
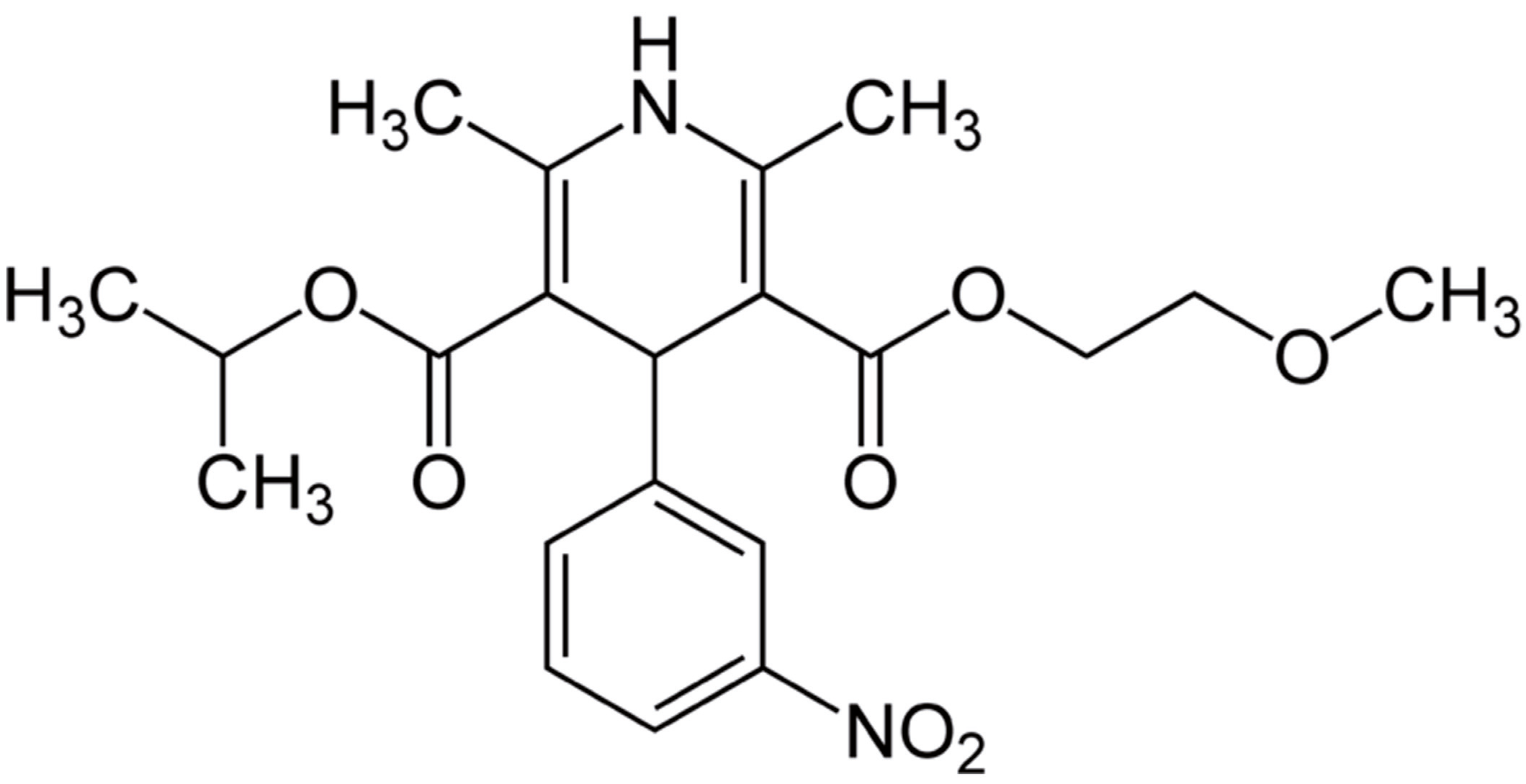
Neuroprotective and Neuroregenerative Effects of Nimodipine in a Model System of Neuronal Differentiation and Neurite Outgrowth

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
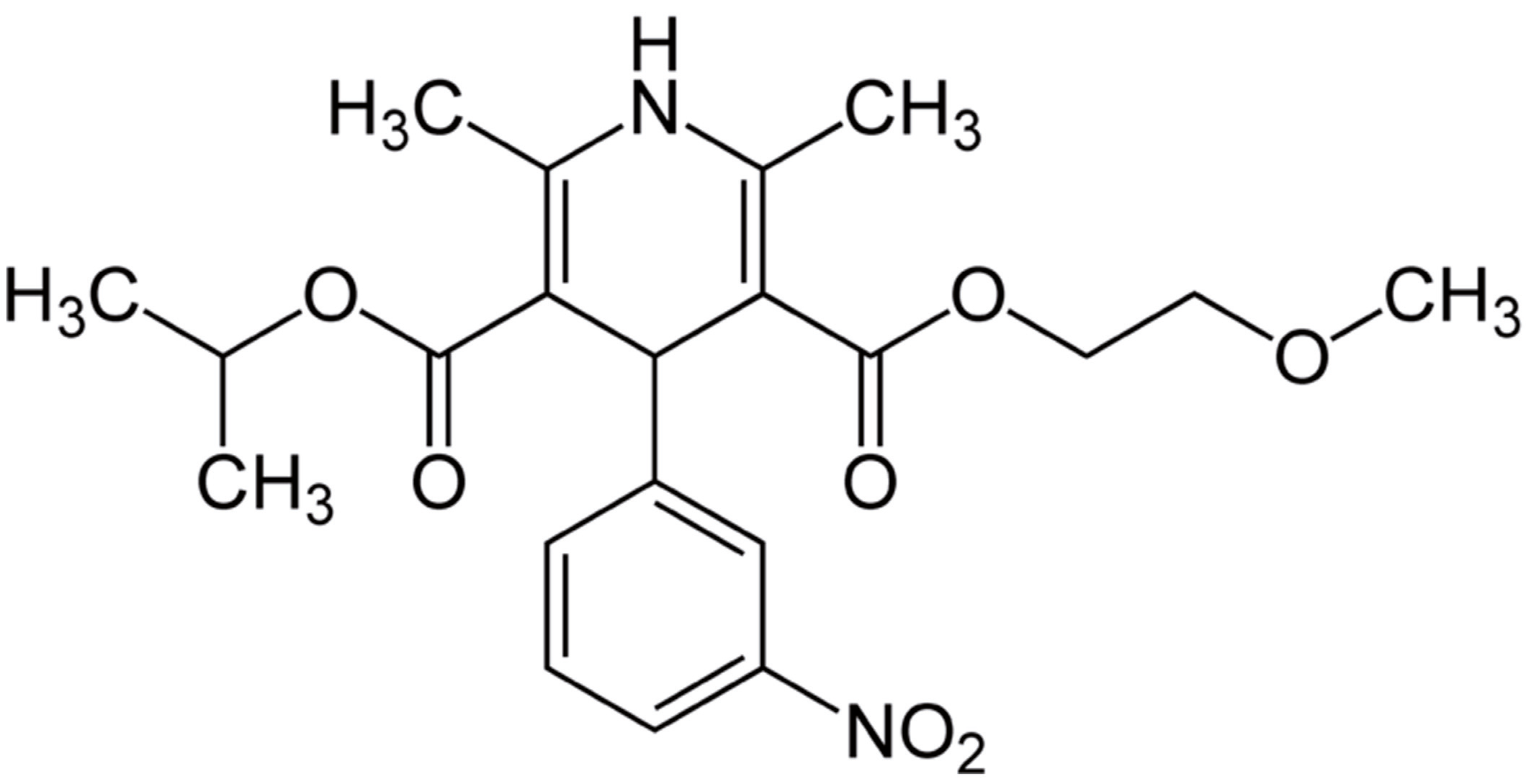
:1. Introduction
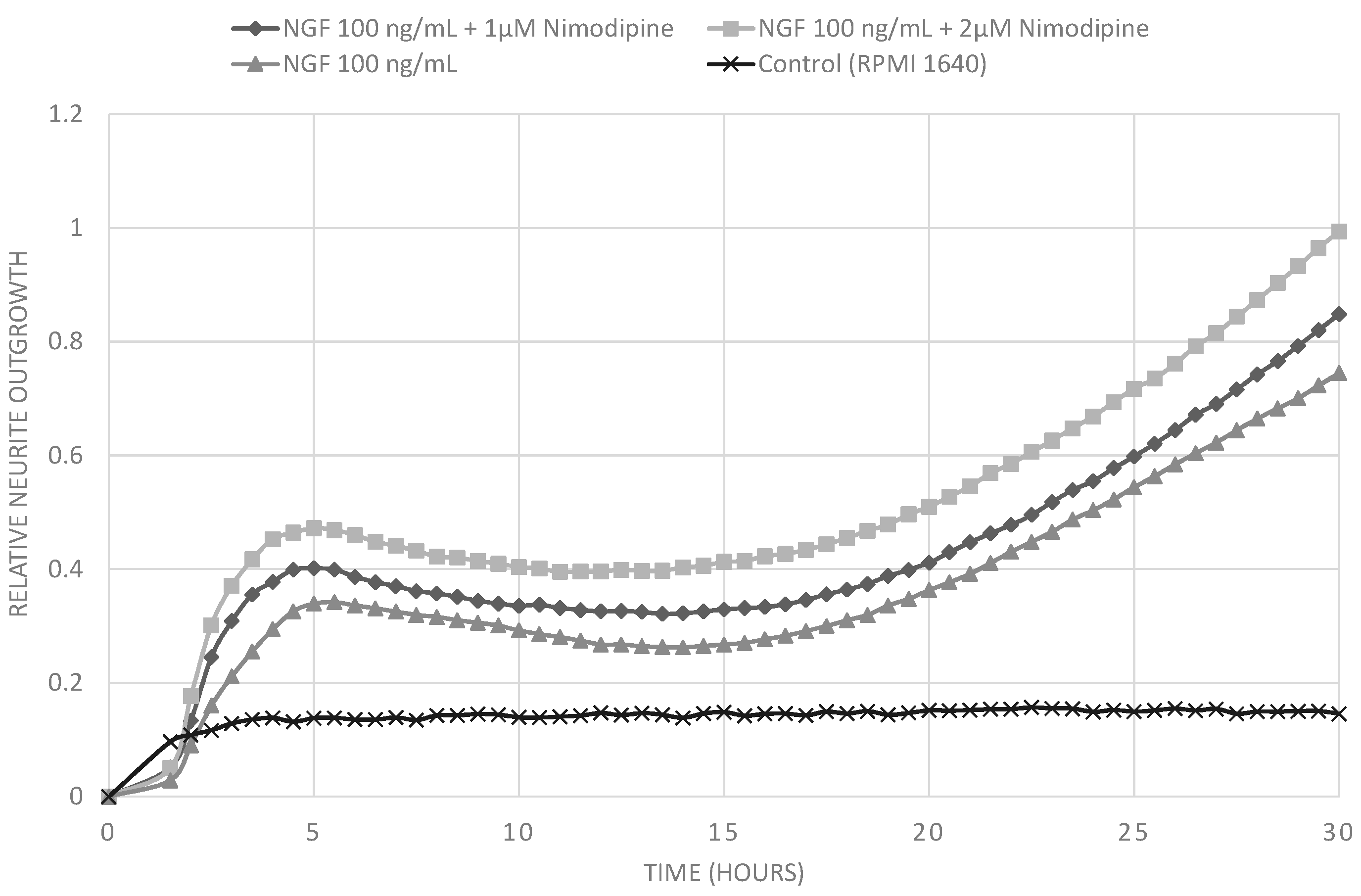
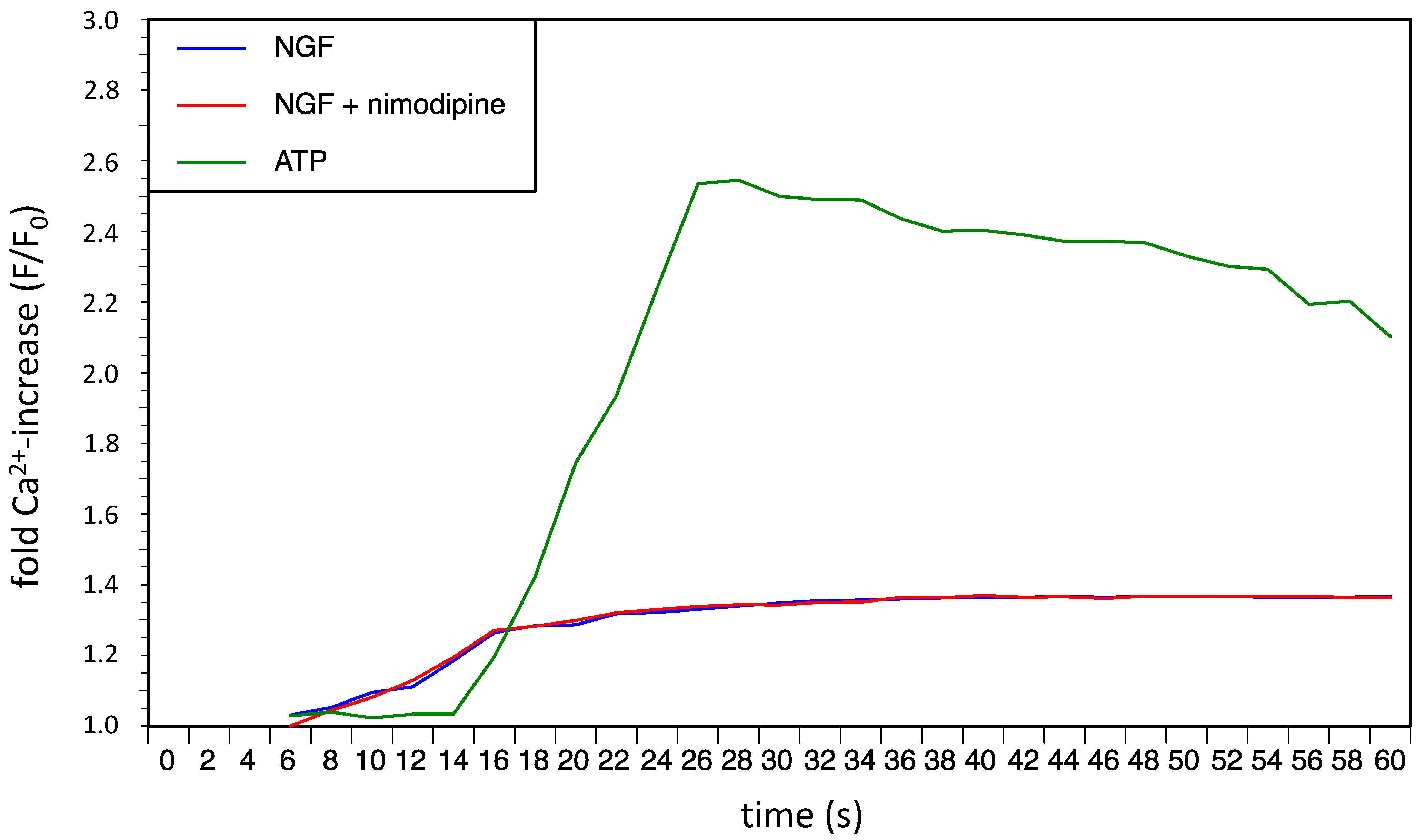
2. Results and Discussion
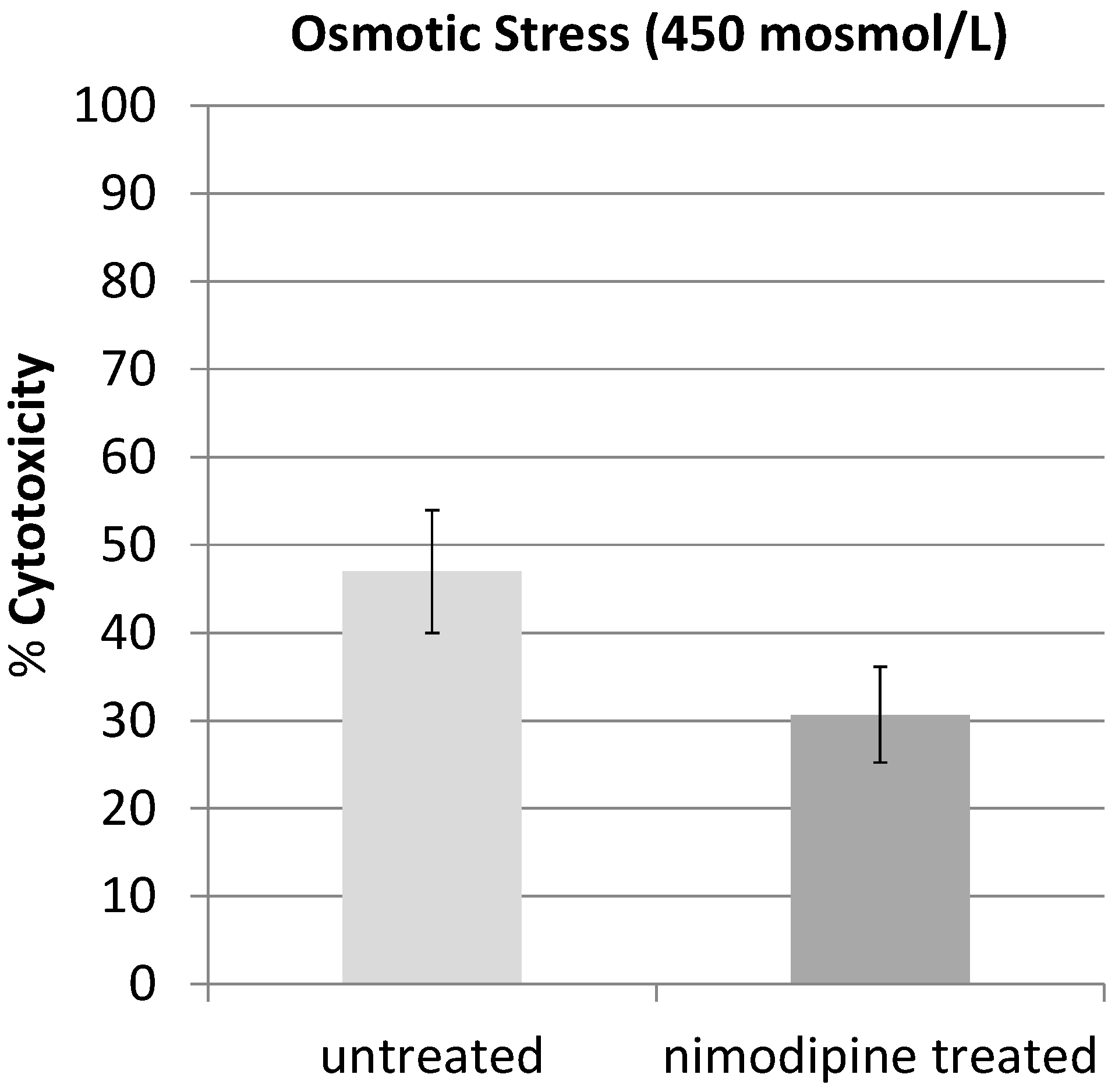
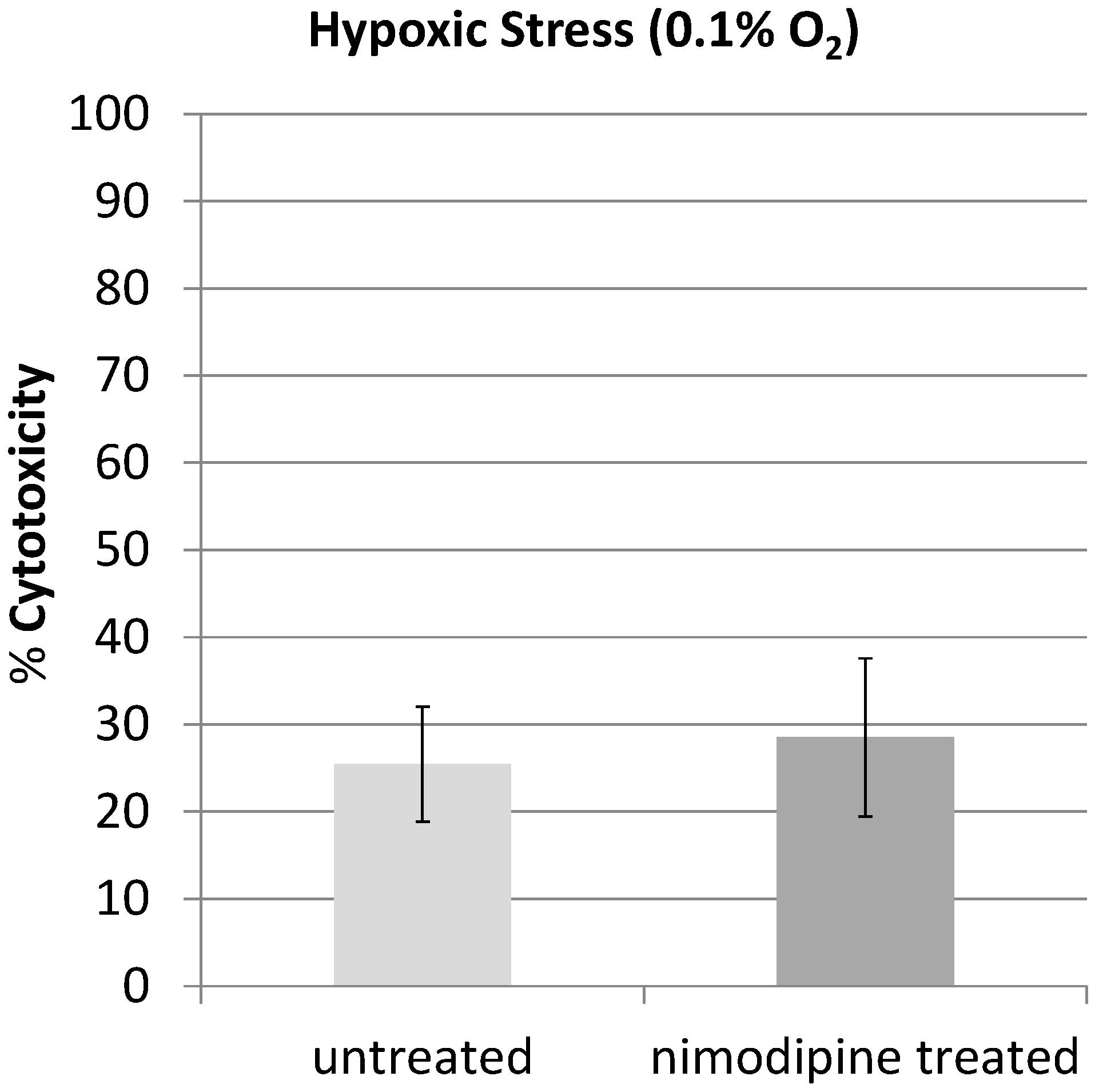
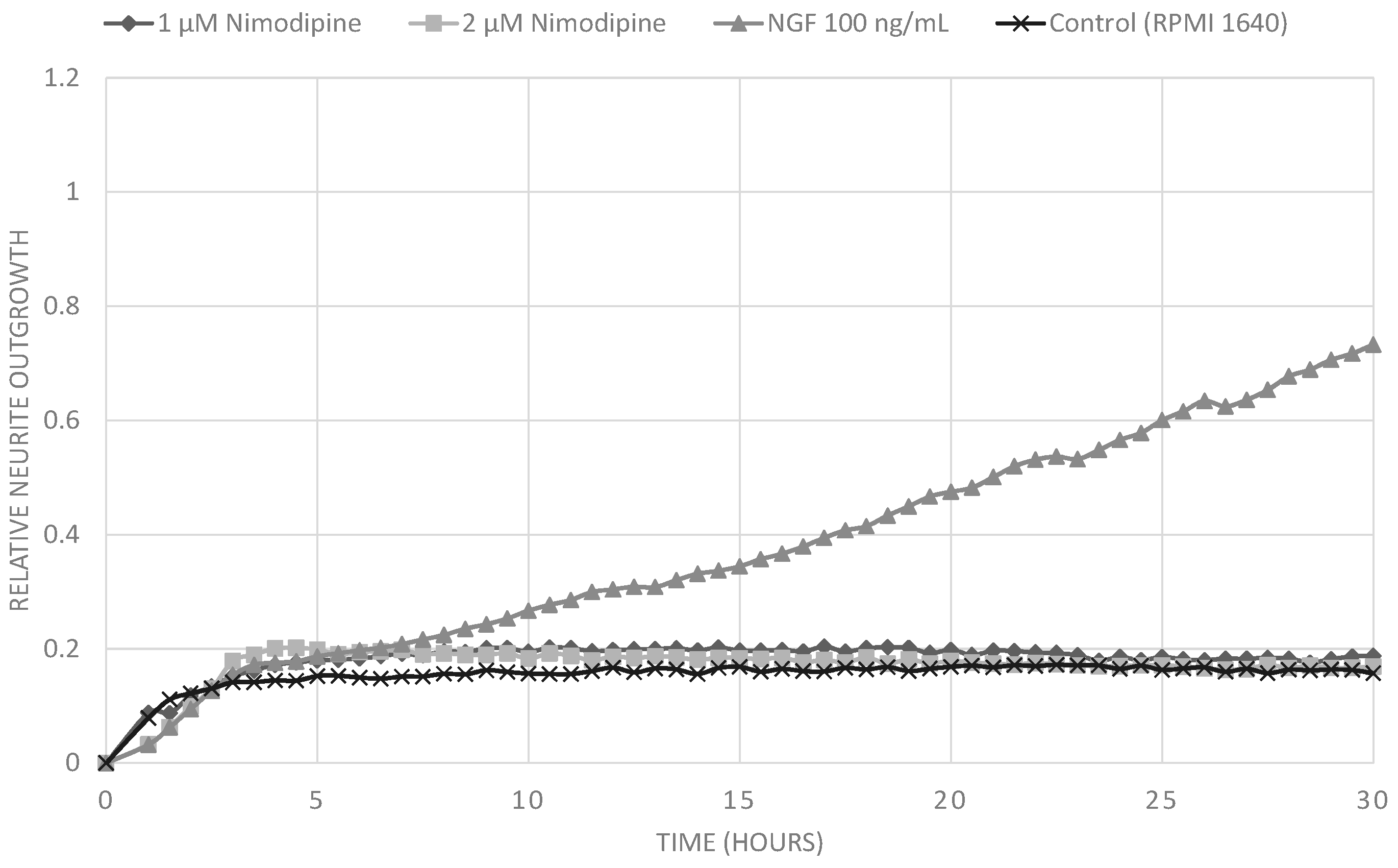
2.1. Results
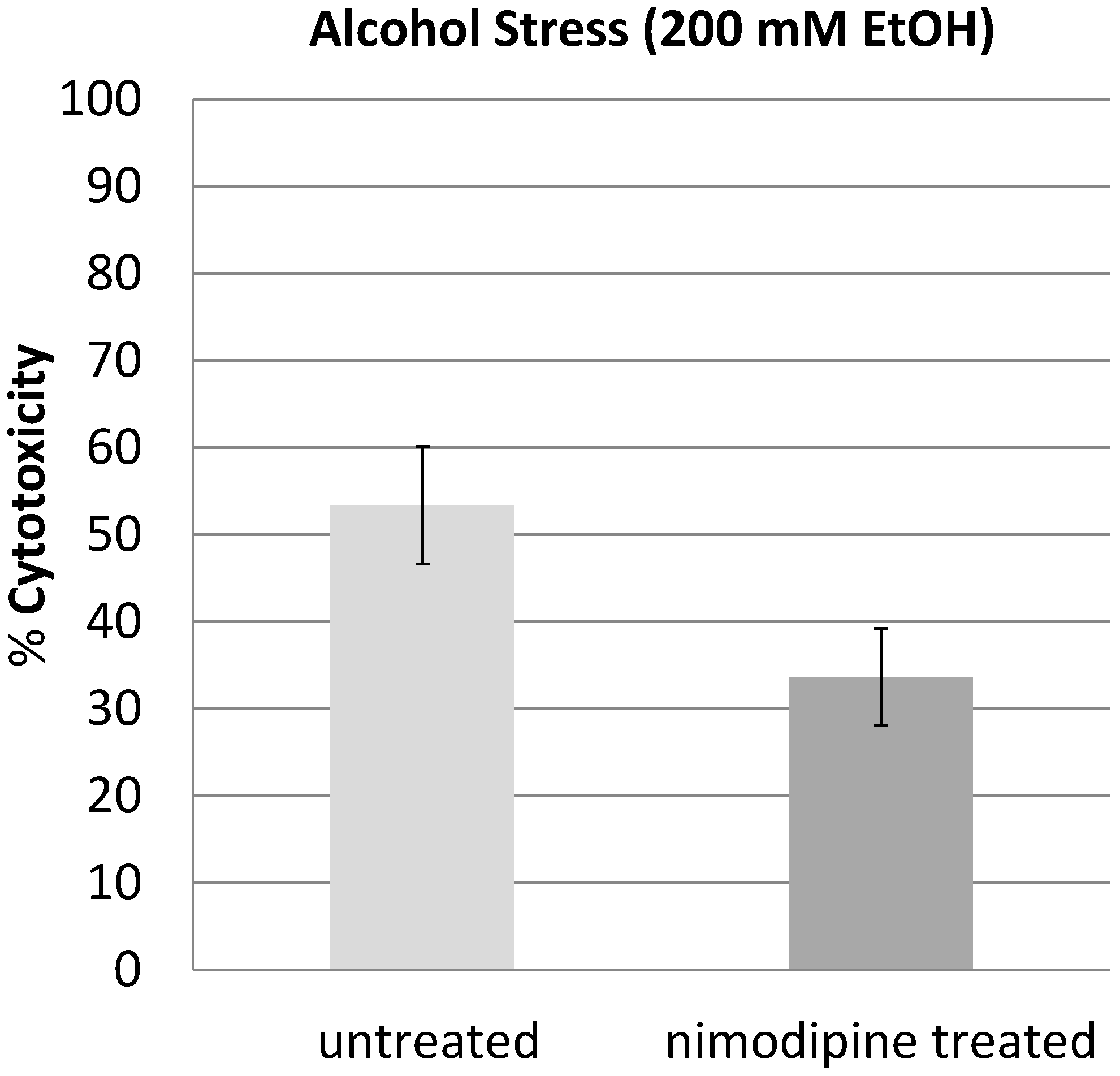
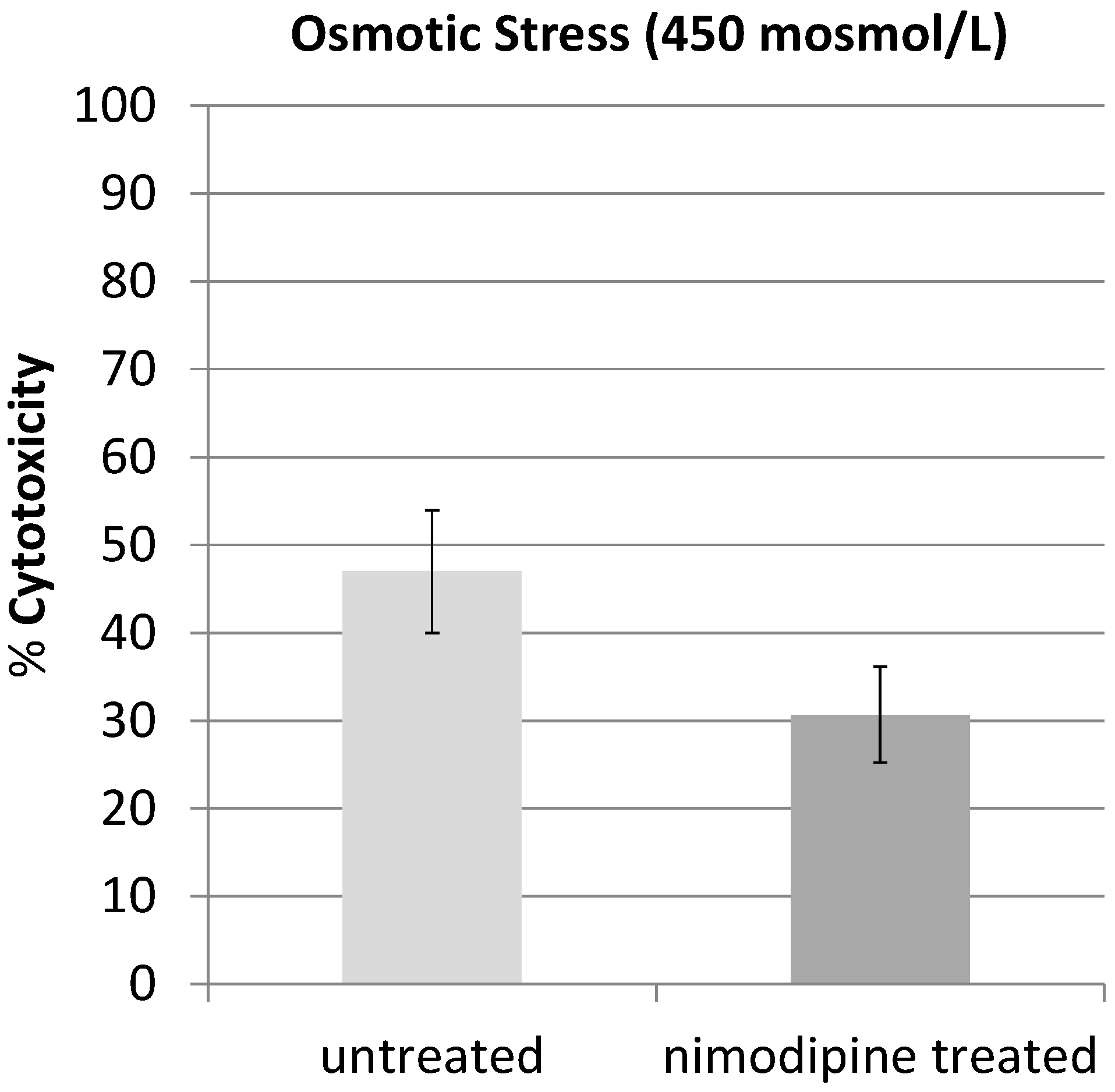
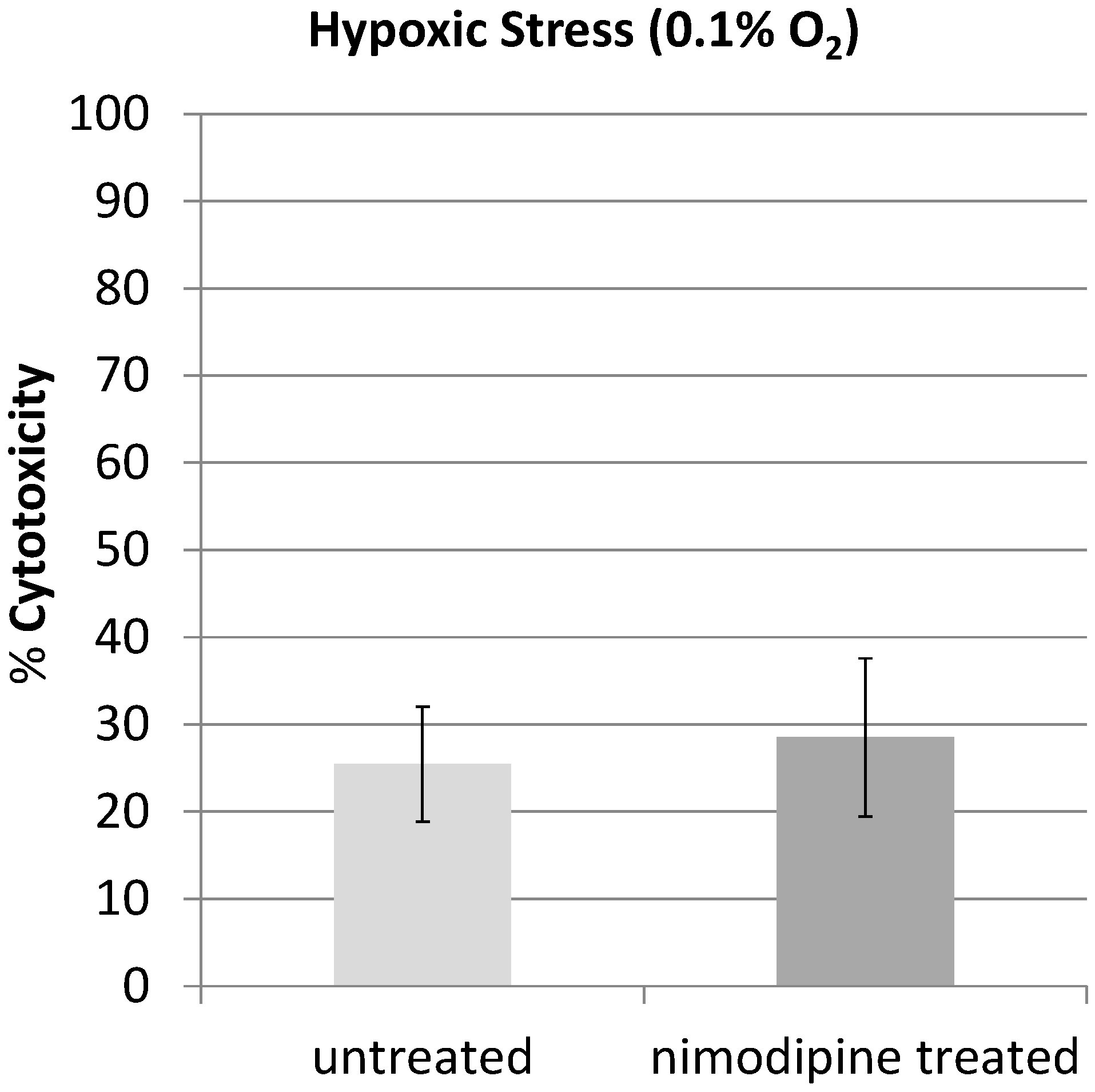
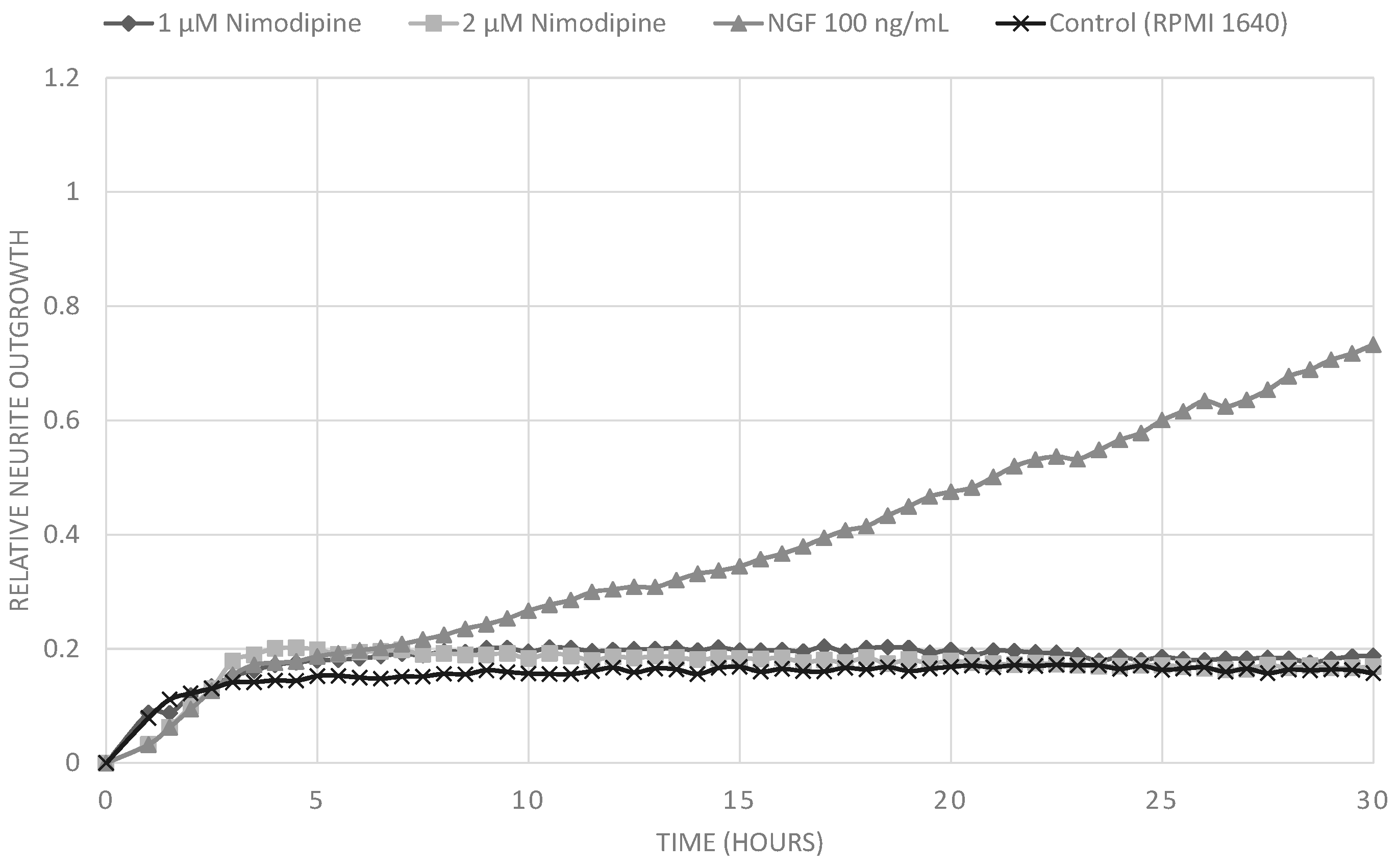
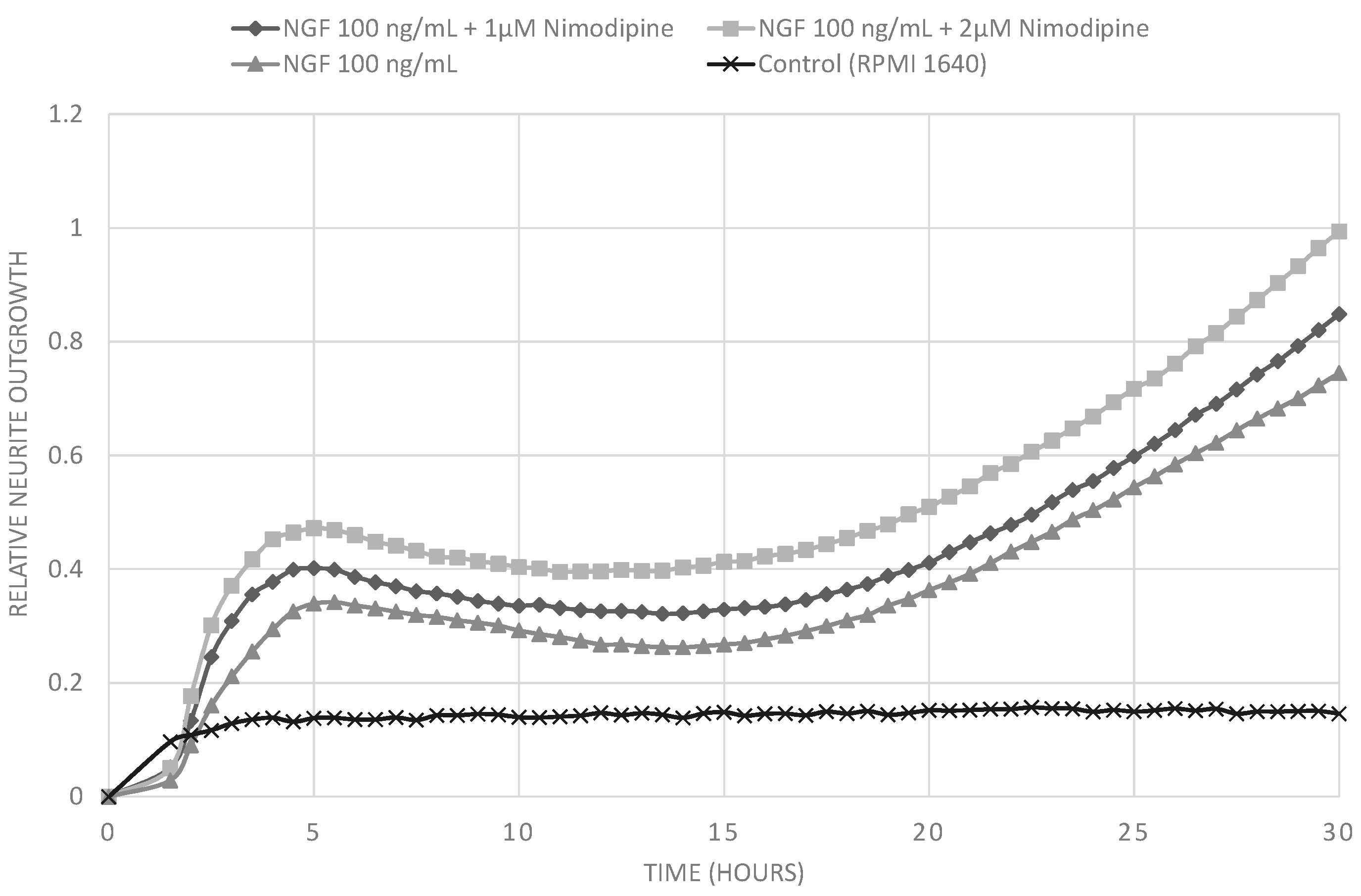
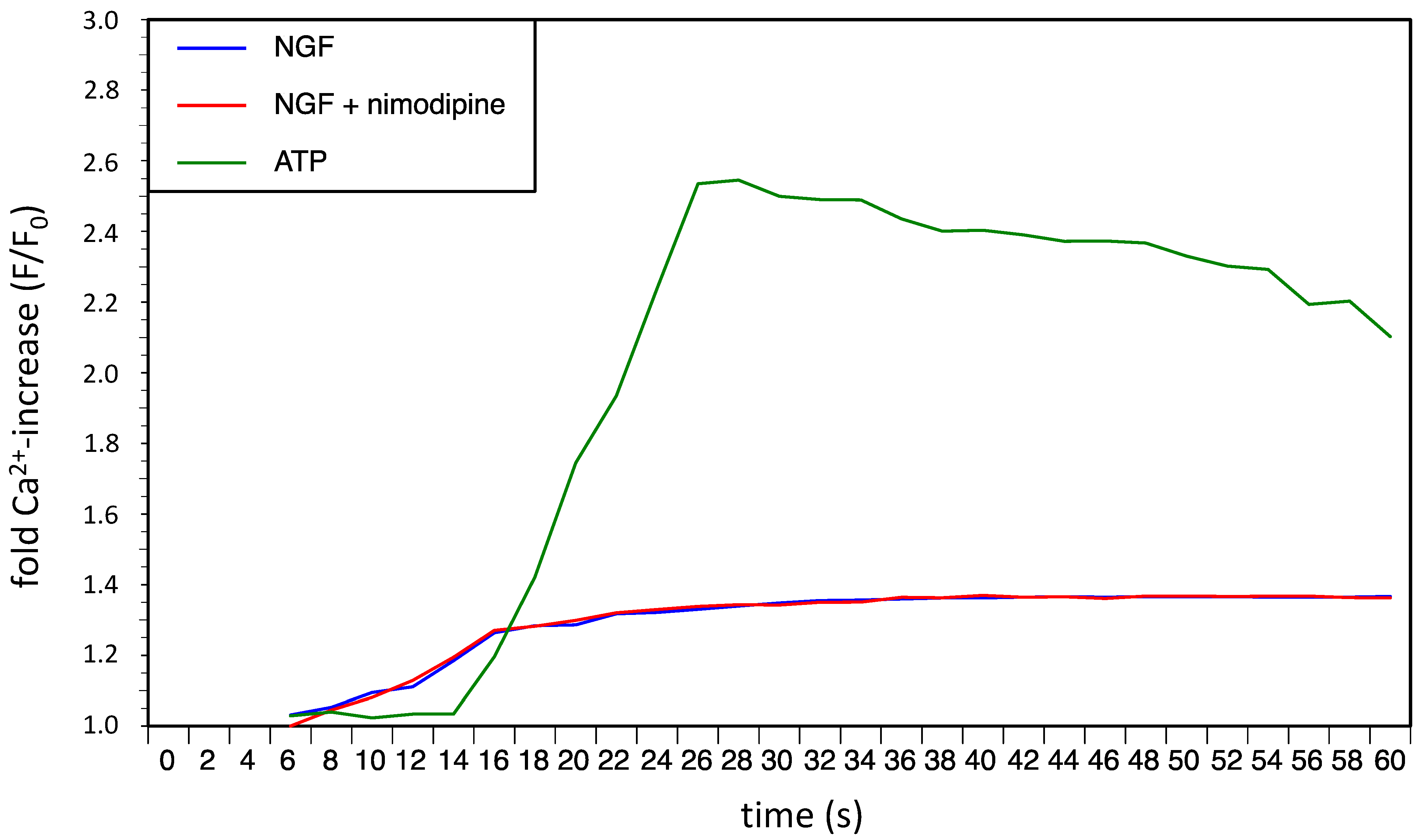
2.2. Discussion
3. Experimental Section
3.1. Cell Line
3.2. Cell Culture
3.3. Stress Assays
3.4. LDH-Assay
3.5. Neurite Outgrowth
3.6. Calcium-Assay
3.7. Cell Proliferation Assay
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kazda, S.; Towart, R. Nimodipine: A new calcium antagonistic drug with a preferential cerebrovascular action. Acta Neurochir. 1982, 63, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Scriabine, A.; van den Kerckhoff, W. Pharmacology of nimodipine. A review. Ann. N. Y. Acad. Sci. 1988, 522, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Dorhout Mees, S.M.; Rinkel, G.J.E.; Feigin, V.L.; Algra, A.; van den Bergh, W.M.; Vermeulen, M.; van Gijn, J. Calcium antagonists for aneurysmal subarachnoid haemorrhage. Cochrane Database Syst. Rev. 2007. [Google Scholar] [CrossRef]
- Toescu, E.C.; Verkhratsky, A. The importance of being subtle: Small changes in calcium homeostasis control cognitive decline in normal aging. Aging Cell 2007, 6, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Thibault, O.; Landfield, P.W. Increase in single l-type calcium channels in hippocampal neurons during aging. Science 1996, 272, 1017–1020. [Google Scholar] [CrossRef] [PubMed]
- López-Arrieta, J.M.; Birks, J. Nimodipine for primary degenerative, mixed and vascular dementia. Cochrane Database Syst. Rev. 2002. [Google Scholar] [CrossRef]
- Hydman, J.; Remahl, S.; Björck, G.; Svensson, M.; Mattsson, P. Nimodipine improves reinnervation and neuromuscular function after injury to the recurrent laryngeal nerve in the rat. Ann. Otol. Rhinol. Laryngol. 2007, 116, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, K.; Kumai, Y.; Sanuki, T.; Minoda, R.; Yumoto, E. The impact of nimodipine administration combined with nerve-muscle pedicle implantation on long-term denervated rat thyroarytenoid muscle. Laryngoscope 2013, 123, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Scheller, C.; Richter, H.-P.; Engelhardt, M.; Köenig, R.; Antoniadis, G. The influence of prophylactic vasoactive treatment on cochlear and facial nerve functions after vestibular schwannoma surgery: A prospective and open-label randomized pilot study. Neurosurgery 2007, 61, 92–97; discussion 97–98. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, B.; Romstöck, J.; Fahlbusch, R.; Buchfelder, M.; Strauss, C. Intraoperative brainstem auditory evoked potential pattern and perioperative vasoactive treatment for hearing preservation in vestibular schwannoma surgery. J. Neurol. Neurosurg. Psychiatry 2008, 79, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Scheller, C.; Wienke, A.; Wurm, F.; Simmermacher, S.; Rampp, S.; Prell, J.; Rachinger, J.; Scheller, K.; Koman, G.; Strauss, C.; et al. Neuroprotective Efficacy of Prophylactic Enteral and Parenteral Nimodipine Treatment in Vestibular Schwannoma Surgery: A Comparative Study. J. Neurol. Surg. Part Cent. Eur. Neurosurg. 2014, 75, 251–258. [Google Scholar]
- Scheller, K.; Scheller, C. Nimodipine promotes regeneration of peripheral facial nerve function after traumatic injury following maxillofacial surgery: An off label pilot-study. J. Cranio-Maxillofac. Surg. 2012, 40, 427–434. [Google Scholar] [CrossRef]
- Li, H.; Yang, X.; Shi, W.; Ma, Z.; Feng, G.; Wang, Q.; Shen, L.; Xie, C. Protective effects of nimodipine on cerebrovascular function in chronic alcoholic encephalopathy. Int. J. Mol. Med. 2014, 33, 201–208. [Google Scholar] [PubMed]
- Rice, D.C. Evidence for delayed neurotoxicity produced by methylmercury. Neurotoxicology 1996, 17, 583–596. [Google Scholar] [PubMed]
- Bailey, J.M.; Hutsell, B.A.; Newland, M.C. Dietary nimodipine delays the onset of methylmercury neurotoxicity in mice. Neurotoxicology 2013, 37, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Greene, L.A.; Tischler, A.S. Establishment of a noradrenergic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proc. Natl. Acad. Sci. USA 1976, 73, 2424–2428. [Google Scholar] [CrossRef] [PubMed]
- Shafer, T.J.; Atchison, W.D. Transmitter, ion channel and receptor properties of pheochromocytoma (PC12) cells: A model for neurotoxicological studies. Neurotoxicology 1991, 12, 473–492. [Google Scholar] [PubMed]
- Scheller, C.; Wienke, A.; Wurm, F.; Vogel, A.-S.; Simmermacher, S.; Prell, J.; Rachinger, J.; Koman, G.; Strauss, C.; Scheller, K. Enteral or parenteral nimodipine treatment: A comparative pharmacokinetic study. J. Neurol. Surg. Part Cent. Eur. Neurosurg. 2014, 75, 84–90. [Google Scholar]
- Mesis, R.G.; Wang, H.; Lombard, F.W.; Yates, R.; Vitek, M.P.; Borel, C.O.; Warner, D.S.; Laskowitz, D.T. Dissociation between vasospasm and functional improvement in a murine model of subarachnoid hemorrhage. Neurosurg. Focus 2006, 21, E4. [Google Scholar] [CrossRef] [PubMed]
- Lecht, S.; Rotfeld, E.; Arien-Zakay, H.; Tabakman, R.; Matzner, H.; Yaka, R.; Lelkes, P.I.; Lazarovici, P. Neuroprotective effects of nimodipine and nifedipine in the NGF-differentiated PC12 cells exposed to oxygen-glucose deprivation or trophic withdrawal. Int. J. Dev. Neurosci. 2012, 30, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Streit, J.; Lux, H.D. Voltage dependent calcium currents in PC12 growth cones and cells during NGF-induced cell growth. Pflüg. Arch. Eur. J. Physiol. 1987, 408, 634–641. [Google Scholar] [CrossRef]
- Bouron, A.; Becker, C.; Porzig, H. Functional expression of voltage-gated Na+ and Ca2+ channels during neuronal differentiation of PC12 cells with nerve growth factor or forskolin. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1999, 359, 370–377. [Google Scholar] [CrossRef]
- Ju, L.; Zhang, X.; Zhang, T.; Zheng, J. Research on recovery function of two drugs combination on rat sciatic nerve injury regeneration model. Pak. J. Pharm. Sci. 2014, 27, 1695–1698. [Google Scholar] [PubMed]
- Wroblewski, F.; Ladue, J.S. Lactic dehydrogenase activity in blood. Proc. Soc. Exp. Biol. Med. 1955, 90, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Pollscheit, J.; Glaubitz, N.; Haller, H.; Horstkorte, R.; Bork, K. Phosphorylation of serine 774 of the neural cell adhesion molecule is necessary for cyclic adenosine monophosphate response element binding protein activation and neurite outgrowth. J. Neurosci. Res. 2012, 90, 1577–1582. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are not available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bork, K.; Wurm, F.; Haller, H.; Strauss, C.; Scheller, C.; Gnanapragassam, V.S.; Horstkorte, R. Neuroprotective and Neuroregenerative Effects of Nimodipine in a Model System of Neuronal Differentiation and Neurite Outgrowth. Molecules 2015, 20, 1003-1013. https://doi.org/10.3390/molecules20011003
Bork K, Wurm F, Haller H, Strauss C, Scheller C, Gnanapragassam VS, Horstkorte R. Neuroprotective and Neuroregenerative Effects of Nimodipine in a Model System of Neuronal Differentiation and Neurite Outgrowth. Molecules. 2015; 20(1):1003-1013. https://doi.org/10.3390/molecules20011003
Chicago/Turabian StyleBork, Kaya, Franziska Wurm, Hannes Haller, Christian Strauss, Christian Scheller, Vinayaga S. Gnanapragassam, and Rüdiger Horstkorte. 2015. "Neuroprotective and Neuroregenerative Effects of Nimodipine in a Model System of Neuronal Differentiation and Neurite Outgrowth" Molecules 20, no. 1: 1003-1013. https://doi.org/10.3390/molecules20011003
APA StyleBork, K., Wurm, F., Haller, H., Strauss, C., Scheller, C., Gnanapragassam, V. S., & Horstkorte, R. (2015). Neuroprotective and Neuroregenerative Effects of Nimodipine in a Model System of Neuronal Differentiation and Neurite Outgrowth. Molecules, 20(1), 1003-1013. https://doi.org/10.3390/molecules20011003