 (2.1H, bs, CH2NHC(S)NHCH2), 5.21 (1.1H, d, J = 3.2 Hz, H4), 5.17 (0.4H, s, H4'), 5.11 (1.1H, dd, J = 3.2, 10.3 Hz, H3), 4.92 (0.4H, d, J = 10.3 Hz, H3'), 4.89 (1.1H, t, J = 9.7 Hz, H2), 4.69 (1.1H, d, J = 7.6 Hz, H1), 4.51 (0.4H, d, J = 8.4 Hz, H1'), 4.12 (1.1H, m), 4.01 (3.7H, m), 3.83 (0.4H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.76 (2.3H, m), 3.27–3.59 (15.9H, m), 3.13 (3.1H, bs), 3.04 (2.6H, bs), 2.61 (5.4H, m), 2.39 (3H, m), 2.16 (4.9H, bs), 2.08 (3H, s), 2.06 (1.5H, s), 1.96 (4.9H, s), 1.87 (2.9H, s), 1.85 (1.4H, s), 1.74 (1.2H, s) ppm. MALDI-TOF (pos) m/z 19,000.
7c: 1H-NMR (500 MHz d6-DMSO) δ 7.95 (1H, bs, amide NH's), 7.79 (0.8H, d, J = 9.0 Hz, NH'Ac), 7.74 (1H, bs, amide NH's), 7.46 (2H, bs, CH2NHC(S)NHCH2), 5.21 (0.8H, d, J = 3.2 Hz, H4), 5.17 (0.7H, s, H4'), 5.11 (0.8H, dd, J = 3.2, 10.3 Hz, H3), 4.92 (0.7H, d, J = 10.3 Hz, H3'), 4.89 (1.3H, t, J = 9.7 Hz, H2), 4.69 (1.3H, d, J = 7.6 Hz, H1), 4.51 (0.7H, d, J = 8.4 Hz, H1'), 4.12 (0.8H, m), 4.01 (3.8H, m), 3.83 (0.7H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.76 (1.9H, m), 3.27–3.59 (15.6H, m), 3.13 (3.2H, bs), 3.04 (2.6H, bs), 2.61 (5.6H, m), 2.39 (3H, m), 2.16 (4.7H, bs), 2.08 (4H, s), 2.00 (4H, s), 1.96 (4H, s), 1.87 (4H, s) ppm. MALDI-TOF (pos) m/z 18,900.
7d: 1H-NMR (500 MHz d6-DMSO) δ 7.95 (1H, bs, amide NH's), 7.79 (1.1H, d, J = 9.0 Hz, NH'Ac), 7.74 (0.9H, bs, amide NH's), 7.46 (2.1H, bs, CH2NHC(S)NHCH2), 5.21 (0.6H, d, J = 3.2 Hz, H4), 5.17 (0.9H, s, H4'), 5.11 (0.6H, dd, J = 3.2, 10.3 Hz, H3), 4.92 (0.9H, d, J = 10.3 Hz, H3'), 4.89 (0.6H, t, J = 9.7 Hz, H2), 4.69 (0.6H, d, J = 7.6 Hz, H1), 4.51 (0.9H, d, J = 8.4 Hz, H1'), 4.12 (0.6H, m), 4.01 (4.2H, m), 3.83 (0.9H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.76 (2.1H, m), 3.27–3.59 (15.7H, m), 3.13 (3.1H, bs), 3.04 (2.4H, bs), 2.61 (5.2H, m), 2.39 (3.1H, m), 2.16 (4.8H, bs), 2.07 (1.9H, s), 2.06 (2.8H, s), 1.98 (0.7H, s), 1.96 (4.9H, s), 1.87 (1.9H, s), 1.85 (2.5H, s), 1.74 (2.5H, s) ppm. MALDI-TOF (pos) m/z 19,300.
7e: 1H-NMR (500 MHz d6-DMSO) δ 7.95 (1H, bs, amide NH's), 7.79 (1.3H, d, J = 9.0 Hz, NH'Ac), 7.74 (0.8H, bs, amide NH's), 7.46 (2.1H, bs, CH2NHC(S)NHCH2), 5.21 (0.3H, d, J = 3.2 Hz, H4), 5.17 (1.1H, s, H4'), 5.11 (0.3H, dd, J = 3.2, 10.3 Hz, H3), 4.92 (1.1H, d, J = 10.3 Hz, H3'), 4.89 (0.3H, t, J = 9.7 Hz, H2), 4.69 (0.3H, d, J = 7.6 Hz, H1), 4.51 (1.1H, d, J = 8.4 Hz, H1'), 4.12 (0.8H, m), 4.01 (3.8H, m), 3.83 (1.1H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.76 (0.3H, m), 3.27–3.59 (15.3H, m), 3.13 (2.8H, bs), 3.04 (2.2H, bs), 2.61 (4.7H, m), 2.39 (2.7H, m), 2.16 (4.3H, bs), 2.07 (1.4H, s), 2.06 (3.4H, s), 1.95 (5H, s), 1.87 (1H, s), 1.85 (3.5H, s), 1.74 (3.3H, s) ppm. MALDI-TOF (pos) m/z 19,000.
7f: 1H-NMR (500 MHz d6-DMSO) δ 7.96 (1H, bs, amide NH's), 7.79 (1.3H, d, J = 9.0 Hz, NH'Ac), 7.74 (1.1H, bs, amide NH's), 7.46 (2.2H, m, CH2NHC(S)NHCH2), 5.17 (1.3H, s, H4'), 4.92 (1.3H, dd, J = 3.0, 10.3 Hz, H3'), 4.51 (1.3H, d, J = 8.4 Hz, H1'), 4.01 (4.7H, m), 3.83 (1.3H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.27–3.59 (16.5H, m), 3.13 (3.2H, bs), 3.04 (2.3H, bs), 2.61 (5.1H, m), 2.39 (2.5H, m), 2.16 (4.1H, bs), 2.06 (4.7H, s), 1.95 (4.7H, s), 1.85 (4.2H, s), 1.74 (4.2H, s) ppm. MALDI-TOF (pos) m/z 19,900.
8a: 1H-NMR (500 MHz d6-DMSO) δ 7.95 (1H, bs, amide NH's), 7.74–7.79 (1.4H, m, amide NH's, NH'Ac), 7.45 (2.1H, bs, CH2NHC(S)NHCH2), 5.21 (1.3H, s, H4), 5.17 (0.2H, s, H4'), 5.12 (1.3H, d, 10.5 Hz, H3), 4.92 (0.2H, m, H3'), 4.89 (1.3H, t, J = 9.7 Hz, H2), 4.69 (1.2H, d, J = 7.6 Hz, H1), 4.51 (0.2H, d, J = 8.4 Hz, H1'), 4.12 (1.3H, m), 4.00 (3.3H, m), 3.83 (0.2H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.76 (2.2H, m), 3.27–3.59 (12.7H, m), 3.13 (2.8H, bs), 3.04 (2.4H, bs), 2.61 (5.1H, m), 2.16 (4.8H, bs), 2.08 (4.7H, s), 2.00 (2.5H, s), 1.96 (5.3H, s), 1.87 (3.3H, s) 1.72 (0.8H, s) ppm. MALDI-TOF (pos) m/z 40,000.
8b: 1H-NMR (500 MHz d6-DMSO) δ 7.95 (1H, bs, amide NH's), 7.79 (0.7H, d, J = 9.0 Hz, NH'Ac), 7.74 (1.0H, bs, amide NH's), 7.46 (2.1H, m, CH2NHC(S)NHCH2), 5.21 (1.2H, s, H4), 5.17 (0.5H, s, H4'), 5.11 (1.3H, m, H3), 4.92 (0.5H, d, J = 10.3 Hz, H3'), 4.89 (1.3H, t, J = 9.7 Hz, H2), 4.69 (1.1H, d, J = 7.6 Hz, H1), 4.51 (0.4H, d, J = 8.4 Hz, H1'), 4.12 (1.3H, m), 4.01 (4.1H, m), 3.83 (0.6H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.76 (2.9H, m), 3.27–3.59 (13.8H, m), 3.13 (3.0H, bs), 3.04 (2.6H, bs), 2.61 (4.4H, m), 2.39 (2.7H, m), 2.16 (4.4H, bs), 2.08 (3.6H, s), 2.06 (1.6H, s), 1.96 (5.9H, s), 1.87 (3.2H, s), 1.85 (1.5H, s), 1.74 (1.3H, s) ppm. MALDI-TOF (pos) m/z 39,700.
8c: 1H-NMR (500 MHz d6-DMSO) δ 7.95 (1H, bs, amide NH's), 7.79 (0.9H, d, J = 9.0 Hz, NH'Ac), 7.74 (0.9H, bs, amide NH's), 7.46 (2.1H, bs, CH2NHC(S)NHCH2), 5.21 (0.7H, d, J = 3.2 Hz, H4), 5.17 (0.7H, s, H4'), 5.11 (0.7H, d, J = 10.3 Hz, H3), 4.92 (0.7H, d, J = 10.3 Hz, H3'), 4.89 (0.7H, t, J = 9.7 Hz, H2), 4.69 (0.6H, d, J = 7.6 Hz, H1), 4.51 (0.7H, d, J = 8.4 Hz, H1'), 4.12 (0.8H, m), 4.01 (3.9H, m), 3.83 (0.7H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.76 (1.9H, m), 3.27–3.59 (13.6H, m), 3.13 (2.9H, bs), 3.04 (2.4H, bs), 2.61 (5.3H, m), 2.39 (2.6H, m), 2.16 (4.6H, bs), 2.08 (5H, s), 2.00 (4.8H, s), 1.96 (1.9H, s), 1.87 (2.4H, s) 1.74 (2.1H, s) ppm. MALDI-TOF (pos) m/z 40,200.
8d: 1H-NMR (500 MHz d6-DMSO) δ 7.95 (1H, bs, amide NH's), 7.80 (1.0H, d, J = 9.0 Hz, NH'Ac), 7.79 (0.8H, bs, amide NH's), 7.46 (2.0H, m, CH2NHC(S)NHCH2), 5.21 (0.5H, s, H4), 5.17 (0.9H, s, H4'), 5.11 (0.4H, d, 10.3 Hz, H3), 4.92 (1H, d, J = 10.3 Hz, H3'), 4.89 (0.4H, t, J = 9.7 Hz, H2), 4.69 (0.4H, d, J = 7.6 Hz, H1), 4.51 (0.9H, d, J = 8.4 Hz, H1'), 4.12 (0.5H, m), 4.01 (4.1H, m), 3.83 (1.1H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.76 (2.0H, m), 3.27–3.59 (13.1H, m), 3.13 (2.7H, bs), 3.04 (2.0H, bs), 2.61 (4.2H, m), 2.39 (2.6H, m), 2.16 (3.8H, bs), 2.07 (1.7H, s), 2.06 (3.9H, s), 1.98 (1H, s), 1.96 (5.3H, s), 1.87 (1.4H, s), 1.85 (2.9H, s), 1.74 (2.8H, s) ppm. MALDI-TOF (pos) m/z 39,700.
8e: 1H-NMR (500 MHz d6-DMSO) δ 7.95 (1H, bs, amide NH's), 7.79 (1.3H, d, J = 9.0 Hz, NH'Ac), 7.74 (0.8H, bs, amide NH's), 7.46 (2.1H, m, CH2NHC(S)NHCH2), 5.21 (0.3H, d, J = 3.2 Hz, H4), 5.17 (1.2H, s, H4'), 5.11 (0.3H, d, J = 10.3 Hz, H3), 4.92 (1.2H, d, J = 10.3 Hz, H3'), 4.89 (0.3H, t, J = 9.7 Hz, H2), 4.69 (0.2H, d, J = 7.6 Hz, H1), 4.51 (1.2H, d, J = 8.4 Hz, H1'), 4.12 (0.3H, m), 4.01 (3.6H, m), 3.83 (1.3H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.76 (2.2H, m), 3.27–3.59 (8.3H, m), 3.13 (3.0H, bs), 3.04 (2.1H, bs), 2.61 (5.0H, m), 2.39 (2.7H, m), 2.16 (4.4H, bs), 2.07 (1.1H, s), 2.06 (3.4H, s), 1.95 (5.3H, s), 1.87 (1H, s), 1.85 (3.9H, s), 1.74 (3.7H, s) ppm. MALDI-TOF (pos) m/z 40,500.
8f: 1H-NMR (500 MHz d6-DMSO) δ 7.96 (1H, bs, amide NH's), 7.81 (1.2H, d, J = 9.0 Hz, NH'Ac), 7.74 (0.8H, bs, amide NH's), 7.46 (2.0H, m, CH2NHC(S)NHCH2), 5.17 (1.3H, s, H4'), 4.92 (1.3H, d, J = 10.3 Hz, H3'), 4.51 (1.4H, d, J = 8.4 Hz, H1'), 4.01 (4.9H, m), 3.83 (1.6H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.27–3.59 (17H, m), 3.13 (2.8H, bs), 3.04 (2.1H, bs), 2.61 (4H, m), 2.39 (2H, m), 2.16 (3.9H, bs), 2.06 (4.7H, s), 1.95 (5H, s), 1.85 (4.5H, s), 1.74 (4.4H, s) ppm. MALDI-TOF (pos) m/z 39,300.
9a: 1H-NMR (500 MHz d6-DMSO) δ 7.95 (1H, bs, amide NH's), 7.74–7.79 (1.3H, m, amide NH's, NH'Ac), 7.45 (1.9H, bs, CH2NHC(S)NHCH2), 5.21 (1H, s, H4), 5.17 (0.2H, s, H4'), 5.11 (1H, d, 10.3 Hz, H3), 4.89 (1H, t, J = 9.7 Hz, H2), 4.69 (1H, d, J = 7.6 Hz, H1), 4.51 (0.2H, d, J = 8.4 Hz, H1'), 4.12 (1H, m), 4.01 (2.6H, m), 3.83 (0.2H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.76 (1.5H, m), 3.27–3.59 (11.4H, m), 3.13 (2.9H, bs), 3.04 (2.3H, bs), 2.61 (4.8H, m), 2.16 (4.1H, bs), 2.08 (4.2H, s), 1.98 (7.6H, s), 1.87 (3.7H, s) 1.72 (0.8H, s) ppm. MALDI-TOF (pos) m/z 119,500.
9b: 1H-NMR (500 MHz d6-DMSO) δ 7.96 (1H, bs, amide NH's), 7.79 (0.6H, d, J = 9.0 Hz, NH'Ac), 7.74 (0.9H, bs, amide NH's), 7.46 (1.9H, bs, CH2NHC(S)NHCH2), 5.21 (2.1H, d, J = 3.3 Hz, H4), 5.17 (0.4H, s, H4'), 5.11 (2.1H, dd, J = 3.2, 10.3 Hz, H3), 4.92 (0.3H, d, J = 10.3 Hz, H3'), 4.89 (2.1H, t, J = 9.7 Hz, H2), 4.69 (1.9H, d, J = 7.6 Hz, H1), 4.51 (0.2H, d, J = 8.4 Hz, H1'), 4.12 (2.1H, m), 4.01 (5.4H, m), 3.83 (0.3H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.76 (5.4H, m), 3.27–3.59 (17.5H, m), 3.13 (3.2H, bs), 3.04 (2.3H, bs), 2.61 (4.8H, m), 2.39 (2.8H, m), 2.16 (4.4H, bs), 2.08 (7.8H, s), 2.06 (4.6H, s), 1.96 (5.5H, s), 1.85 (1.6H, s), 1.74 (1.4H, s) ppm. MALDI-TOF (pos) m/z 119,000.
9c: 1H-NMR (500 MHz d6-DMSO) δ 7.95 (1H, bs, amide NH's), 7.81 (0.7H, d, J = 9.0 (H2') Hz, NH'Ac), 7.74 (0.9H, bs, amide NH's), 7.46 (2H, bs, CH2NHC(S)NHCH2), 5.21 (0.7H, d, J = 3.2 Hz, H4), 5.17 (0.6H, s, H4'), 5.11 (0.7H, d, J = 10.3 Hz, H3), 4.92 (0.6H, d, J = 10.3 Hz, H3'), 4.89 (0.7H, t, J = 9.7 Hz, H2), 4.69 (0.6H, d, J = 7.6 Hz, H1), 4.51 (0.5H, d, J = 8.4 Hz, H1'), 4.12 (0.8H, m), 4.01 (3.2H, m), 3.83 (0.7H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.76 (1.7H, m), 3.27–3.59 (12H, m), 3.13 (3H, bs), 3.04 (2.1H, bs), 2.61 (5.4H, m), 2.39 (2H, m), 2.16 (4H, bs), 2.08 (3.7H, s), 2.00 (5.5H, s), 1.96 (1.7H, s), 1.87 (1.6H, s), 1.74 (1.4H, s) ppm. MALDI-TOF (pos) m/z 119,000.
9d: 1H-NMR (500 MHz d6-DMSO) δ 7.96 (1H, bs, amide NH's), 7.79 (0.8H, d, J = 9.0 Hz, NH'Ac), 7.74 (0.7H, bs, amide NH's), 7.46 (1.8H, m, CH2NHC(S)NHCH2), 5.21 (0.5H, s, H4), 5.17 (0.6H, s, H4'), 5.11 (0.5H, d, J = 10.3 Hz, H3), 4.92 (0.6H, d, J = 10.3 Hz, H3'), 4.89 (0.5H, t, J = 9.7 Hz, H2), 4.69 (0.4H, d, J = 7.6 Hz, H1), 4.51 (0.6H, d, J = 8.4 Hz, H1'), 4.12 (0.6H, m), 4.01 (3.1H, m), 3.83 (0.8H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.76 (1.5H, m), 3.27–3.59 (10H, m), 3.13 (2.8H, bs), 3.04 (1.7H, bs), 2.61 (4H, m), 2.39 (2.2H, m), 2.16 (3.5H, bs), 2.07 (3.5H, s), 1.98 (0.7H, s), 1.96 (4.4H, s), 1.87 (1.2H, s), 1.85 (2H, s), 1.74 (1.9H, s) ppm. MALDI-TOF (pos) m/z 121,500.
9e: 1H-NMR (500 MHz d6-DMSO) δ 7.95 (1H, bs, amide NH's), 7.79 (0.8H, d, J = 9.0 Hz, NH'Ac), 7.74 (0.7H, bs, amide NH's), 7.46 (1.6H, bs, CH2NHC(S)NHCH2), 5.21 (0.2H, d, J = 3.2 Hz, H4), 5.17 (0.7H, s, H4'), 5.11 (0.3H, m, Hz, H3), 4.92 (0.7H, d, J = 10.3 Hz, H3'), 4.89 (0.2H, t, J = 9.7 Hz, H2), 4.69 (0.2H, d, J = 7.6 Hz, H1), 4.51 (0.7H, d, J = 8.4 Hz, H1'), 4.12 (0.3H, m), 4.01 (2.9H, m), 3.83 (0.9H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.7 (1.2H, m), 3.27–3.59 (8.6H, m), 3.04–3.13 (2.8H, m), 2.61 (3.2H, m), 2.39 (1.4H, m), 2.16 (2.9H, bs), 2.07 (3.2H, s), 1.95 (3.9H, s), 1.85 (2.9H, s), 1.74 (2.2H, s) ppm. MALDI-TOF (pos) m/z 121,500.
9f: 1H-NMR (500 MHz d6-DMSO) δ 7.96 (1H, bs, amide NH's), 7.79 (1.1H, d, J = 9.0 Hz, NH'Ac), 7.74 (0.8H, bs, amide NH's), 7.46 (2.8H, m, CH2NHC(S)NHCH2), 5.17 (1.1H, s, H4'), 4.92 (1.1H, d, J = 10.3 Hz, H3'), 4.51 (1.2H, d, J = 8.4 Hz, H1'), 4.01 (4.2H, m), 3.83 (1.2H, app q, J = 8.4, 9.0, 10.3 Hz, H2'), 3.27–3.59 (12.7H, m), 3.04–3.13 (4.2H, m), 2.61 (3.9H, m), 2.16 (3.1H, bs), 2.06 (4.4H, s), 1.95 (4.1H, s), 1.85 (3.7H, s), 1.74 (3.8H, s) ppm. MALDI-TOF (pos) m/z 125,000.
Deacetylated
7a: 1H-NMR (500 MHz d6-DMSO) δ 8.00 (bs, 1H), 7.81 (bs, 1H), 7.67 (d, J = 8.6 Hz, 0.2H), 7.50 (bs, 2.1H), 4.95 (bs, 0.7H), 4.60–4.70 (m, 1.5H), 4.43 (bs, 0.8H), 4.27 (d, J = 8.3 Hz, 0.2H), 4.11 (d, J = 5.2 Hz, 0.8H), 3.84 (m, 1.0H), 3.38–3.65 (m, 27H), 3.17 (bs, 2.8H), 3.08 (bs, 2.6H), 2.66 (bs, 4.0H), 2.42 (bs, 2.3H), 2.20 (bs, 4.3H), 1.80 (m, 0.9H) ppm. MALDI-TOF (pos) m/z 15,000.
7b: 1H-NMR (500 MHz d6-DMSO) δ 8.00 (bs, 1H), 7.80 (bs, 0.9H), 7.67 (d, J = 8.6 Hz, 0.3H), 7.50 (m, 1.5H), 4.95 (bs, 0.6H), 4.50–4.75 (m, 2.1H), 4.43 (bs, 0.7H), 4.27 (d, J = 8.3 Hz, 0.4H), 4.11 (d, J = 5.2Hz, 0.7H), 3.84 (m, 0.9H), 3.38–3.65 (m, 24H), 3.17 (bs, 2.3H), 3.08 (bs, 1.9H), 2.66 (bs, 3.4H), 2.42 (bs, 1.8H), 2.20 (bs, 3.7H), 1.80 (m, 1.0H) ppm. MALDI-TOF (pos) m/z 15,100.
7c: 1H-NMR (500 MHz d6-DMSO) δ 8.01 (bs, 1H), 7.83 (bs, 0.9H), 7.67 (d, J = 8.6 Hz, 0.5H), 7.59 (bs, 0.5H), 7.50 (bs, 1.2H), 4.95 (bs, 0.5H), 4.60–4.70 (m, 2.2H), 4.43 (bs, 0.7H), 4.27 (d, J = 8.3 Hz, 0.7H), 4.11 (d, J = 5.2 Hz, 0.6H), 3.84 (m, 1.0H), 3.38–3.65 (m, 30H), 3.17 (bs, 3.1H), 3.08 (bs, 2.1H), 2.66 (bs, 4.0H), 2.42 (bs, 1.8H), 2.20 (bs, 3.9H), 1.80 (m, 1.6H) ppm. MALDI-TOF (pos) m/z 7,900.
7d: 1H-NMR (500 MHz d6-DMSO) δ 8.02 (bs, 1H), 7.83 (bs, 0.8H), 7.67 (d, J = 8.6 Hz, 0.7H), 7.59 (bs, 0.6H), 7.50 (m, 1.1H), 4.95 (bs, 0.4H), 4.60–4.70 (m, 2.4H), 4.43 (bs, 0.5H), 4.27 (d, J = 8.3 Hz, 0.9H), 4.11 (d, J = 5.2 Hz, 0.5H), 3.38–3.65 (m, 30H), 3.17 (bs, 2.9H), 3.08 (bs, 2.0H), 2.66 (bs, 3.9H), 2.42 (bs, 1.9H), 2.20 (bs, 3.9H), 1.80 (m, 2.0H) ppm. MALDI-TOF (pos) m/z 15,700.
7e: 1H-NMR (500 MHz d6-DMSO) δ 8.01 (bs, 1H), 7.82 (bs, 0.9H), 7.67 (d, J = 8.6 Hz, 0.9H), 7.60 (bs, 0.8H), 7.50 (bs, 0.7H), 4.95 (bs, 0.2H), 4.60–4.70 (m, 2.6H), 4.43 (bs, 0.2H), 4.27 (d, J = 8.3 Hz, 1.0H), 4.11 (d, J = 5.2 Hz, 0.3H), 3.38–3.65 (m, 23H), 3.17 (bs, 2.6H), 3.08 (bs, 2.61), 2.66 (bs, 3.9H), 2.42 (bs, 1.7H), 2.20 (bs, 3.7H), 1.80 (m, 2.6H) ppm. MALDI-TOF (pos) m/z 15,800.
7f: 1H-NMR (500 MHz d6-DMSO) δ 8.01 (bs, 1H), 7.82 (bs, 0.9H), 7.68 (d, J = 8.6 Hz, 1.0H), 7.59 (bs, 0.9H), 7.50 (m, 1.0H), 4.60–4.70 (m, 2.8H), 4.27 (d, J = 8.3 Hz, 1.1H), 3.77 (m, 3.3H), 3.38–3.65 (m, 30H), 3.17 (bs, 3.3H), 3.10 (bs, 1.9H), 2.66 (bs, 4.1H), 2.42 (bs, 1.9H), 2.20 (bs, 3.8H), 1.80 (m, 3.0H) ppm. MALDI-TOF (pos) m/z 16,400.
8a: 1H-NMR (500 MHz d6-DMSO) δ 8.01 (bs, 1H), 7.81 (bs, 1H), 7.67 (m, 0.2H), 7.50 (bs, 1.8H), 4.95 (bs, 0.6H), 4.60–4.70 (m, 2.0H), 4.43 (bs, 0.9H), 4.27 (m, 0.2H), 4.11 (m, 0.8H), 3.84 (m, 1.0H), 3.38–3.65 (m, 30H), 3.17 (bs, 2.5H), 3.08 (bs, 2.6H), 2.66 (bs, 3.9H), 2.42 (bs, 2.1H), 2.20 (bs, 4.1H), 1.80 (m, 0.7H) ppm. 31,200.
8b: 1H-NMR (500 MHz d6-DMSO) δ 8.03 (bs, 1H), 7.85 (bs, 0.9H), 7.73 (d, J = 8.6 Hz, 0.4H), 7.50 (m, 1.6H), 4.95 (bs, 0.4H), 4.50–4.90 (m, 2.3H), 4.52 (bs, 0.6H), 4.27 (d, J = 8.3 Hz, 0.5H), 4.12 (s, 0.9H), 3.38–3.70 (m, 30H), 3.17 (bs, 2.7H), 3.08 (bs, 2.1H), 2.66 (bs, 3.8H), 2.42 (bs, 1.8H), 2.20 (bs, 3.9H), 1.80 (m, 1.2H) ppm. MALDI-TOF (pos) m/z 31,700.
8c: 1H-NMR (500 MHz d6-DMSO) δ 8.02 (bs, 1H), 7.83 (bs, 0.9H), 7.71 (m, 0.5H), 7.58 (bs, 0.6H), 7.50 (bs, 1.0H), 4.95 (bs, 0.3H), 4.60–4.70 (m, 1.7H), 4.58 (bs, 0.5H), 4.48 (bs, 0.5H), 4.27 (d, J = 8.3 Hz, 0.5H), 4.11 (m, 0.4H), 3.84 (m, 1.6H), 3.38–3.65 (m, 30H), 3.17 (bs, 2.4H), 3.08 (bs, 2.1H), 2.66 (bs, 3.8H), 2.42 (bs, 1.7H), 2.20 (bs, 4.0H), 1.80 (m, 1.8H) ppm. MALDI-TOF (pos) m/z 33,000.
8d: 1H-NMR (500 MHz d6-DMSO) δ 8.02 (bs, 1H), 7.83 (bs, 0.8H), 7.67 (d, J = 8.6 Hz, 0.8H), 7.58 (bs, 0.7H), 7.43 (m, 0.9H), 4.95 (bs, 0.1H), 4.69 (bs, 1.7H),4.58 (bs, 0.8H), 4.43 (bs, 0.2H), 4.27 (d, J = 8.3 Hz, 0.8H), 4.11 (m, 0.1H), 3.38–3.65 (m, 30H), 3.17 (bs, 2.5H), 3.08 (bs, 1.8H), 2.66 (bs, 4.1H), 2.42 (bs, 1.8H), 2.21 (bs, 3.8H), 1.80 (m, 2.2H) ppm. MALDI-TOF (pos) m/z 34,300.
8e: 1H-NMR (500 MHz d6-DMSO) δ 8.02 (bs, 1H), 7.83 (bs, 0.8H), 7.67 (d, J = 8.6 Hz, 0.9H), 7.58 (bs, 0.8H), 7.44 (bs, 0.8H), 4.68 (m, 1.5H), 4.58 (1.0H), 4.27 (d, J = 8.3 Hz, 0.9H), 3.38–3.65 (m, 23H), 3.17 (bs, 2.4H), 3.08 (bs, 1.9H), 2.66 (bs, 3.8H), 2.42 (bs, 1.6H), 2.20 (bs, 3.8H), 1.80 (m, 2.8H) ppm. MALDI-TOF (pos) m/z 34,300.
8f: 1H-NMR (500 MHz d6-DMSO) δ 8.02 (bs, 1H), 7.83 (bs, 0.8H), 7.69 (d, J = 8.6 Hz, 0.8H), 7.58 (bs, 0.8H), 7.43 (m, 0.9H), 4.67 (m, 1.6H), 4.58 (bs, 0.8H), 4.27 (d, J = 8.3 Hz, 0.8H), 3.38–3.65 (m, 30H), 3.10–3.17 (m, 4.6H), 2.66 (bs, 3.7H), 2.42 (bs, 1.6H), 2.20 (bs, 3.6H), 1.80 (m, 2.4H) ppm. MALDI-TOF (pos) m/z 33,300.
9a: 1H-NMR (500 MHz d6-DMSO) δ 8.01 (bs, 1H), 7.83 (bs, 0.7H), 7.67 (m, 0.2H), 7.48 (bs, 1.5H), 5.04 (bs, 0.7H), 4.60–4.70 (m, 1.7H), 4.43 (bs, 0.8H), 4.28 (m, 0.2H), 4.11 (s, 1.0H), 3.84 (m, 2.0H), 3.38–3.65 (m, 30H), 3.17 (bs, 2.1H), 3.08 (bs, 1.7H), 2.65 (bs, 3.2H), 2.42 (bs, 1.4H), 2.20 (bs, 3.5H), 1.80 (m, 0.6H) ppm. MALDI-TOF (pos) m/z 100,000.
9b: 1H-NMR (500 MHz d6-DMSO) δ 8.03 (bs, 1H), 7.85 (bs, 0.7H), 7.73 (s, 0.3H), 7.47 (m, 1.4H), 5.07 (bs, 0.5H), 4.68–4.90 (m, 1.7H), 4.63 (bs, 0.4H), 4.54 (bs, 0.6H), 4.27 (s, 0.4H), 4.12 (s, 0.9H), 3.38–3.70 (m, 30H), 3.17 (bs, 2.2H), 3.08 (bs, 1.6H), 2.66 (bs, 2.8H), 2.20 (bs, 3.4H), 1.80 (m, 1.2H) ppm. MALDI-TOF (pos) m/z 101,000.
9c: 1H-NMR (500 MHz d6-DMSO) δ 8.02 (bs, 1H), 7.84 (bs, 0.7H), 7.72 (s, 0.5H), 7.58 (bs, 0.5H), 7.44 (bs, 0.9H), 5.04 (bs, 0.3H), 4.72 (m, 1.7H), 4.61 (bs, 0.8H), 4.28 (s, 0.6H), 4.13 (m, 0.4H), 3.38–3.80 (m, 30H), 3.10–3.17 (m, 3.5H), 2.66 (bs, 3.0H), 2.42 (bs, 1.0H), 2.20 (bs, 3.4H), 1.80 (m, 1.5H) ppm. MALDI-TOF (pos) m/z 101,500.
9d: 1H-NMR (500 MHz d6-DMSO) δ 8.02 (bs, 1H), 7.84 (bs, 0.8H), 7.72 (s, 0.4H), 7.57 (bs, 0.5H), 7.48 (m, 0.9H), 5.05 (bs, 0.4H), 4.69 (bs, 1.6H), 4.58 (bs, 0.5H), 4.43 (bs, 0.2H), 4.27 (s, 0.5H), 4.13 (m, 0.5H), 3.38–3.65 (m, 30H), 3.10–3.17 (bs, 3.6H), 2.66 (bs, 3.0H), 2.42 (bs, 1.3H), 2.21 (bs, 3.3H), 1.82 (m, 1.4H) ppm. MALDI-TOF (pos) m/z 102,000.
9e: 1H-NMR (500 MHz d6-DMSO) δ 8.02 (bs, 1H), 7.82 (bs, 0.7H), 7.67 (s, 0.7H), 7.58 (bs, 0.7H), 7.44 (bs, 0.8H), 5.05 (bs, 0.2H), 4.68 (m, 1.7H), 4.60 (0.7H), 4.28 (s, 0.7H), 3.38–3.65 (m, 23H), 3.10–3.17 (m, 3.6H), 2.66 (bs, 2.9H), 2.42 (bs, 0.8H), 2.21 (bs, 3.2H), 1.80 (m, 1.8H) ppm. MALDI-TOF (pos) m/z 106,500.
9f: 1H-NMR (500 MHz d6-DMSO) δ 8.03 (bs, 1H), 7.84 (bs, 0.7H), 7.72 (s, 0.8H), 7.58 (bs, 0.7H), 7.43 (m, 0.9H), 4.73 (m, 1.6H), 4.62 (bs, 0.9H), 4.27 (s, 0.8H), 3.38–3.65 (m, 30H), 3.10–3.17 (m, 3.8H), 2.66 (bs, 3.0H), 2.42 (bs, 1.2H), 2.21 (bs, 3.5H), 1.80 (m, 2.2H) ppm. MALDI-TOF (pos) m/z 107,500.



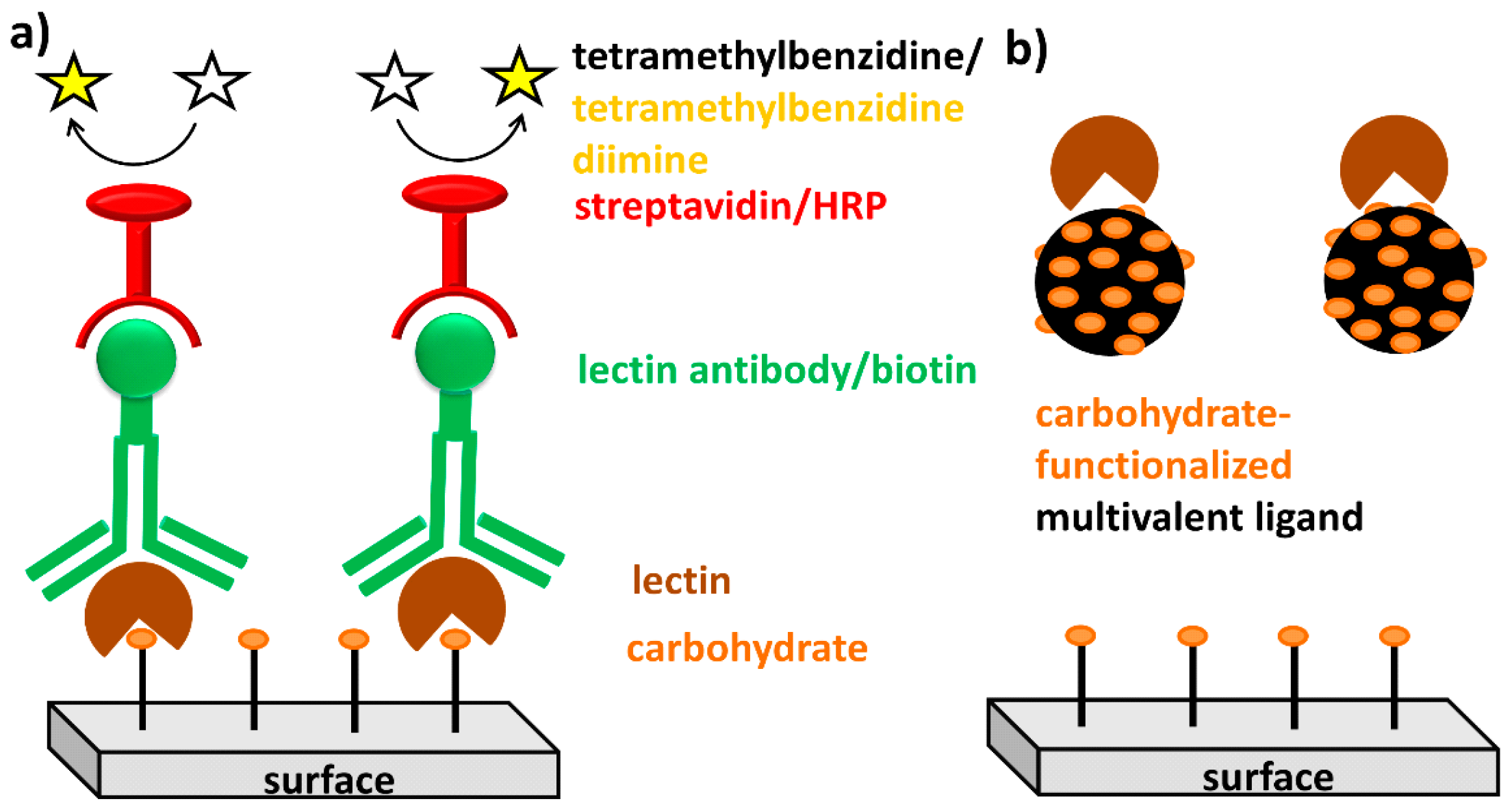
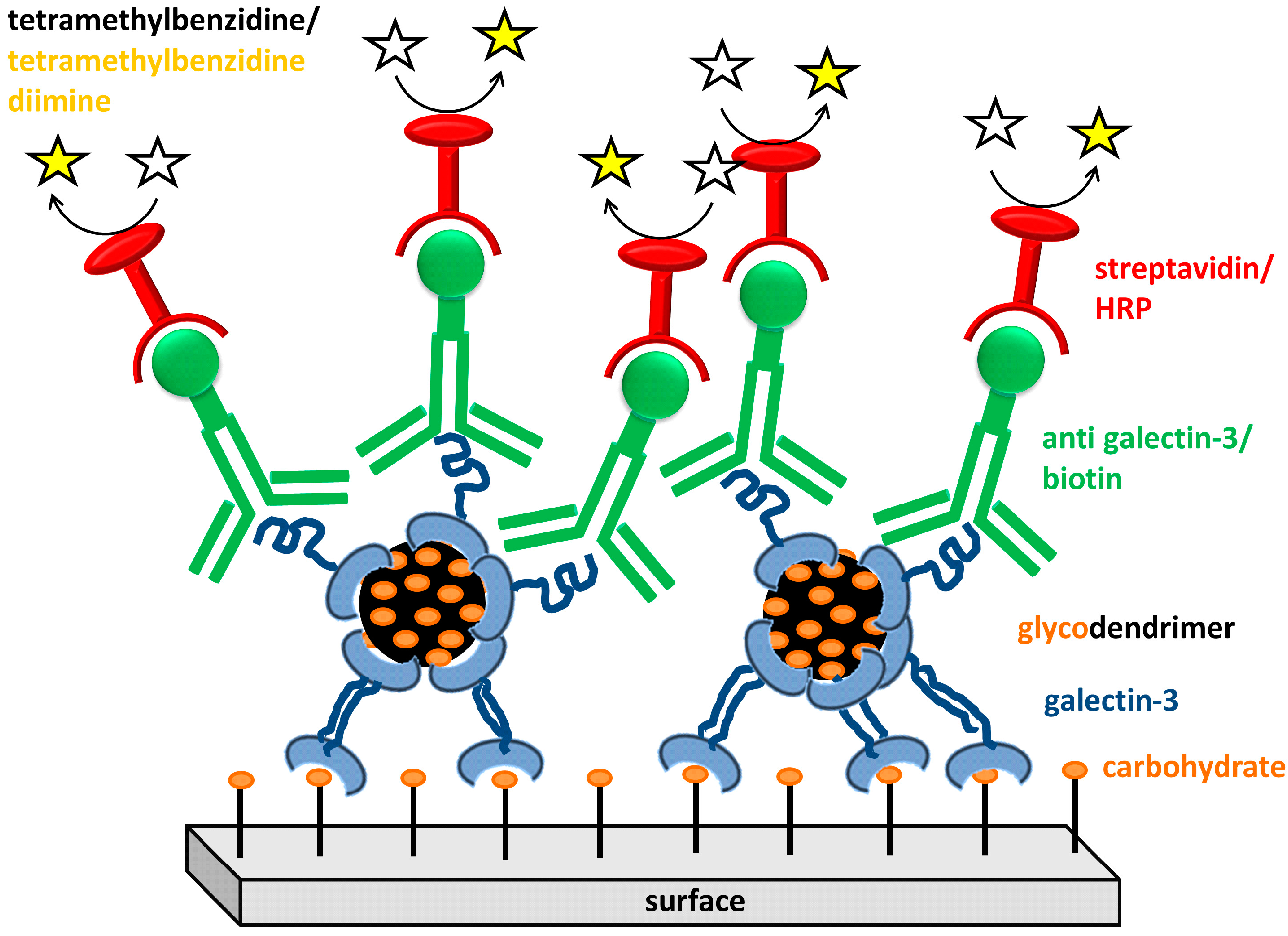
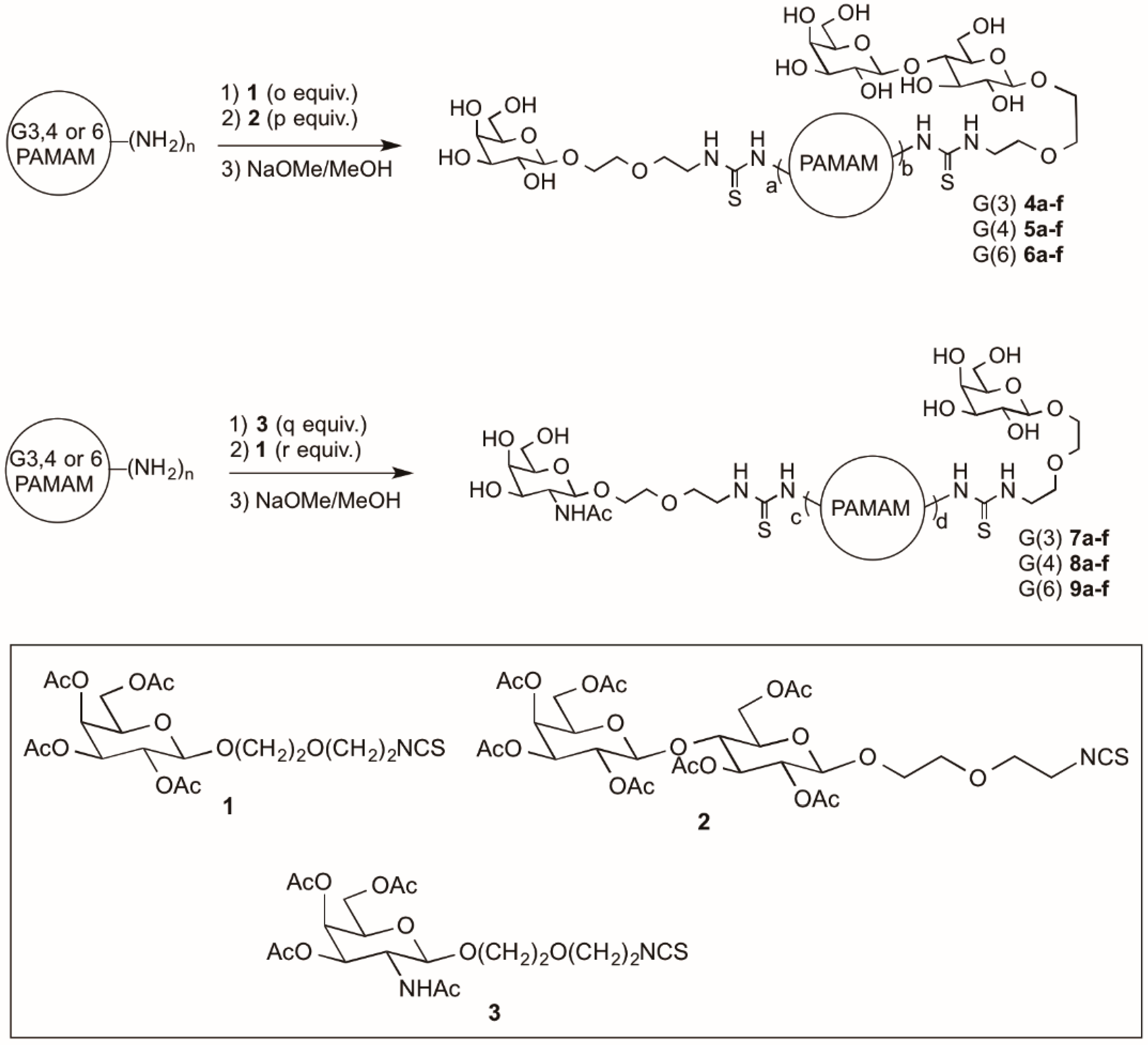

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}