
Cross-Amplification of Vicia sativa subsp. sativa Microsatellites across 22 Other Vicia Species

Abstract
:1. Introduction
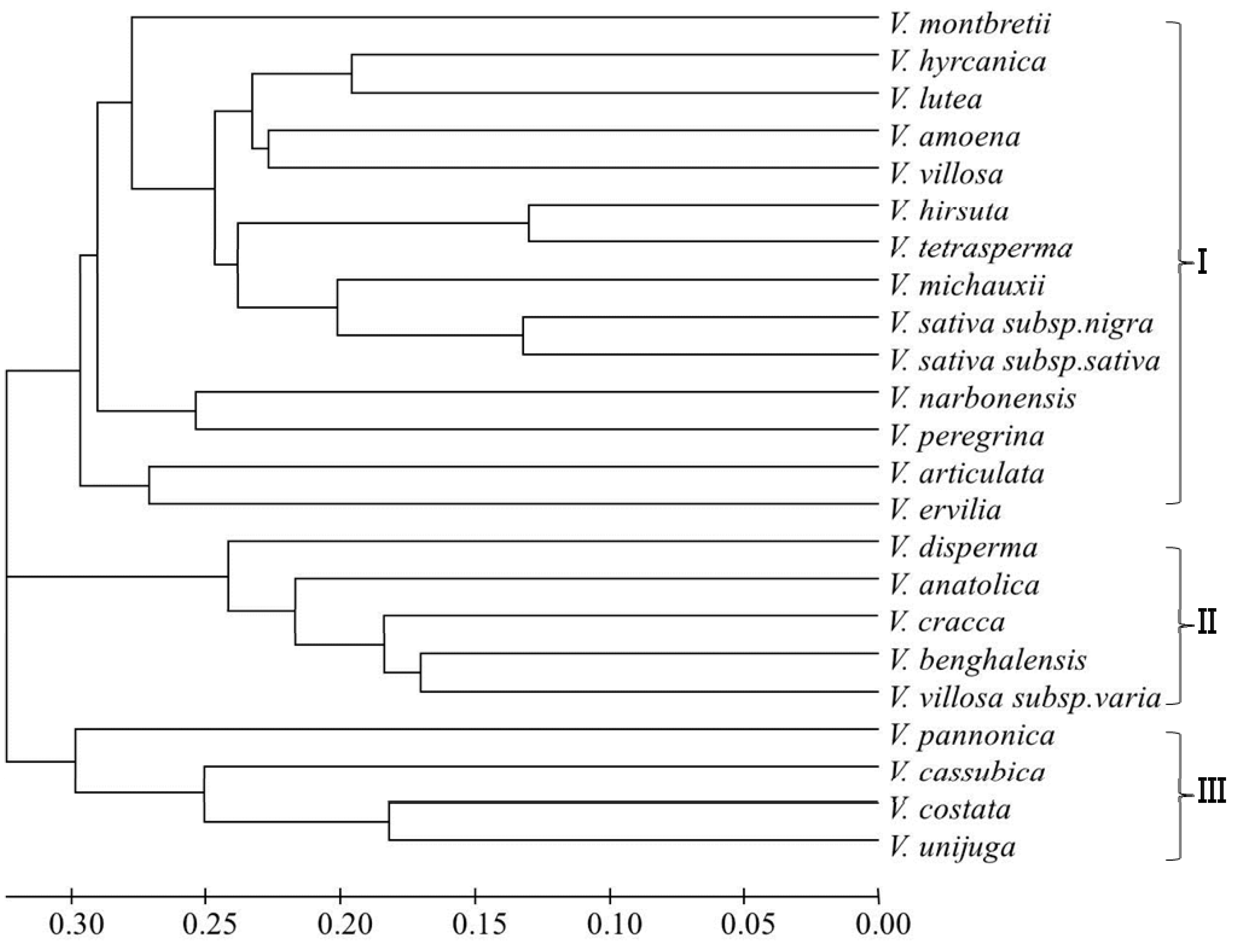
2. Results and Discussion

{kind=link}
| Species (Sample Size) | Theoretical Amplicons (No.) | Amplicons (No.) | Transferability (%) |
|---|---|---|---|
| V. amoena (4) | 144 | 77 | 53 |
| V. anatolica (2) | 72 | 36 | 50 |
| V. articulata (4) | 144 | 51 | 35 |
| V. benghalensis (4) | 144 | 72 | 50 |
| V. cassubica (2) | 72 | 28 | 39 |
| V. costata (2) | 72 | 25 | 35 |
| V. cracca (4) | 144 | 73 | 51 |
| V. disperma (2) | 72 | 30 | 42 |
| V. ervilia (4) | 144 | 47 | 33 |
| V. hirsuta (2) | 72 | 40 | 56 |
| V. hyrcanica (4) | 144 | 85 | 59 |
| V. lutea (2) | 72 | 53 | 74 |
| V. michauxii (4) | 144 | 84 | 58 |
| V. montbretii (4) | 144 | 90 | 62 |
| V. narbonensis (4) | 144 | 66 | 46 |
| V. pannonica (2) | 72 | 31 | 43 |
| V. peregrina (4) | 144 | 74 | 51 |
| V. sativa subsp. nigra (8) | 288 | 236 | 82 |
| V. sativa subsp. sativa (14) | 504 | 473 | 94 |
| V. tetrasperma (4) | 144 | 78 | 54 |
| V. unijuga (2) | 72 | 35 | 49 |
| V. villosa (4) | 144 | 95 | 66 |
| V. villosa subsp. varia (2) | 72 | 43 | 60 |
| Mean | 54 |
| Marker | NA | MAF | HO | HE | PIC |
|---|---|---|---|---|---|
| GBSSR-VSspS-020 | 3 | 0.83 | 0.00 | 0.30 | 0.26 |
| GBSSR-VSspS-023 | 4 | 0.42 | 0.09 | 0.70 | 0.63 |
| GBSSR-VSspS-073 | 5 | 0.52 | 0.19 | 0.70 | 0.60 |
| GBSSR-VSspS-076 | 4 | 0.66 | 0.11 | 0.50 | 0.43 |
| GBSSR-VSspS-080 | 5 | 0.47 | 0.11 | 0.70 | 0.63 |
| GBSSR-VSspS-079 | 6 | 0.63 | 0.05 | 0.60 | 0.53 |
| GBSSR-VSspS-090 | 9 | 0.30 | 0.06 | 0.80 | 0.79 |
| GBSSR-VSspS-099 | 6 | 0.47 | 0.13 | 0.70 | 0.62 |
| GBSSR-VSspS-107 | 10 | 0.50 | 0.23 | 0.70 | 0.64 |
| GBSSR-VSspS-102 | 5 | 0.52 | 0.10 | 0.60 | 0.56 |
| GBSSR-VSspS-125 | 6 | 0.50 | 0.13 | 0.70 | 0.66 |
| GBSSR-VSspS-119 | 6 | 0.64 | 0.17 | 0.50 | 0.45 |
| GBSSR-VSspS-140 | 3 | 0.48 | 0.00 | 0.60 | 0.56 |
| GBSSR-VSspS-126 | 3 | 0.50 | 0.00 | 0.60 | 0.55 |
| GBSSR-VSspS-166 | 2 | 0.72 | 0.00 | 0.40 | 0.32 |
| GBSSR-VSspS-179 | 12 | 0.27 | 0.16 | 0.90 | 0.84 |
| GBSSR-VSspS-181 | 9 | 0.61 | 0.35 | 0.60 | 0.57 |
| GBSSR-VSspS-203 | 3 | 0.61 | 0.09 | 0.50 | 0.45 |
| GBSSR-VSspS-247 | 3 | 0.82 | 0.00 | 0.30 | 0.28 |
| GBSSR-VSspS-301 | 4 | 0.67 | 0.00 | 0.50 | 0.45 |
| GBSSR-VSspS-310 | 7 | 0.54 | 0.36 | 0.60 | 0.60 |
| GBSSR-VSspS-313 | 2 | 0.97 | 0.00 | 0.10 | 0.06 |
| GBSSR-VSspS-067 | 4 | 0.94 | 0.03 | 0.10 | 0.12 |
| GBSSR-VSspS-251 | 20 | 0.46 | 0.69 | 0.80 | 0.74 |
| GBSSR-VSspS-037 | 11 | 0.37 | 0.25 | 0.80 | 0.77 |
| GBSSR-VSspS-268 | 4 | 0.53 | 0.11 | 0.60 | 0.56 |
| GBSSR-VSspS-042 | 7 | 0.49 | 0.10 | 0.70 | 0.67 |
| GBSSR-VSspS-305 | 10 | 0.44 | 0.13 | 0.70 | 0.60 |
| GBSSR-VSspS-088 | 2 | 0.76 | 0.08 | 0.40 | 0.30 |
| GBSSR-VSspS-308 | 6 | 0.91 | 0.05 | 0.20 | 0.17 |
| GBSSR-VSspS-024 | 3 | 0.90 | 0.00 | 0.20 | 0.18 |
| GBSSR-VSspS-028 | 2 | 0.90 | 0.07 | 0.20 | 0.16 |
| GBSSR-VSspS-075 | 6 | 0.38 | 0.08 | 0.70 | 0.68 |
| GBSSR-VSspS-091 | 1 | 1.00 | 0.00 | 0.00 | 0.00 |
| GBSSR-VSspS-115 | 17 | 0.35 | 0.27 | 0.80 | 0.82 |
| GBSSR-VSspS-129 | 16 | 0.25 | 0.21 | 0.90 | 0.83 |
| Mean | 6.3 | 0.59 | 0.12 | 0.50 | 0.50 |
3. Experimental Section
3.1. Plant Materials
3.2. Amplification of SSRs
3.3. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Frediani, M.; Maggini, F.; Gelati, M.T.; Cremonini, R. Repetitive DNA sequences as probes for phylogenetic analysis in vicia genus. Caryologia 2004, 57, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Hanelt, P.; Mettin, D. Biosystematics of the genus Vicia L. (leguminosae). Annu. Rev. Ecol. Syst. 1989, 20, 199–223. [Google Scholar]
- Haider, A.S.; El-Shanshoury, A.R. Variability of storage proteins and esterase isozymes in vicia sativa subspecies. Biol. Plant. 2000, 43, 205–209. [Google Scholar] [CrossRef]
- Naranjo, C.A.; Ferrari, M.R.; Palermo, A.M.; Poggio, L. Karyotype, DNA content and meiotic behaviour in five south american species of vicia (fabaceae). Ann. Bot. Lond. 1998, 82, 757–764. [Google Scholar] [CrossRef] [Green Version]
- Maxted, N.; Callimassia, M.A.; Bennett, M.D. Cytotaxonomic studies of eastern mediterranean vicia species (leguminosae). Plant Syst. Evol. 1991, 177, 221–234. [Google Scholar] [CrossRef]
- Navratilova, A.; Neumann, P.; Macas, J. Karyotype analysis of four vicia species using in situ hybridization with repetitive sequences. Ann. Bot. 2003, 91, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Gil, J.; Cubero, J.I. Multivariate analysis of the vicia sativa l. Aggregate. Bot. J. Linn. Soc. 1993, 113, 389–400. [Google Scholar] [CrossRef]
- Gupta, S.K.; Niranjan, D.G.; Agrawal, S.S.; Srivastava, S.; Saxena, R. Recent advances in pharmacotherapy of glaucoma. Indian J. Pharmacol. 2008, 40, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Erpelding, J.E.; Blake, N.K.; Blake, T.K.; Talbert, L.E. Transfer of sequence tagged site pcr markers between wheat and barley. Genome 1996, 39, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Sigmund, R.; Borner, A.; Korzun, V.; Stein, N.; Sorrells, M.E.; Langridge, P.; Graner, A. Interspecific transferability and comparative mapping of barley est-ssr markers in wheat, rye and rice. Plant Sci. 2005, 168, 195–202. [Google Scholar] [CrossRef]
- Gutierrez, M.V.; Patto, M.C.V.; Huguet, T.; Cubero, J.I.; Moreno, M.T.; Torres, A.M. Cross-species amplification of medicago truncatula microsatellites across three major pulse crops. Theor. Appl. Genet. 2005, 110, 1210–1217. [Google Scholar] [CrossRef] [PubMed]
- Powell, W.; Machray, G.C.; Provan, J. Polymorphism revealed by simple sequence repeats. Trends Plant Sci. 1996, 1, 215–222. [Google Scholar] [CrossRef]
- Suresh, S.; Park, J.H.; Cho, G.T.; Lee, H.S.; Baek, H.J.; Lee, S.Y.; Chung, J.W. Development and molecular characterization of 55 novel polymorphic cdna-ssr markers in faba bean (Vicia faba L.) using 454 pyrosequencing. Molecules 2013, 18, 1844–1856. [Google Scholar]
- Scott, K.D.; Eggler, P.; Seaton, G.; Rossetto, M.; Ablett, E.M.; Lee, L.S.; Henry, R.J. Analysis of ssrs derived from grape ests. Theor. Appl. Genet. 2000, 100, 723–726. [Google Scholar] [CrossRef]
- Fredslund, J.; Madsen, L.H.; Hougaard, B.K.; Sandal, N.; Stougaard, J.; Bertioli, D.; Schauser, L. Gemprospector—Online design of cross-species genetic marker candidates in legumes and grasses. Nucleic Acids Res. 2006, 34, W670–W675. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jafari, N.; Behroozi, R.; Bagheri, A.; Moshtaghi, N. Determination of genetic diversity of cultivated chickpea (cicer arietinum l.) using medicago truncatula est-ssrs. J. Plant Mol. Breed. 2013, 2, 1–16. [Google Scholar]
- Weeden, N.F.; Muehlbauer, F.J.; Ladizinsky, G. Extensive conservation of linkage relationships between pea and lentil genetic maps. J. Hered. 1992, 83, 123–129. [Google Scholar]
- Pandian, A.; Ford, R.; Taylor, P.J. Transferability of sequence tagged microsatellite site (stms) primers across four major pulses. Plant Mol. Biol. Rep. 2000, 18, 395–395. [Google Scholar] [CrossRef]
- Kalo, P.; Seres, A.; Taylor, S.A.; Jakab, J.; Kevei, Z.; Kereszt, A.; Endre, G.; Ellis, T.H.N.; Kiss, G.B. Comparative mapping between medicago sativa and pisum sativum. Mol. Genet. Genomics 2004, 272, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, S.; Sethy, N.K.; Shokeen, B.; Bhatia, S. Development of chickpea est-ssr markers and analysis of allelic variation across related species. Theor. Appl. Genet. 2009, 118, 591–608. [Google Scholar] [CrossRef] [PubMed]
- Akash, M.W.; Myers, G.O. The development of faba bean expressed sequence tag-simple sequence repeats (est-ssrs) and their validity in diversity analysis. Plant Breed. 2012, 131, 522–530. [Google Scholar] [CrossRef]
- Liu, K.J.; Muse, S.V. Powermarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.W.; Kim, T.S.; Suresh, S.; Lee, S.Y.; Cho, G.T. Development of 65 novel polymorphic cdna-ssr markers in common vetch (vicia sativa subsp sativa) using next generation sequencing. Molecules 2013, 18, 8376–8392. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, E.J.; Padua, J.G.; Zucchi, M.I.; Vencovsky, R.; Vieira, M.L.C. Origin, evolution and genome distribution of microsatellites. Genet. Mol. Biol. 2006, 29, 294–307. [Google Scholar] [CrossRef]
- Riley, M. Functions of the gene-products of Escherichia coli. Microbiol. Rev. 1993, 57, 862–952. [Google Scholar] [PubMed]
- Peakall, R.; Smouse, P.E. Genalex 6.5: Genetic analysis in excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar]
- Sample Availability: Samples are available from authors Institute.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raveendar, S.; Lee, G.-A.; Jeon, Y.-A.; Lee, Y.J.; Lee, J.-R.; Cho, G.-T.; Cho, J.-H.; Park, J.-H.; Ma, K.-H.; Chung, J.-W. Cross-Amplification of Vicia sativa subsp. sativa Microsatellites across 22 Other Vicia Species. Molecules 2015, 20, 1543-1550. https://doi.org/10.3390/molecules20011543
Raveendar S, Lee G-A, Jeon Y-A, Lee YJ, Lee J-R, Cho G-T, Cho J-H, Park J-H, Ma K-H, Chung J-W. Cross-Amplification of Vicia sativa subsp. sativa Microsatellites across 22 Other Vicia Species. Molecules. 2015; 20(1):1543-1550. https://doi.org/10.3390/molecules20011543
Chicago/Turabian StyleRaveendar, Sebastin, Gi-An Lee, Young-Ah Jeon, Yun Jeong Lee, Jung-Ro Lee, Gyu-Taek Cho, Joon-Hyeong Cho, Jong-Hyun Park, Kyung-Ho Ma, and Jong-Wook Chung. 2015. "Cross-Amplification of Vicia sativa subsp. sativa Microsatellites across 22 Other Vicia Species" Molecules 20, no. 1: 1543-1550. https://doi.org/10.3390/molecules20011543
APA StyleRaveendar, S., Lee, G.-A., Jeon, Y.-A., Lee, Y. J., Lee, J.-R., Cho, G.-T., Cho, J.-H., Park, J.-H., Ma, K.-H., & Chung, J.-W. (2015). Cross-Amplification of Vicia sativa subsp. sativa Microsatellites across 22 Other Vicia Species. Molecules, 20(1), 1543-1550. https://doi.org/10.3390/molecules20011543