A Review on Anti-Inflammatory Activity of Phenylpropanoids Found in Essential Oils

Abstract
:1. Introduction

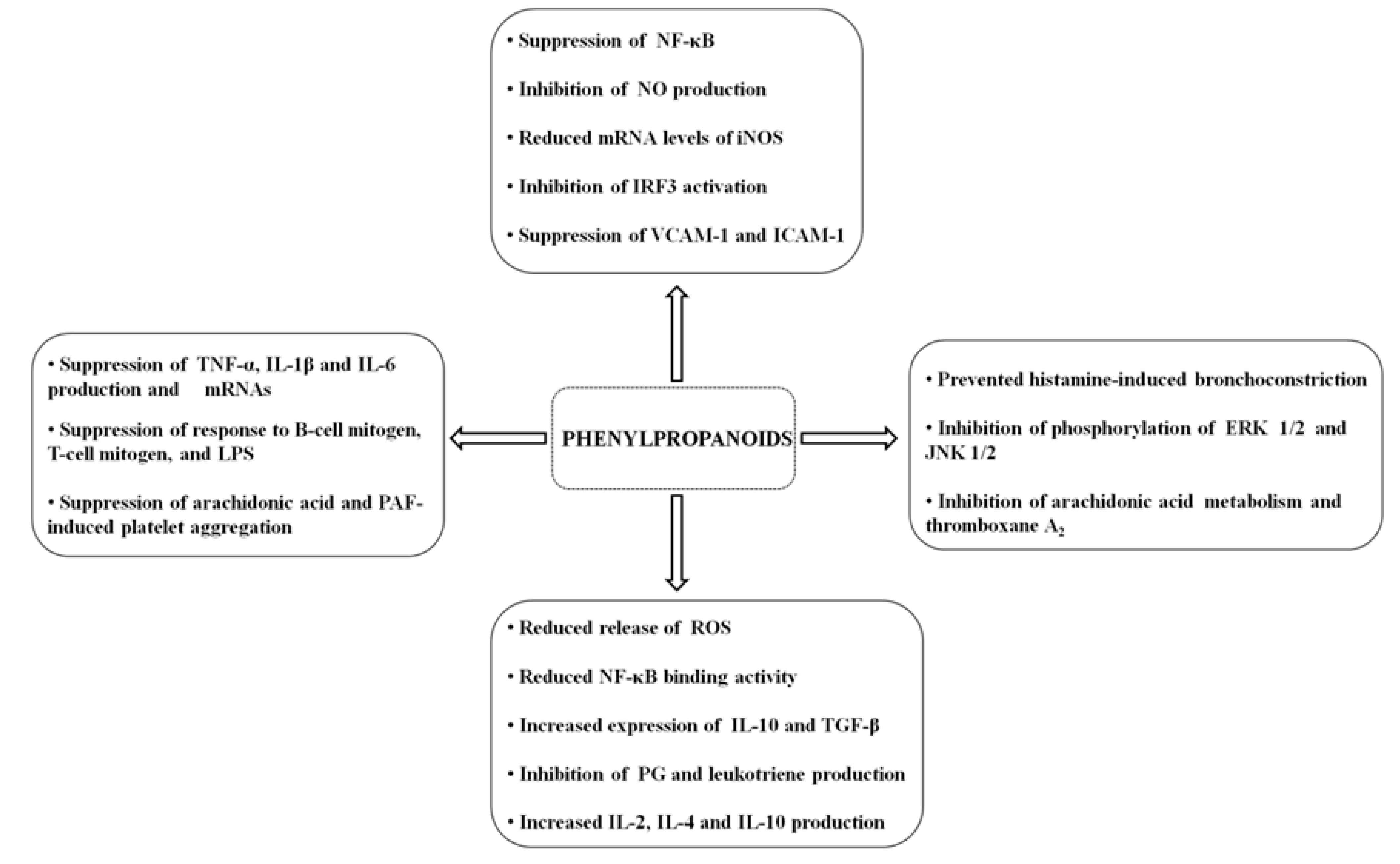
{kind=link}
{kind=link}
| Compound | Experimental protocol | Anti-inflammatory activity and/or mechanism | Animal/cell line tested | Reference |
|---|---|---|---|---|
| 2′-hydroxycinnamaldehyde (2-propenal, 3-(2-hydroxyphenyl)-cinnamaldehyde) 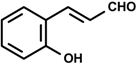 | LPS-induced NO Production | Inhibitory of NO production | RAW 264.7 macrophage | [13] |
| NF-kB-luciferase activity and NF-kB DNA binding activity | Suppression of luciferase activity | |||
| LPS-induced NF-kB activation and degradation of IkB | Inhibition of the production of nitrate, COX-2 and TNF-α | |||
| Cinnamaldehyde [(2E)-3-phenylprop-2-enal] 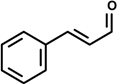 | 10 day-treatment (mixed on food) | Suppression of NF-κB activation | Fischer 344 rat | [14] |
| Inactivation of IKK | ||||
| Upregulation of IKBα and IKBβ | ||||
| Inhibition of ERK and p38MAPK pathway | ||||
| LPS-activated macrophage | Inhibition of NO and PGE2 production | RAW 264.7 macrophage | [15] | |
| LPS- or LTA-stimulated macrophage | Inhibition of IL-1β and TNF-α | Murine J774A-1 macrophage | [16] | |
| Suppression of pro-IL-1β production | Murine J774A-1 macrophage | |||
| Reduced release of ROS | ||||
| Inhibition of phosphorylation of ERK 1/2 and JNK 1/2 | ||||
| OVA-sensitization | Increased IL-2, IL-4 and IL-10 production | Balb/c mice | [17] | |
| Cytokine-induced monocyte/human endothelial cell interaction | Suppression of VCAM-1 and ICAM-1 | Human endothelial cell | [18] | |
| Suppression of NF-κB | ||||
| LPS-activated BV2 microglia | Inhibition of NO production | BV2 microglia | [19] | |
| Decreases the expression of IL-1β, IL-6, and TNF-α | ||||
| Reduced mRNA levels of iNOS | ||||
| LPS-activated macrophage | Inhibiton of NF-κB and IRF3 activation | RAW 264.7 macrophage | [20] | |
| IL-1β-activation of COX-2 and PGE2 | Reduction of COX-2 activity and PGE2 production | Rat cerebral microvascular endothelial cell | [21] | |
| LPS-induction of NO production | Reduction of NO release | J774 macrophage | [22] | |
| LPS-induction of COX-2 expression | Suppression of COX-2 expression | |||
| Carrageenan-induced paw edema | Reduction of paw edema | Rat | ||
Cinnamyl acetate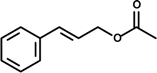 | LPS-activated macrophage | Inhibition of NO and PGE2 production | RAW 264.7 macrophage | [15] |
Cinnamic acid [(E)-3-phenylprop-2-enoic acid]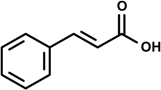 | LPS-induction NO production | Inhibition of NO production | RAW 264.7 macrophage | [23] |
Safrole (4-Allyl-1,2-methylenedioxybenzene)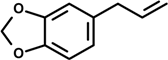 | 14 day-oral treatment | LPS inhibition of spleen cell suspension | Balb/c mice | [24] |
| Suppression of response to B-cell mitogen, T-cell mitogen, and LPS | ||||
| Splenocyte culture | Inhibition of response to LPS | |||
Dillapiole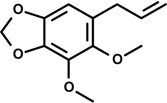 | Carrageenan-induced paw edema | Inhibition of paw edema | Rat | [25] |
Dihydrodillapiole | ||||
Eugenol (4-allyl-2-methoxyphenol)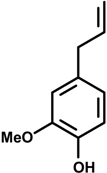 | Croton oil-induced edema | Inhibition of edema | Mice | [26] |
| Tooth pulp microsomes and homogenates | Inhibition of PG and leukotriene production | Rat tooth pulp homogenates | [27] | |
| Leukocyte | Human leukocytes | |||
| Kidney medulla | Rabbit and bovine kidney medulla | |||
| IL-1β-stimulated gingival fibroblast | Proinflammatory mediator in gingivitis | Gingival fibroblasts | [28] | |
| Platelet aggregation | Suppression of AA and PAF-induced platelet aggregation | Human platelet | [29] | |
| In vivo effects on AA and PAF-induced shock | Prevented the lethal effects of intravenous PAF or AA in a dose-dependent fashion | Rabbits | ||
| Carrageenan-induced paw edema | Reduction of edema | Rat | ||
| Platelet aggregation induced by AA | Inhibition of AA metabolism and thromboxane A2 | Rat | [30] | |
Methyleugenol (4-allyl-1,2-dimethoxybenzene)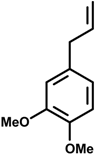 | Cerebral ischemic model | Inhibition of NO and iNOS production | Rat | [31] |
| Down-regulation of IL-1β, IL-6 and TNF-α | ||||
| Increased expression of IL-10 and TGF-β | ||||
Myristicin (5-allyl-3-methoxy-1,2-methylenedioxybenzene) | Double-stranded RNA-stimulated macrophage | Suppression of NO, IL-6, IL-10, GM-CSF, MIP-1α, MIP-1β, LIF and calcium production | RAW 264.7 macrophage | [32] |
Elimicin (5-allyl-1,2,3-trimethoxybenzene) | Basophil leukemia cells | Inhibition of 5-LOX | Rat | [33] |
Asarone (1,2,4-trimethoxy-5-[(E)-prop-1-enyl]benzene)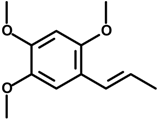 | COX- enzymes inhibitory assay | Inhibition of COX-I and COX-II enzymes | Rat | [34] |
Anethole [1-methoxy-4-(1-propenyl)benzene]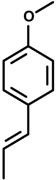 | Carrageenan and persistent inflammation induced by Complete Freund’s Adjuvant | Suppressed paw edema | Mice | [35] |
| Ear edema induced by croton oil | Inhibited the activity of myeloperoxidase | Mouse | [36] | |
| Carrageenan-induced pleurisy | Inhibiting the levels of NO and PGE2 | Rat | ||
| LPS-induced acute lung injury | Reduced total protein concentrations | BALB/c mice | [37] | |
| Reduced numbers of inflammatory cells, such as neutrophils and macrophages | ||||
| Reduced the production of inflammatory mediators matrix metalloproteinase-9 (MMP-9), TNF-α and NO | ||||
Phenethyl alcohol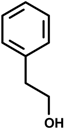 | In vitro on the contraction of isolated tracheal smooth muscle | Prevented histamine-induced bronchoconstriction | Tracheal smooth muscle in guinea pig | [38] |
2. Results and Discussion
2.1. Cinnamaldehyde (trans-Cinnamaldehyde, 2′-hydroxycinnamaldehyde) and Cinnamic Acid
2.2. Safrole, Dillapide, Dihydrodillapide, Eugenol and Methyleugenol
2.3. Myristicin, Elemicin, Asarone, Anethole and Phenethyl Alcohol
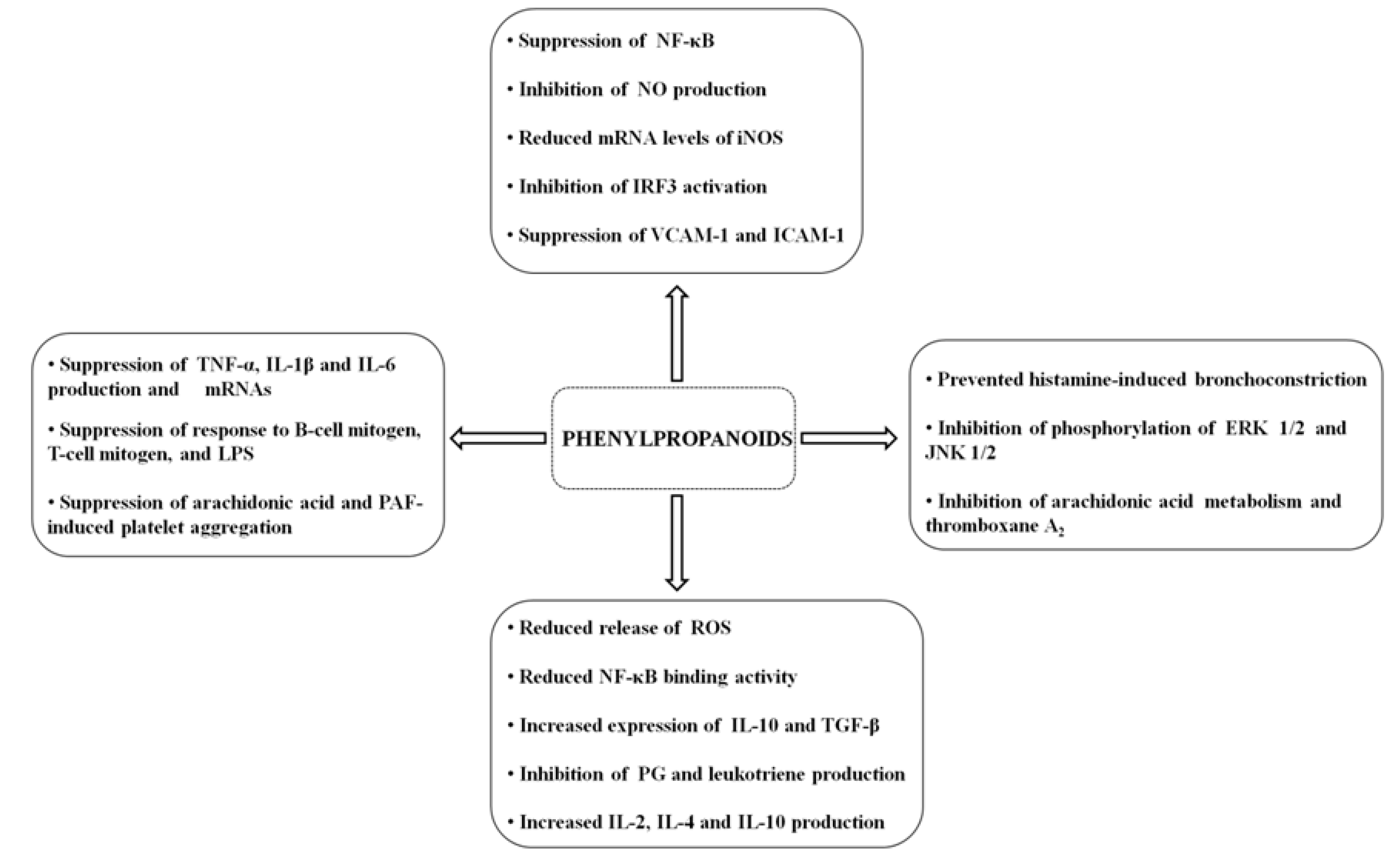
3. Conclusions
Abbreviations
| AA | Arachidonic acid |
| c-AMP | Cyclic adenosine monophosphate |
| COX-1 | Cyclo-oxygenase-1 |
| COX-2 | Cyclo-oxygenase-2 |
| ERK | Extracellular signal-regulated kinases |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| IC | Inhibitory concentration |
| ICAM-1 | Intercellular Adhesion Molecule 1 |
| IFN-γ | Interferon gama |
| IL1-β | Interleukin 1β |
| IL-6 | Interleukin 6 |
| IL-12 | Interleukin 12 |
| IRF3 | Interferon regulatory transcription factor 3 |
| LPS | Gram negative bacteria lipopolysaccharide |
| LOX | Lipoxygenase |
| LTB4 | Leukotriene B4 |
| LTC4 | Leukotriene C4 |
| LTD4 | Leukotriene D4 |
| LTE4 | Leukotriene E4 |
| MAPK | Mitogen-activated protein kinases |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MCP-3 | Monocyte chemoattractant protein-3 |
| NF-Κb | fator nuclear kappa B |
| NO | Nitric oxide |
| NOS | Nitric oxide synthase |
| mRNA | Messenger RNA |
| PAF | Platelet-activating factor |
| PGE2 | Prostaglandin E2 |
| ROS | Reactive oxygen species |
| TNF-α | Tumor necrosis factor-alpha |
| TGF-β | Transforming growth factor-beta |
Acknowledgments
Conflicts of Interest
References
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- De Sousa, D.P. Medicinal Essential Oils: Chemical, Pharmacological and Therapeutic Aspects, 1st ed.; Nova Science Publishers: New York, NY, USA, 2012; p. 236. [Google Scholar]
- Hahlbrock, K.; Scheel, D. Physiology and molecular biology of phenylpropanoid metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 347–369. [Google Scholar] [CrossRef]
- Friedrich, H. Phenylpropanoid constituents of essential oils. Lloydia 1976, 39, 1–7. [Google Scholar]
- Ferrero-Miliani, L.; Nielsen, O.H.; Andersen, P.S.; Girardin, S.E. Chronic inflammation: Importance of NOD2 and NALP3 in interleukin-1beta generation. Clin. Exp. Immunol. 2007, 147, 227–235. [Google Scholar]
- Abbas, A.B.; Lichtman, A.H. Innate Immunity. In Basic Immunology, Functions and Disorders of the Immune System, 3rd ed.; Abbas, A.B., Lichtman, A.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Barton, G.M. A calculated response: Control of inflammation by the innate immune system. J. Clin. Investig. 2008, 118, 413–420. [Google Scholar] [CrossRef]
- Saitoh, S.; Akashi, S.; Yamada, T.; Tanimura, N.; Kobayashi, M.; Konno, K. Lipid A antagonist, lipid IVa, is distinct from lipid A in interaction with Toll-like receptor 4 (TLR4)-MD-2 and ligand-induced TLR4 oligomerization. Int. Immunol. 2004, 16, 961–969. [Google Scholar] [CrossRef]
- Hopkins, S.J. The pathophysiological role of cytokines. Leg. Med. 2003, 5, S45–S57. [Google Scholar] [CrossRef]
- Vandekerckhove, F.; Opdenakker, G.; Vanranst, M.; Lenaerts, J.P.; Put, W.; Billiau, A.; van Damme, J. Bradykinin induces interleukin-6 and synergizes with interleukin-1. Lymphokine Cytokine Res. 1991, 10, 285–289. [Google Scholar]
- Sá, R.C.S.; Andrade, L.N.; de Sousa, D.P. A review on anti-inflammatory activity of monoterpenes. Molecules 2013, 18, 1227–1254. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, S.Y.; Son, D.J.; Lee, H.; Yoo, H.S.; Song, S.; Oh, K.W.; Han, D.C.; Kwon, B.M.; Hong, J.T. Inhibitiory effect of 2′-hydroxycinnamaldehyde on nitric oxide production through inhibition of NF-kappaB activation in RAW 264.7 cells. Biochem. Pharmacol. 2005, 69, 791–799. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, C.H.; Kim, M.S.; Kim, J.Y.; Jung, K.J.; Chung, J.H.; An, W.G.; Lee, J.W.; Yu, B.P.; Chung, H.Y. Suppression of age-related inflammatory NF-κB activation by cinnamaldehyde. Biogerontology 2007, 8, 545–554. [Google Scholar] [CrossRef]
- Tung, Y.T.; Chua, M.T.; Wang, S.Y.; Chang, S.T. Anti-inflammation activities of essential oil and its constituents from indigenous cinnamon (Cinnamomum osmophloeum). Bioresour. Technol. 2008, 99, 3908–3912. [Google Scholar] [CrossRef]
- Chao, L.K.; Hua, K.F.; Hsu, H.Y.; Cheng, S.S.; Lin, I.F.; Chen, C.J.; Chen, S.T.; Chang, S.T. Cinnamaldehyde inhibits pro-inflammatory cytokines secretion from monocytes/macrophages through suppression of intracellular signaling. Food Chem. Toxicol. 2008, 46, 220–231. [Google Scholar] [CrossRef]
- Lin, S.S.C.; Lu, T.M.; Chao, P.C.; Lai, Y.Y.; Tsai, H.T.; Chen, C.S.; Lee, Y.P.; Chen, S.C.; Chou, M.C.; Yang, C.C. In vivo cytokine modulatory effects of cinnamaldehyde, the major constituent of leaf essential oil from Cinnamomum osmophloeum kaneh. Phytother. Res. 2011, 25, 1511–1518. [Google Scholar] [CrossRef]
- Yu, A.L.; Lu, C.Y.; Wang, T.S.; Tsai, C.W.; Liu, K.L.; Cheng, Y.P.; Chang, H.C.; Lii, C.K.; Chen, H.W. Induction of heme oxygenase 1 and inhibition of tumor necrosis factor alpha-induced intercellular adhesion molecule expression by andrographolide in EA.hy926 cells. J. Agric. Food Chem. 2010, 58, 7641–7648. [Google Scholar] [CrossRef]
- Ho, S.C.; Chang, K.S.; Chang, P.W. Inhibition of neuroinflammation by cinnamon and its main components. Food Chem. 2013, 138, 2275–2282. [Google Scholar] [CrossRef]
- Youn, H.S.; Lee, J.K.; Choi, Y.J.; Saitoh, S.I.; Miyake, K.; Hwang, D.H.; Lee, J.Y. Cinnamaldehyde suppresses toll-like receptor 4 activation mediated through the inhibition of receptor oligomerization. Biochem. Pharmacol. 2008, 75, 494–502. [Google Scholar] [CrossRef]
- Guo, J.Y.; Huo, H.R.; Zhao, B.S.; Liu, H.B.; Li, L.F.; Ma, Y.Y.; Guo, S.Y.; Jiang, T.L. Cinnamaldehyde reduces IL-1β-induced cyclooxygenase-2 activity in rat cerebral microvascular endothelial cells. Eur. J. Pharmacol. 2006, 537, 174–180. [Google Scholar] [CrossRef]
- Ballabeni, V.; Tognolini, M.; Giorgio, C.; Bertoni, S.; Bruni, R.; Barocelli, E. Ocotea quixos Lam. essential oil: In vitro and in vivo investigation on its anti-inflammatory properties. Fitoterapia 2010, 81, 289–295. [Google Scholar] [CrossRef]
- Kong, Y.H.; Lee, Y.C.; Choi, S.Y. Neuroprotective and anti-inflammatory effects of phenolic compounds in Panax ginseng CA Meyer. J. Ginseng. Res. 2009, 33, 111–114. [Google Scholar] [CrossRef]
- Kim, B.S.; Jeong, T.C.; Choe, S.Y.; Yang, K.H. Immunosuppressive effects of safrole in Balb/C mice. Korean J. Toxicol. 1993, 8, 191–203. [Google Scholar]
- Parise-Filho, R.; Pastrello, M.; Camerlingo, C.E.P.; Silva, G.J.; Agostinho, L.A.; de Souza, T.; Magri, F.M.M.; Ribeiro, R.R.; Brandt, C.A.; Polli, M.C. The anti-inflammatory activity of dillapiole and some semisynthetic analogues. Pharm. Biol. 2011, 49, 1173–1179. [Google Scholar] [CrossRef]
- Dohi, T.; Terada, H.; Anamura, S.; Okamoto, H.; Tsujimoto, A. The anti-inflammatory effects of phenolic dental medicaments as determined by mouse ear edema assay. Jpn. J. Pharmacol. 1989, 49, 535–539. [Google Scholar] [CrossRef]
- Anamura, S. Effects of phenolic dental medications on arachidonic acid metabolism and their anti-inflammatory action. Hiroshima Daigaku Shigaku Zasshi 1989, 21, 147–162. [Google Scholar]
- Koh, T.; Murakami, Y.; Tanaka, S.; Machino, M.; Sakagami, H. Re-evaluation of anti-inflammatory potential of eugenol in IL-1β-stimulated gingival fibroblast and pulp cells. In Vivo 2013, 27, 269–273. [Google Scholar]
- Saeed, S.A.; Simjee, R.U.; Shamim, G.; Gllani, A.H. Eugenol: A dual inhibitor of platelet-activating factor and arachidonic acid metabolism. Phytomedicine 1995, 2, 23–28. [Google Scholar] [CrossRef]
- Chen, S.J.; Wang, M.H.; Chen, I.J. Antiplatelet and calcium inhibitory properties of eugenol and sodium eugenol acetate. Gen. Pharmacol. 1996, 27, 629–633. [Google Scholar] [CrossRef]
- Choi, Y.K.; Cho, G.S.; Hwang, S.; Kim, B.W.; Lim, J.H.; Lee, J.C.; Kim, H.C.; Kim, W.K.; Kim, Y.S. Methyleugenol reduces cerebral ischemic injury by suppression of oxidative injury and inflammation. Free Radic. Res. 2010, 44, 925–935. [Google Scholar] [CrossRef]
- Lee, J.Y.; Park, W. Anti-inflammatory effect of myristicin on RAW 264.7 macrophages stimulated with polyinosinic-polycytidylic acid. Molecules 2011, 16, 7132–7142. [Google Scholar] [CrossRef]
- Hashimoto, K.; Yanagisawa, T.; Okui, Y.; Ikeya, Y.; Maruno, M.; Fujita, T. Studies on anti-allergic components in the roots of Asiasarum sieboldi. Planta Med. 1994, 60, 124–127. [Google Scholar] [CrossRef]
- Momin, R.A.; de Witt, D.L.; Nair, M.G. Inhibition of cyclooxygenase (COX) enzymes by compounds from Daucus carota L. Seeds. Phytother. Res. 2003, 17, 976–979. [Google Scholar] [CrossRef]
- Ritter, A.M.V.; Domiciano, T.P.; Verri, W.A.J.; Zarpelon, A.C.; da Silva, L.G.; Barbosa, C.P.; Natali, M.R.M.; Cuman, R.K.N.; Bersani-Amado, C.A. Antihypernociceptive activity of anethole in experimental inflammatory pain. Inflammopharmacology 2013, 21, 187–197. [Google Scholar] [CrossRef]
- Domiciano, T.P.; Dalalio, M.M.O.; Silva, E.L.; Ritter, A.M.V.; Estevão-Silva, C.F.; Ramos, F.; Caparroz-Assef, S.M.; Cuman, R.K.N.; Bersani-Amado, C.A. Inhibitory effect of anethole in nonimmune acute inflammation. Naunyn Schmiedebergs Arch. Pharmacol. 2013, 386, 331–338. [Google Scholar] [CrossRef]
- Kang, P.; Kim, K.Y.; Lee, H.S.; Min, S.S.; Seol, G.H. Anti-inflammatory effects of anethole in lipopolysaccharide-induced acute lung injury in mice. Life Sci. 2013, 93, 955–961. [Google Scholar] [CrossRef]
- Chi, T.; Ji, X.; Xia, M.; Rong, Y.; Qiu, F.; Zou, L. Effect of six extractions from Wuhu decoction on isolated tracheal smoothe muscle in Guinea pig. Zhongguo Shiyan Fangjixue Zazhi 2009, 15, 52–55. [Google Scholar]
- Yu, B.P. Aging and oxidative stress: Modulation by dietary restriction. Free Radic. Biol. Med. 1996, 21, 651–668. [Google Scholar] [CrossRef]
- Shishodia, S.; Aggarwal, B.B. Cyclooxygenase (COX)-2 inhibitor celecoxib abrogates activation of cigarette smoke-induced nuclear factor (NF)-kappaB by suppressing activation of IkappaBalpha kinase in human non-small cell lung carcinoma: Correlation with suppression of cyclin D1, COX-2, and matrix metalloproteinase-9. Cancer Res. 2004, 64, 5004–5012. [Google Scholar] [CrossRef]
- Chung, H.Y.; Sung, B.; Jung, K.J.; Zou, Y.; Yu, B.P. The molecular inflammatory process in aging. Antioxid. Redox Signal. 2006, 8, 572–581. [Google Scholar] [CrossRef]
- Chang, S.T.; Chen, P.F.; Chang, S.C. Antibacterial activity of leaf essential oils and components from Cinnamomum osmophloeum. J. Ethnopharmacol. 2001, 77, 123–127. [Google Scholar] [CrossRef]
- Cheng, S.S.; Liu, J.Y.; Hsui, Y.R.; Chang, S.T. Chemical polymorphism and antifungal activity of essential oils from leaves of different provenances of indigenous cinnamom (Cinnamomum osmophloeum). Bioresour. Technol. 2006, 97, 306–312. [Google Scholar] [CrossRef]
- Chen, M.J.; Yang, Y.H.; Shieh, T.Y. Evaluation of a self-rating screening test for areca quid abusers in Taiwan. Public Health 2002, 116, 195–200. [Google Scholar]
- Hsu, H.Y.; Wen, M.H. Lipopolysaccharide-mediated reactive oxygen species and signal transduction in the regulation of interleukin-1 gene expression. J. Biol. Chem. 2002, 277, 22131–22139. [Google Scholar] [CrossRef]
- Dunn, D.E.; Herold, K.C.; Otten, G.R. Interleukin 2 and concanavalin A stimulate interferon-gamma production in a murine cytolytic cell clone by different pathways. J. Immunol. 1987, 139, 3942–3948. [Google Scholar]
- Beissert, S.; Schwartz, A.; Schwarz, T. Regulatory T cells. J. Investig. Dermatol. 2006, 126, 15–24. [Google Scholar] [CrossRef]
- Kwon, H.K.; Hwang, J.S.; So, J.S.; Lee, C.G.; Sahoo, A.; Ryu, J.H. Cinnamon extract induces tumor cell death through inhibition of NF-jB and AP1. BMC Cancer 2010, 10, 392–401. [Google Scholar] [CrossRef]
- Liao, J.C.; Deng, J.S.; Chiu, C.S.; Hou, W.C.; Huang, S.S.; Shie, P.H.; Huang, G.J. Anti-inflammatory activities of Cinnamomum cassia constituents in vitro and in vivo. Evid. Based Complement. Alternat. Med. 2012. [Google Scholar] [CrossRef]
- Iiyama, K.; Hajra, L.; Iiyama, M.; Li, H.; DiChiara, M.; Medoff, B.D.; Cybulsky, M.I. Patterns of vascular cell adhesion molecule-1 and intercellular adhesion molecule-1 expression in rabbit and mouse atherosclerotic lesions and at sites predisposed to lesion formation. Circ. Res. 1999, 85, 199–207. [Google Scholar] [CrossRef]
- Glass, C.K.; Witztum, J.L. Atherosclerosis: The road ahead. Cell 2001, 104, 503–516. [Google Scholar] [CrossRef]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Libby, P. Inflammation in atherosclerosis. Nature 2002, 420, 868–874. [Google Scholar] [CrossRef]
- Liao, B.C.; Hsieh, C.W.; Liu, Y.C.; Tzeng, T.T.; Sun, Y.W.; Wung, B.S. Cinnamaldehyde inhibits the tumor necrosis factor-α-induced expression of cell adhesion molecules in endothelial cells by suppressing NF-κB activation: Effects upon IκB and Nrf2. Toxicol.Appl. Pharmacol. 2008, 229, 161–171. [Google Scholar]
- Glass, C.K.; Saijo, K.; Winner, B.; Marchetto, M.C.; Gage, F.H. Mechanisms underlying inflammation in neurodegeneration. Cell 2010, 140, 918–934. [Google Scholar] [CrossRef]
- Collins, S.E.; Noyce, R.S.; Mossman, K.L. Innate cellular response to virus particle entry requires IRF3 but not virus replication. J. Virol. 2004, 78, 1706–1717. [Google Scholar] [CrossRef]
- Lee, J.Y.; Hwang, D.H. The modulation of inflammatory gene expression by lipids: Mediation through Toll-like receptors. Mol. Cells 2006, 21, 174–185. [Google Scholar]
- Uracz, W.; Uracz, D.; Olszanec, R.; Gryglewski, R.J. Interleukin 1beta induces functional prostaglandin E synthase in cultured human umbilical vein endothelial cells. J. Physiol.Pharmacol. 2002, 53, 643–654. [Google Scholar]
- Sehic, E.; Szekely, M.; Ungar, A.L.; Oladehin, A.; Blatteis, C.M. Hypothalamic prostaglandin E2 during lipopolysaccharide-induced fever in guinea pigs. Brain Res. Bull. 1996, 39, 391–399. [Google Scholar] [CrossRef]
- Naranjo, P.; Kijjoa, A.; Giesbrecht, A.M.; Gottlieb, O.R. Ocotea quixos, American cinnamon. J. Ethnopharmacol. 1981, 4, 233–236. [Google Scholar] [CrossRef]
- Ballabeni, V.; Tognolini, M.; Bertoni, S.; Bruni, R.; Guerrini, A.; Rueda, G.M. Antiplatelet and antithrombotic activities of essential oil from wild Ocotea quixos (Lam.) Kosterm. (Lauraceae) calices from Amazonian Ecuador. Pharmacol. Res. 2007, 55, 23–30. [Google Scholar] [CrossRef]
- Kim, B.H.; Lee, Y.G.; Lee, J.; Lee, J.Y.; Cho, J.Y. Regulatory effect of cinnamaldehyde on monocyte/macrophage-mediated inflammatory responses. Mediat. Inflamm. 2010. [Google Scholar] [CrossRef]
- Samuelsson, B. Arachidonic acid metabolism: Role in inflammation. Z. Rheumatol. 1991, 50, 3–6. [Google Scholar]
- Zhang, K.; Liu, C.; Kim, K.; Lee, S.Y.; Song, Y.; Jin, C.; Ryu, J.C.; Kim, M.; Park, J. Anti-inflammatory compounds inhibiting formation of leukotriene B4 in rat neutrophils. Environ. Mutagens Carcinog. 1994, 14, 34–42. [Google Scholar]
- Yun, T.K. Brief introduction of Panax ginseng CA Meyer. J. Korean Med. Sci. 2001, 16, S3–S5. [Google Scholar]
- Imamichi, T.; Nakamura, T.; Hayashi, K.; Kaneko, K.; Koyama, J. Different effects of cinnamicacid on the superoxide anion generation by guinea pig macrophages stimulated with a chemotactic peptide and immune complex. J. Pharmacobiodyn. 1990, 13, 344–352. [Google Scholar] [CrossRef]
- Ni, W.F.; Tsai, C.H.; Yang, S.F.; Chang, Y.C. Elevated expression of NF-κB in oral submucous fibrosis—Evidence for NF-κB induction by safrole in human buccal mucosal fibroblasts. Oral Oncol. 2007, 43, 557–562. [Google Scholar] [CrossRef]
- Pruthi, J.S. Species and Condiments, 1st ed.; National Book Trust: New Delhi, India, 1976; pp. 91–98. [Google Scholar]
- Chainy, G.B.N.; Manna, S.K.; Chaturvedi, M.M.; Aggarwal, B.B. Anethole blocks both early and late cellular responses transduced by tumor necrosis factor: Effect on NF-κB, AP-1, JNK, MAPKK and apoptosis. Oncogene 2000, 19, 2943–2950. [Google Scholar] [CrossRef]
- Rasheed, A.; Laekeman, G.; Totte, J.; Vlietinck, A.J.; Herman, A.G. Eugenol and prostaglandin biosynthesis. N. Engl. J. Med. 1984, 310, 50–51. [Google Scholar]
- Chan, A.S.L.; Pang, H.; Yip, E.C.H.; Tam, Y.K.; Wong, Y.H. Carvacrol and eugenol differentially stimulate intracellular CA2+ mobilization and mitogen-activated protein kinases in Jurkat T-cells and monocytic THP-1 cells. Planta Med. 2005, 71, 634–639. [Google Scholar] [CrossRef]
- Singer, A.L.; Koretzky, G.A. Control of T cell function by positive and negative regulators. Science 2002, 296, 1639–1640. [Google Scholar]
- Burhop, K.E.; Garcia, J.G.N.; Selig, W.M.; Lo, S.K.; Vander Zee, H.; Kaplam, J.E.; Malik, S.B. Platelet-activating factor increases lung vascular permeability to protein. Appl. Physiol. 1986, 61, 2210–2217. [Google Scholar]
- Pollock, W.K.; Armstrong, R.A.; Brydon, L.J.; Jones, R.L.; Maclntyre, D.E. Thromboxane-induced phosphatidate formation in human platelets. Biochem. J. 1984, 219, 833–842. [Google Scholar]
- Ahn, G.S.; Jang, H.U.; Jung, G.Y.; Kim, J.H.; Lee, H.G.; Lee, I.S.; Oh, S.R.; Park, S.H. Essential Oil Component Having Inhibition Activity of Leukotriene Production. Korea Patent 1020010086473, 12 September 2001. [Google Scholar]
- Luster, M.I.; Simeonova, P.P.; Gallucci, R.M.; Matheson, J.M.; Yucesoy, B. Immunotoxicology: Role of inflammation in chemical-induced hepatotoxicity. Int. J. Immunopharmacol. 2000, 22, 1143–1147. [Google Scholar] [CrossRef]
- Yogalakshmi, B.; Viswanathan, P.; Anuradha, C.V. Investigation of antioxidant, anti-inflammatory and DNA-protective properties of eugenol in thioacetamide-induced liver injury in rats. Toxicology 2010, 268, 204–212. [Google Scholar] [CrossRef]
- Ito, M.; Murakami, K.; Yoshino, M. Antioxidant action of eugenol compounds: Role of metal ion in inhibiton of lipid peroxidation. Food Chem. Toxicol. 2005, 43, 461–466. [Google Scholar] [CrossRef]
- Yano, S.; Suzuki, Y.; Yuzurihara, M.; Kase, Y.; Takeda, S.; Watanabe, S.; Aburada, M.; Miyamoto, K. Antinociceptive effect of methyeugenol on formalin-induced hyperlagesia in mice. Eur. J. Pharmacol. 2006, 553, 99–103. [Google Scholar] [CrossRef]
- Nishizawa, Y. Glutamate release and neuronal damage in ischemia. Life Sci. 2001, 69, 369–381. [Google Scholar] [CrossRef]
- Martins, C.; Doran, C.; Laires, A.; Rueff, J.; Rodrigues, A.S. Genotoxic and apoptotic activities of the food flavourings myristicin and eugenol in AA8 and XRCC1 deficient EM9 cells. Food Chem. Toxicol. 2011, 49, 385–392. [Google Scholar] [CrossRef]
- Narasimhan, B.; Dhake, A.S. Antibacterial principles from Myristica fragrans seeds. J. Med. Food 2006, 9, 395–399. [Google Scholar] [CrossRef]
- Cuschieri, J.; Maier, R.V. Oxidative stress, lipid rafts, and macrophage reprogramming. Antioxid. Redox. Signal. 2007, 9, 1485–1497. [Google Scholar] [CrossRef]
- Serhan, C.N.; Chiang, N.; van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and proresolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef]
- Ishihara, K.; Hirano, T. IL-6 in autoimmune disease and chronic inflammatory proliferative disease. Cytokine. Growth. Factor Rev. 2002, 13, 357–368. [Google Scholar] [CrossRef]
- Mallavarapu, G.R.; Ramesh, S. Composition of essential oils of nutmeg and mace. J. Med. Aromat. Plant Sci. 1998, 20, 746–748. [Google Scholar]
- Radmark, O.P. The molecular biology and regulation of 5-lipoxygenase. Am. J. Respir. Crit. Care Med. 2000, 161, S11–S15. [Google Scholar] [CrossRef]
- Albert, D.; Zündorf, I.; Dingermann, T.; Müller, W.E.; Steinhilber, D.; Werz, O. Hyperforin is a dual inhibitor of cyclooxygenase-1 and 5-lipoxygenase. Biochem. Pharmacol. 2012, 15, 1767–1775. [Google Scholar]
- De, M.; de, A.K.; Sen, P.; Banerjee, A.B. Antimicrobial properties of star anise (Illicium verum Hook f). Phytother. Res. 2002, 16, 94–95. [Google Scholar] [CrossRef]
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
De Cássia da Silveira e Sá, R.; Andrade, L.N.; Dos Reis Barreto de Oliveira, R.; De Sousa, D.P. A Review on Anti-Inflammatory Activity of Phenylpropanoids Found in Essential Oils. Molecules 2014, 19, 1459-1480. https://doi.org/10.3390/molecules19021459
De Cássia da Silveira e Sá R, Andrade LN, Dos Reis Barreto de Oliveira R, De Sousa DP. A Review on Anti-Inflammatory Activity of Phenylpropanoids Found in Essential Oils. Molecules. 2014; 19(2):1459-1480. https://doi.org/10.3390/molecules19021459
Chicago/Turabian StyleDe Cássia da Silveira e Sá, Rita, Luciana Nalone Andrade, Rafael Dos Reis Barreto de Oliveira, and Damião Pergentino De Sousa. 2014. "A Review on Anti-Inflammatory Activity of Phenylpropanoids Found in Essential Oils" Molecules 19, no. 2: 1459-1480. https://doi.org/10.3390/molecules19021459
APA StyleDe Cássia da Silveira e Sá, R., Andrade, L. N., Dos Reis Barreto de Oliveira, R., & De Sousa, D. P. (2014). A Review on Anti-Inflammatory Activity of Phenylpropanoids Found in Essential Oils. Molecules, 19(2), 1459-1480. https://doi.org/10.3390/molecules19021459