Riboflavin Accumulation and Molecular Characterization of cDNAs Encoding Bifunctional GTP Cyclohydrolase II/3,4-Dihydroxy-2-Butanone 4-Phosphate Synthase, Lumazine Synthase, and Riboflavin Synthase in Different Organs of Lycium chinense Plant



Abstract
:
1. Introduction
2. Results and Discussion
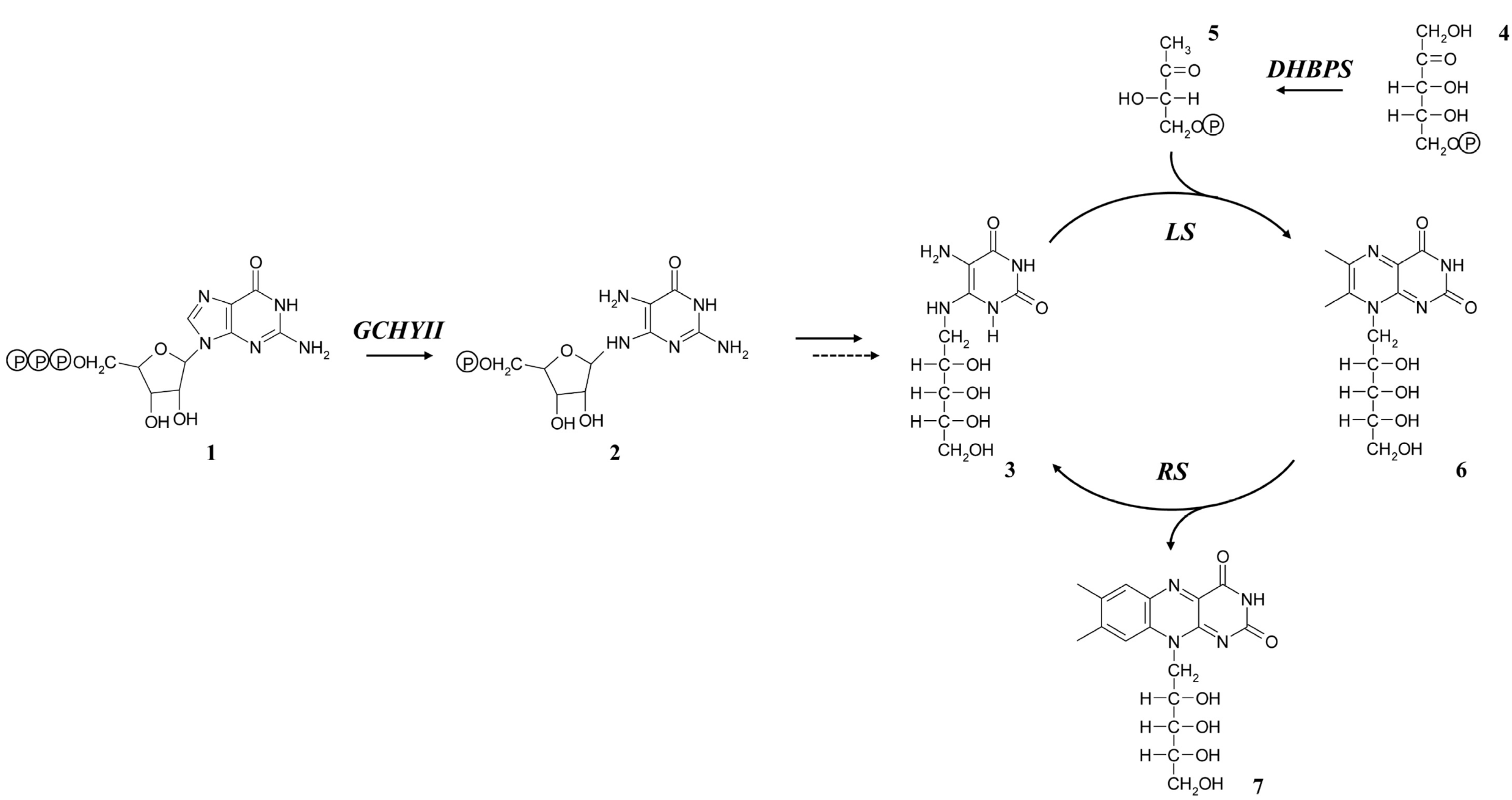
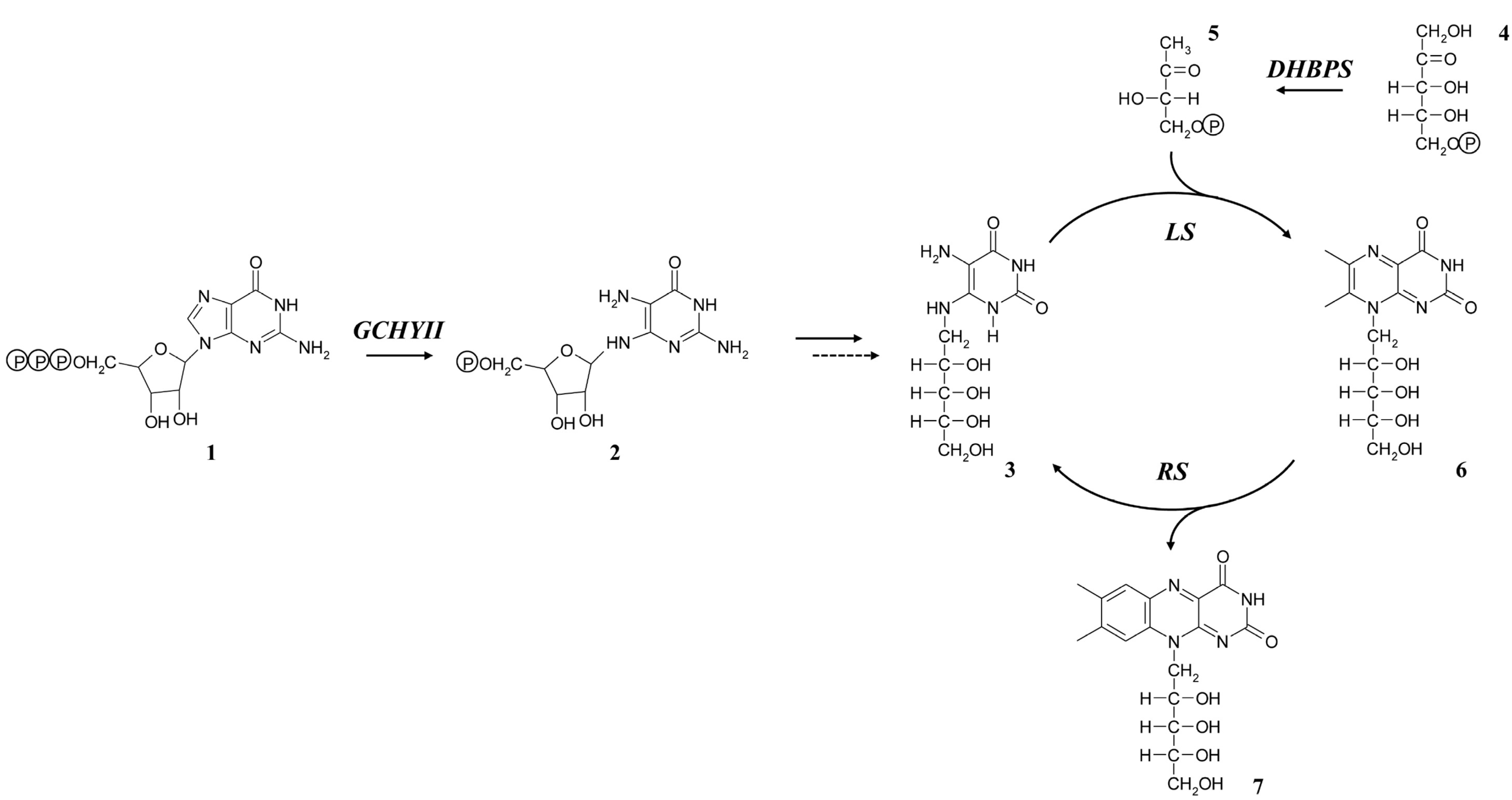
2.1. Sequence Analyses of Riboflavin Biosynthetic Genes from L. chinense



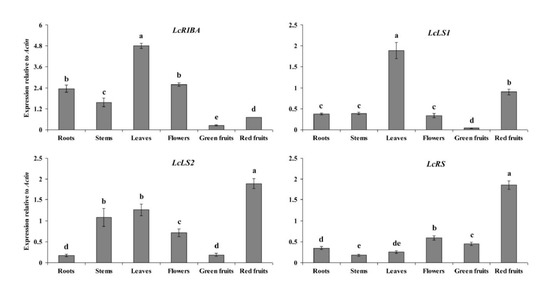
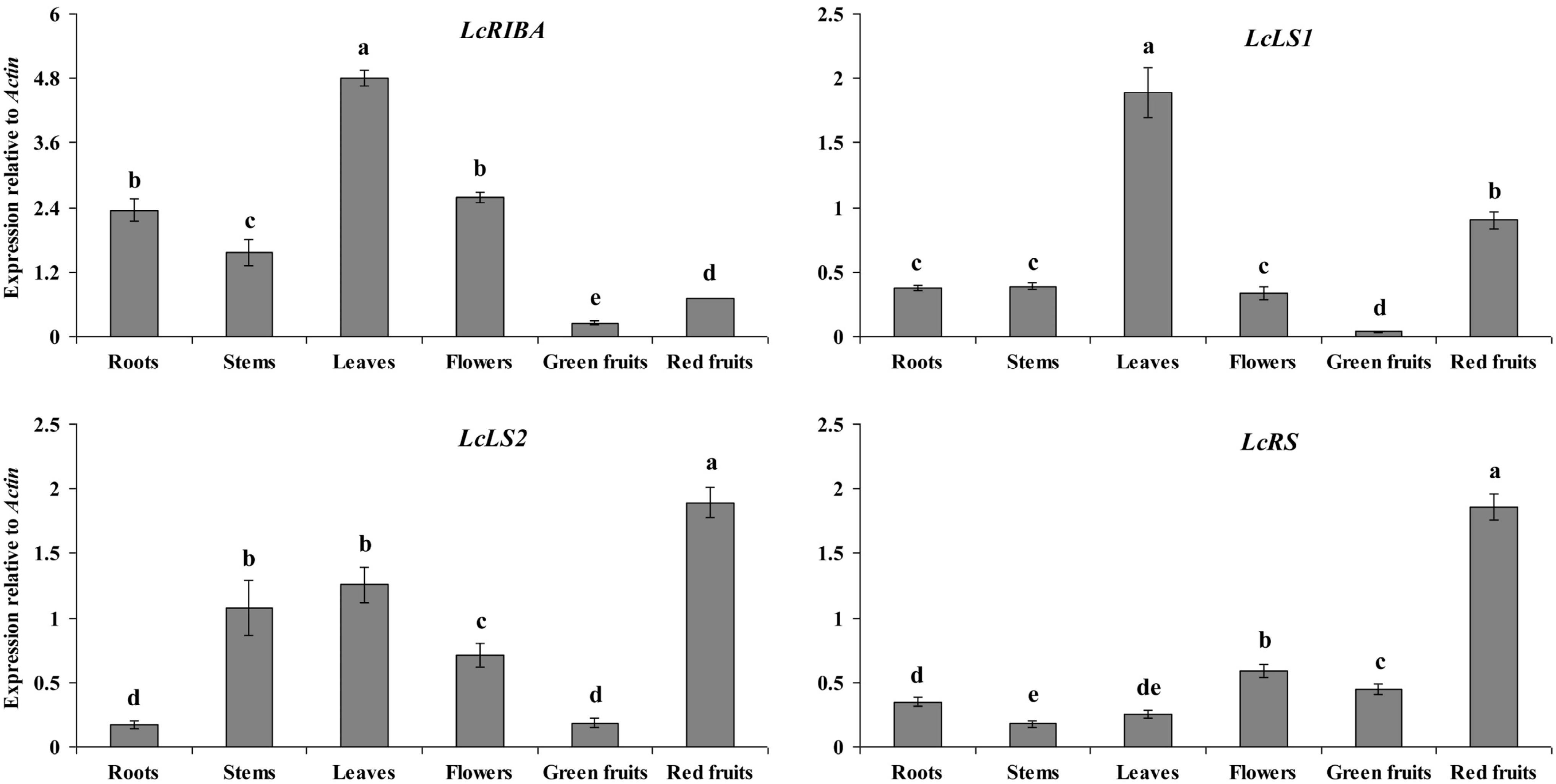
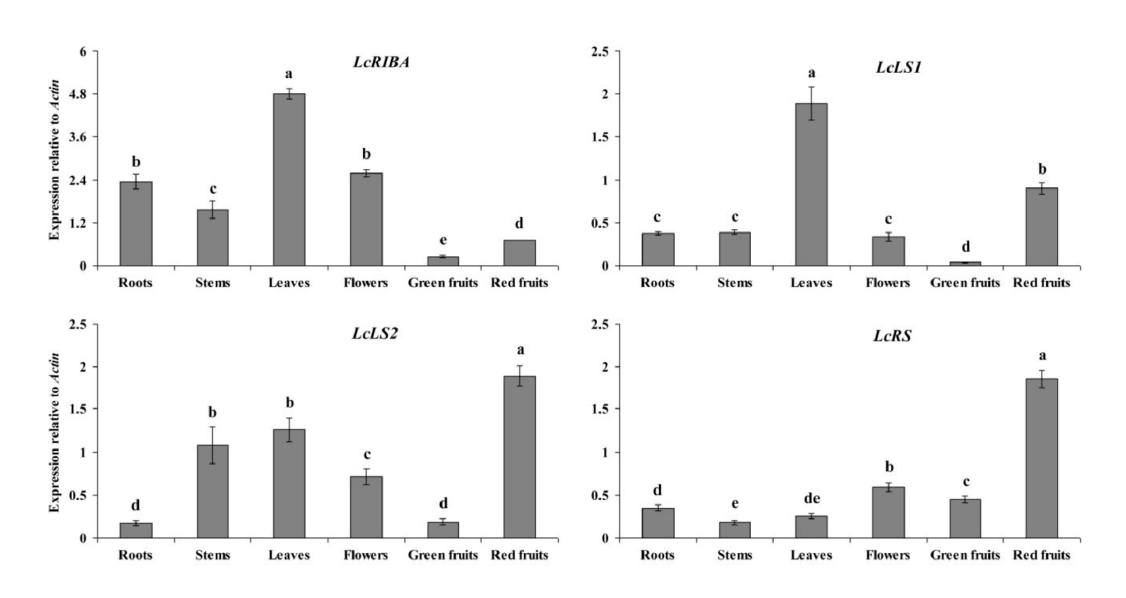
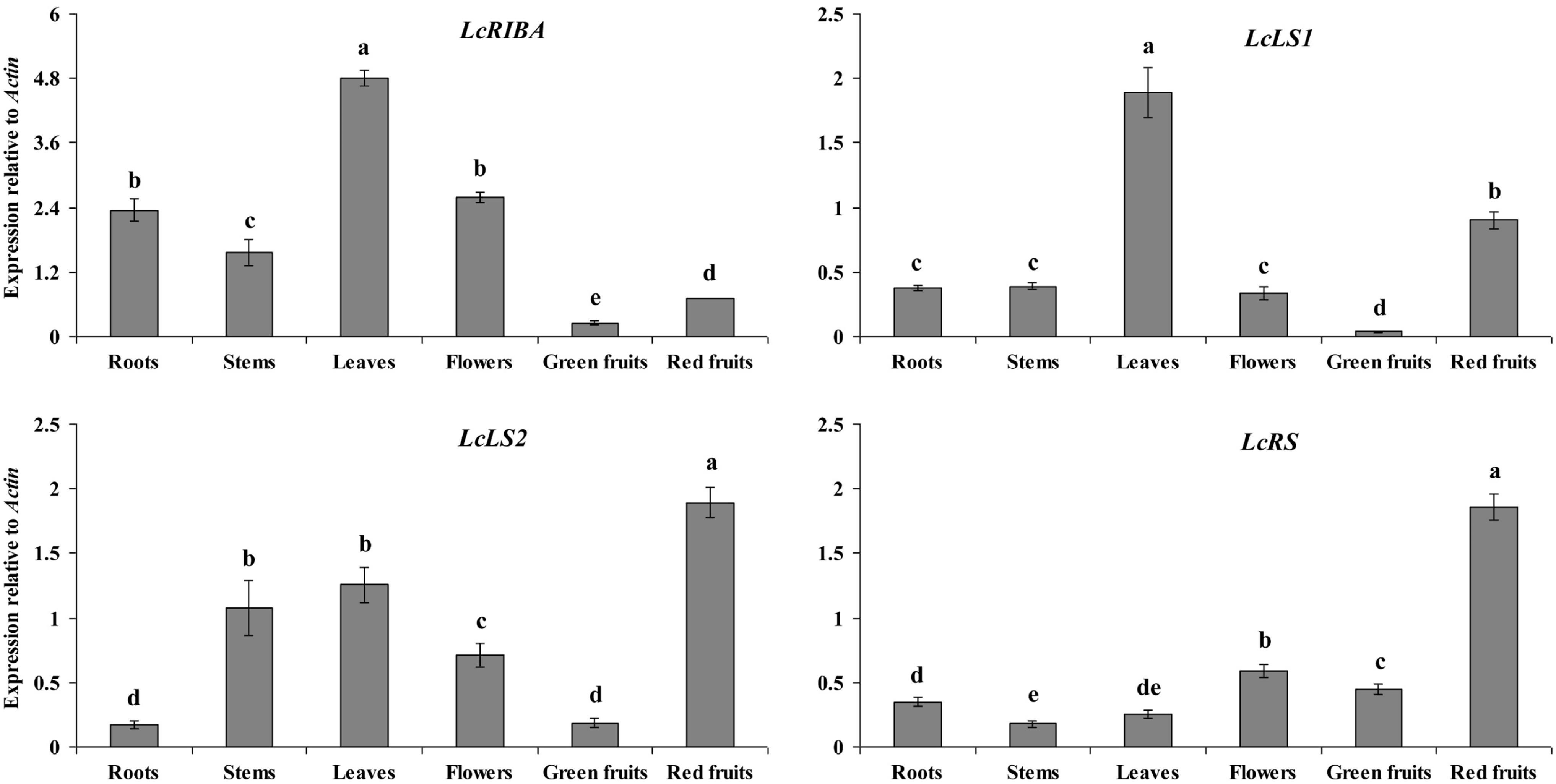
2.2. Expression Levels of LcRIBA, LcLS1, LcLS2, and LcRS in Different Organs of L. chinense
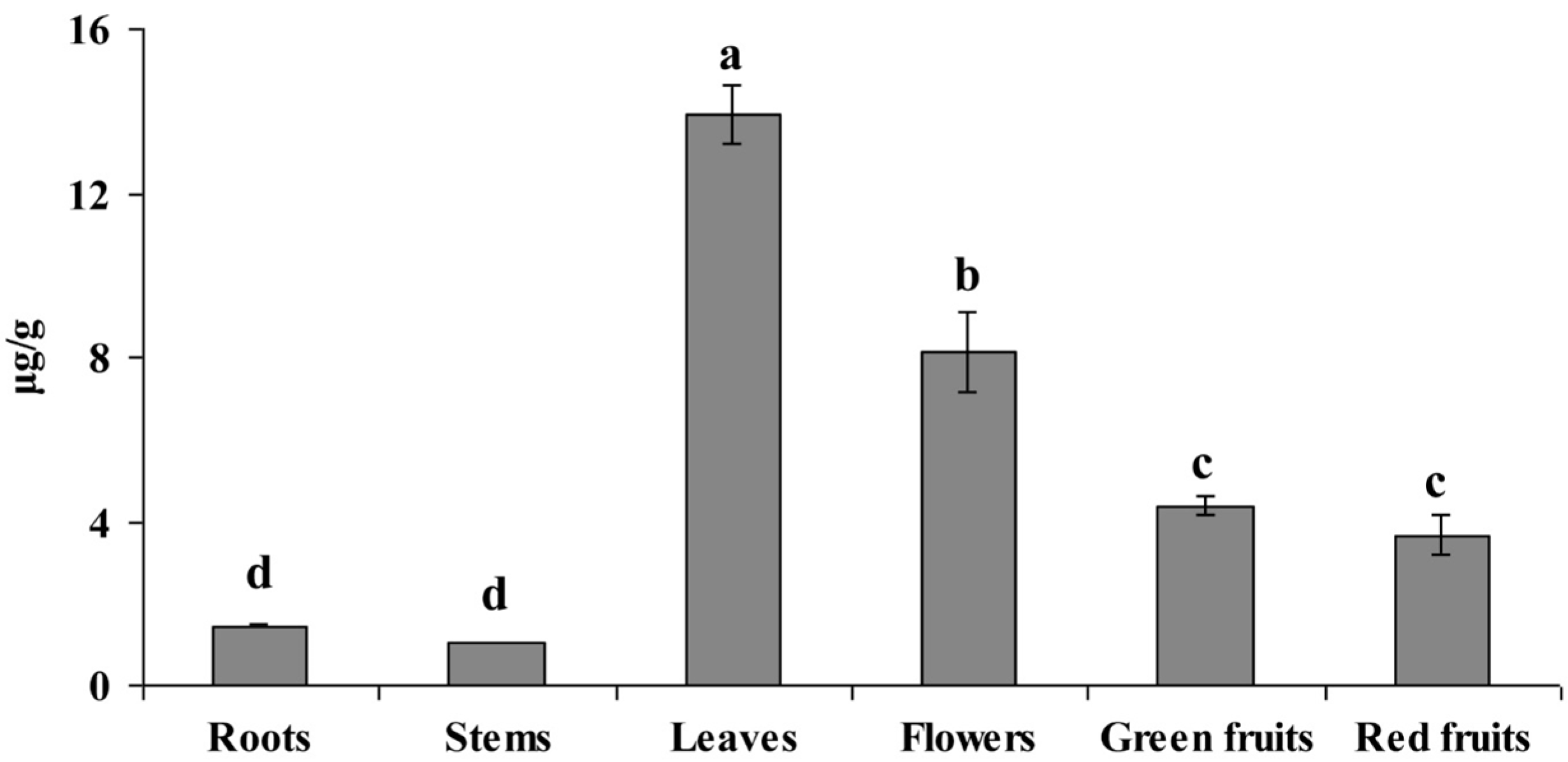
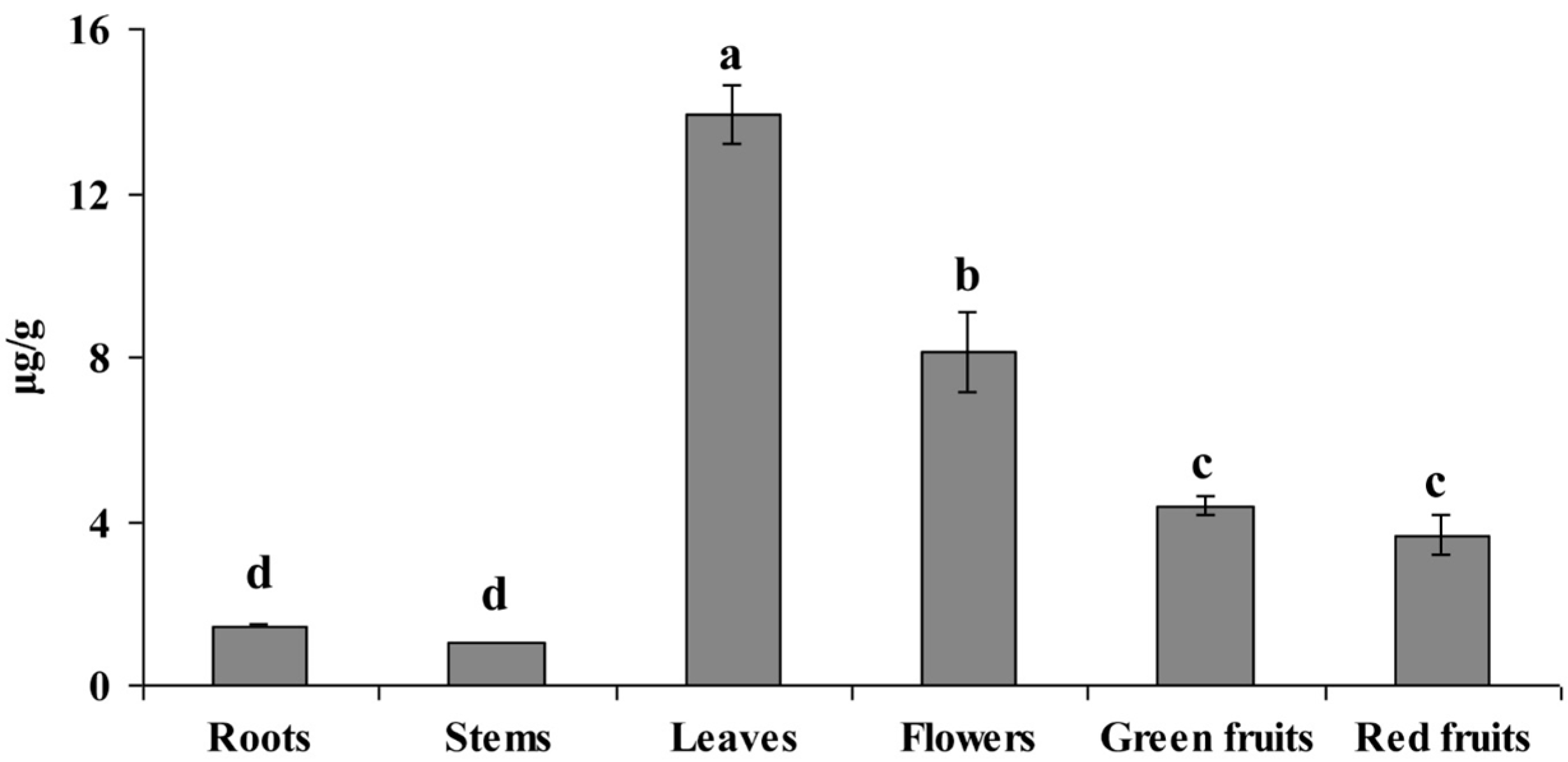
2.3. Anaysis of Riboflavin Content in Different Organs of L. chinense
3. Experimental Section
3.1. Plant Materials
3.2. RNA Isolation and cDNA Synthesis
3.3. Isolation of cDNAs Encoding Enzymes Involved in Riboflavin Biosynthetic Pathway
3.4. Sequence Analysis
3.5. Quantitative Real-Time PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5' to 3') | Amplicon (Base Pairs) |
|---|---|---|
| LcRIBA F | CTGGCTTAGACCCTGTTGGAGTAAT | 169 |
| LcRIBA R | GAAGCATGCTCTACCAACTGATCTC | |
| LcLS1 F | CAACTGTAATAAATCCTACGCAACG | 157 |
| LcLS1 R | CATGTTGTAAACCGGTTTAGATCCT | |
| LcLS2 F | CAATCCTTCACAGTTGCAACATTT | 180 |
| LcLS2 R | ACAGCTGATGTTTGAACTAAATCCC | |
| LcRS F | TTGAGCTTAAAACTGAAGGGGATTC | 165 |
| LcRS R | AAGCCACCAACATGAAGTTAAAACA | |
| LcActin F | ACCACTTGTTTGTGACAATGGAACT | 198 |
| LcActin R | TCAATTGGGTATTTCAAGGTCAAGA |
3.6. Riboflavin Extraction and Analysis
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- De Souza, A.C.; Kodach, L.; Gadelha, F.R.; Bos, C.L.; Cavagis, A.D.; Aoyama, H.; Peppelenbosch, M.P.; Ferreira, C.V. A promising action of riboflavin as a mediator of leukaemia cell death. Apoptosis 2006, 11, 1761–1771. [Google Scholar]
- Powers, H.J. Riboflavin (vitamin B-2) and health. Am. J. Clin. Nutr. 2003, 77, 1352–1360. [Google Scholar]
- Skalka, H.W.; Prchal, J.T. Cataracts and riboflavin deficiency. Am. J. Clin. Nutr. 1981, 34, 861–863. [Google Scholar]
- Massey, V. The chemical and biological versatility of riboflavin. Biochem. Soc. Trans. 2000, 28, 283–296. [Google Scholar] [CrossRef]
- Gregory, J.F.I. Nutritional properties and significance of vitamin glycosides. Annu. Rev. Nutr. 1998, 18, 277–296. [Google Scholar] [CrossRef]
- Gastaldi, G.; Laforenza, U.; Gasirola, D.; Ferrari, G.; Tosco, M.; Rindi, G. Energy depletion differently affects membrane transport nd intracellular metabolism of riboflavin taken up by isolated rat enterocytes. J. Nutr. 1999, 129, 406–409. [Google Scholar]
- Fischer, M.; Bacher, A. Biosynthesis of vitamin B2: Structure and mechanism of riboflavin synthase. Arch. Biochem. Biophys. 2008, 474, 252–265. [Google Scholar] [CrossRef]
- Jordan, D.B.; Bacot, K.O.; Carlson, T.J.; Kessel, M.; Viitanen, P.V. Plant riboflavin biosynthesis. Cloning, chloroplast localization, expression, purification, and partial characterization of spinach lumazine synthase. J. Biol. Chem. 1999, 274, 22114–22121. [Google Scholar] [CrossRef]
- Herz, S.; Eberhardt, S.; Bacher, A. Biosynthesis of riboflavin in plants. The ribA gene of Arabidopsis thaliana specifies a bifunctional GTP cyclohydrolase II/3,4-dihydroxy-2-butanone 4-phosphate synthase. Phytochemistry 2000, 53, 723–731. [Google Scholar] [CrossRef]
- Schramek, N.; Haase, I.; Fischer, M.; Bacher, A. Biosynthesis of riboflavin. Single turnover kinetic analysis of 6,7-dimethyl-8-ribityllumazine synthase. J. Am. Chem. Soc. 2003, 125, 4460–4466. [Google Scholar] [CrossRef]
- Illarionov, B.; Haase, I.; Bacher, A.; Fischer, M.; Schramek, N. Presteady state kinetic analysis of riboflavin synthase. J. Biol. Chem. 2003, 278, 47700–47706. [Google Scholar] [CrossRef]
- Fischer, M.; Bacher, A. Biosynthesis of vitamin B2 in plants. Physiol. Plant. 2006, 126, 304–318. [Google Scholar] [CrossRef]
- Tuan, P.A.; Kim, J.K.; Lee, S.; Chae, S.C.; Park, S.U. Riboflavin accumulation and characterization of cDNAs encoding lumazine synthase and riboflavin synthase in bitter melon (Momordica charantia). J. Agric. Food Chem. 2012, 60, 11980–11986. [Google Scholar] [CrossRef]
- Xiao, P.-G.; Xing, S.-T.; Wang, L.-W. Immunological aspects of Chinese medicinal plants as antiageing drugs. J. Ethnopharmacol. 1993, 38, 159–165. [Google Scholar] [CrossRef]
- Chin, Y.-W.; Lim, S.W.; Kim, S.-H.; Shin, D.-Y.; Suh, Y.-G.; Kim, Y.-B.; Kim, Y.C.; Kim, J. Hepatoprotective pyrrole derivatives of Lycium chinense fruits. Bioorg. Med. Chem. Lett. 2003, 13, 79–81. [Google Scholar] [CrossRef]
- Shin, Y.G.; Cho, K.H.; Kim, J.M.; Park, M.K.; Park, J.H. Determination of betaine in Lycium chinense fruits by liquid chromatography–electrospray ionization mass spectrometry. J. Chromatogr. A 1999, 857, 331–335. [Google Scholar] [CrossRef]
- Pottérat, O. Goji (Lycium barbarum and L. chinense): Phytochemistry, pharmacology and safety in the perspective of traditional uses and recent popularity. Planta Med. 2010, 76, 7–19. [Google Scholar] [CrossRef]
- Zhou, X.; Xu, G.; Wang, Q. Studies on the chemical constituents in the roots of Lycium chinense mill. Zhong Guo Zhong Yao Za Zhi 1996, 21, 675–676. [Google Scholar]
- Kim, H.P.; Lee, E.J.; Kim, Y.C.; Kim, J.; Kim, H.K.; Park, J.H.; Kim, S.Y.; Kim, Y.C. Zeaxanthin dipalmitate from Lycium chinense fruit reduces experimentally induced hepatic fibrosis in rats. Biol. Pharm. Bull. 2002, 25, 390–392. [Google Scholar] [CrossRef]
- Hsu, H.Y.; Yang, J.J.; Ho, Y.H.; Lin, C.C. Difference in the effects of radioprotection between aerial and root parts of Lycium chinense. J. Ethnopharmacol. 1999, 64, 101–108. [Google Scholar] [CrossRef]
- Yamada, P.; Nemoto, M.; Shigemori, H.; Yokota, S.; Isoda, H. Isolation of 5-(hydroxymethyl) furfural from Lycium chinense and its inhibitory effect on the chemical mediator release by basophilic cells. Planta Med. 2011, 77, 434–440. [Google Scholar] [CrossRef]
- Kim, H.P.; Kim, S.Y.; Lee, E.J.; Kim, Y.C.; Kim, Y.C. Zeaxanthin dipalmitate from Lycium chinense has hepatoprotective activity. Res. Commun. Mol. Pathol. Pharmacol. 1997, 97, 301–314. [Google Scholar]
- Richter, G.; Ritz, H.; Katzenmeier, G.; Volk, R.; Kohnle, A.; Lottspeich, F.; Allendorf, D.; Bacher, A. Biosynthesis of riboflavin: Cloning, sequencing, mapping, and expression of the gene coding for GTP cyclohydrolase II in Escherichia coli. J. Bacteriol. 1993, 175, 4045–4051. [Google Scholar]
- Richter, G.; Volk, R.; Krieger, C.; Lahm, H.W.; Rothlisberger, U.; Bacher, A. Biosynthesis of riboflavin: Cloning, sequencing, and expression of the gene coding for 3,4-dihydroxy-2-butanone 4-phosphate synthase of Escherichia coli. J. Bacteriol. 1992, 174, 4050–4056. [Google Scholar]
- Fischer, M.; Romisch, W.; Saller, S.; Illarionov, B.; Richter, G.; Rohdich, F.; Eisenreich, W.; Bacher, A. Evolution of vitamin B2 biosynthesis: Structural and functional similarity between pyrimidine deaminases of eubacterial and plant origin. J. Biol. Chem. 2004, 279, 36299–36308. [Google Scholar] [CrossRef]
- Gerhardt, S.; Schott, A.K.; Kairies, N.; Cushman, M.; Illarionov, B.; Eisenreich, W.; Bacher, A.; Huber, R.; Steinbacher, S.; Fischer, M. Studies on the reaction mechanism of riboflavin synthase: X-Ray crystal structure of a complex with 6- carboxyethyl-7-oxo-8-ribityllumazine. Structure 2002, 10, 1371–1381. [Google Scholar] [CrossRef]
- Roje, S. Vitamin B biosynthesis in plants. Phytochemistry 2007, 68, 1904–1921. [Google Scholar] [CrossRef]
- Fischer, M.; Haase, I.; Feicht, R.; Schramek, N.; Köhler, P.; Schieberle, P.; Bacher, A. Evolution of vitamin B2 biosynthesis: Riboflavin synthase of Arabidopsis thaliana and its inhibition by riboflavin. Biol. Chem. 2005, 386, 417–428. [Google Scholar] [CrossRef]
- Zhao, S.; Tuan, P.A.; Li, X.; Kim, Y.B.; Kim, H.; Park, C.G.; Yang, J.; Li, C.H.; Park, S.U. Identification of phenylpropanoid biosynthetic genes and phenylpropanoid accumulation by transcriptome analysis of Lycium chinense. BMC Genomics 2013, 14, 802. [Google Scholar] [CrossRef]
- Protein Molecular Weight Calculator. Available online: http://www.sciencegateway.org/tools/proteinmw.htm (accessed on 20 July 2014).
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Esteve, M.J.; Farré, R.; Frígola, A.; García-Cantabella, J.M. Simultaneous determination of thiamin and riboflavin in mushrooms by liquid chromatography. J. Agric. Food Chem. 2001, 49, 1450–1454. [Google Scholar] [CrossRef]
- Perkins, J.B.; Pero, J.G.; Sloma, A. Riboflavin Overproducing Strains of Bacteria. European Patent Application 405370, January 1991. [Google Scholar]
- Heefner, D.L.; Weaver, C.A.; Yarus, M.J.; Burdzinski, L.A. Method for Producing Riboflavin with Candida famata. U.S. Patent 5,164,303, 17 November 1992. [Google Scholar]
- Sample Availability: In general, samples of the compounds analyzed herein are unavailable from the authors due to their isolation on a small scale. They are readily analyzed using the procedures described.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuan, P.A.; Zhao, S.; Kim, J.K.; Kim, Y.B.; Yang, J.; Li, C.H.; Kim, S.-J.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.U. Riboflavin Accumulation and Molecular Characterization of cDNAs Encoding Bifunctional GTP Cyclohydrolase II/3,4-Dihydroxy-2-Butanone 4-Phosphate Synthase, Lumazine Synthase, and Riboflavin Synthase in Different Organs of Lycium chinense Plant. Molecules 2014, 19, 17141-17153. https://doi.org/10.3390/molecules191117141
Tuan PA, Zhao S, Kim JK, Kim YB, Yang J, Li CH, Kim S-J, Arasu MV, Al-Dhabi NA, Park SU. Riboflavin Accumulation and Molecular Characterization of cDNAs Encoding Bifunctional GTP Cyclohydrolase II/3,4-Dihydroxy-2-Butanone 4-Phosphate Synthase, Lumazine Synthase, and Riboflavin Synthase in Different Organs of Lycium chinense Plant. Molecules. 2014; 19(11):17141-17153. https://doi.org/10.3390/molecules191117141
Chicago/Turabian StyleTuan, Pham Anh, Shicheng Zhao, Jae Kwang Kim, Yeon Bok Kim, Jingli Yang, Cheng Hao Li, Sun-Ju Kim, Mariadhas Valan Arasu, Naif Abdullah Al-Dhabi, and Sang Un Park. 2014. "Riboflavin Accumulation and Molecular Characterization of cDNAs Encoding Bifunctional GTP Cyclohydrolase II/3,4-Dihydroxy-2-Butanone 4-Phosphate Synthase, Lumazine Synthase, and Riboflavin Synthase in Different Organs of Lycium chinense Plant" Molecules 19, no. 11: 17141-17153. https://doi.org/10.3390/molecules191117141
APA StyleTuan, P. A., Zhao, S., Kim, J. K., Kim, Y. B., Yang, J., Li, C. H., Kim, S.-J., Arasu, M. V., Al-Dhabi, N. A., & Park, S. U. (2014). Riboflavin Accumulation and Molecular Characterization of cDNAs Encoding Bifunctional GTP Cyclohydrolase II/3,4-Dihydroxy-2-Butanone 4-Phosphate Synthase, Lumazine Synthase, and Riboflavin Synthase in Different Organs of Lycium chinense Plant. Molecules, 19(11), 17141-17153. https://doi.org/10.3390/molecules191117141