Susceptibility of Opportunistic Burkholderia glumae to Copper Surfaces Following Wet or Dry Surface Contact


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
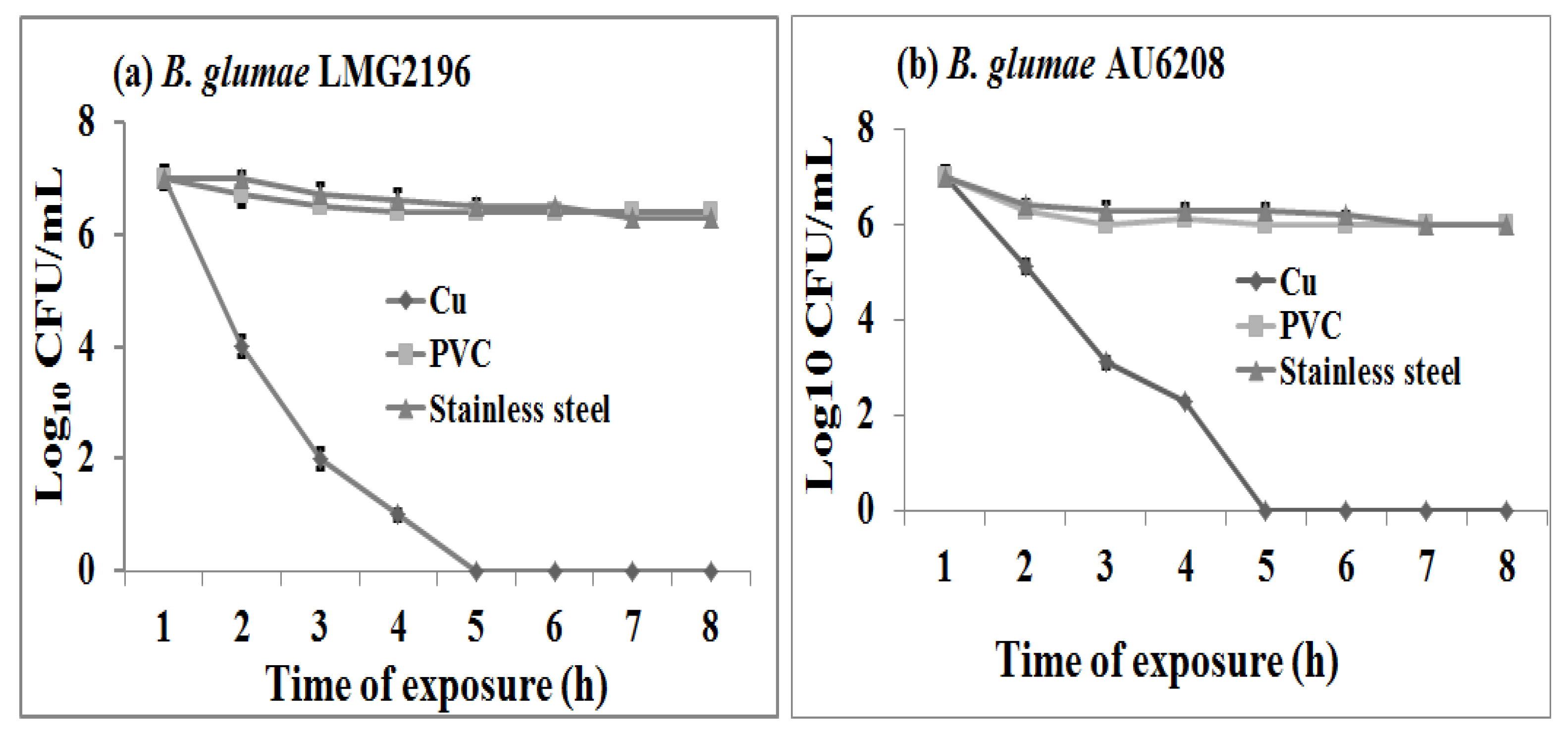
2.1. Susceptibility of B. glumae to Copper Surfaces
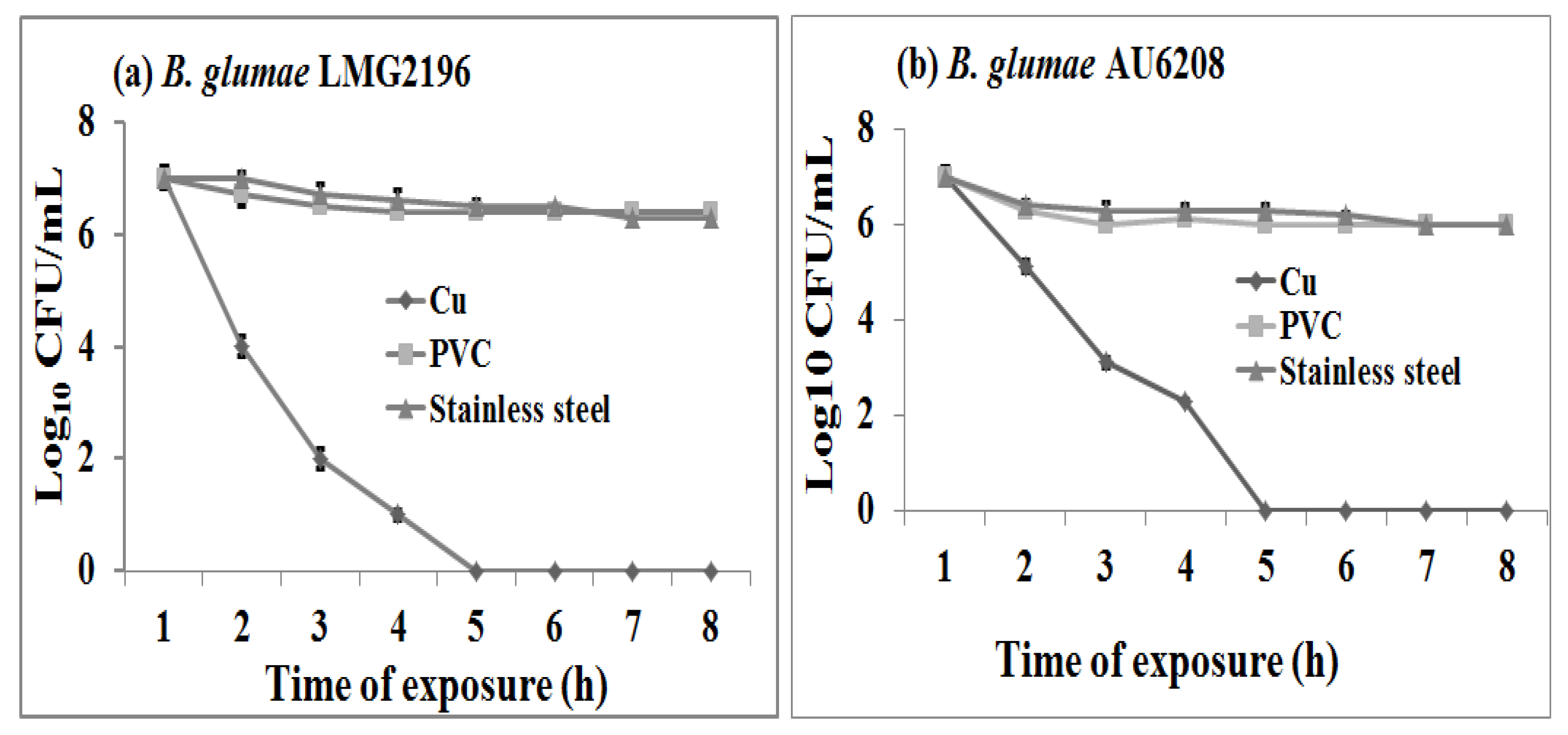
2.2. Acute Toxicity of Copper to B. glumae
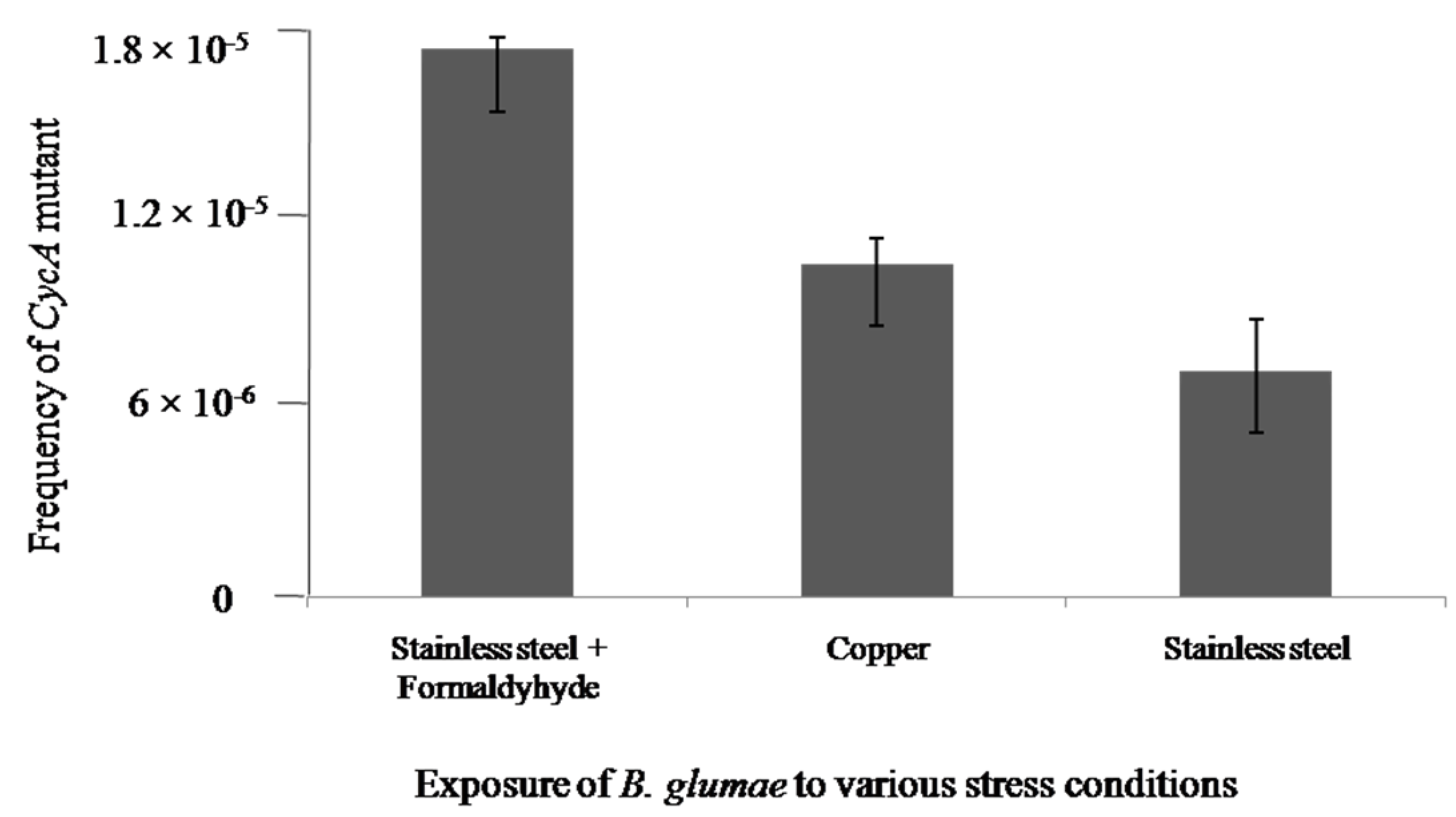
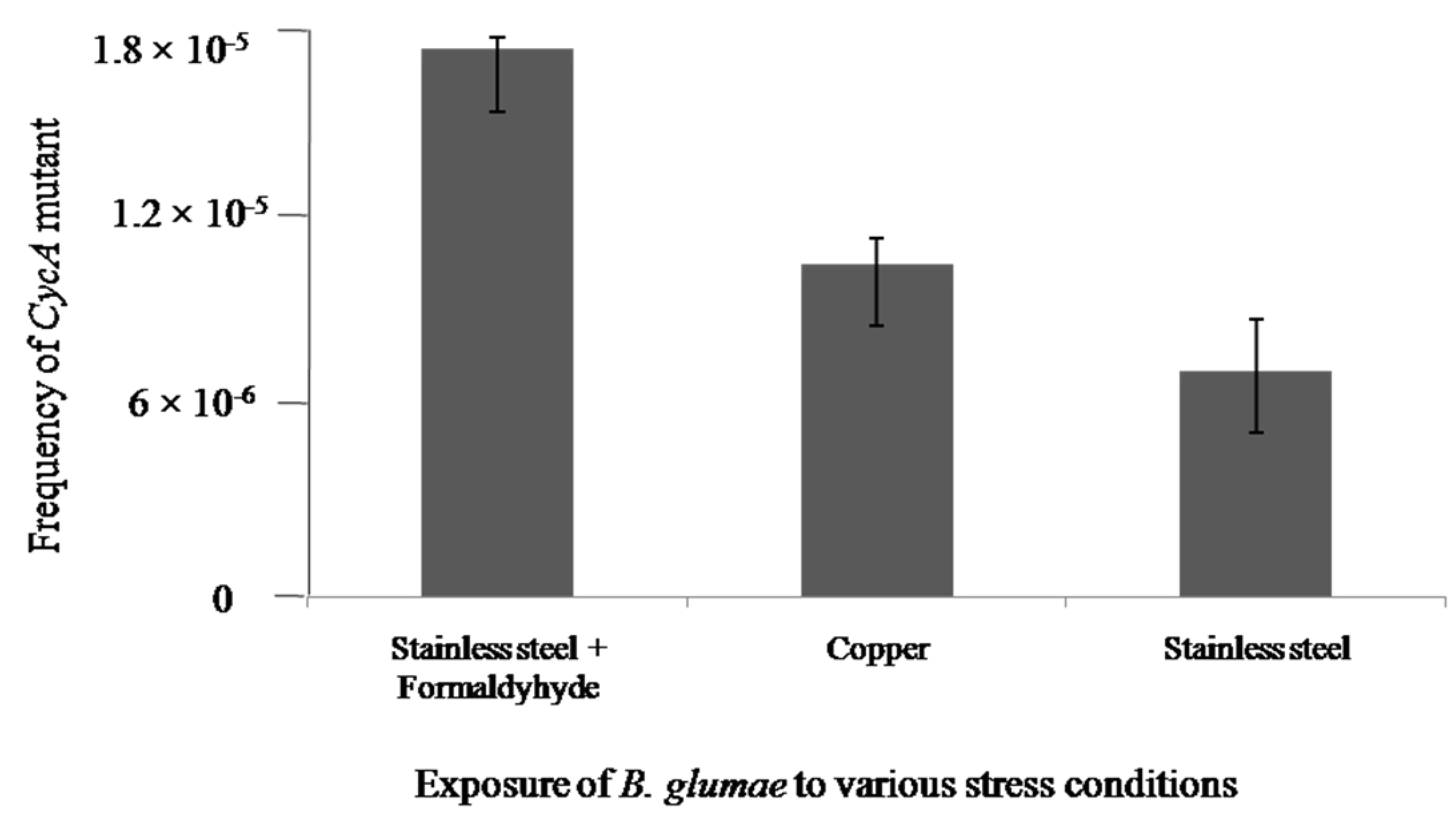
2.3. DNA Damage

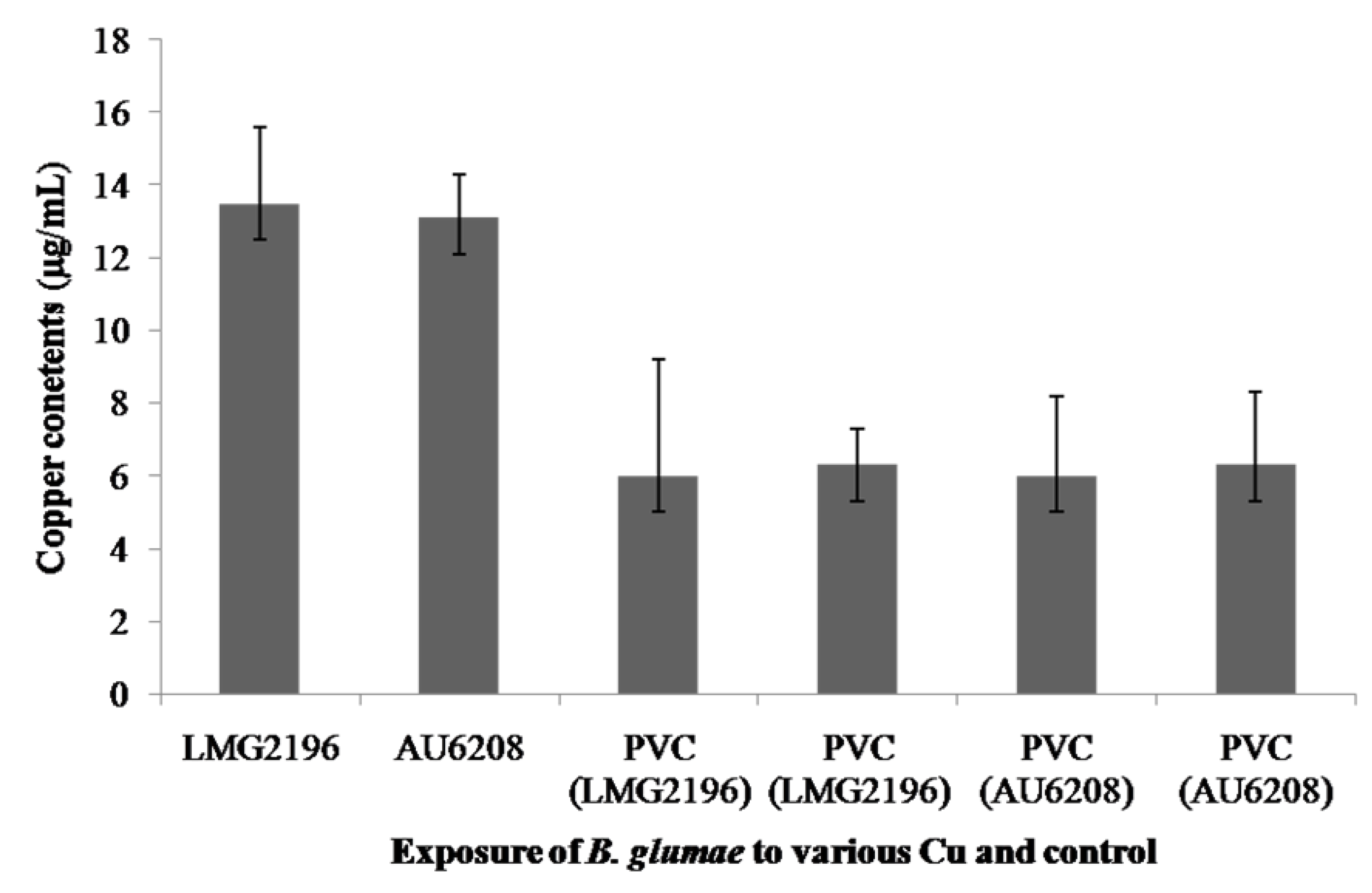
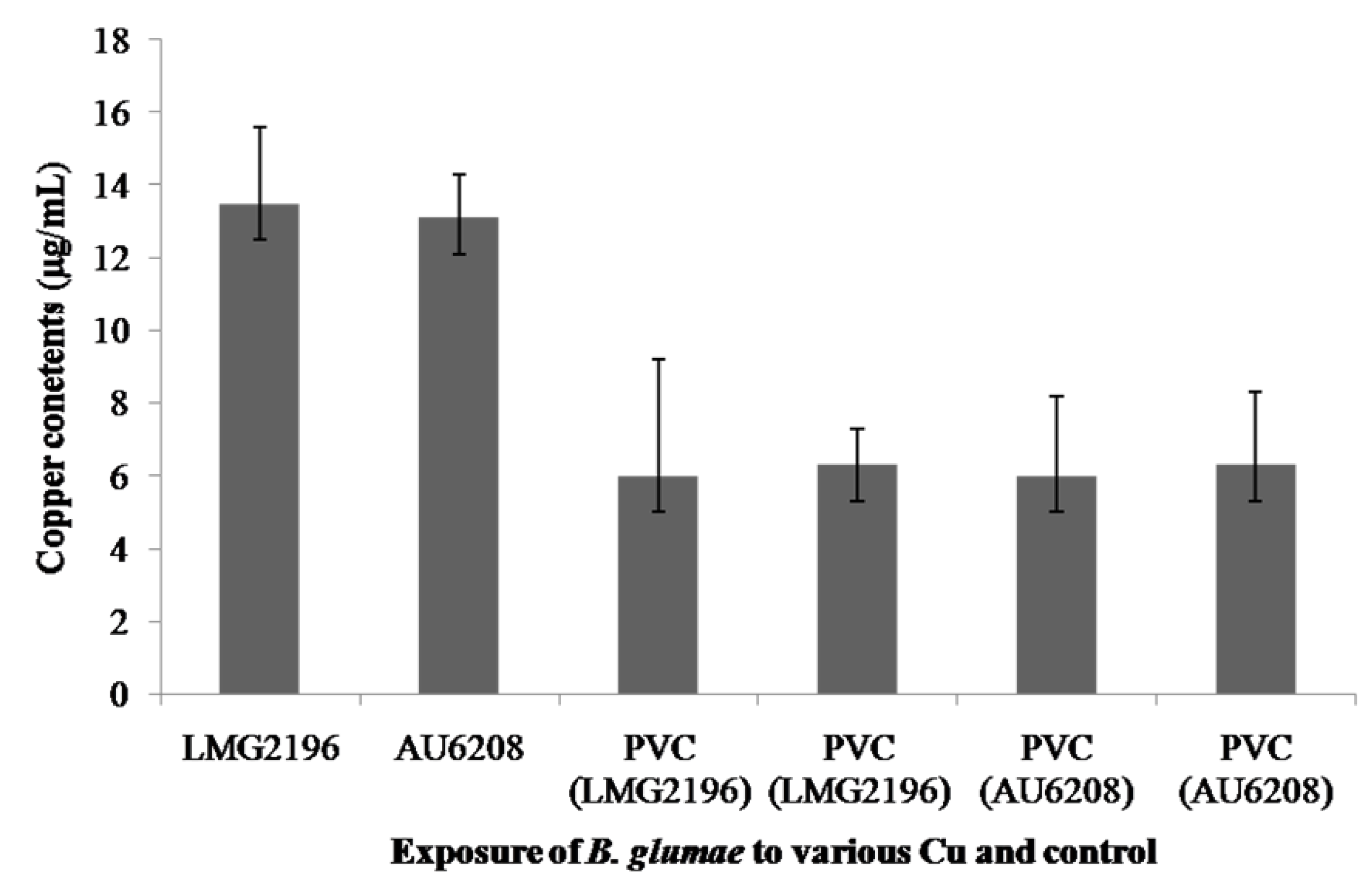
2.4. Accumulation of Copper Ions in Cells
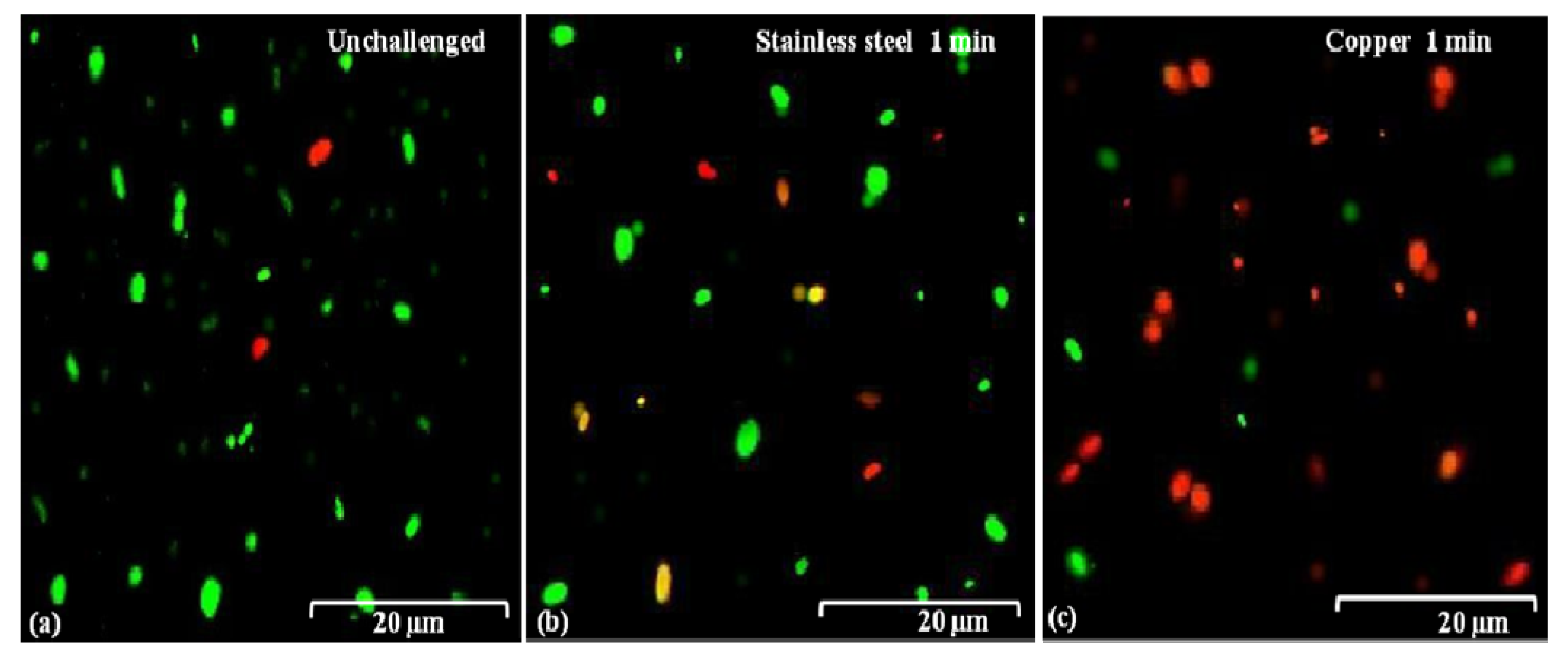
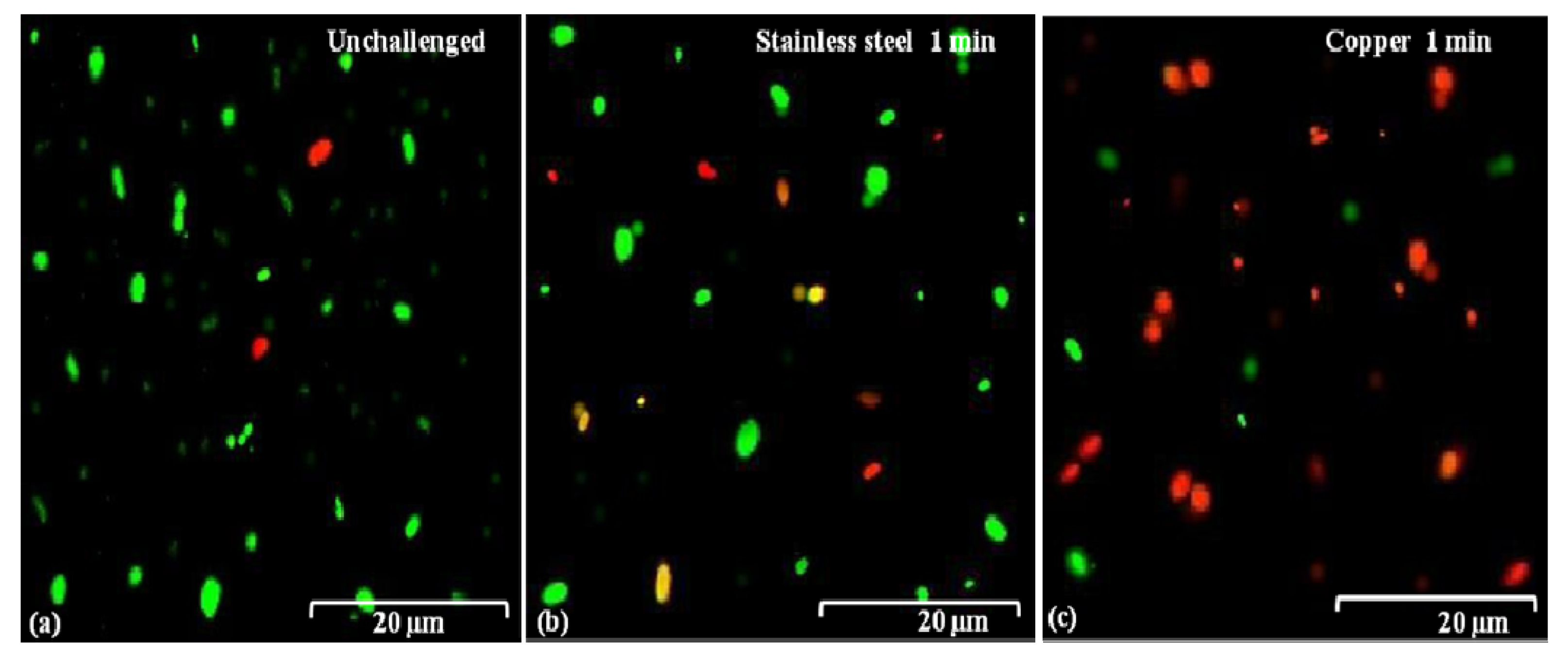
2.5. Damage in Cell Membranes
3. Experimental
3.1. Bacterial Strains and Culture Conditions
3.2. Metallic Copper Surfaces
3.3. Viability Assays
3.4. Mutagenicity Assay
3.5. Single Cell Gel Electrophoresis (SCGE)
3.6. ICP-MS Analysis
3.7. Membrane Damage
3.8. Statistical Analyses
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mahenthiralingam, E.; Urban, T.A.; Goldberg, J.B. The multifarious, multireplicon Burkholderia cepacia complex. Nat. Rev. Microbiol. 2005, 3, 144–156. [Google Scholar] [CrossRef]
- Coenye, T.; Vandamme, T. Diversity and significance of Burkholderia species occupying diverse ecological niches. Environ. Microbial. 2003, 5, 719–729. [Google Scholar] [CrossRef]
- Kim, S.; Park, J.; Kim, J.H.; Lee, J.; Bang, B.; Hwang, I.; Seo, Y.S. RNAseq-based transcriptome analysis of Burkholderia glumae quorum sensing. Plant Pathol. J. 2013, 29, 249–259. [Google Scholar] [CrossRef]
- Kato, T.; Morohoshi, T.; Tsushima, S.; Ikeda, T. Phenotypic characterization of colony morphological mutants of Burkholderia glumae that emerged during subculture. J. Gen. Plant Pathol. 2013, 79, 249–259. [Google Scholar] [CrossRef]
- Karki, H.S.; Shrestha, B.K.; Han, J.W.; Groth, D.E.; Barphagha, I.K.; Rush, M.C.; Melanson, R.A.; Kim, B.S.; Ham, J.H. Diversities in virulence, antifungal activity, pigmentation and DNA fingerprint among strains of Burkholderia glumae. PLoS One 2012, 7, e45376. [Google Scholar] [CrossRef]
- Ham, J.H.; Melanson, R.A.; Rush, M.C. Burkholderia glumae: Next major pathogen of rice? Mol. Plant Pathol. 2011, 12, 329–339. [Google Scholar] [CrossRef]
- Weinberg, J.B.; Alexander, B.D.; Majure, J.M.; Williams, L.W.; Kim, J.Y.; Vandamme, P.; LiPuma, J.J. Burkholderia glumae infection in an infant with chronic granulomatous disease. J. Clin. Microbiol. 2007, 45, 662–665. [Google Scholar] [CrossRef]
- Luo, J.; Xie, G.; Li, B.; Xu, L.H. First report of Burkholderia glumae isolated from symptomless rice seeds in china. Plant Dis. 2007, 91, 1363. [Google Scholar]
- Kirzinger, M.W.B.; Nadarasah, G.; Stavrinides, J. Insights into cross-kingdom plant pathogenic bacteria. Genes 2011, 2, 980–997. [Google Scholar] [CrossRef]
- Azanza, M.P.V.; Ortega, M.P. Microbiology of day-old chicks: A Philippine street food. Food Control 2004, 15, 245–252. [Google Scholar] [CrossRef]
- Airey, P.; Verran, J. Potential use of copper as a hygienic surface; problems associated with cumulative soiling and cleaning. J. Hosp. Infect. 2007, 67, 272–278. [Google Scholar]
- Hengge-Aronis, R. Signal transduction and regulatory mechanisms involved in control of the σS (RpoS) subunit of RNA polymerase. Microbiol. Mol. Biol. Rev. 2002, 66, 373–395. [Google Scholar] [CrossRef]
- Espirito Santo, C.; Lam, E.W.; Elowsky, C.G.; Quaranta, D.; Domaille, D.W.; Chang, C.J.; Grass, G. Bacterial killing by dry metallic copper surfaces. Appl. Environ. Microbiol. 2011, 77, 794–800. [Google Scholar] [CrossRef]
- Dollwet, H.H.; Sorenson, J.R. Historic uses of copper compounds in medicine. Trace Elem. Biol. 1985, 2, 80–87. [Google Scholar]
- Faundez, G.; Troncoso, M.; Navarrete, P.; Figueroa, G. Antimicrobial activity of copper surfaces against suspensions of Salmonella enterica and Campylobacter jejuni. BMC Microbiol. 2004, 4, 19. [Google Scholar] [CrossRef]
- Elguindi, J.; Wagner, J.; Rensing, C. Genes involved in copper resistance influence survival of Pseudomonas aeruginosa on copper surfaces. J. Appl. Microbial. 2009, 106, 1448–1455. [Google Scholar] [CrossRef]
- Noyce, J.O.; Michels, H.; Keevil, C.W. Potential use of copper surfaces to reduce survival of epidemic methicillin-resistant Staphylococcus aureus in the healthcare environment. J. Hosp. Infect. 2006, 63, 289–297. [Google Scholar] [CrossRef]
- Borkow, G.; Gabbay, J. Copper as a biocidal tool. Curr. Med. Chem. 2005, 12, 2163–2175. [Google Scholar] [CrossRef]
- Grass, G.; Rensing, C.; Solioz, M. Metallic copper as an antimicrobial surface. Appl. Environ. Microbiol. 2011, 77, 1541–1547. [Google Scholar] [CrossRef]
- Ohsumi, Y.; Kitamoto, K.; Anraku, Y. Changes induced in the permeability barrier of the yeast plasma membrane by cupric ion. J. Bacteriol. 1998, 170, 2676–2682. [Google Scholar]
- Espirito Santo, C.; Morais, P.V.; Grass, G. Isolation and characterization of bacteria resistant to metallic copper surfaces. Appl. Environ. Microbiol. 2010, 76, 1341–1348. [Google Scholar] [CrossRef]
- Mehtar, S.; Wiid, I.; Todorov, S.D. The antimicrobial activity of copper and copper alloys against nosocomial pathogens and Mycobacterium tuberculosis isolated from healthcare facilities in the Western Cape, an in vitro study. J. Hosp. Infect. 2008, 68, 45–51. [Google Scholar] [CrossRef]
- Ibrahim, M.; Wang, F.; Lou, M.; Xie, G.; Li, B.; Bo, Z.; Zhang, G.; Liu, H.; Wareth, A. Copper as an antibacterial agent for human pathogenic multidrug resistant Burkholderia cepacia complex bacteria. J. Biosc. Bioengin. 2011, 112, 570–576. [Google Scholar] [CrossRef]
- Tian, W.X.; Yu, S.; Ibrahim, M.; Almonaofy, A.W.; Liu, H.; Qiu, H.; Zhu, B.; Li, B.; Xie, G.L. Copper as an antimicrobial agent against opportunistic pathogenic and multidrug resistant Enterobacter bacteria. J. Microbiol. 2012, 50, 586–593. [Google Scholar] [CrossRef]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells for DNA damage in individual cells. Exp. Cell Res. 1998, 175, 184–191. [Google Scholar]
- Sample Availability: Samples of the copper are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, Z.; Ibrahim, M.; Yang, C.; Fang, Y.; Annam, H.; Li, B.; Wang, Y.; Xie, G.-L.; Sun, G. Susceptibility of Opportunistic Burkholderia glumae to Copper Surfaces Following Wet or Dry Surface Contact. Molecules 2014, 19, 9975-9985. https://doi.org/10.3390/molecules19079975
Cui Z, Ibrahim M, Yang C, Fang Y, Annam H, Li B, Wang Y, Xie G-L, Sun G. Susceptibility of Opportunistic Burkholderia glumae to Copper Surfaces Following Wet or Dry Surface Contact. Molecules. 2014; 19(7):9975-9985. https://doi.org/10.3390/molecules19079975
Chicago/Turabian StyleCui, Zhouqi, Muhammad Ibrahim, Chunlan Yang, Yuan Fang, Hussain Annam, Bin Li, Yanli Wang, Guan-Lin Xie, and Guochang Sun. 2014. "Susceptibility of Opportunistic Burkholderia glumae to Copper Surfaces Following Wet or Dry Surface Contact" Molecules 19, no. 7: 9975-9985. https://doi.org/10.3390/molecules19079975
APA StyleCui, Z., Ibrahim, M., Yang, C., Fang, Y., Annam, H., Li, B., Wang, Y., Xie, G.-L., & Sun, G. (2014). Susceptibility of Opportunistic Burkholderia glumae to Copper Surfaces Following Wet or Dry Surface Contact. Molecules, 19(7), 9975-9985. https://doi.org/10.3390/molecules19079975