
Breeding Vegetables with Increased Content in Bioactive Phenolic Acids

 ,
, 

 ,
, 
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetable | Total Phenolic Acids [mg/100 g fw] | Major Soluble Phenolic Acids |
|---|---|---|
| Eggplant (Solanum melongena) | 32.0 | chlorogenic |
| Carrot (Daucus carota) | 29.5 | chlorogenic, caffeic, protocatechuic |
| Red beet (Beta vulgaris) | 27.0 | ferulic |
| Basil (Ocimum basilicum) | 22.0 | chlorogenic |
| Broccoli (Brassica oleracea var. italica) | 15.0 | sinapic, caffeic |
| Radish (Raphanus sativus var. sativus) | 12.0 | p-coumaric, ferulic |
| Spinach (Spinacia oleracea) | 11.0 | chlorogenic, protocatechuic, gallic |
| Chinese cabbage (Brassica pekinensis) | 7.7 | sinapic, chlorogenic |
| Parsley (Petroselinum crispum) | 6.2 | protocatechuic |
| Parsnip (Pastinaca sativa) | 5.7 | chlorogenic |
| Lettuce (Lactuca sativa var. capitata) | 5.1 | chlorogenic |
| Pepper (Capsicum annuum) | 4.7 | chlorogenic, p-coumaric, ferulic, protocatechuic |
| Cauliflower (Brassica oleracea var. botrytis) | 4.6 | p-coumaric, sinapic, chlorogenic |
| Turnip (Brassica rapa) | 4.6 | sinapic, ferulic, chlorogenic |
| White cabbage (Brassica oleracea var. capitata f. alba) | 3.8 | sinapic, p-coumaric |
| Grean bean (Phaseolus vulgaris) | 3.5 | chlorogenic, protocatechuic |
| Tomato (Solanum esculentum) | 3.5 | chlorogenic |
| Pea (Pisum sativum) | 1.3 | sinapic |
| Onion (Allium cepa) | 1.0 | protocatechuic, p-coumaric |
| Zucchini (Cucurbita pepo) | 0.9 | p-coumaric, caffeic |
| Cucumber (Cucumis sativus) | 0.1 | p-coumaric, ferulic |
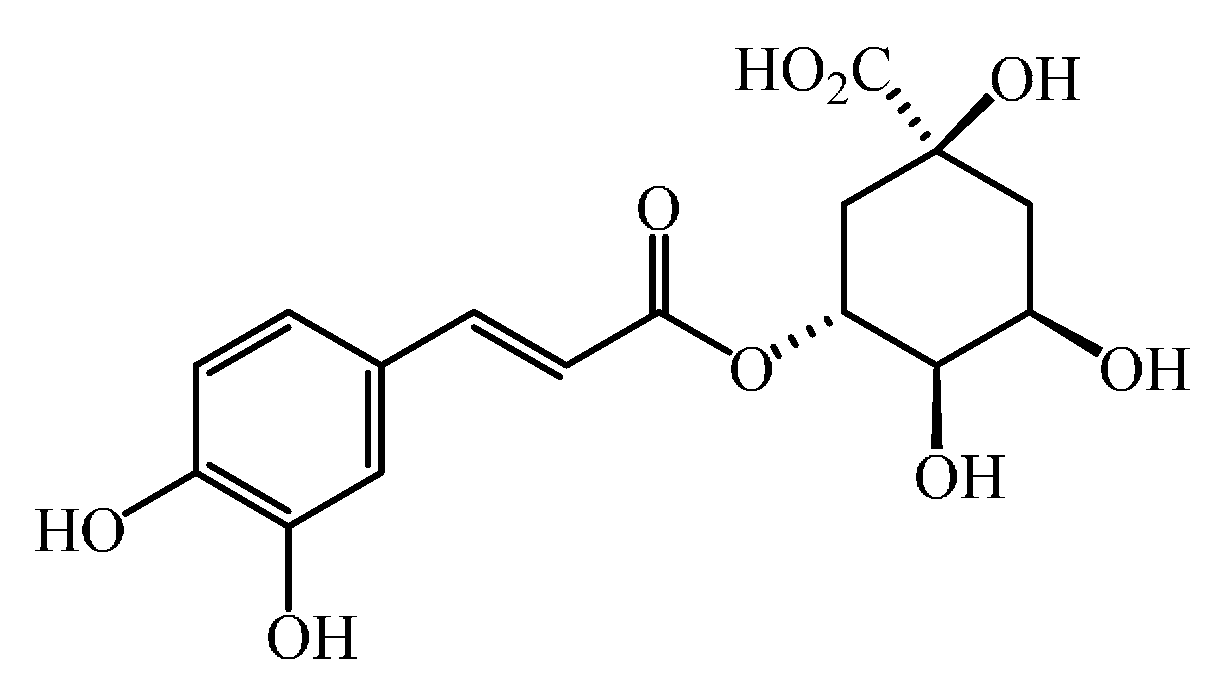
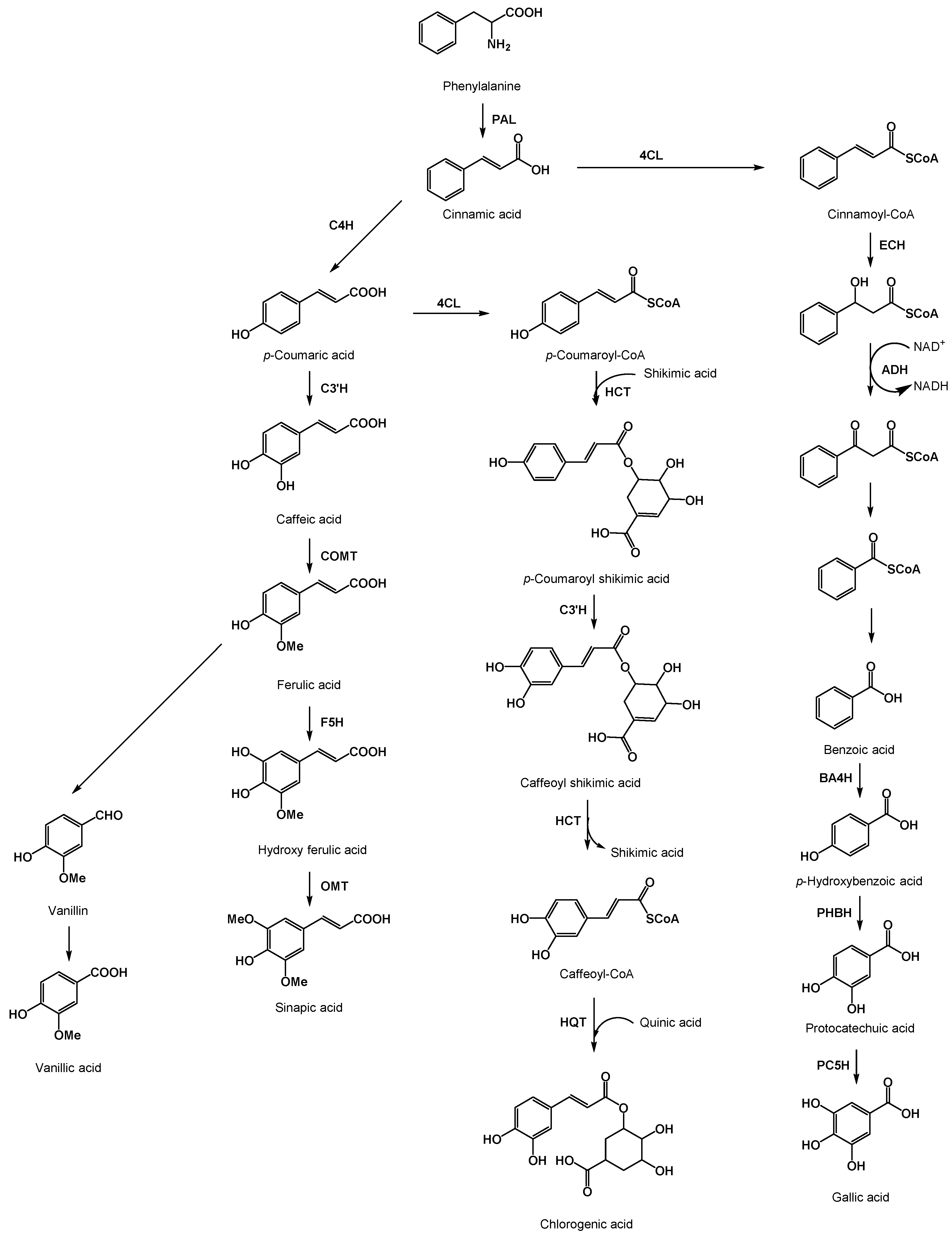
2. What Are Phenolic Acids?
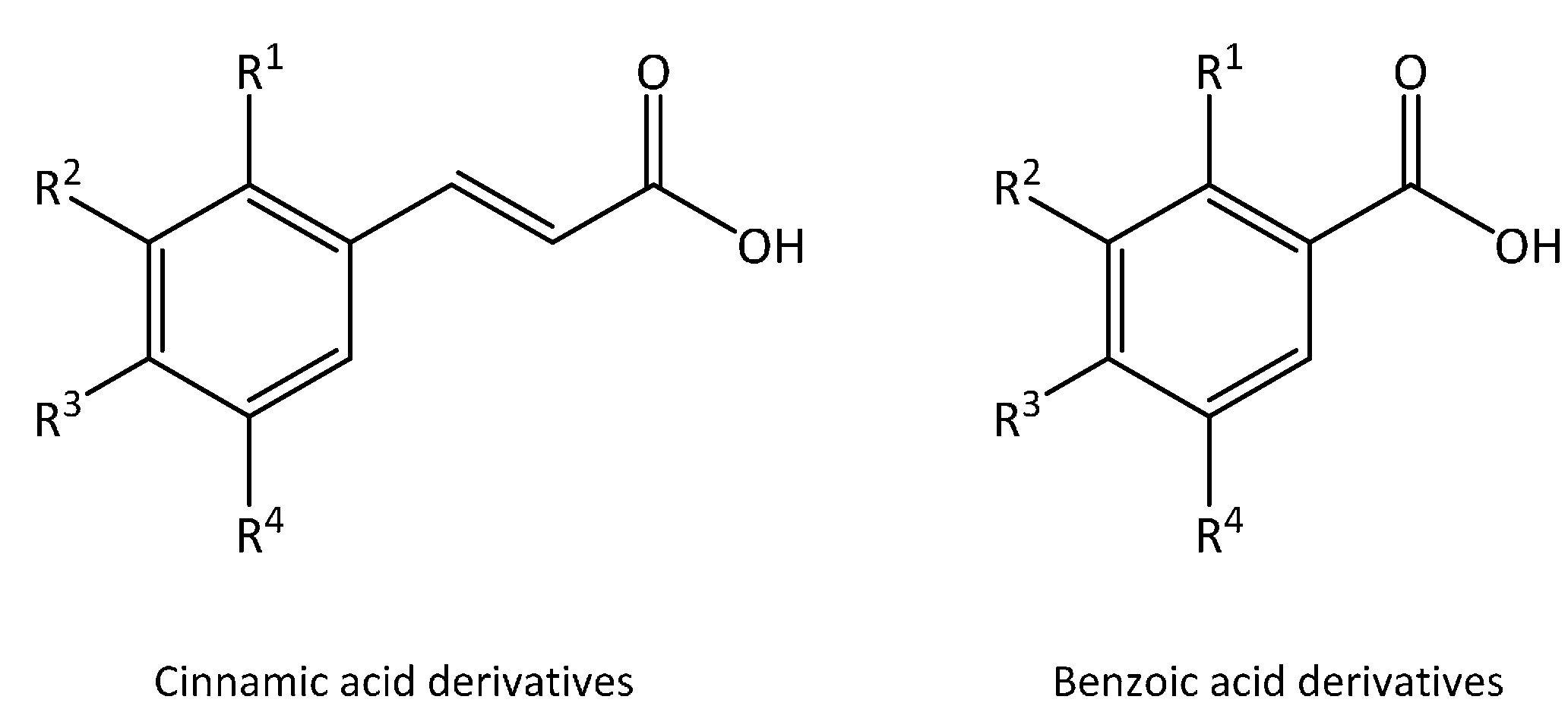
| Substitution | Cinnamic Acid Derivatives | Benzoic Acid Derivatives |
|---|---|---|
| R3 = OH | p-Coumaric acid | p-Hydroxybenzoic acid |
| R3 = R4 = OH | Caffeic acid | Protocatechuic acid |
| R2 = OCH3, R3 = OH | Ferulic acid | Vanillic acid |
| R2 = R3 = R4 = OH | Gallic acid | |
| R2 = R4 = OCH3, R3= OH | Sinapic acid | Syringic acid |
| R2 = R3 = OH [plus the carboxylic group being esterified with quinic acid] | 5-O-caffeoylquinic acid |
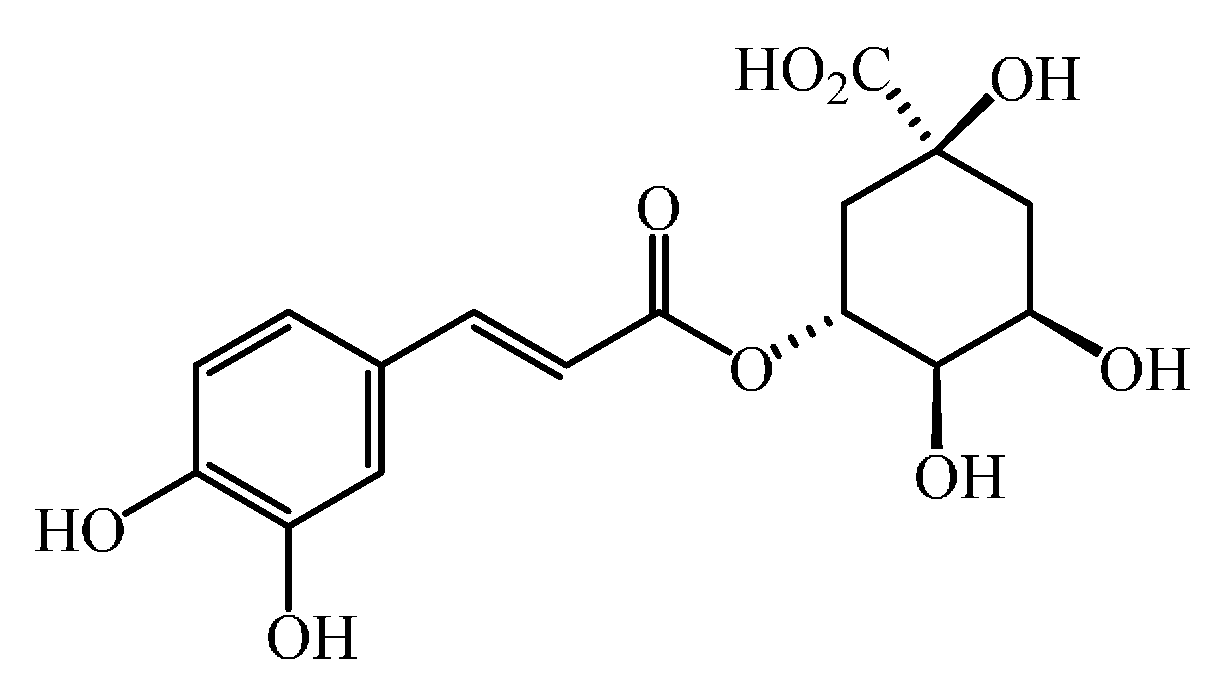
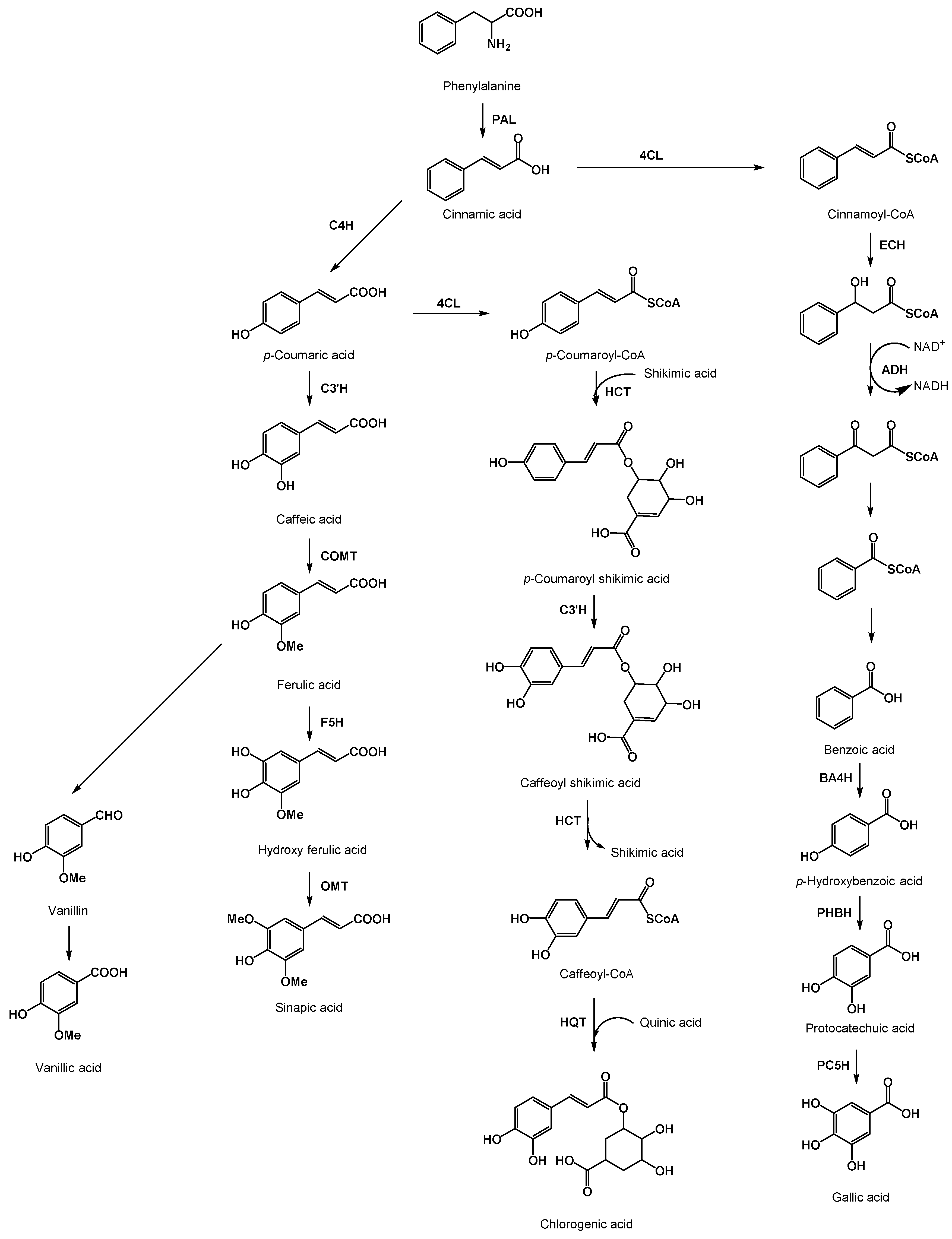
3. Bioactive Properties of Phenolic Acids
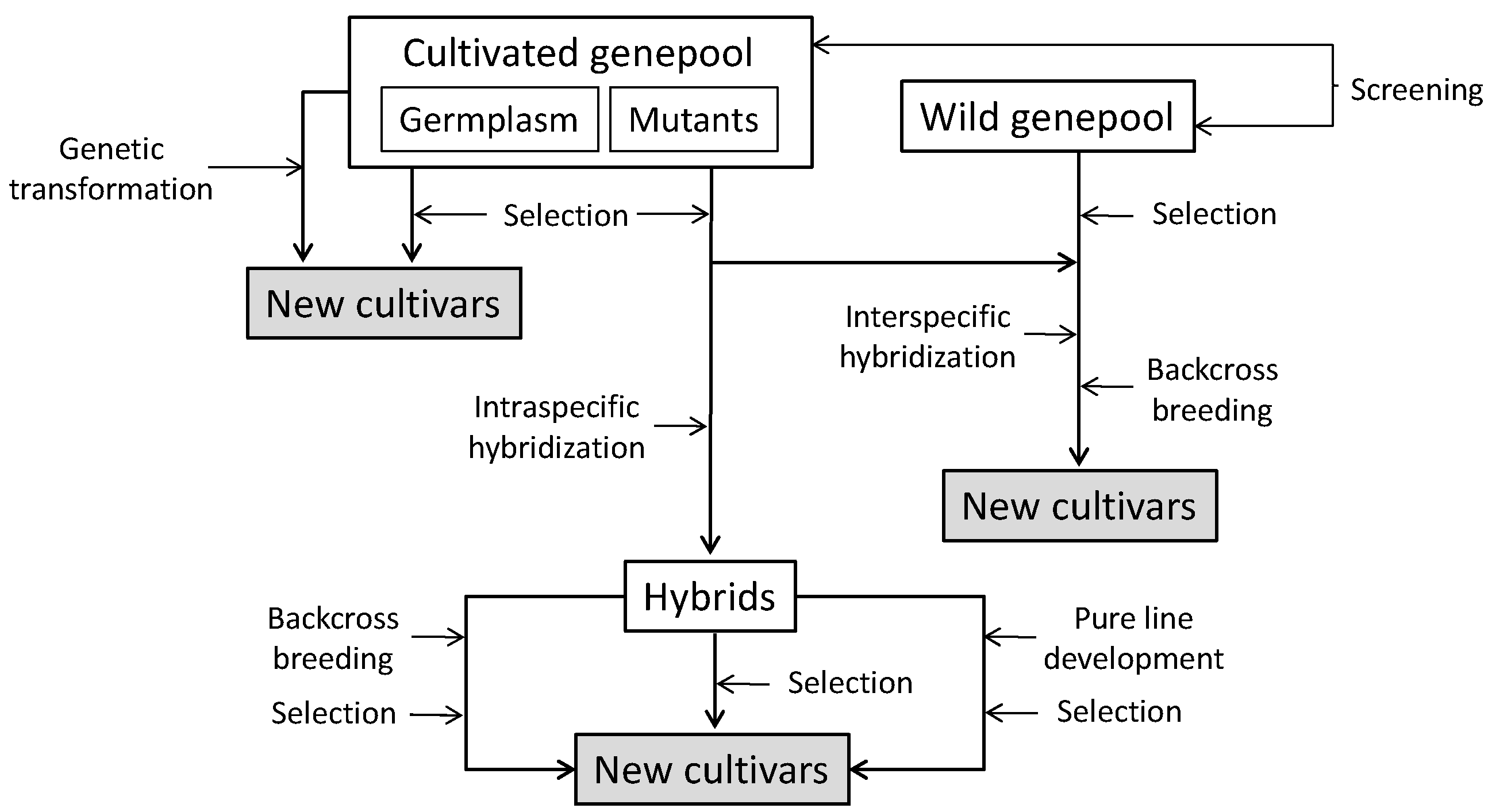
4. Breeding for Increased Phenolic Acids Content
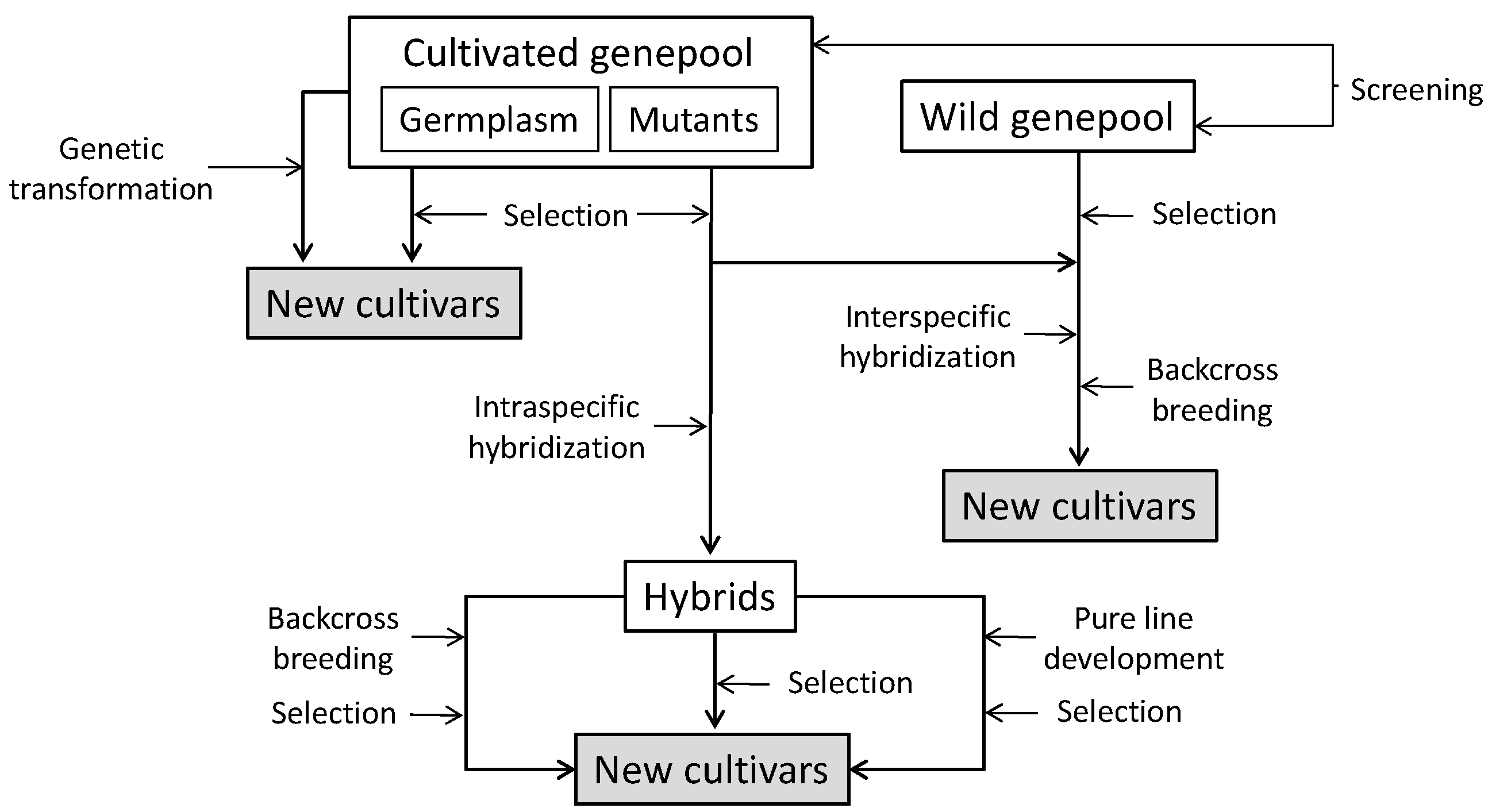
4.1. Identification of Sources of Variation
| Vegetable | Chlorogenic Acid [g·kg−1] | References |
|---|---|---|
| Artichoke (Cynara scolymus L.) | 0.4–7.3 | [45] |
| Carrot (Daucus carota) | 0.3–18.8 | [46] |
| Chicory (Cichorium intybus L.) | 0.1–0.9 | [47] |
| Eggplant (Solanum melongena) | 1.4–28.0 | [48,49] |
| Lettuce (Lactuca sativa L.) | 0.1–0.3 | [50] |
| Pepper (Capsicum annuum) | 0.7–0.9 | [51] |
| Tomato (Solanum esculentum) | 0.2–0.4 | [52] |
4.2. Gene Action and Heritability
4.3. QTL and Candidate Genes for Phenolic Acids Content
5. Genetic Transformation for Increasing Phenolic Acids Content
6. Collateral Effects of Breeding for Phenolic Acids in Vegetables
6.1. Biotic and Abiotic Stresses
6.2. Browning
6.3. Flavour
7. Future Prospects and Challenges
Acknowledgments
Conflicts of Interest
References
- Boeing, H.; Bechthold, A.; Bub, A.; Ellinger, S.; Haller, D.; Kroke, A.; Leschik-Bonnet, E.; Müller, M.J.; Oberritter, H.; Schulze, M.; et al. Critical review: Vegetables and fruit in the prevention of chronic diseases. Eur. J. Nutr. 2012, 51, 637–663. [Google Scholar] [CrossRef] [PubMed]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants, and functional foods: Impact on human health. Pharmacogn Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Fulton, S.L.; McKinley, M.C.; Young, I.S.; Cardwell, C.R.; Woodside, J.V. The effect of increasing fruit and vegetable consumption on overall diet: A systematic review and meta-analysis. Crit. Rev. Food Sci. Nutr. 2014, in press. [Google Scholar] [CrossRef] [PubMed]
- Leopoldini, M.; Marino, T.; Russo, N.; Toscano, M. Antioxidant properties of phenolic compounds: H-atom versus electron transfer mechanism. J. Phys. Chem. A 2004, 108, 4916–4922. [Google Scholar] [CrossRef]
- Piazzon, A.; Vrhovsek, U.; Masuero, D.; Mattivi, F.; Mandoj, F.; Nardini, M. Antioxidant activity of phenolic acids and their metabolites: Synthesis and antioxidant properties of the sulfate derivatives of ferulic and caffeic acids and of the acyl glucuronide of ferulic acid. J. Agric. Food Chem. 2012, 60, 12312–12323. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Kapoor, H.C. Antioxidants in fruits and vegetables the millennium’s health. Int. J. Food Sci. Technol. 2001, 36, 703–725. [Google Scholar] [CrossRef]
- Gülçin, I. Antioxidant activity of food constituents-An overview. Arch. Toxicol. 2012, 86, 345–396. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, A.; Klein, K. Antioxidants in Food, Vitamins and Supplements; Elsevier Inc.: San Diego, CA, USA, 2014. [Google Scholar]
- Volden, J.; Borge, G.I.A.; Bengtsson, G.B.; Hansen, M.; Thygesen, I.E.; Wicklund, T. Effect of thermal treatment on glucosinolates and antioxidant related parameters in red cabbage (Brassica oleracea L ssp. capitata f. rubra). Food Chem. 2008, 109, 595–605. [Google Scholar] [CrossRef]
- Balasundram, N.; Sundram, K.; Samman, S. Phenolic compounds in plants and agri-industrial by-products: Antioxidant activity, occurrence, and potential uses. Food Chem. 2006, 99, 191–203. [Google Scholar] [CrossRef]
- Herrmann, K.; Nagel, C.W. Occurrence and content of hydroxycinnamic and hydroxybenzoic acid compounds in foods. Crit. Rev. Food Sci. 1989, 28, 315–347. [Google Scholar] [CrossRef] [PubMed]
- Mattila, P.; Hellstrom, J. Phenolic acids in potatoes, vegetables, and some of their products. J. Food Compos. Anal. 2007, 20, 152–160. [Google Scholar] [CrossRef]
- Sands, D.C.; Morris, C.E.; Dratz, E.A.; Pilgeram, A. Elevating optimal human nutrition to a central goal of plant breeding and production of plant-based foods. Plant Sci. 2009, 177, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Diamanti, J.; Battino, M.; Mezzetti, B. Breeding for fruit nutritional and nutraceutical quality. In Breeding for Fruit Quality; Jenks, M.A., Bebeli, J.P., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 61–80. [Google Scholar]
- Treutter, D. Managing Phenol Contents in Crop Plants by Phytochemical Farming and Breeding Visions and Constraints. Int. J. Mol. Sci. 2010, 11, 807–857. [Google Scholar] [PubMed]
- Stalikas, C.D. Extraction, Separation, and Detection Methods for Phenolic Acids and Flavonoids. J. Sep. Sci. 2007, 30, 3268–3295. [Google Scholar] [CrossRef] [PubMed]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Poységu, L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chem. Int. 2011, 50, 586–621. [Google Scholar] [CrossRef] [PubMed]
- Goleniowski, M.; Bonfill, M.; Cusido, R.; Palazón, J. Phenolic Acids. In Natural Products: Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes; Ramawat, K.G., Mérillon, J.M., Eds.; Springer-Verlag: Berlin Heidelbergen, Germany, 2013; pp. 1951–1973. [Google Scholar]
- Mattila, P.; Hellstrom, J.; Torronen, R. Phenolic acids in berries, fruits and beverages. J. Agric. Food Chem. 2006, 54, 7193–7199. [Google Scholar] [CrossRef] [PubMed]
- Andjelkovic, M.; Camp, J.V.; Meulenaer, B.D.; Depaemelaere, G.; Socaciu, C.; Verloo, M.; Verhe, R. Iron-chelation properties of phenolic acids bearing catechol and galloyl groups. Food Chem. 2005, 98, 23–31. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; El-Said, A.M.A.; Khalifa, S.A.M.; Goransson, F.L.U.; Bohlin, L.; Borg-Karlson, K.A.; Verpoorte, R. Biosynthesis, natural sources, dietary intake, pharmacokinetic properties, and biological activities of hydroxycinnamic acids. J. Agric. Food Chem. 2012, 60, 10877–10895. [Google Scholar] [CrossRef] [PubMed]
- Prohens, J.; Rodríguez-Burruezo, A.; Raigón, M.D.; Nuez, F. Total phenolic concentration and browning susceptibility in a collection of different varietal types and hybrids of eggplant: Implications for breeding for higher nutritional quality and reduced browning. J. Am. Soc. Hortic. Sci. 2007, 132, 638–646. [Google Scholar]
- Upadhyay, R.; Mohan Rao, L.J. An outlook on chlorogenic acids-occurrence, chemistry, technology, and biological activities. Crit. Rev. Food Sci. Nutr. 2013, 53, 968–984. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.M.; Chakraborty, D.; Dey, S. Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signal Behav. 2010, 5, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Heleno, A.S.; Martins, A.; Queiroz, P.R.J.M.; Ferreira, R.F.C.I. Bioactivity of phenolic acids: Metabolites versus parent compounds: A review. Food Chem. 2015, 173, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Naczk, M. Phenolics in Food and Nutraceuticals; CRC Press: Boca Raton, FL, USA, 2004; pp. 1–3. [Google Scholar]
- Qualley, A.V.; Wildham, J.R.; Adebesin, F.; Kish, C.M.; Dudareva, N. Completion of the core β-oxidative pathway of benzoic acid biosynthesis in plants. Proc. Natl. Acad. Sci. USA 2012, 109, 16383–16388. [Google Scholar] [CrossRef] [PubMed]
- Gross, G.G. Biosynthesis and metabolism of phenolic acids and monolignols. In Biosynthesis and Biodegradation of Wood Components; Higuchi, T., Ed.; Academic Press: Orlando, FL, USA, 1985; pp. 229–271. [Google Scholar]
- Comino, C.; Lanteri, S.; Portis, E.; Acquadro, A.; Romani, A.; Hehn, A.; Larbat, R.; Bourgaud, F. Isolation and functional characterization of a cDNA coding a hydroxycinnamoyltransferase involved in phenylpropanoid biosynthesis in Cynara cardunculus L. Plant Sci. 2010, 179, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Niggeweg, R.; Michael, A.J.; Martin, C. Engineering plants with increased levels of the antioxidant chlorogenic acid. Nat. Biotechnol. 2004, 22, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Stratil, P.; Klejdus, B.; Kuban, V. Determination of total content of phenolic compounds and their antioxidant activity in vegetables-evaluation of spectrophotometric methods. J. Agric. Food Chem. 2006, 54, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R.; Deng, Z. Separation procedures for naturally occurring antioxidant phytochemicals. J. Chromatogr. B 2004, 812, 85–99. [Google Scholar] [CrossRef]
- Nardini, M.; Natella, F.; Scaccini, C.; Ghiselli, A. Phenolic acids from beer are absorbed and extensively metabolized in humans. J. Nutr. Biochem. 2006, 17, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Wittemer, S.M.; Ploch, M.; Windeck, T.; Muller, S.C.; Drewelow, B.; Derendorf, H.; Veit, M. Bioavailability and pharmacokinetics of caffeoylquinic acids and flavonoids after oral administration of artichoke leaf extracts in humans. Phytomedicine 2005, 12, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Roche, A.; Ross, E.; Walsh, N.; O’Donnell, K.; Williams, A.; Klapp, M.; Fullard, N.; Edelstein, S. Representative literature on the phytonutrients category: Phenolic acids. Crit. Rev. Food Sci. Nutr. 2015. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; Biswas, S.K.; Kirkham, P.A. Regulation of inflammation and redox signaling by dietary polyphenols. Biochem. Pharmacol. 2006, 72, 1439–1452. [Google Scholar] [CrossRef] [PubMed]
- Cho, A.S.; Jeon, S.M.; Kim, M.J.; Yeo, J.; Seo, K.I.; Choi, M.S.; Lee, M.K. Chlorogenic acid exhibits anti-obesity property and improves lipid metabolism in high-fat diet-induced-obesemice. Food Chem. Toxicol. 2010, 48, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Sova, M. Antioxidant and antimicrobial activities of cinnamic acid derivatives. Mini-Rev. Med. Chem. 2012, 12, 749–767. [Google Scholar] [CrossRef] [PubMed]
- Manuja, R.; Sachdeva, S.; Jain, A.; Chaudhary, J. A comprehensive Review on Biological Activities of P-Hydroxy Benzoic Acid and Its Derivatives. Int. J. Pharm. Sci. Rev. Res. 2013, 22, 109–115. [Google Scholar]
- Balyan, H.S.; Gupta, P.K.; Kumar, S.; Dhariwal, R.; Jaiswal, V.; Tyagi, S.; Agarwal, P.; Gahlaut, V.; Kumari, S. Genetic improvement of grain protein content and other health-related constituents of wheat grain. Plant Breed. 2013, 132, 446–457. [Google Scholar] [CrossRef]
- Farneti, B.; Masuero, D.; Costa, F.; Magnago, P.; Malnoy, M.; Costa, G.; Vrhovsek, U.; Mattivi, F. Is there room for improving the nutraceutical composition of apple? J. Agric. Food Chem. 2015, 63, 2750–2759. [Google Scholar] [CrossRef] [PubMed]
- Parr, A.J.; Bolwell, G.P. Phenols in the plant and in man: The potential for possible nutritional enhancement of the diet by modifying the phenols content or profile. J. Sci. Food Agric. 2000, 80, 985–1012. [Google Scholar] [CrossRef]
- Stommel, R.J.; Whitaker, D.B.; Haynes, G.K.; Prohens, J. Genotype x environment interactions in eggplant for fruit phenolic acid content. Euphytica 2015, 205, 823–836. [Google Scholar] [CrossRef]
- Yao, Y.; Sang, W.; Zhou, M.J.; Ren, G.X. Phenolic composition and antioxidant activities of 11 celery cultivars. J. Food Sci. 2010, 75, 9–13. [Google Scholar] [CrossRef]
- Alarcón-Flores, I.M.; Romero-González, R.; Martinez Vidal, LJ.; Garrido Frenich, A. Determination of phenolic compounds in artichoke, garlic and spinach by Ultra-High-Performance Liquid Chromatography coupled to Tandem Mass Spectrometry. Food Anal. Methods 2014, 7, 2095–2106. [Google Scholar] [CrossRef]
- Sun, T.; Simon, P.W.; Tanumihardjo, S.A. Antioxidant phytochemicals and antioxidant capacity of biofortified carrots (Daucus carota L.) of various colors. J. Agric. Food Chem. 2009, 57, 4142–4147. [Google Scholar] [CrossRef] [PubMed]
- Sinkovic, L.; Demsar, L.; Znidarcic, D.; Vidrih, R.; Hribar, J.; Treutter, D. Phenolic profiles in leaves of chicory cultivars (Cichorium intybus L.) as influenced by organic and mineral fertilizers. Food Chem. 2015, 166, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Luthria, D.L. A simplified UV spectral scan method for the estimation of phenolic acids and antioxidant capacity in eggplant pulp extracts. J. Funct. Foods 2012, 4, 238–242. [Google Scholar] [CrossRef]
- Mennella, G.; Lo Scalzo, R.; Fibiani, M.; D’Alessandro, A.; Francese, G.; Toppino, L.; Acciarri, N.; de Almeida, A.E.; Rotino, G.L. Chemical and bioactive quality traits during fruit ripening in eggplant (S. melongena L.) and allied species. J. Agric. Food Chem. 2012, 60, 11821–11831. [Google Scholar] [CrossRef] [PubMed]
- López, A.; Javier, G.A.; Fenoll, J.; Hellín, P.; Flores, P. Chemical composition and antioxidant capacity of lettuce: Comparative study of regular-sized (Romaine) and baby-sized (Little Gem and Mini Romaine) types. J. Food Compos. Anal. 2014, 33, 39–48. [Google Scholar] [CrossRef]
- Hallmann, E.; Rembiałkowska, E. Characterisation of antioxidant compounds in sweet bell pepper (Capsicum annuum L.) under organic and conventional growing systems. J. Sci. Food Agric. 2012, 92, 2409–2415. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, E. The influence of organic and conventional cultivation systems on the nutritional value and content of bioactive compounds in selected tomato types. J. Sci. Food Agric. 2012, 92, 2840–2848. [Google Scholar] [CrossRef] [PubMed]
- Leja, M.; Kamińska, I.; Kramer, M.; Maksylewicz-Kaul, A.; Kammerer, D.; Carle, R.; Baranski, R. The content of phenolic compounds and radical scavenging activity varies with carrot origin and root color. Plant Foods Hum. Nutr. 2013, 68, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Vera-Guzmán, M.A.; Chávez-Servia, L.J.; Carrillo-Rodríguez, C.J.; López, G.M. Phytochemical evaluation of wild and cultivated pepper (Capsicum annuum L. and C. pubescens Ruiz & Pav.) from Oaxaca, Mexico. Chilean J. Agric. Res. 2011, 71, 578–585. [Google Scholar]
- Stushnoff, C.; Holm, D.; Thomson, M.D.; Jiang, W.; Thomson, H.J.; Joyce, N.I.; Wilson, P. Antioxidant properties of cultivars and selections from the Colorado potato breeding program. Am. J. Potato. Res. 2008, 85, 267–276. [Google Scholar] [CrossRef]
- Meyer, S.R.; Whitaker, D.B.; Little, P.D.; Wu, B.S.; Kennelly, J.E.; Long, L.C.; Little, A. Parallel reductions in phenolic constituents resulting from the domestication of eggplant. Phytochemistry 2015, 115, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Millar, D.J.; Kimura, Y.; Donovan, G.; Rees, J.; Fraser, P.D.; Bramley, P.M.; Bolwell, G.P. Metabolite profiling of carotenoid and phenolic pathways in carotenoid and transgenic pathways in mutant and transgenic lines of tomato: Identification of a high antioxidant fruit line. Phytochemistry 2006, 67, 1750–1757. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.D.; Pogrebnyak, N.; Kuhn, P.; Krueger, C.G.; Johnson, W.D.; Raskin, I. Development and phytochemical characterization of high polyphenol red lettuce with anti-diabetic properties. PLoS ONE 2014, 9, e91571. [Google Scholar] [CrossRef] [PubMed]
- Meléndez-Martínez, J.A.; Fraser, D.P.; Bramley, M.P. Accumulation of health promoting phytochemicals in wild relatives of tomato and their contribution to in vitro antioxidant activity. Phytochemistry 2010, 71, 1104–1114. [Google Scholar]
- Prohens, J.; Whitaker, D.B.; Plazas, M.; Vilanova, S.; Hurtado, M.; Blasco, M.; Gramazio, P.; Stommel, R.J. Genetic diversity in morphological characters and phenolic acids content resulting from an interspecific cross between eggplant, Solanum melongena, and its wild ancestor (S. incanum). Ann. Appl. Biol. 2013, 162, 242–257. [Google Scholar] [CrossRef]
- Gramazio, P.; Prohens, J.; Plazas, M.; Andújar, I.; Castillo, E.; Herraiz, F.J.; Knapp, S.; Meyer, S.R.; Vilanova, S. Location of the chlorogenic acid biosynthesis pathway and polyphenol oxidase genes in a new interspecific anchored linkage map of eggplant. BMC Plant. Biol. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Mennella, G.; Rotino, G.L.; Fibiani, M.; D’Alessandro, A.; Francese, G.; Toppino, L.; Cavallanti, F.; Acciarri, N.; Lo Scalzo, R. Characterization of health-related compounds in eggplant (Solanum melongena L.) lines derived from introgression of allied species. J. Agric. Food Chem. 2010, 58, 7597–7603. [Google Scholar] [CrossRef] [PubMed]
- Kalloo, G.; Chowdhury, J.B. Distant Hybridization of Crop Plants; Springer: Verlag Berlin, Heidelberg, Germany, 1992; Volume 16, p. 271. [Google Scholar]
- Hajjar, R.; Hodgkin, T. The use of wild relatives in crop improvement: A survey of developments over the last 20 years. Euphytica 2007, 156, 1–13. [Google Scholar] [CrossRef]
- Plazas, M.; Prohens, J.; Cuñat, N.A.; Vilanova, S.; Gramazio, P.; Herraiz, J.F.; Andújar, I. Reducing capacity, chlorogenic acid content and biological activity in a collection of scarlet (Solanum aethiopicum) and gboma (S. macrocarpon) eggplants. Int. J. Mol. Sci. 2014, 15, 17221–17241. [Google Scholar] [CrossRef] [PubMed]
- Pék, Z.; Daood, H.; Nagyné, M.G.; Neményi, A.; Helyes, L. Effect of environmental conditions and water status on the bioactive compounds of broccoli. Cent. Eur. J. Biol. 2013, 8, 777–787. [Google Scholar] [CrossRef]
- Helyes, L.; Lugasi, A.; Daood, H.G.; Pék, Z. The simultaneous effect of water supply and genotype on yield quantity, antioxidants content and composition of processing tomatoes. Not. Bot. Horti. Agrobo. 2014, 42, 143–149. [Google Scholar] [CrossRef]
- El-Soda, M.; Malosetti, M.; Zwaan, B.J.; Koornneef, M.; Aarts, M.G. Genotype × environment interaction QTL mapping in plants: Lessons from Arabidopsis. Trends Plant Sci. 2014, 19, 390–398. [Google Scholar] [CrossRef]
- Payyavula, S.R.; Shakya, R.; Sengoda, G.V.; Munyaneza, E.J.; Swamy, P.; Navarre, A.D. Synthesis and regulation of chlorogenic acid in potato: Rerouting phenylpropanoid flux in HQT-silenced lines. Plant. Biotechnol. J. 2015, 13, 551–564. [Google Scholar] [CrossRef]
- Di Matteo, A.; Ruggieri, V.; Sacco, A.; Rigano, M.M.; Carriero, F.; Bolger, A.; Fernie, A.R.; Frusciante, L.; Barone, A. Identification of candidate genes for phenolics accumulation in tomato fruit. Plant Sci. 2013, 205–206, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Schoch, G.; Goepfert, S.; Morant, M.; Hehn, A.; Meyer, D.; Ullmann, P.; Werck-Reichhart, D. CYP98A3 from Arabidopsis thaliana is a 3-hydroxylase of phenolic esters, a missing link in the phenylpropanoid pathway. J. Biol. Chem. 2001, 276, 36566–36574. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Naqvi, S.; Gomez-Galera, S.; Pelacho, A.M.; Capell, T.; Christou, P. Transgenic strategies for the nutritional enhancement of plants. Trends Plant Sci. 2007, 12, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Amaya, I.; Osorio, S.; Martinez-Ferri, E.; Lima-Silva, V.; Doblas, G.V.; Fernández-Muñoz, R.; Fernie, R.A.; Botella, A.M.; Valpuesta, V. Increased antioxidant capacity in tomato by ectopic expression of the strawberry D-galacturonate reductase gene. Biotechnol. J. 2015, 10, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Lukaszewicz, M.; Matysiak-Kata, I.; Skala, J. Antioxidant capacity manipulation in transgenic potato tuber by changes in phenolic content. J. Agric. Food Chem. 2004, 52, 1526–1533. [Google Scholar] [CrossRef] [PubMed]
- Nicolia, A.; Manzo, A.; Veronesi, F.; Rosellini, D. An overview of the last 10 years of genetically engineered crop safety research. Crit. Rev. Biotechnol. 2014, 34, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, E.; Schouten, H.J. Cisgenesis strongly improves introgression breeding and induced translocation breeding of plants. Trends Biotechnol. 2007, 25, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Robbins, R.J. Phenolic acids in foods: An overview of analytical methodology. J. Agric. Food Chem. 2003, 51, 2866–2887. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.C. Determinants of eating quality in fruits and vegetables. Proc. Nutr. Soc. Aust. 2000, 24, 183–190. [Google Scholar]
- Dixon, A.R. Progress natural products and plant disease resistance. Nature 2001, 411, 843–847. [Google Scholar] [CrossRef]
- Singhai, K.P.; Sarma, K.B.; Srivastava, S.J. Phenolic acid content in potato peel determines natural infection of common scab caused by Streptomyces spp. World J. Microbiol. Biotechnol. 2011, 27, 1559–1567. [Google Scholar] [CrossRef]
- De Ascensao, A.R.; Dubery, I.A. Soluble and wall bound phenolic polymers in Musa acuminata roots exposed to elicitors from Fusarium oxysporum f.sp. cubense. Phytochemistry 2003, 63, 679–686. [Google Scholar] [CrossRef]
- Ding, H.; Lamb, R.J.; Ames, N. Inducible production of phenolic acids in wheat and antibiotic resistance to Sitodiplosis mosellana. J. Chem. Ecol. 2000, 26, 969–985. [Google Scholar] [CrossRef]
- Leiss, K.A.; Maltese, F.; Choi, Y.H.; Verpoorte, R.; Klinkhamer, P.G. Identification of chlorogenic acid as a resistance factor for thrips in chrysanthemum. Plant Physiol. 2009, 150, 1567–1575. [Google Scholar] [CrossRef]
- Shivashankar, S.; Sumathi, M.; Krishnakumar, K.N.; Rao, K.V. Role of phenolic acids and enzymes of phenylpropanoid pathway in resistance of chayote fruit (Sechium edule) against infestation by melon fly (Bactrocera cucurbitae). Ann. Appl. Biol. 2015, 166, 420–433. [Google Scholar] [CrossRef]
- Chitwood, D.J. Phytochemical based strategies for nematode control. Annu. Rev. Phytopathol. 2002, 40, 221–249. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, H.; Baatour, O.; Huang, J.; Nasri, N.; Salah, B.I.; Tarchoun, I.; Zaghdoudi, M.; Gruber, Y.M.; Lachaal, M.; Ouerghi, Z.; et al. Verte and Romaine lettuce varieties (Lactuca sativa L.) show differential responses to high NaCl concentrations. J. Plant Nutr. Soil Sci. 2012, 175, 641–648. [Google Scholar] [CrossRef]
- Lattanzio, V.; Cardinali, A.; di Venere, D.; Linsalata, V.; Palmeri, S. Browning phenomena in stored artichoke (Cynara scolymus L.) heads: Enzymic or chemical reactions? Food Chem. 1994, 50, 1–7. [Google Scholar] [CrossRef]
- Queiroz, C.; Lopes, M.L.M.; Fialho, E.; Valente-Mesquita, V.L. Polyphenol oxidase: Characteristics and mechanisms of browning control. Food Rev. Int. 2008, 24, 361–375. [Google Scholar] [CrossRef]
- Fruit Phenolics; Macheix, J.J.; Fleuriet, A.; Billot, J. (Eds.) CRC Press: Boca Raton, FL, USA, 1990; pp. 1–126.
- Plazas, M.; López-Gresa, M.P.; Vilanova, S.; Torres, C.; Hurtado, M.; Gramazio, P.; Andújar, I.; Herráiz, F.J.; Bellés, J.M.; Prohens, J. Diversity and relationships in key traits for functional and apparent quality in a collection of eggplant: Fruit phenolics content, antioxidant activity, polyphenol oxidase activity, and browning. J. Agric. Food Chem. 2013, 61, 8871–8879. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Nishimura, M.; Murai, N.; Haruta, M.; Homma, S.; Itoh, Y. A transgenic apple callus showing reduced polyphenol oxidase activity and lower browning potential. Biosci. Biotechnol. Biochem. 2001, 65, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Chi, M.; Bhagwat, B.; Lane, W.D.; Tang, G.L.; Su, Y.Q.; Sun, R.C.; Oomah, B.D.; Wiersma, P.A.; Xiang, Y. Reduced polyphenol oxidase gene expression and enzymatic browning in potato (Solanum tuberosum L.) with artificial microRNAs. BMC Plant Biol. 2014, 14, 62. [Google Scholar] [CrossRef] [PubMed]
- Buffo, R.A.; Cardelli-Freire, C. Coffee flavour: An overview. Flavour Frag. J. 2004, 19, 99–104. [Google Scholar] [CrossRef]
- Nagel, C.W.; Herrick, I.W.; Graber, W.R. Is chlorogenic acid bitter? J. Food Sci. 1987, 52, 213–213. [Google Scholar] [CrossRef]
- Dawid, C.; Hofmann, T. Quantitation and bitter taste contribution of saponins in fresh and cooked white asparagus (Asparagus officinalis L.). Food Chem. 2014, 145, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Kreutzmann, S.; Christensen, L.P.; Edelenbos, M. Investigation of bitterness in carrots (Daucus carota L.) based on quantitative chemical and sensory analyses. LWT—Food Sci. Technol. 2008, 41, 193–205. [Google Scholar] [CrossRef]
- Tordoff, M.G.; Sandell, M.A. Vegetable bitterness is related to calcium content. Appetite 2009, 52, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Barkley, N.A.; Wang, M.L. Application of TILLING and EcoTILLING as reverse genetic approaches to elucidate the function of genes in plants and animals. Curr. Genomics. 2008, 9, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Morrell, P.L.; Buckler, E.S.; Ross-Ibarra, J. Crop genomics: Advances and applications. Nat. Rev. Genet. 2012, 13, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Juste, J.; Cutler, R.S. Plant genome engineering in full bloom. Trends Plant Sci. 2014, 19, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Araki, M.; Ishii, T. Towards social acceptance of plant breeding by genome editing. Trends Plant Sci. 2015, 20, 145–149. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaushik, P.; Andújar, I.; Vilanova, S.; Plazas, M.; Gramazio, P.; Herraiz, F.J.; Brar, N.S.; Prohens, J. Breeding Vegetables with Increased Content in Bioactive Phenolic Acids. Molecules 2015, 20, 18464-18481. https://doi.org/10.3390/molecules201018464
Kaushik P, Andújar I, Vilanova S, Plazas M, Gramazio P, Herraiz FJ, Brar NS, Prohens J. Breeding Vegetables with Increased Content in Bioactive Phenolic Acids. Molecules. 2015; 20(10):18464-18481. https://doi.org/10.3390/molecules201018464
Chicago/Turabian StyleKaushik, Prashant, Isabel Andújar, Santiago Vilanova, Mariola Plazas, Pietro Gramazio, Francisco Javier Herraiz, Navjot Singh Brar, and Jaime Prohens. 2015. "Breeding Vegetables with Increased Content in Bioactive Phenolic Acids" Molecules 20, no. 10: 18464-18481. https://doi.org/10.3390/molecules201018464
APA StyleKaushik, P., Andújar, I., Vilanova, S., Plazas, M., Gramazio, P., Herraiz, F. J., Brar, N. S., & Prohens, J. (2015). Breeding Vegetables with Increased Content in Bioactive Phenolic Acids. Molecules, 20(10), 18464-18481. https://doi.org/10.3390/molecules201018464