Goat Milk Fat Naturally Enriched with Conjugated Linoleic Acid Increased Lipoproteins and Reduced Triacylglycerol in Rats

Abstract
:1. Introduction
2. Results and Discussion
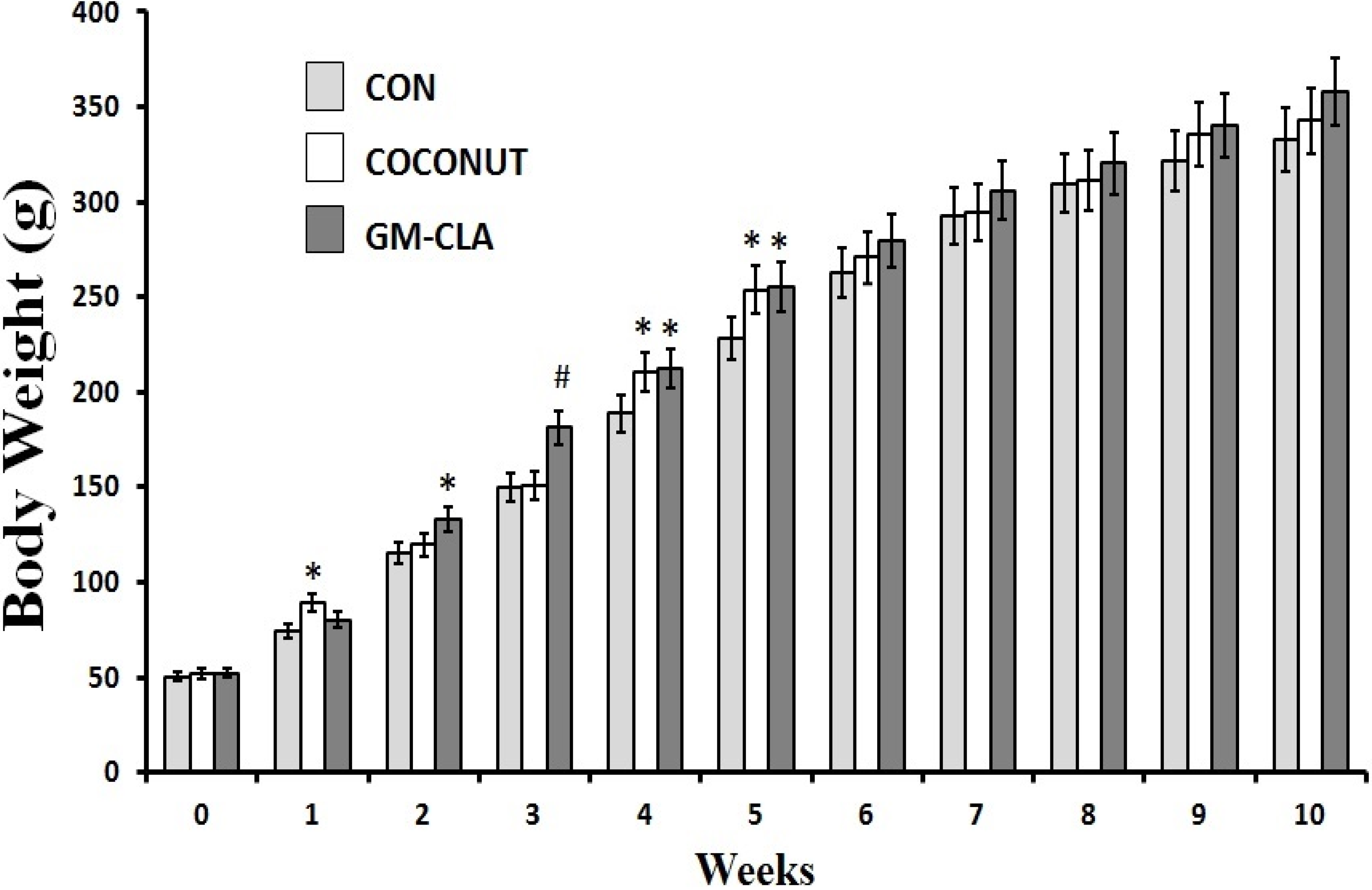
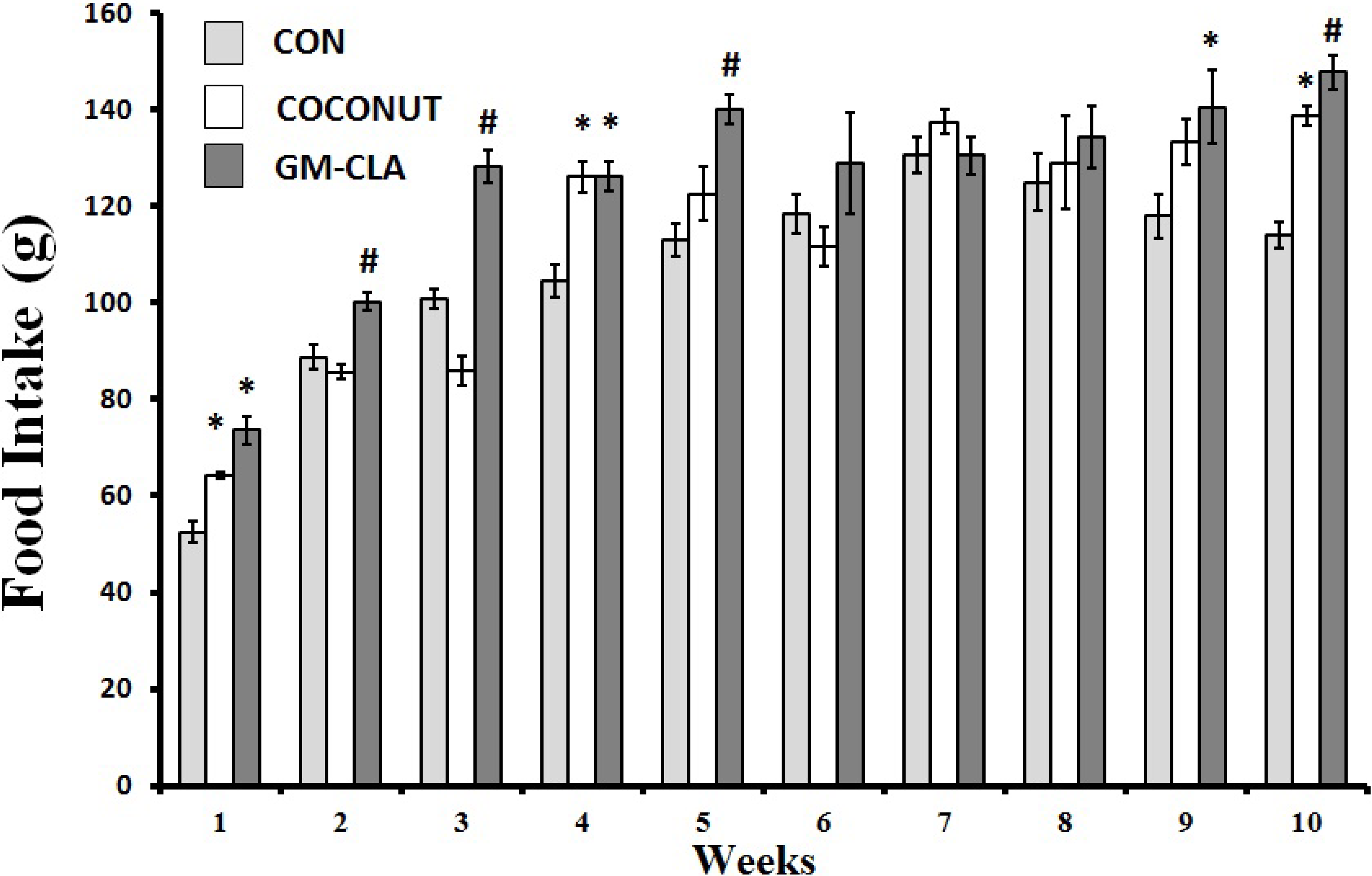
2.1. Body Weight and Feed Intake
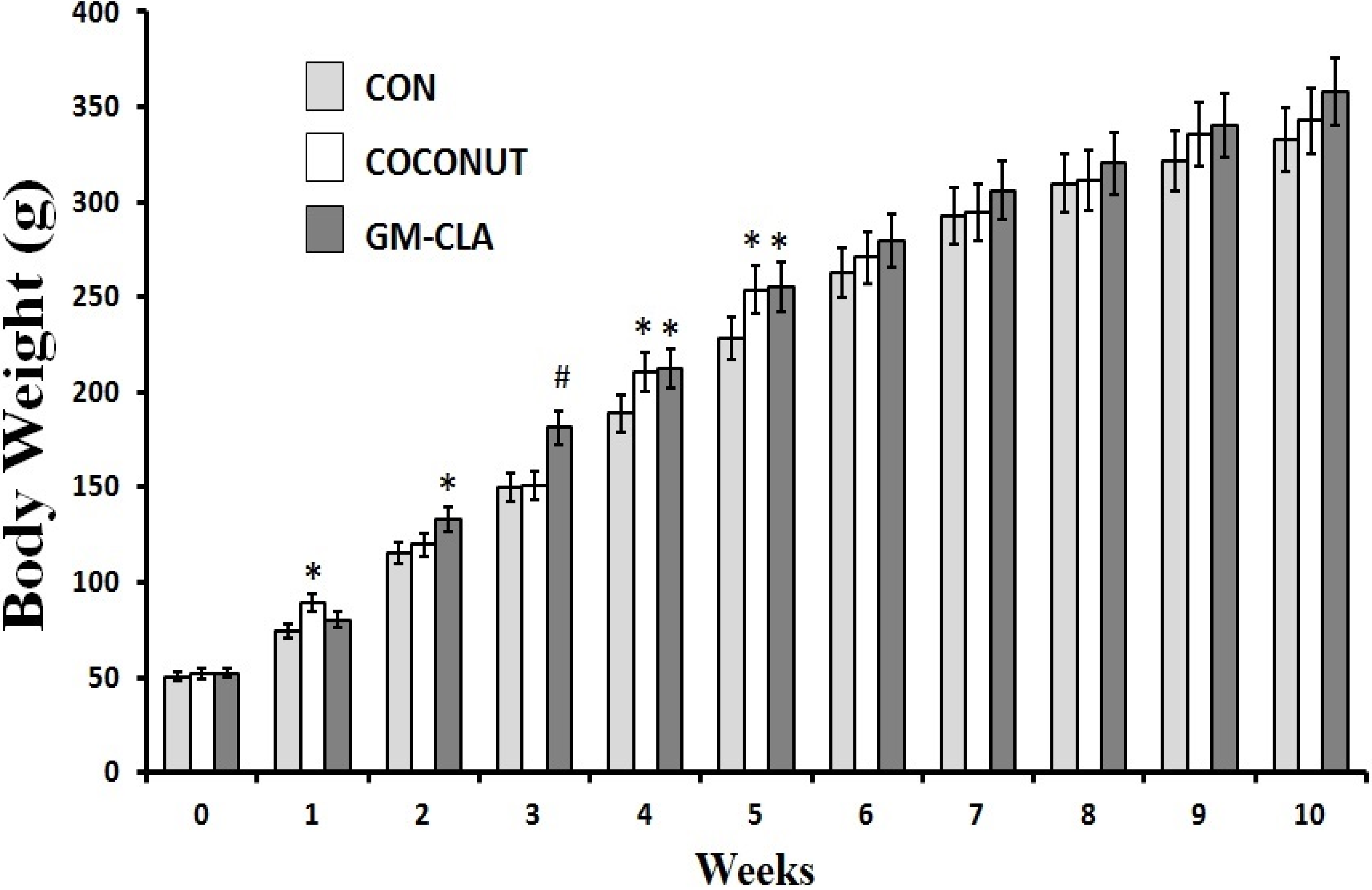
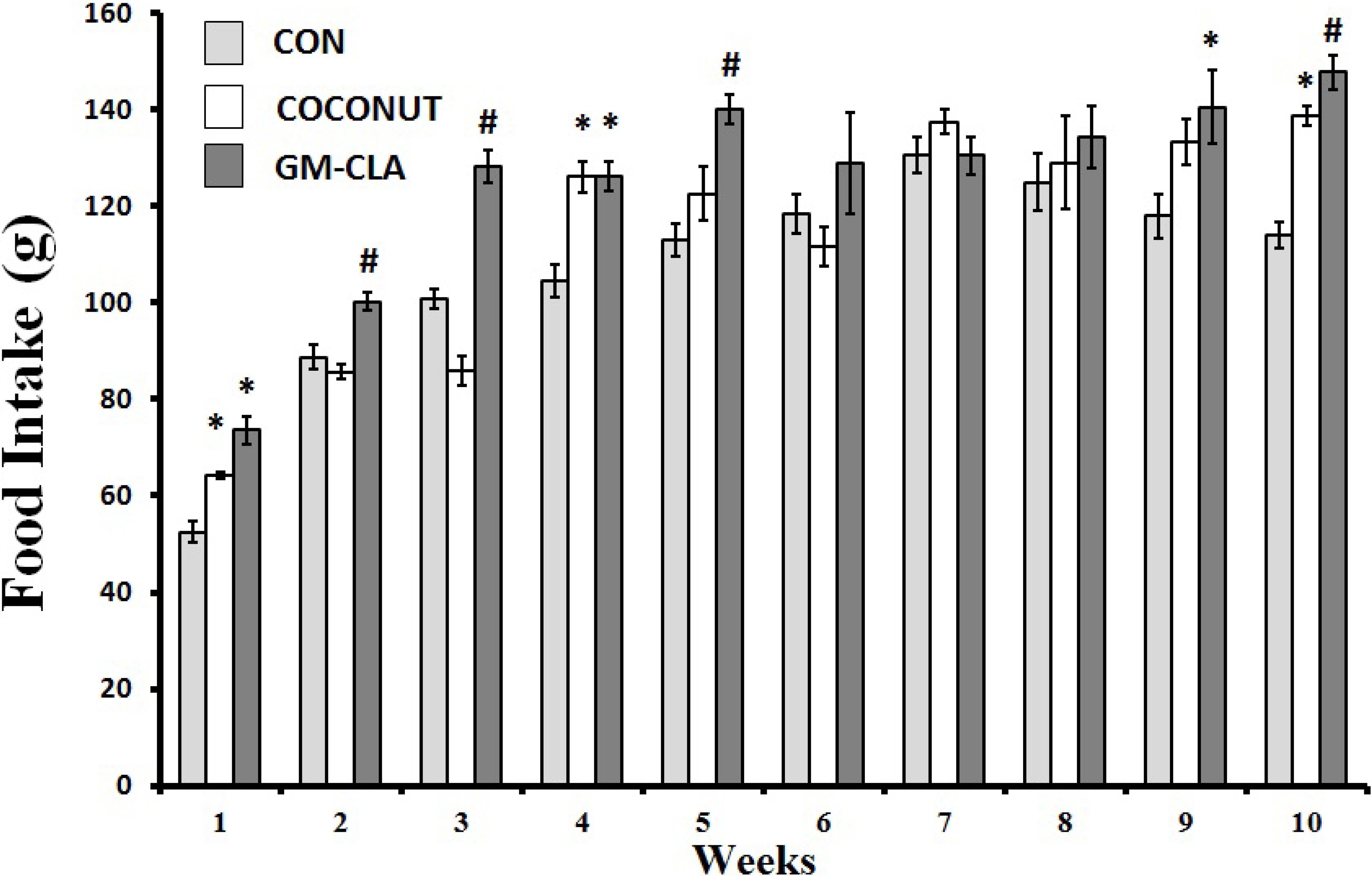
2.2. Biochemical Analysis

{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | Coconut Oil | Goat Milk Fat |
|---|---|---|---|
| Total Cholesterol | 67.17 (±7.60) | 73.83 (±5.00) | 87.00 (±11.38) # |
| HDL | 16.08 (±1.86) | 15.22 (±1.58) | 26.63 (±3.16) # |
| LDL | 50.01 (±18.08) | 42.81 (±8.45) | 53.27 ±8.59) |
| TAG | 40.78 (±4.84) | 64.00 (±4.12) * | 35.50(±3.66) # |
| TG/HDL | 2.82 (±1.22) | 4.06 (±0.71) * | 1.36(±0.26) # |
| Glucose | 81.33(±7.94) | 64.86(±4.56) | 107.88(± 8.5) § |
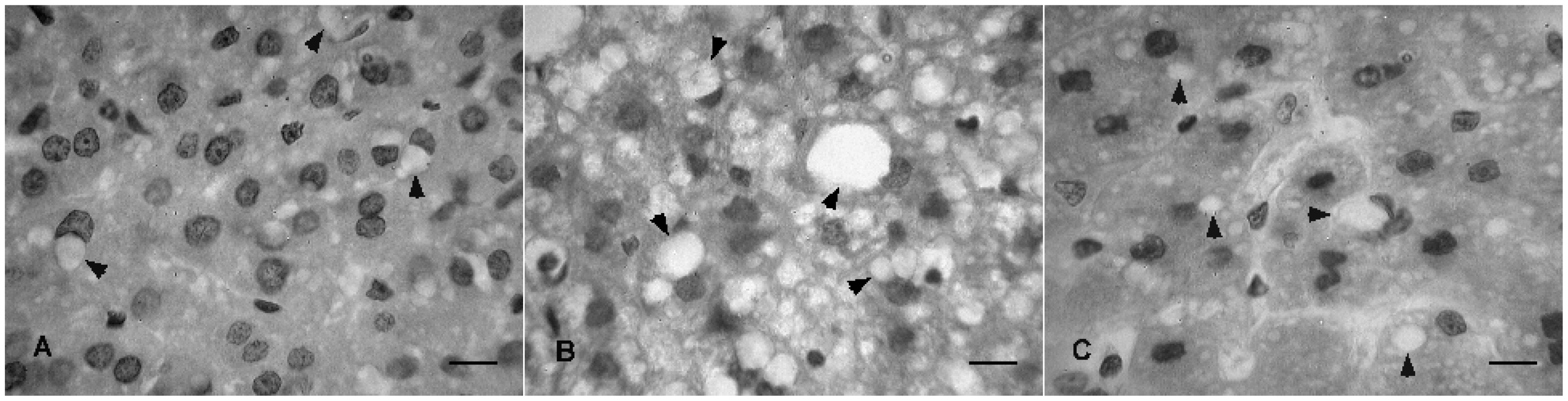
2.3. Histopathological Analysis
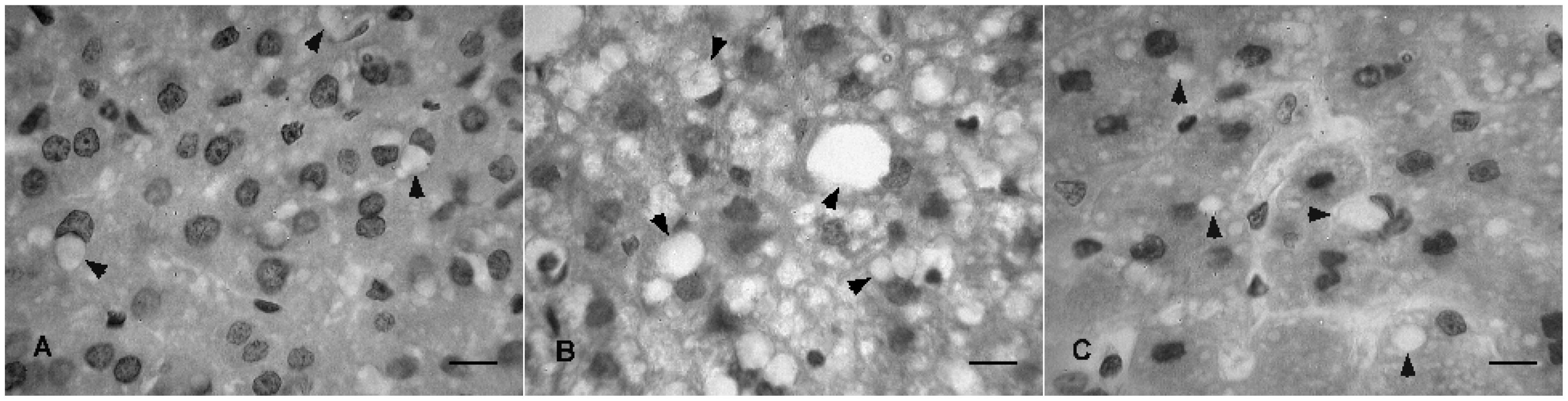
3. Experimental
3.1. Obtainment of the Lipids from Goat Milk
| Fatty acids | Control | Coconut Oil | Goat Milk Fat |
|---|---|---|---|
| C4:0 | - | - | - |
| C6:0 | - | - | 0.96 |
| C8:0 | - | - | 0.99 |
| C10:0 | - | - | 3.60 |
| C12:0 | - | 47.92 | 2.10 |
| C14:0 | 0.42 | 14.83 | 6.40 |
| C15:0 | - | - | 0.63 |
| C16:0 | 17.1 | 9.34 | 24.26 |
| C16:1 | - | - | 0.34 |
| C17:0 | 0.34 | - | 0.53 |
| C18:0 | 5.93 | 4.25 | 20.94 |
| C18:1(n9) | 28.53 | 19.2 | 28.79 |
| C18:2(n6) | 42.56 | 4.45 | 5.97 |
| C18:2 *CLA | - | - | 1.20 |
| C18:3(n3) | 4.89 | - | 0.63 |
| Not Determined | 0.23 | 0.01 | 2.66 |
3.2. Animals and Diet
3.3. Measurement of Dietary Fatty Acids
3.4. Biochemical Analysis
3.5. Tissue Preparation
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflictts of Interest
References
- Jones, D.S.; Podolsky, S.H.; Greene, J.A. The burden of disease and the changing task of medicine. N. Engl. J. Med. 2012, 366, 2333–2338. [Google Scholar] [CrossRef]
- Kostogrys, R.B.; Pisulewski, P.M. Effect of conjugated linoleic acid (CLA) onlipid profile and liver histology in laboratory rats fed high-fructose diet. Environ. Toxicol. Pharmacol. 2010, 30, 245–250. [Google Scholar] [CrossRef]
- Chandrashekar, P.; Lokesh, B.R.; Krishna, A.G.G. Hypolipidemic effect of blends of coconut oil with soybean oil or sunflower oil in experimental rats. Food Chem. 2010, 123, 728–733. [Google Scholar] [CrossRef]
- Dietschy, J.M. Dietary fatty acids and the regulation of plasma Low Density Lipoprotein cholesterol concentrations. J. Nutr. 1998, 128, 444–448. [Google Scholar]
- Pariza, M.W.; Hargraves, W.A. A beef-derived mutagenesis modulator inhibits initiation of mouse epidermal tumors by 7,12-dimethylbenz[a]anthracene. Carcinogenesis 1985, 6, 591–593. [Google Scholar]
- Hernandez-Diaz, G.; Alexander-Aguilera, A.; Arzaba-Villalba, A.; Soto-Rodriguez, I.; Garcia, G. Effect of conjugated linoleic acid on body fat, tumornecrosis factor alpha and resistin secretion in spontaneously hypertensive rats. Prostag. Leukotr. Ess. 2010, 82, 105–109. [Google Scholar] [CrossRef]
- Parra, P.; Serra, F.; Palou, A. Moderate doses of conjugated linoleic acid isomers mix contribute to lowering body fat content maintaining insulin sensitivity and a noninflammatory pattern in adipose tissue in mice. J. Nutr. Biochem. 2010, 21, 107–115. [Google Scholar] [CrossRef]
- Terpstra, A.H.M. Effect of conjugated linoleic acid on body composition and plasma lipids in humans: An overview of the literature. Am J. Clin. Nutr. 2004, 79, 352–361. [Google Scholar]
- Kennedy, A.; Martinez, K.; Schmidt, S.; Mandrup, S.; LaPoint, K.; McIntosh, M. Antiobesity mechanisms of action of conjugated linoleic acid. J. Nutr. Biochem. 2010, 21, 171–179. [Google Scholar] [CrossRef]
- Arbones-Mainar, J.M.; Navarro, M.A.; Guzman, M.A.; Arnal, C.; Surra, J.C.; Acin, S.; Carnicer, R.; Osada, J.; Roche, H.M. Selective effect of conjugated linoleic acid isomers on atherosclerotic lesion development in apolipoprotein E knockout mice. Atherosclerosis 2006, 189, 318–327. [Google Scholar] [CrossRef]
- Kelley, N.S.; Hubbard, N.E.; Erickson, K.L. Conjugated linoleic acid isomers and cancer. J. Nutr. 2008, 137, 2599–2607. [Google Scholar]
- Lee, K.W.; Lee, H.J.; Cho, H.Y.; Kim, Y.J. Role of the conjugated linoleic acid in the prevention of cancer. Crit. Rev. Food Sci. Nutr. 2005, 45, 135–144. [Google Scholar] [CrossRef]
- Haenlein, G.F.W. Goat milk in human nutrition. Small Rumin. Res. 2004, 51, 155–163. [Google Scholar] [CrossRef]
- Alferez, M.J.M.; Lopez-Aliaga, I.; Nestares, T.; Diaz-Castro, J.; Barrionuevo, M.; Ros, P.B.; Campos, M.S. Dietary goat milk improves iron bioavailability in rats with induced ferropenic anaemia in comparison with cow milk. Int. Dairy J. 2006, 16, 813–821. [Google Scholar] [CrossRef]
- Barrionuevo, M.; Alferez, M.J.M.; Lopez-Aliaga, I.; Sanz Sampelayo, M.R.; Campos, M.S. Beneficial effect of goat milk on nutritive utilization of iron and copper in malabsorption syndrome. J. Dairy Sci. 2002, 85, 657–664. [Google Scholar] [CrossRef]
- Lopez-Aliaga, I.; Alferez, M.J.M.; Barrionuevo, M.; Lisbona, F.; Campos, M.S. Influence of goat and cow milk on the digestive and metabolic utilization of calcium and iron. J. Physiol. Biochem. 2006, 56, 201–208. [Google Scholar]
- Tsiplakou, E.; Mountzouris, K.C.; Zervas, G. Concentration of conjugated linoleic acid in grazing sheep and goat milk fat. Livest. Sci. 2006, 103, 74–84. [Google Scholar] [CrossRef]
- Bouattour, M.A.; Casals, R.; Albanell, E.; Such, X.; Caja, G. Feeding soybean oil to dairy goats increases conjugated linoleic acid in milk. J. Dairy Sci. 2008, 91, 2399–2407. [Google Scholar] [CrossRef]
- Soares, J.K.B.; Rocha-De-Melo, A.P.; Medeiros, M.C.; Queiroga, R.C.R.E.; Bomfim, M.A.D.; Souza, A.F.O.D.; Nascimento, A.L.V.; Guedes, R.C.A. Conjugated linoleic acid in the maternal diet differentially enhances growth and cortical spreading depression in the rat progeny. Biochim. Biophys. Acta 2012, 1820, 1490–1495. [Google Scholar] [CrossRef]
- He, M.L.; Mir, P.S.; Okine, E.K.; Napadajlo, H. Effect of conjugated linoleic acids from beef or industrial hydrogenation on growth and adipose tissue characteristics of rats. Nutr. Metab. 2009, 6. [Google Scholar] [CrossRef]
- Chin, S.F.; Storksoh, J.M.; Albright, K.J.; Cook, M.E.; Pariza, M.W. Conjugated linoleic acid is a growth factor for rats as shown by enhanced weight gain and improved feed efficiency. J. Nutr. 1994, 124, 2344–2349. [Google Scholar]
- Ramirez-Santana, C.; Castellote, C.; Castell, M.; Molto-Puigmarti, C.; Rivero, M.; Perez-Cano, F.J.; Franch, A. Enhancement of antibody synthesis in rats by feeding cis-9, trans-11 conjugated linoleic acid during early life. J. Nutr. Biochem. 2011, 22, 495–501. [Google Scholar] [CrossRef]
- Hayashi, A.A.; Medeiros, S.R.; Carvalho, M.H.; Lanna, D.P. Conjugated linoleic acid (CLA) effects on pups growth, milk composition and lipogenic enzymes in lactating rats. J. Dairy Res. 2007, 74, 160–166. [Google Scholar] [CrossRef]
- Park, P.; Albright, K.J.; Storkson, J.M.; Liu, W.; Pariza, M.W. Effects of dietary conjugated linoleic acid (CLA) on spontaneously hypertensive rats. J. Fun. Foods 2010, 2, 54–59. [Google Scholar] [CrossRef]
- Inoue, N.; Nagao, K.; Hirata, J.; Wang, Y.M.; Yanagita, T. Conjugated linoleic acid prevents the development of essential hypertension in spontaneously hypertensive rats. Biochem. Biophys. Res. Commun. 2004, 323, 679–684. [Google Scholar] [CrossRef]
- Halade, G.V.; Rahman, M.M.; Fernandes, G. Differential effects of conjugated linoleic acid isomers in insulin-resistant female C57Bl/6J mice. J. Nutr. Biochem. 2010, 21, 332–337. [Google Scholar] [CrossRef]
- Declercq, V.; Zahradka, P.; Taylor, C.G. Dietary t10,c12-CLA but not c9,t11 CLA reduces adipocyte size in the absence of changes in the adipose renin–angiotensin system in fa/fa Zucker rats. Lipids 2010, 45, 1025–1033. [Google Scholar] [CrossRef]
- So, M.H.H.; Tse, I.M.Y.; Li, E.T.S. Dietary fat concentration influences the effects of trans-10, cis-12 conjugated linoleic acid on temporal patterns of energy intake and hypothalamic expression of appetite-controlling genes in mice. J. Nutr. 2009, 139, 145–151. [Google Scholar]
- Lopez-Aliaga, M.J.M.; Alferez, M.T.; Nestares, P.B.; Ros, M.; Barrionuevo, M.; Campos, M.S. Goat milk feeding causes an increase in biliary secretion of cholesterol and a decrease in plasma cholesterol levels in rats. J. Dairy Sci. 2005, 88, 1024–1030. [Google Scholar] [CrossRef]
- Takahashi, Y.; Kushiro, M.; Shinohara, K.I.T. Dietary conjugated linoleic acid reduces body fat mass and affects gene expression of proteins regulating energy metabolism in mice. Comp. Bioch. Phys. Bioch. Mol. Bio. 2002, 133, 395–404. [Google Scholar]
- Shimano, H.; Yakagi, N.; Amemiya-Kudo, M.; Hasty, A.H.; Osuga, J.; Tamura, Y.; Shionoiri, F.; Iizuka, Y.; Ohashi, K.; Harada, K.; et al. Sterol regulatory element-binding protein-1 is a key transcription factor for nutritional induction of lipogenic enzyme genes. J. Biol. Chem. 1999, 274, 35832–35839. [Google Scholar] [CrossRef]
- Granlund, L.; Juvet, L.K.; Pedersen, J.I.; Nebb, H.I. Trans 10, cis 12-conjugated linoleic acid prevents triacylglycerol accumulation in adipocytes by acting as a PPARγ modulator. J. Lipid Res. 2003, 44, 1441–1452. [Google Scholar] [CrossRef]
- Mitchell, P.L.; Langille, M.A.; Currie, D.L.; McLeod, R.S. Effect of conjugated linoleic acid isomers on lipoproteins and atherosclerosis in the Syrian Golden hamster. Biochim. Biophys. Acta 2005, 1734, 269–276. [Google Scholar]
- Nestel, P.; Fujii, A.; Allen, T. The cis-9,trans-11 isomer of conjugated linoleic acid (CLA) lowers plasma triglyceride and raises HDL cholesterol concentrations but does not suppress aortic atherosclerosis in diabetic apoE-deficient. Atherosclerosis 2006, 189, 282–287. [Google Scholar]
- Diniz, Y.W.; Santos, P.P.; Assalin, H.B.; Souza, G.A.; Rocha, K.K.; Ebaid, G.M.; Seiva, F.R.; Amauchi, J.F.; Novelli-Filho, J.L.V.; Novelli, E.L. Conjugated linoleic acid and cardiac health: Oxidative stress and energetic metabolism in standard and sucroserich diets. Eur. J. Pharmacol. 2008, 579, 318–325. [Google Scholar] [CrossRef]
- Cooper, M.H.; Miller, J.R.; Mitchell, P.L.; Currie, D.L.; McLeod, R.S. Conjugated linoleic acid isomers have no effect on atherosclerosis and adverse effects on lipoprotein and liver lipid metabolism in apoE−/− mice fed a high-cholesterol diet. Atherosclerosis 2008, 200, 294–302. [Google Scholar] [CrossRef]
- Park, Y. Conjugated linoleic acid (CLA): Good or bad trans fat? J. Food Compos. Anal. 2009, 22S, S4–S12. [Google Scholar] [CrossRef]
- Branco, A.C.S.C.; Diniz, M.F.F.M.; Almeida, R.N.; Santos, H.B.; Oliveira, K.M.; Ramalho, J.A.; Dantas, J.G. Biochemical and hematological parameters of wistar rats and swiss mice in the professor Thomas George animal laboratory. Rev. Bras. Ciênc. Saúde 2011, 15, 209–214. [Google Scholar]
- Ashwell, M.S.; Ceddia, R.P.; House, R.L.; Cassady, J.P.; Eisen, E.J.; Eling, T.E.; Collins, J.B.; Grissom, S.F.; Odle, J. Trans-10, cis-12-conjugated linoleic acid alters hepatic gene expression in a polygenic obese line of mice displaying hepatic lipidosis. J. Nutr. Biochem. 2009, 21, 848–855. [Google Scholar]
- Wendel, A.A.; Martha, A.B. Effects of conjugated linoleic acid and troglitazone on lipid accumulation and composition in lean and Zucker diabetic fatty (fa/fa) rats. Lipids 2006, 41, 241–247. [Google Scholar] [CrossRef]
- Andreoli, M.F.; Illesca, P.G.; Gonzalez, M.A.; Bernal, C.A. Conjugated linoleic acid reduces hepatic steatosis and restores liver triacylglycerol secretion and the fatty acid profile during protein repletion in rats. Lipids 2010, 45, 1035–1045. [Google Scholar] [CrossRef]
- Musso, G.; Gambino, R.; Cassader, M. Recent insights into hepatic lipid metabolism in non-alcoholic fatty liver disease (NAFLD). Prog. Lip. Res. 2009, 48, 1–26. [Google Scholar] [CrossRef]
- Purushotam, A.; Shrode, G.E.; Wendel, A.A.; Liu, L.F.; Belury, M.A. Conjugated linoleic acid does not reduce body fat but decreases hepatic steatosis in adult Wistar rats. J. Nutr. Biochem. 2007, 18, 676–684. [Google Scholar] [CrossRef]
- Silva, M.A.F.; Ataide, T.R.; Oliveira, S.L.; Sant’Ana, A.E.G.; Cabral Junior, C.R.; Balwani, M.C.L.V.; Oliveira, F.G.S.; Santos, M.C. Hepatoprotective effect of diheptanoin and tritreptanoin chronic consumption against steatosis in rats. Arq. Bras. Endocrinol. Metab. 2008, 52, 1145–1155. [Google Scholar]
- Stanciuc, N.; Damitrascu, G.; Rapeanu, S. Glutamul transferase inactivation in milk and cream: A comparative kinetic study. Innov. Food Sci. Emerg. Technol. 2011, 12, 56–61. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, J.R. AIN-93 purified diets for laboratory rodents: Final report of American Institute of Nutrition ad hoc writing Committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar]
- Bligh, E.C.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can.J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Rautela, G.S.; Liedtke, R.J. Automed enzymic measurement of total cholesterol in serum. Clin. Chem. 1978, 24, 108–114. [Google Scholar]
- Friedewald, W.T.; Levy, R.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma without use of preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rodrigues, R.; Soares, J.; Garcia, H.; Nascimento, C.; Medeiros, M.; Bomfim, M.; Medeiros, M.C.; Queiroga, R. Goat Milk Fat Naturally Enriched with Conjugated Linoleic Acid Increased Lipoproteins and Reduced Triacylglycerol in Rats. Molecules 2014, 19, 3820-3831. https://doi.org/10.3390/molecules19033820
Rodrigues R, Soares J, Garcia H, Nascimento C, Medeiros M, Bomfim M, Medeiros MC, Queiroga R. Goat Milk Fat Naturally Enriched with Conjugated Linoleic Acid Increased Lipoproteins and Reduced Triacylglycerol in Rats. Molecules. 2014; 19(3):3820-3831. https://doi.org/10.3390/molecules19033820
Chicago/Turabian StyleRodrigues, Raphaela, Juliana Soares, Hugo Garcia, Claudenice Nascimento, Maria Medeiros, Marco Bomfim, Maria Carmo Medeiros, and Rita Queiroga. 2014. "Goat Milk Fat Naturally Enriched with Conjugated Linoleic Acid Increased Lipoproteins and Reduced Triacylglycerol in Rats" Molecules 19, no. 3: 3820-3831. https://doi.org/10.3390/molecules19033820
APA StyleRodrigues, R., Soares, J., Garcia, H., Nascimento, C., Medeiros, M., Bomfim, M., Medeiros, M. C., & Queiroga, R. (2014). Goat Milk Fat Naturally Enriched with Conjugated Linoleic Acid Increased Lipoproteins and Reduced Triacylglycerol in Rats. Molecules, 19(3), 3820-3831. https://doi.org/10.3390/molecules19033820