Fatty Acids from Pool Lipids as Possible Precursors of the Male Marking Pheromone in Bumblebees


Abstract
:1. Introduction
2. Results and Discussion
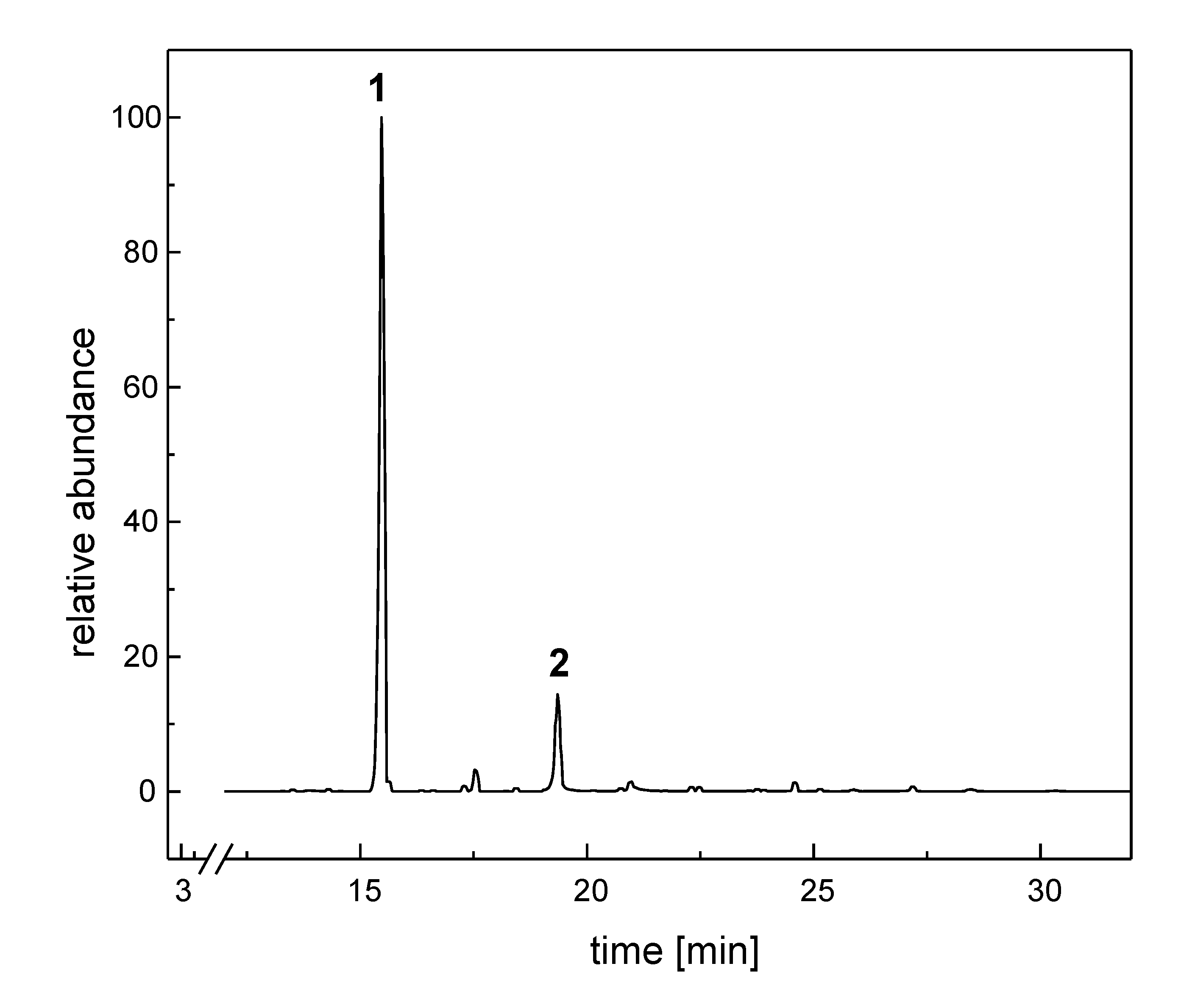

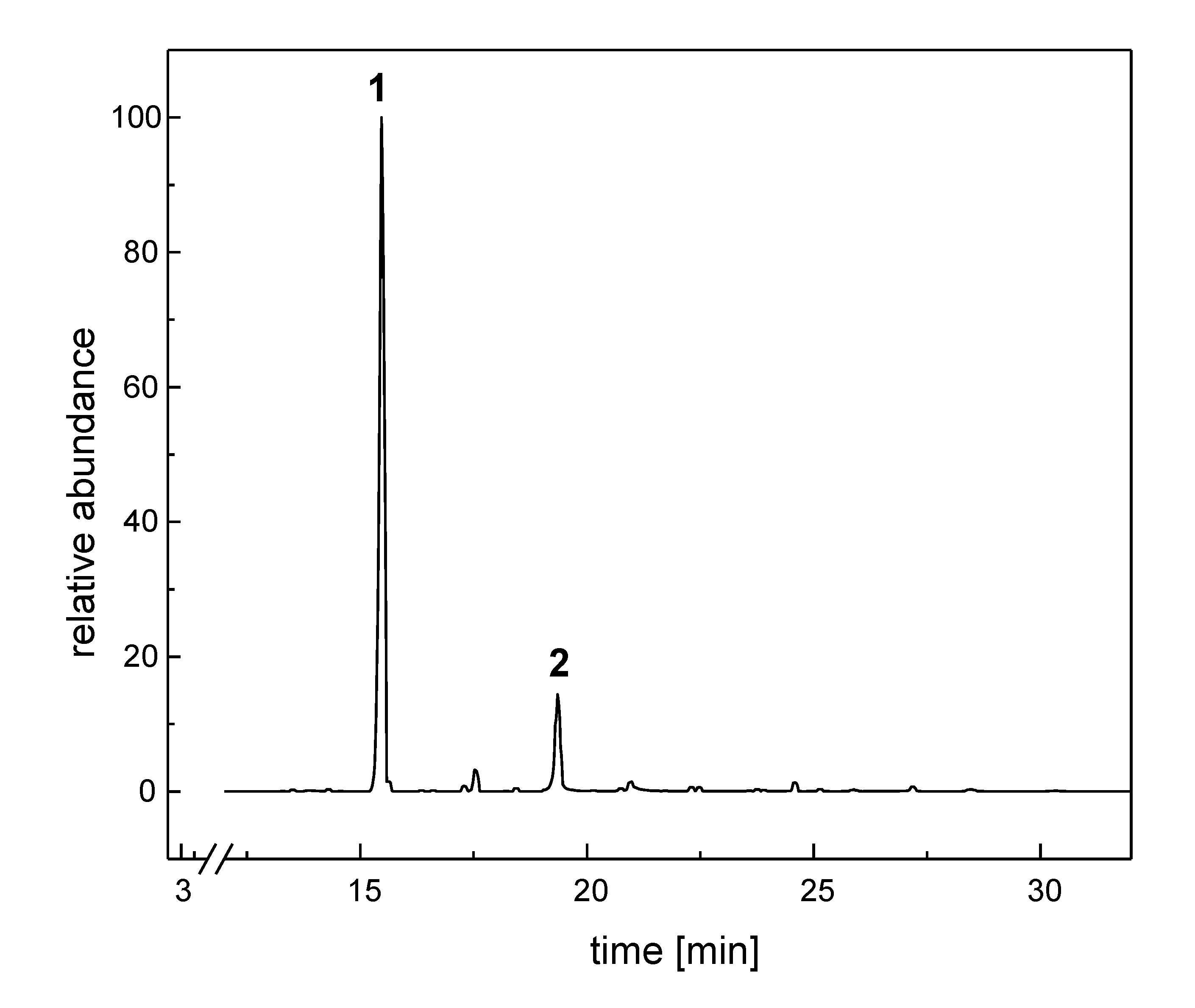{kind=link}
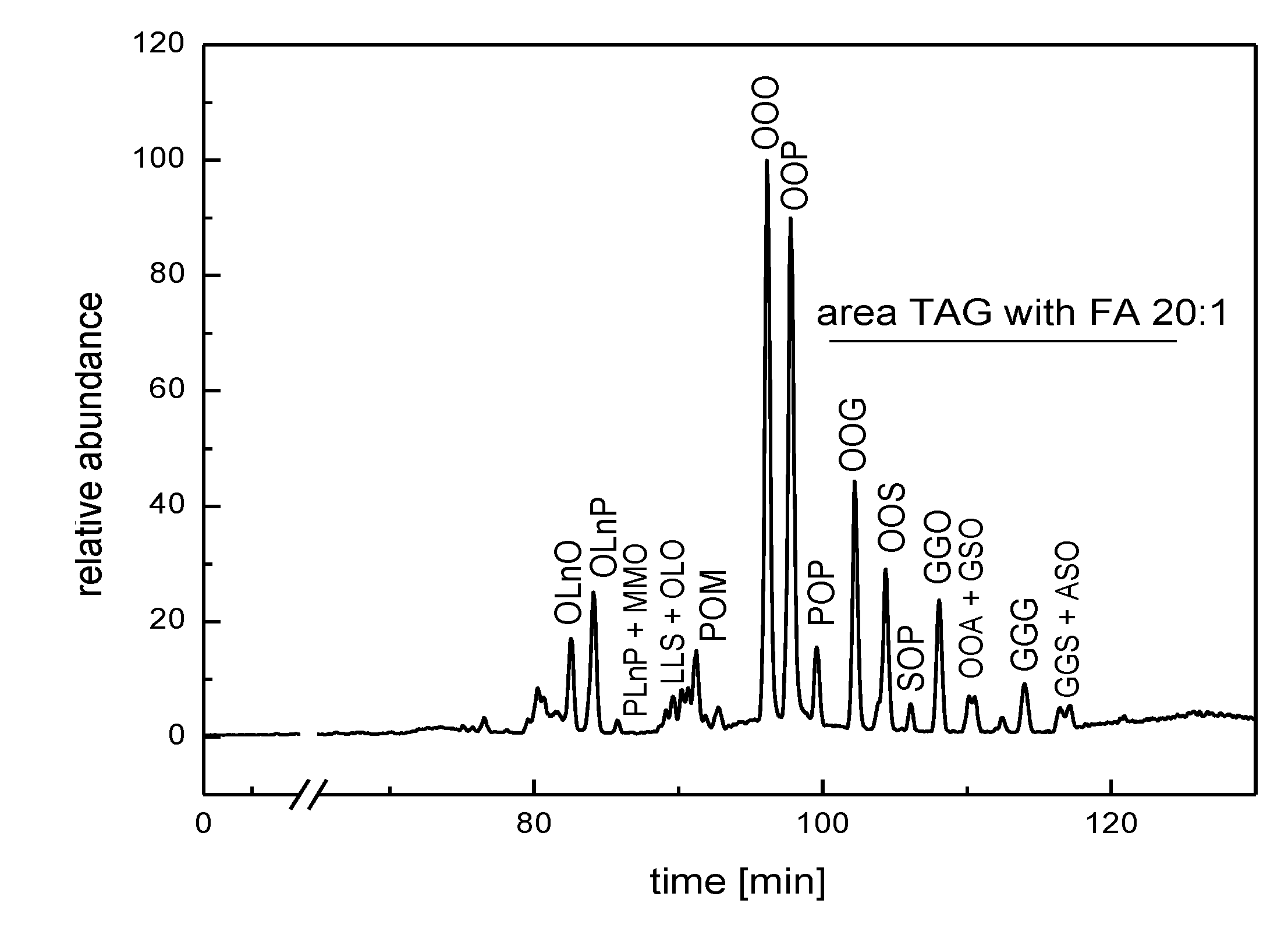
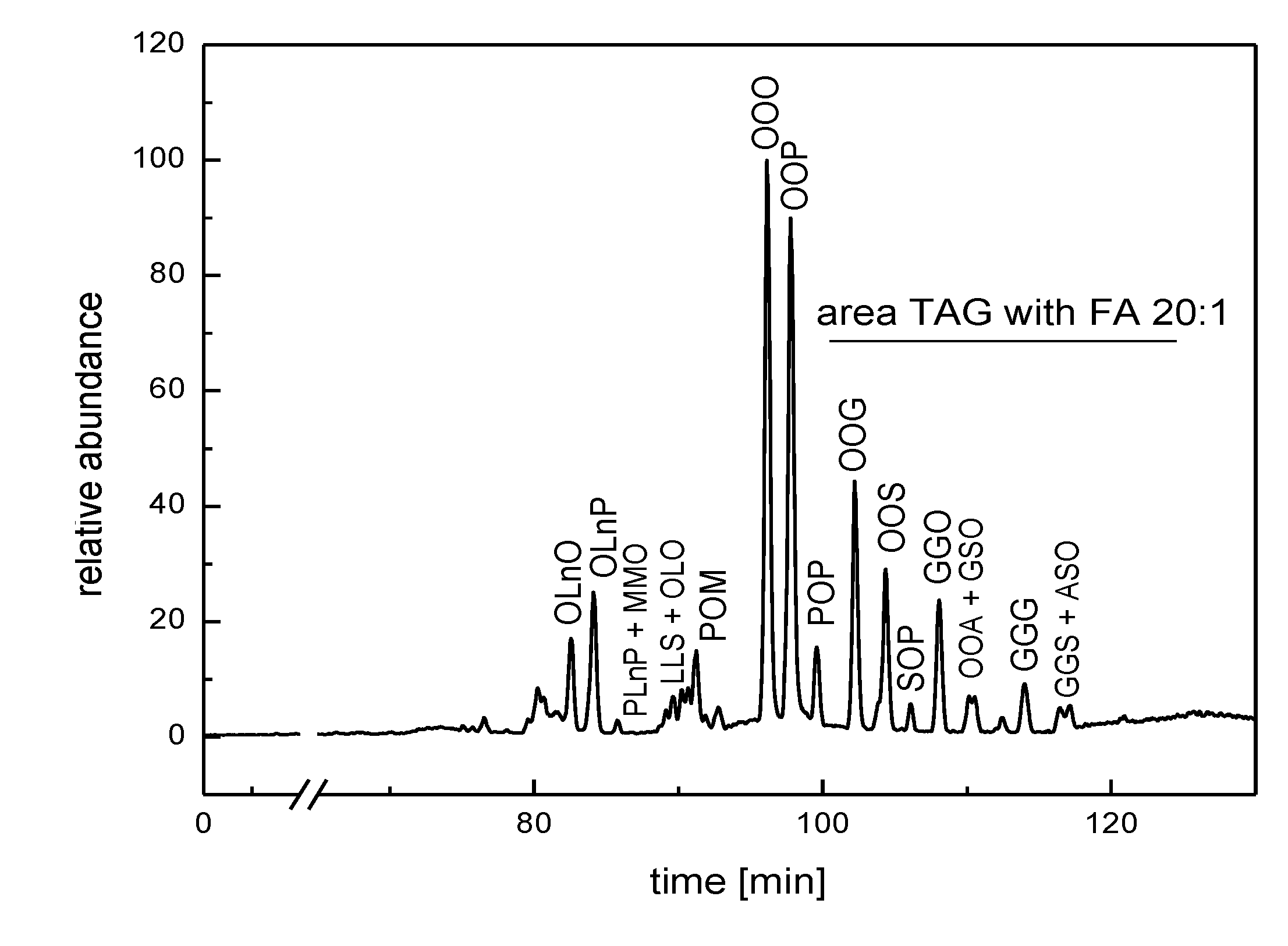{kind=link}
| Compound | Mean (%) | Median (%) | Standard deviation (%) |
|---|---|---|---|
| (E,E)-Farnesol | 0.7 | 0.02 | 1.4 |
| Octadec-9-ene | 0.2 | 0.1 | 0.2 |
| Nonadec-9-ene | 76.7 | 78.0 | 8.4 |
| Nonadecane | 0.9 | 0.6 | 0.5 |
| Henicos-9-ene | 0.4 | 0.2 | 0.4 |
| Henicosane | 1.2 | 0.8 | 0.8 |
| Icos-15-en-1-ol | 15.0 | 16.4 | 8.3 |
| Tricosane | 3.6 | 3.1 | 2.4 |
| Pentacos-9-ene | 0.1 | 0.07 | 0.2 |
| Pentacosane | 0.4 | 0.2 | 0.3 |
| Docosen-1-ol a | 0.1 | 0.0 | 0.1 |
| Heptacos-9-ene | 0.2 | 0.1 | 0.4 |
| Heptacosane | 0.1 | 0.1 | 0.1 |
| Farnesyl tetradecenoate b | 0.4 | 0.07 | 0.7 |
| Hentriacont-9-ene | 0.1 | 0.1 | 0.1 |
| Farnesyl octadecadienoate | 0.2 | 0.1 | 0.3 |
| Icosenyl tetradecenoate c | 0.2 | 0.05 | 0.5 |
| ECN | CN:DB | Triacylglycerol (TG) a | Peak area relative % (mean ± SD; N = 4) |
|---|---|---|---|
| 40 | 48:4 | 18:3_18:1_12:0; 18:3_14:0_16:1 | 0.1 ± 0.1 |
| 46:3 | 16:0_18:3_12:0; 14:0_14:0_18:3 | 0.1 ± 0.1 | |
| 42 | 54:6 | 18:1_18:3_18:2; 18:2_18:2_18:2 | 0.2 ± 0.1 |
| 52:5 | 18:3_18:1_16:1; 18:2_18:2_16:1; 18:2_16:0_18:3 | 0.8 ± 0.2 | |
| 50:4 | 18:3_18:1_14:0; 18:3_16:0_16:1 | 0.8 ± 0.2 | |
| 48:3 | 16:0_18:3_14:0; 18:2_18:1_12:0; 18:2_14:0_16:1 | 0.4 ± 0.4 | |
| 46:2 | 16:1_18:1_12:0; 16:1_16:1_14:0 | 0.1 ± 0.1 | |
| 44:1 | 14:0_18:1_12:0; 14:0_16:1_14:0 | 0.1 ± 0.1 | |
| 44 | 54:5 | 18:1_18:3_18:1 | 3.3 ± 1.8 |
| 52:4 | 18:1_18:3_16:0 | 5.1 ± 2.1 | |
| 50:3 | 16:0_18:3_16:0; 18:2_16:1_16:0 | 1.6 ± 0.2 | |
| 48:2 | 18:1_18:1_12:0; 18:1_16:1_14:0; 16:1_16:1_16:0 | 1.3 ± 0.5 | |
| 46:1 | 16:0_18:1_12:0; 14:0_14:0_18:1 | 0.7 ± 0.4 | |
| 46 | 56.5 | 18:1_18:1_20:3 | 0.7 ± 0.3 |
| 54:4 | 18:2_18:2_18:0; 18:1_18:1_18:2 | 2.2 ± 0.2 | |
| 52:3 | 18:2_18:1_16:0; 18:2_18:0_16:1; 18:1_18:1_16:1 | 7.1 ± 1.6 | |
| 50:2 | 18:1_18:1_14:0; 18:1_16:1_16:0 | 6.6 ± 3.2 | |
| 48:1 | 16:0_18:1_14:0 | 2.4 ± 1.4 | |
| 48 | 58:5 | 18:2_18:2_22:1 | 0.6 ± 0.1 |
| 56.4 | 18:1_18:1_20:2 | 0.5 ± 0.2 | |
| 54:3 | 18:1_18:1_18:1 | 19.4 ± 2.5 | |
| 52:2 | 18:1_18:1_16:0 | 22.8 ± 4.7 | |
| 50:1 | 16:0_18:1_16:0 | 4.0 ± 1.6 | |
| 50 | 56:3 | 18:1_18:1_20:1 | 4.9 ± 2.5 |
| 54:2 | 18:1_18:1_18:0 | 5.0 ± 0.6 | |
| 52:1 | 18:0_18:1_16:0 | 1.4 ± 0.5 | |
| 52 | 58:3 | 20:1_20:1_18:1; 18:1_22:1_18:1 | 2.5 ± 1.7 |
| 56:2 | 20:1_18:0_18:1; 18:1_18:1_20:0 | 1.1 ± 0.7 | |
| 54:1 | 20:0_16:0_18:1; 18:0_18:0_18:1 | 0.5 ± 0.1 | |
| 54 | 60:3 | 20:1_20:1_20:1; 20:1_22:1_18:1; 18:1_18:1_24:1 | 0.9 ± 0.8 |
| 58:2 | 20:1_20:1_18:0; 18:1_18:1_22:0 | 0.9 ± 0.7 | |
| 56:1 | 20:0_18:1_18:0 | 0.2 ± 0.1 | |
| 56 | 62:3 | 18:1_18:1_26:1 | 0.5 ± 0.5 |
| 60:2 | 18:1_18:1_24:0 | 0.4 ± 0.3 | |
| 58:1 | 22:0_18:1_18:0; 18:0_26:1_14:0; 18:0_24:0_16:1 | 0.1 ± 0.1 | |
| 58 | 64:3 | 18:1_18:1_28:1 | 0.2 ± 0.2 |
| 62:2 | 22:1_20:0_20:1; 18:1_18:1_26:0 | 0.2 ± 0.2 | |
| 60 | 66:3 | 18:1_18:1_30:1 | 0.2 ± 0.2 |
| 64:2 | 18:1_18:1_28:0 | 0.2 ± 0.1 |
| Fatty acid (Trivial name) | Code | Bombus species | ||
|---|---|---|---|---|
| ruderatus | campestris | bohemicus | ||
| Decanoic (Capric) | 10:0 | 0.3 | trace | 0.1 |
| Dodecanoic (Lauric) | 12:0 | - | 0.4 | 10.1 |
| Tetradec-9-enoic (Myristoleic) | 9-14:1 | 0.2 | 0.1 a | 6.0 |
| Tetradecanoic (Myristic) | 14:0 | 1.3 | 1.6 | 15.6 |
| Hexadec-9-enoic (Palmitoleic) | 9-16:1 | 5.1 | 1.2 | 14.3 |
| Hexadec-11-enoic (Palmitvaccenic) | 11-16:1 | 1.1 | - | 2.3 |
| Hexadecanoic (Palmitic) | 16:0 | 14.9 | 15.3 | 22.3 |
| Octadecadienoic a | 18:2 | 0.1 | 2.3 | 2.5 |
| Octadec-9-enoic (Oleic) | 9-18:1 | 68.5 | 56.1 | 23.5 |
| Octadec-11-enoic (cis-Vaccenic) | 11-18:1 | - | 4.3 | 1.2 |
| Octadec-13-enoic (No trivial name) | 13-18:1 | trace | - | 0.1 a |
| Octadecanoic (Stearic) | 18:0 | 4.1 | 12.6 | 1.6 |
| Icosadienoic a | 20:2 | - | - | trace |
| Icos-11-enoic (Gondoic) | 11-20:1 | 3.3 | 1.7 | 0.1 a |
| Icos-15-enoic (No trivial name) | 15-20:1 | trace | - | - |
| Icosanoic (Arachidic) | 20:0 | 0.5 | 1.4 | 0.1 |
| Docos-13-enoic (Erucic) | 13-22:1 | 0.5 | - | - |
| Docosanoic (Behenic) | 22:0 | 0.1 | 0.4 | - |
| Tetracosenoic a | 24:1 | 0.1 | - | - |
| Tetracosanoic (Lignoceric) | 24:0 | trace | 0.2 | - |
| ECN | CN:DB | Triacylglycerol (TG) a | Peak area relative % (mean ± SD; N = 5) |
|---|---|---|---|
| 40 | 54:7 | 18:2_18:3_18:2; 18:3_18:3_18:1 | 0.1 ± 0.1 |
| 52:6 | 18:2_18:3_16:1; 18:3_18:3_16:0 | 0.1 ± 0.1 | |
| 48:4 | 12:0_18:2_18:2 | 0.1 ± 0.0 | |
| 42 | 54:6 | 18:2_18:2_18:2; 18:3_18:1_18:2 | 0.2 ± 0.2 |
| 52:5 | 18:2_18:2_16:1; 16:1_18:3_18:1 | 0.3 ± 0.2 | |
| 50:4 | 14:0_18:2_18:2; 18:1_14:0_18:3 | 0.3 ± 0.1 | |
| 48:3 | 16:1_16:1_16:1; 16:0_18:3_14:0; 18:2_14:0_16:1 | 0.1 ± 0.0 | |
| 44 | 54:5 | 18:1_18:3_18:1 | 7.4 ± 4.0 |
| 52:4 | 18:1_18:3_16:0 | 6.1 ± 2.4 | |
| 50:3 | 16:1_14:0_20:2; 16:1_16:1_18:1; 16:0_16:1_18:2 | 0.2 ± 0.1 | |
| 48:2 | 18:1_18:1_12:0 | 0.4 ± 0.1 | |
| 46:1 | 12:0_18:1_16:0; 14:0_14:0_18:1; 16:0_16:1_14:0 | 0.3 ± 0.1 | |
| 46 | 56:5 | 18:2_18:2_20:1; 18:1_18:1_20:3; 18:1_18:2_20:2 | 0.7 ± 0.2 |
| 54:4 | 18:1_18:1_18:2; 18:3_18:1_18:0 | 3.8 ± 0.7 | |
| 52:3 | 16:1_18:1_18:1; 16:0_18:1_18:2 | 2.7 ± 0.2 | |
| 50:2 | 18:1_14:0_18:1; 16:0_18:1_16:1 | 3.4 ± 0.4 | |
| 48:1 | 16:0_16:1_16:0; 14:0_18:1_16:0 | 0.6 ± 0.2 | |
| 48 | 54:3 | 18:1_18:1_18:1 | 24.5 ± 2.3 |
| 52:2 | 16:0_18:1_18:1 | 27.4 ± 2.3 | |
| 50:1 | 16:0_18:1_16:0; 14:0_18:0_18:1 | 1.3 ± 0.5 | |
| 50 | 56:3 | 18:1_20:1_18:1 | 2.1 ± 1.0 |
| 54:2 | 18:1_18:0_18:1; 18:0_18:2_18:0 | 11.6 ± 2.3 | |
| 52:1 | 16:0_18:1_18:0; 18:0_16:1_18:0 | 2.7 ± 1.2 | |
| 52 | 58:3 | 18:1_20:1_20:1 | 0.1 ± 0.1 |
| 56:2 | 18:1_20:0_18:1 | 1.6 ± 0.7 | |
| 54:1 | 18:0_18:0_18:1 | 1.1 ± 0.5 |
| ECN | CN:DB | Triacylglycerol (TG) a | Peak area relative % (mean ± SD; N = 6) |
|---|---|---|---|
| 38 | 54:8 | 18:3_18:3_18:2 | 0.1 ± 0.0 |
| 52:7 | 16:1_18:3_18:3 | 0.2 ± 0.0 | |
| 50:6 | 14:0_18:3_18:3 | 0.2 ± 0.0 | |
| 48:5 | 14:1_16.1_18:3 | 0.1 ± 0.0 | |
| 46:4 | 12:0_16:1_18:3 | 0.2 ± 0.1 | |
| 44:3 | 14:0_12:0_18:3 | 0.2 ± 0.1 | |
| 40 | 54:7 | 18.3_18:1_18:3; 18:2_18:3_18:2 | 1.0 ± 0.3 |
| 52:6 | 18:3_18:3_16:0 | 1.0 ± 0.3 | |
| 50:5 | 16:1_16:1_18:3 | 0.8 ± 0.2 | |
| 48:4 | 12:0_18:3_18:1; | 1.9 ± 0.7 | |
| 46:3 | 16:1_16:1_14:1; 16:0_18:3_12:0 | 0.9 ± 0.3 | |
| 44:2 | 16:1_16:1_12:0 | 0.2 ± 0.1 | |
| 42 | 54:6 | 18:2_18:2_18:2; 18:3_18:1_18:2 | 1.3 ± 0.3 |
| 52:5 | 18:2_18:2_16:1; 16:1_18:3_18:1 | 4.5 ± 0.5 | |
| 50:4 | 14:0_18:2_18:2; 16:1_16:1_18:2; 18:3_16:0_16:1 | 5.6 ± 1.0 | |
| 48:3 | 16:1_16:1_16:1; 14:0_14:1_20:2; 14:0_16:0_18:3 | 2.5 ± 0.9 | |
| 46:2 | 16:1_18:1_12:0; 16:1_16:1_14:0 | 1.1 ± 0.6 | |
| 44:1 | 12:0_18:1_14:0; 14:0_16:1_14:0; 12:0_16:1_16:0 | 0.9 ± 0.4 | |
| 44 | 54:5 | 18:2_18:1_18:2; 18:1_18:3_18:1 | 9.4 ± 4.2 |
| 52:4 | 18:3_18:1_16:0; 18:2_18:2_16:0; 16:1_16:1_20:2 | 12.0 ± 3.2 | |
| 50:3 | 16:1_16:1_18:1; 14:0_16:0_20:2; 16:0_16:0_18:3 | 4.6 ± 1.4 | |
| 48:2 | 18:1_18:1_12:0; 18:1_16:1_14:0; 16:1_16:1_16:0 | 4.1 ± 1.4 | |
| 46:1 | 12:0_18:1_16:0; 14:0_14:0_18:1; 16:0_16:1_14:0 | 2.8 ± 0.9 | |
| 46 | 56:5 | 20:2_16:1_20:2; 18:1_18:1_20:3 | 0.3 ± 0.3 |
| 54:4 | 18:1_18:1_18:2; 18:2_16:0_20:2 | 3.6 ± 1.1 | |
| 52:3 | 16:1_18:1_18:1; 16:0_18:1_18:2 | 6.5 ± 0.5 | |
| 50:2 | 18:1_14:0_18:1; 16:0_18:1_16:1 | 8.1 ± 2.0 | |
| 48:1 | 16:0_16:1_16:0; 14:0_18:1_16:0 | 3.5 ± 1.6 | |
| 48 | 54:3 | 18:1_18:1_18:1 | 7.1 ± 2.9 |
| 52:2 | 16:0_18:1_18:1 | 9.7 ± 0.9 | |
| 50:1 | 16:0_18:1_16:0 | 2.6 ± 1.2 | |
| 50 | 56:3 | 18:1_18:1_20:1; 18.2_20:0_18:1 | 0.2 ± 0.2 |
| 54:2 | 18:1_18:0_18:1; 18:0_18:2_18:0 | 1.3 ± 0.3 | |
| 52:1 | 16:0_18:1_18:0; 18:0_16:1_18:0 | 0.6 ± 0.3 | |
| 52 | 58:3 | 18:1_18:1_22:1 | 0.1 ± 0.1 |
| 56:2 | 18:1_18:1_20:0 | 0.2 ± 0.1 | |
| 54:1 | 18:0_18:0_18:1; 20:0_16:0_18:1 | 0.2 ± 0.1 |
3. Experimental
3.1. Biological Materials
3.2. Sample Preparation
3.3. GC/MS Analysis of the Labial Gland Extracts
3.4. HPLC/APCI-MS Analysis of TGs
3.5. Transesterification of TG and GC/MS Analysis of FAMEs
4. Conclusions
Abbreviations
| APCI-MS | atmospheric pressure chemical ionisation mass spectrometry |
| CI | chemical ionisation |
| CN | number of carbon atoms |
| DB | number of double bonds |
| ECN | equivalent carbon number |
| FA | fatty acid |
| FAME | fatty acid methyl ester |
| FAR | fatty acyl-CoA reductase |
| FB | fat body |
| GC/MS | gas chromatography and mass spectrometry |
| HPLC | high performance liquid chromatography |
| LG | labial gland |
| SD | standard deviation |
| TG | triacylglycerol |
Supplementary Materials
Acknowledgments
Conflicts of Interest
References
- Calam, D.H. Species and sex-specific compounds from the heads of male bumblebees (Bombus spp.). Nature 1969, 221, 856–857. [Google Scholar] [CrossRef]
- Kullenberg, B.; Bergström, G.; Bringer, B.; Calberg, B.; Cederberg, B. Observations on scent marking by Bombus Latr. and Psithyrus Lep. males (Hym., Apidae) and localisation of site of production of the secretion. Zoon Suppl. 1973, 1, 23–25. [Google Scholar]
- Bergman, P.; Bergstrom, G. Scent marking, scent origin and species-specificity in male premating behavior of two Scandinavian bumblebees. J. Chem. Ecol. 1997, 23, 1235–1251. [Google Scholar] [CrossRef]
- Terzo, M.; Urbanová, K.; Valterová, I.; Rasmont, P. Intra and interspecific variability of the cephalic labial glands’ secretions in male bumblebees: the case of Bombus (Thoracobombus) ruderarius and B. (Thoracobombus) sylvarum [Hymenoptera, Apidae]. Apidologie 2005, 36, 85–96. [Google Scholar] [CrossRef]
- Bergman, P. Chemical Communication in Bumblebee Premating Behaviour. Ph.D. Dissertation, Göteborg University, Göteborg, Sweden, 1997. [Google Scholar]
- Coppée, A.; Mathy, T.; Cammaerts, M.-C.; Verheggen, F.; Terzo, M.; Iserbyt, S.; Valterová, I.; Rasmont, P. Age-dependent attractivity of males’ sexual pheromones in Bombus terrestris (L.) [Hymenoptera, Apidae]. Chemoecology 2011, 21, 75–82. [Google Scholar] [CrossRef]
- Žáček, P.; Kalinová, B.; Šobotník, J.; Hovorka, O.; Ptáček, V.; Coppée, A.; Verheggen, F.; Valterová, I. Comparison of age-dependent quantitative changes in the male labial gland secretion of Bombus terrestris and Bombus lucorum. J. Chem. Ecol. 2009, 35, 698–705. [Google Scholar] [CrossRef]
- Kullenberg, B.; Bergström, G.; Ställberg-Stenhagen, S. Volatile components of the cephalic marking secretion of male bumblebees. Acta Chem. Scand. 1970, 24, 1481–1483. [Google Scholar] [CrossRef]
- Bergström, G.; Svensson, B.G.; Appelgren, M.; Groth, I. Biosystematics of Social Insects; Elsevier: Waltham, MA, USA, 1981; p. 175. [Google Scholar]
- Cederberg, B.; Svensson, B.G.; Bergström, G.; Appelgren, M.; Groth, I. Male marking pheromones in North European cuckoo bumble bees, Psithyrus (Hymenoptera, Apidae). Nova Acta Regiae Soc. Ups. 1984, 3, 161–166. [Google Scholar]
- Hovorka, O.; Urbanová, K.; Valterová, I. Premating behavior of Bombus confusus males and analysis of their labial gland secretion. J. Chem. Ecol. 1998, 24, 183–193. [Google Scholar] [CrossRef]
- Hovorka, O.; Valterová, I.; Rasmont, P.; Terzo, M. Male cephalic labial gland secretions of two bumblebee species of the subgenus Cullumanobombus (Hymenoptera: Apidae: Bombus Latreille) and their distribution in Central Europe. Chem. Biodivers. 2006, 3, 1015–1022. [Google Scholar] [CrossRef]
- Urbanová, K.; Valterová, I.; Hovorka, O.; Kindl, J. Chemotaxonomical characterisation of males of Bombus lucorum collected in the Czech Republic. Eur. J. Entomol. 2001, 98, 111–115. [Google Scholar] [CrossRef]
- Urbanová, K.; Halík, J.; Hovorka, O. Kindl, J.; Valterová, I. Marking pheromones of the cuckoo bumblebee males (Hymenoptera, Apoidea, Bombus Latreille): Compositions of labial gland secretions of six species occurring in the Czech Republic. Biochem. Syst. Ecol. 2004, 32, 1025–1045. [Google Scholar] [CrossRef]
- Terzo, M.; Valterová, I.; Rasmont, P. Atypical secretions of the male cephalic labial glands in bumblebees: The case of Bombus (Rhodobombus) mesomelas Gerstaecker (Hymenoptera, Apidae). Chem. Biodivers. 2007, 4, 1466–1471. [Google Scholar] [CrossRef]
- Bertsch, A.; Schweer, H.; Titze, A.; Tanaka, H. Male labial gland secretions and mitochondrial DNA markers support species status of Bombus cryptarum and B. magnus (Hymenoptera, Apidae). Insectes Soc. 2005, 52, 45–54. [Google Scholar] [CrossRef]
- Valterová, I.; Urbanová, K. Chemical signals of bumblebees. Chem. Listy 1997, 91, 846–857. [Google Scholar]
- Terzo, M.; Valterová, I.; Urbanová, K.; Rasmont, P. De la nécessité de redécrire les phéromones sexuelles des mâles de bourdons [Hymenoptera, Apidae, Bombini] publiées avant 1996 pour leur utilisation en analyse phylogénétique. Phytoprotection 2003, 84, 39–51. [Google Scholar] [CrossRef]
- Morgan, D.E. Biosynthesis in Insects; RSC Press: Cambridge, UK, 2004. [Google Scholar]
- Tillman, J.A.; Seybold, S.J.; Jurenka, R.A.; Blomquist, G.J. Insect pheromones—An overview of biosynthesis and endocrine regulation. Insect Biochem. Mol. Biol. 1999, 29, 481–514. [Google Scholar] [CrossRef]
- Matsumoto, S.; Hull, J.J.; Ohnishi, A.; Moto, K.; Fónagy, A. Molecular mechanisms underlying sex pheromone production in the silkmoth, Bombyx mori: Characterization of the molecular components involved in bombykol biosynthesis. J. Insect Physiol. 2007, 53, 752–759. [Google Scholar] [CrossRef]
- Mann, J. Chemical Aspects of Biosynthesis; Oxford University Press: Oxford, UK, 1994. [Google Scholar]
- Lanne, B.S.; Bergström, G.; Wassgren, A.-B.; Törnbäck, B. Biogenetic pattern of straight chain marking compounds in male bumble bees. Comp. Biochem. Physiol. 1987, 88, 631–636. [Google Scholar]
- ek, P.; Prchalová-Horňáková, D.; Tykva, R.; Kindl, J.; Vogel, H.; Svatoš, A.; Pichová, I.; Valterová, I. ŽáčDe novo biosynthesis of sexual pheromone in the labial gland of bumblebee males. ChemBioChem 2013, 14, 361–371. [Google Scholar] [CrossRef]
- Luxová, A.; Valterová, I.; Stránský, K.; Hovorka, O.; Svatoš, A. Biosynthetic studies on marking pheromones of bumblebee males. Chemoecology 2003, 13, 81–87. [Google Scholar]
- Žáček, P.; Frišonsová, K.; Kindl, J.; Hovorka, O.; Kowalczuk, T.; Valterová, I. Application of the labeled precursors and analysis of their metabolites by means of GCxGC-MS—Tool for biosynthesis of the bumblebee male’s sexual pheromone research. ChemPlusChem 2014. submitted. [Google Scholar]
- Kofroňová, E.; Cvačka, J.; Vrkoslav, V.; Hanus, R.; Jiroš, P.; Kindl, J.; Hovorka, O.; Valterová, I. Comparison of HPLC/MS and MALDI-MS for characterizing triacylglycerols in insects: Specie-specific composition of lipids in fat bodies of bumblebee males. J. Chromatogr. B 2009, 877, 3878–3884. [Google Scholar] [CrossRef]
- Jiroš, P.; Cvačka, J.; Hanus, R.; Kindl, J.; Kofroňová, E.; Valterová, I. Changes in composition of triacylglycerols in the fat body of bumblebee males during their lifetime. Lipids 2011, 46, 863–871. [Google Scholar] [CrossRef]
- Cvačka, J.; Kofroňová, E.; Vašíčková, S.; Stránský, K.; Jiroš, P.; Hovorka, O.; Kindl, J.; Valterová, I. Unusual fatty acids in the fat body of the early bumblebee. Bombus pratorum. Lipids 2008, 43, 441–450. [Google Scholar] [CrossRef]
- Cvačka, J.; Hovorka, O.; Jiroš, P.; Kindl, J.; Stránský, K.; Valterová, I. Analysis of triacylglycerols in fat body of bumblebees by chromatographic methods. J. Chromatogr. A 2006, 1101, 226–237. [Google Scholar]
- Appelgren, M.; Bergström, G.; Svensson, B.G.; Cederberg, B. Marking pheromones of Megabombus bumble bee males. Acta Chem. Scand. 1991, 45, 972–974. [Google Scholar]
- Rasmont, P.; Terzo, M.; Coppée, A.; Valterová, I. The Species Specific Cephalic Secretions of the Homoplasic and Oligolectic Bumblebees Bombus consobrinus and Bombus gerstaeckeri. In Proceedings of the Third European Conference of Apidology, Belfast, UK, 8–11 September 2008.
- Valterová, I.; Terzo, M.; Rasmont, P. Institute of Organic Chemistry and Biochemistry, Academy of Sciences of the Czech Republic, Prague, Czech Republic. Unpublished work. 2003. [Google Scholar]
- Liebisch, G.; Vizcaíno, J.A.; Köfeler, H.; Trötzmüller, M.; Griffiths, W.J.; Schmitz, G.; Spener, F.; Wakelam, M.J.O. Shorthand notation for lipid structures derived from mass spectrometry. J. Lipid Res. 2013, 54, 1523–1530. [Google Scholar] [CrossRef]
- Oldham, N.J.; Svatoš, A. Determination of the double bond position in functionalized monoenes by chemical ionization ion-trap mass spectrometry using acetonitrile as a reagent gas. Rapid Commun. Mass Spec. 1999, 13, 331–336. [Google Scholar] [CrossRef]
- Antony, B.; Fujii, T.; Mot, K.; Matsumoto, S.; Fukuzawa, M.; Nakano, R.; Tatsuki, S.; Ishikawa, Y. Pheromone-gland-specific fatty-acyl reductase in the adzuki bean borer, Ostrinia scapulalis (Lepidoptera: Crambidae). Insect Biochem. Mol. Biol. 2009, 39, 90–95. [Google Scholar] [CrossRef]
- Tomčala, A.; Bártů, I.; Šimek, P.; Kodrík, D. Locust adipokinetic hormones mobilize diacylglycerols selectively. Comp. Biochem. Physiol. B 2010, 156, 26–32. [Google Scholar] [CrossRef]
- Bergman, P.; Bergström, G.; Appelgren, M. Labial gland marking secretion in males of two Scandinavian cuckoo bumblebee species (genus Psithyrus). Chemoecology 1996, 7, 140–145. [Google Scholar] [CrossRef]
- Vioque, J.; Kolattukudy, P.E. Resolution and purification of an aldehyde-generating and an alcohol-generating fatty acyl-CoA reductase from pea leaves (Pisum sativum L.). Arch. Biochem. Biophys. 1997, 340, 64–72. [Google Scholar] [CrossRef]
- Francis, G.W. Alkylthiolation for the determination of double bond positions in unsaturated fatty acid esters. Chem. Phys. Lipids 1981, 29, 369–374. [Google Scholar] [CrossRef]
- Kofroňová, E.; Cvačka, J.; Jiroš, P.; Sýkora, D.; Valterová, I. Analysis of insect triacylglycerols using liquid chromatography—Atmospheric pressure chemical ionization mass spectrometry. Eur. J. Lipid Sci. Technol. 2009, 111, 519–525. [Google Scholar] [CrossRef]
- Cvačka, J.; Krafková, E.; Jiroš, P.; Valterová, I. Computer-assisted interpretation of atmospheric pressure chemical ionization mass spectra of triacylglycerols. Rapid Commun. Mass Spectrom. 2006, 20, 3586–3594. [Google Scholar] [CrossRef]
- Stránský, K.; Jursík, T. Simple quantitative transesterification of lipids. 1. Introduction. Fett/Lipid 1996, 98, 65–71. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kofroňová, E.; Nekola, A.; Cvačka, J.; Kindl, J.; Valterová, I. Fatty Acids from Pool Lipids as Possible Precursors of the Male Marking Pheromone in Bumblebees. Molecules 2014, 19, 2330-2343. https://doi.org/10.3390/molecules19022330
Kofroňová E, Nekola A, Cvačka J, Kindl J, Valterová I. Fatty Acids from Pool Lipids as Possible Precursors of the Male Marking Pheromone in Bumblebees. Molecules. 2014; 19(2):2330-2343. https://doi.org/10.3390/molecules19022330
Chicago/Turabian StyleKofroňová, Edita, Adam Nekola, Josef Cvačka, Jiří Kindl, and Irena Valterová. 2014. "Fatty Acids from Pool Lipids as Possible Precursors of the Male Marking Pheromone in Bumblebees" Molecules 19, no. 2: 2330-2343. https://doi.org/10.3390/molecules19022330
APA StyleKofroňová, E., Nekola, A., Cvačka, J., Kindl, J., & Valterová, I. (2014). Fatty Acids from Pool Lipids as Possible Precursors of the Male Marking Pheromone in Bumblebees. Molecules, 19(2), 2330-2343. https://doi.org/10.3390/molecules19022330