Stimulation of Natural Killer T Cells by Glycolipids


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
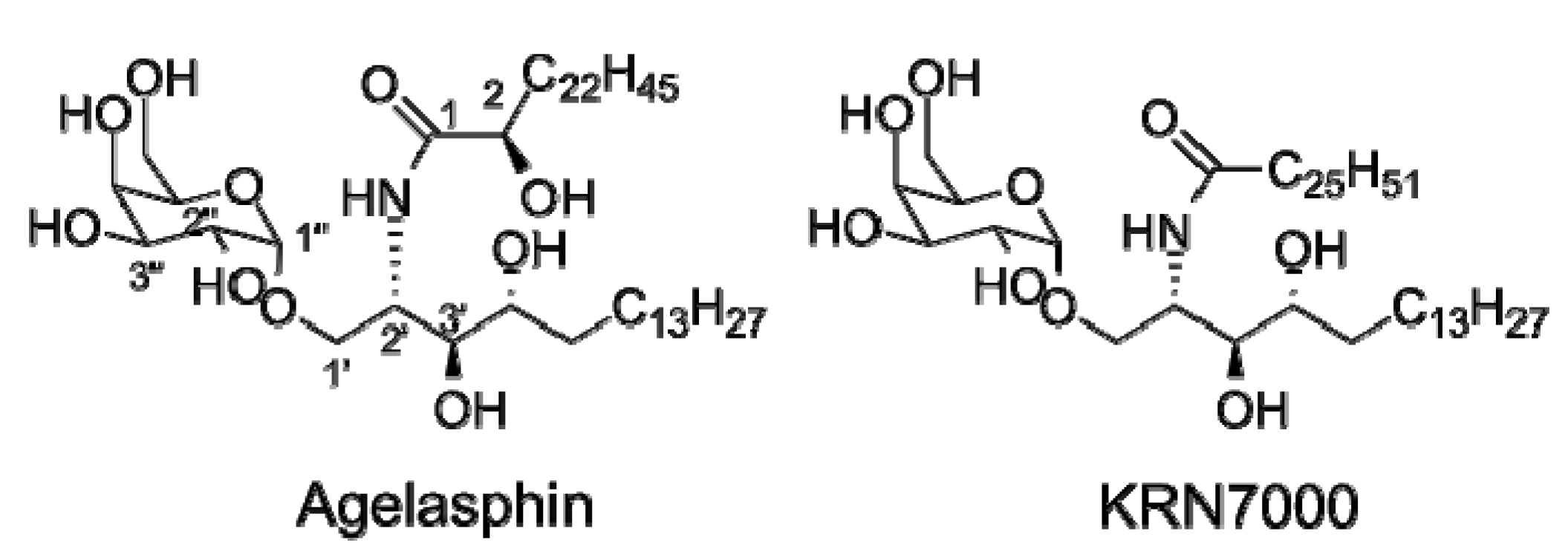
1.1. The Model iNKT Antigen: α-GalCer
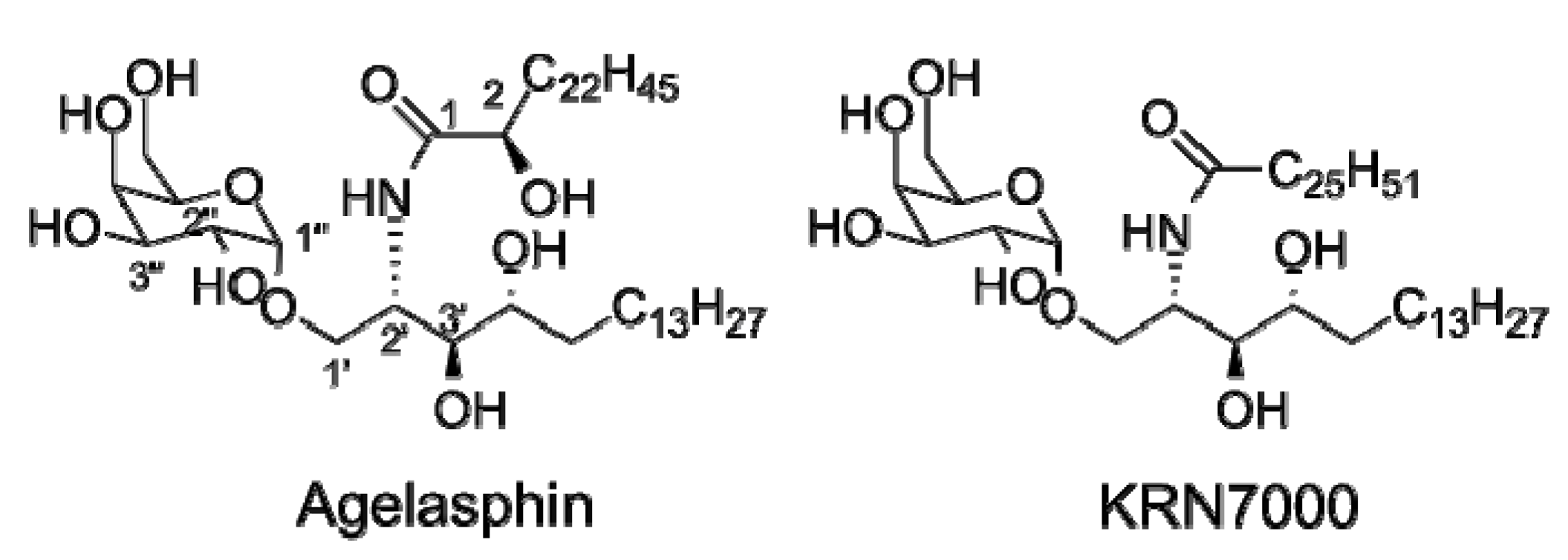
1.2. The Continuing Search for iNKT Cell Antigens
2. Exogenous Antigens for iNKT Cells
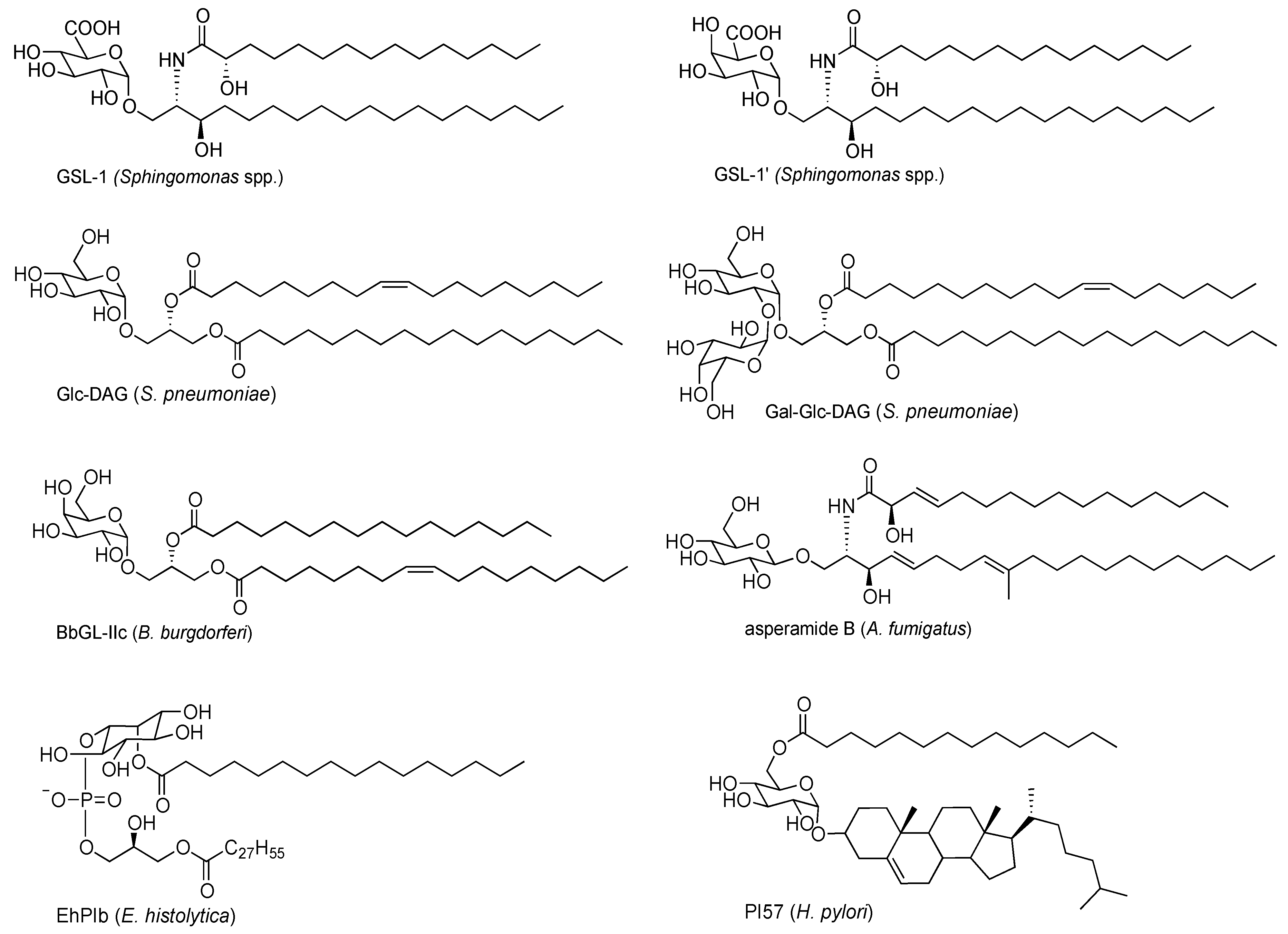
2.1. Sphingomonas Glycosphingolipids
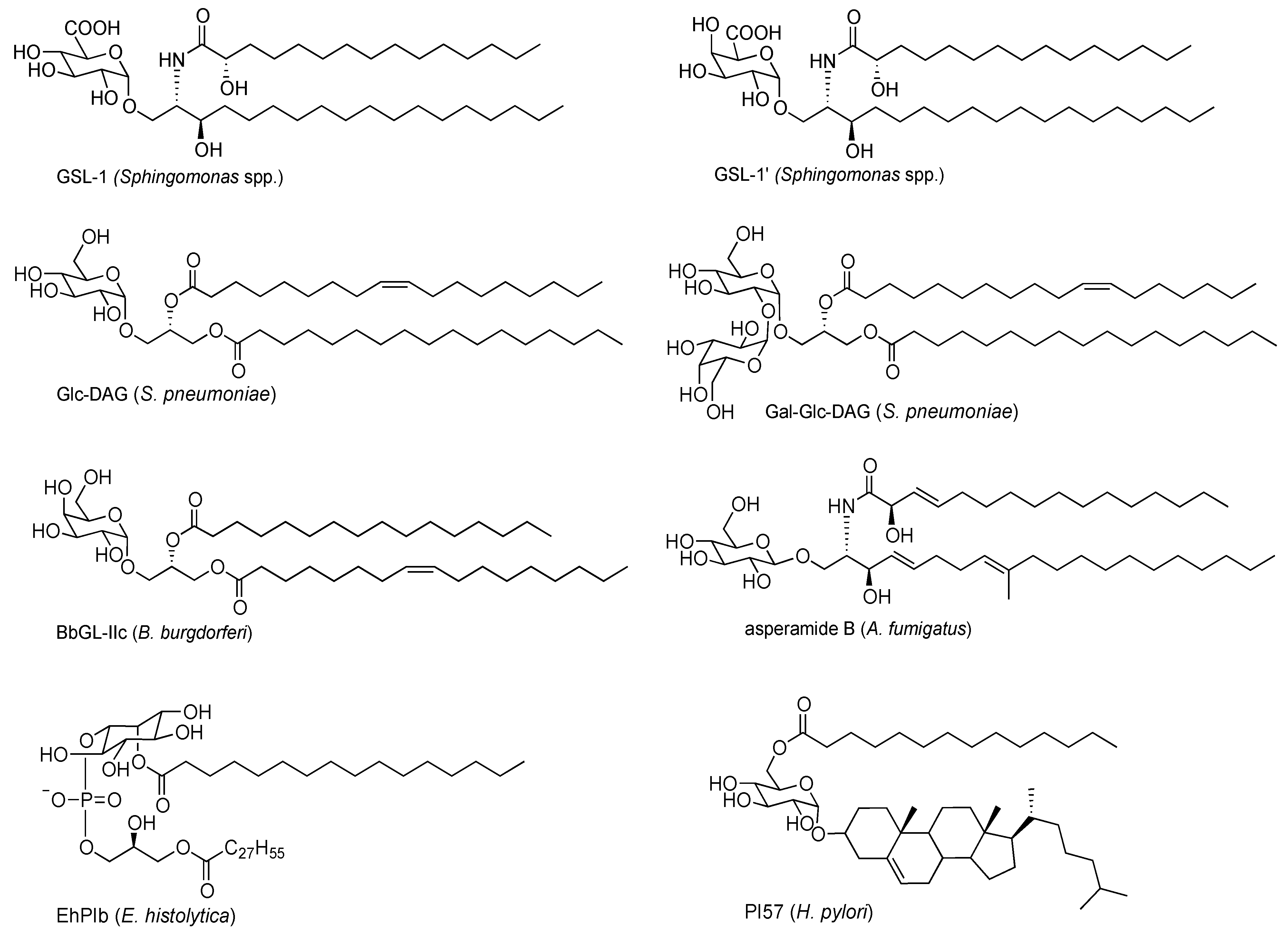
2.2. Bacterial Diacylglycerols
2.3. Protozoan Phosphoinositols
2.4. Fungal Glycolipids
2.5. H. pylori Glycolipid
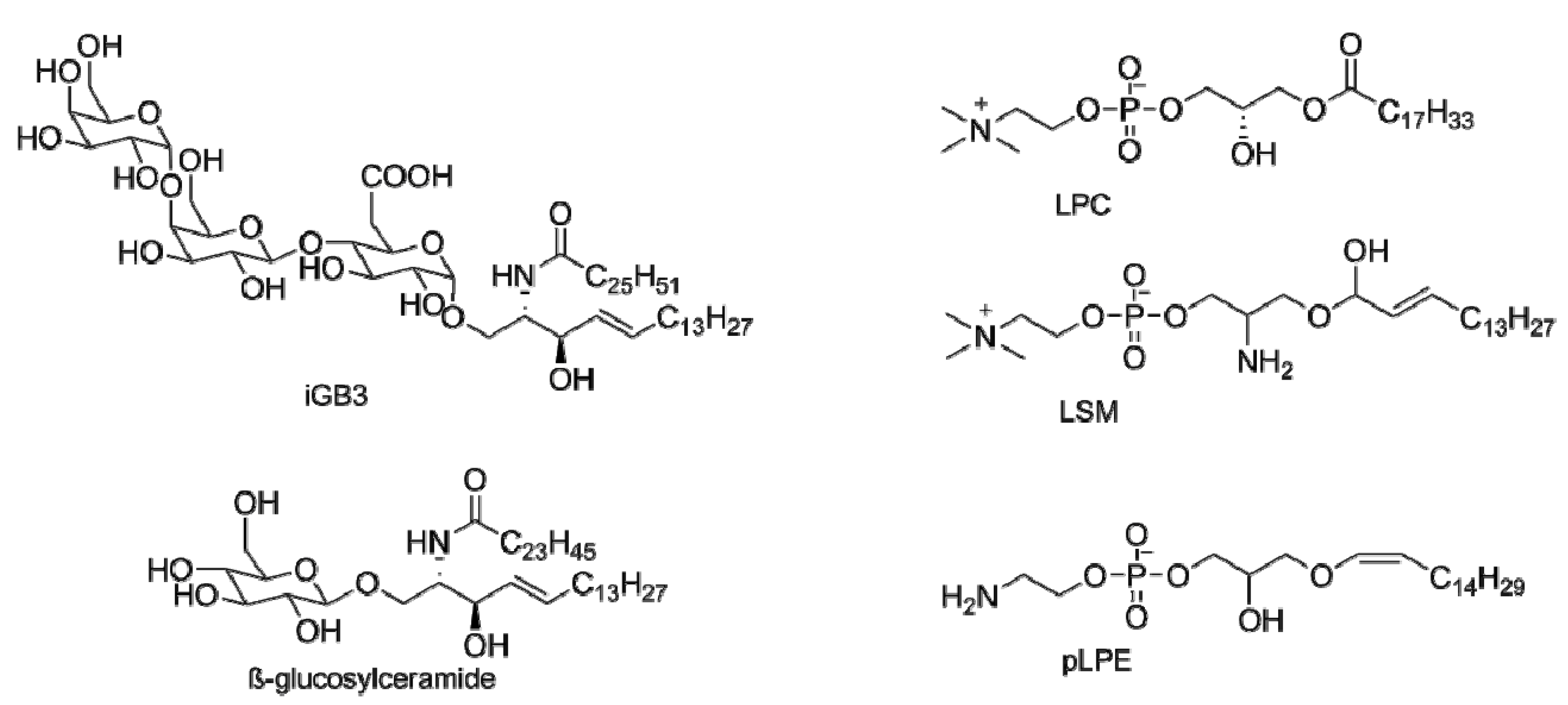
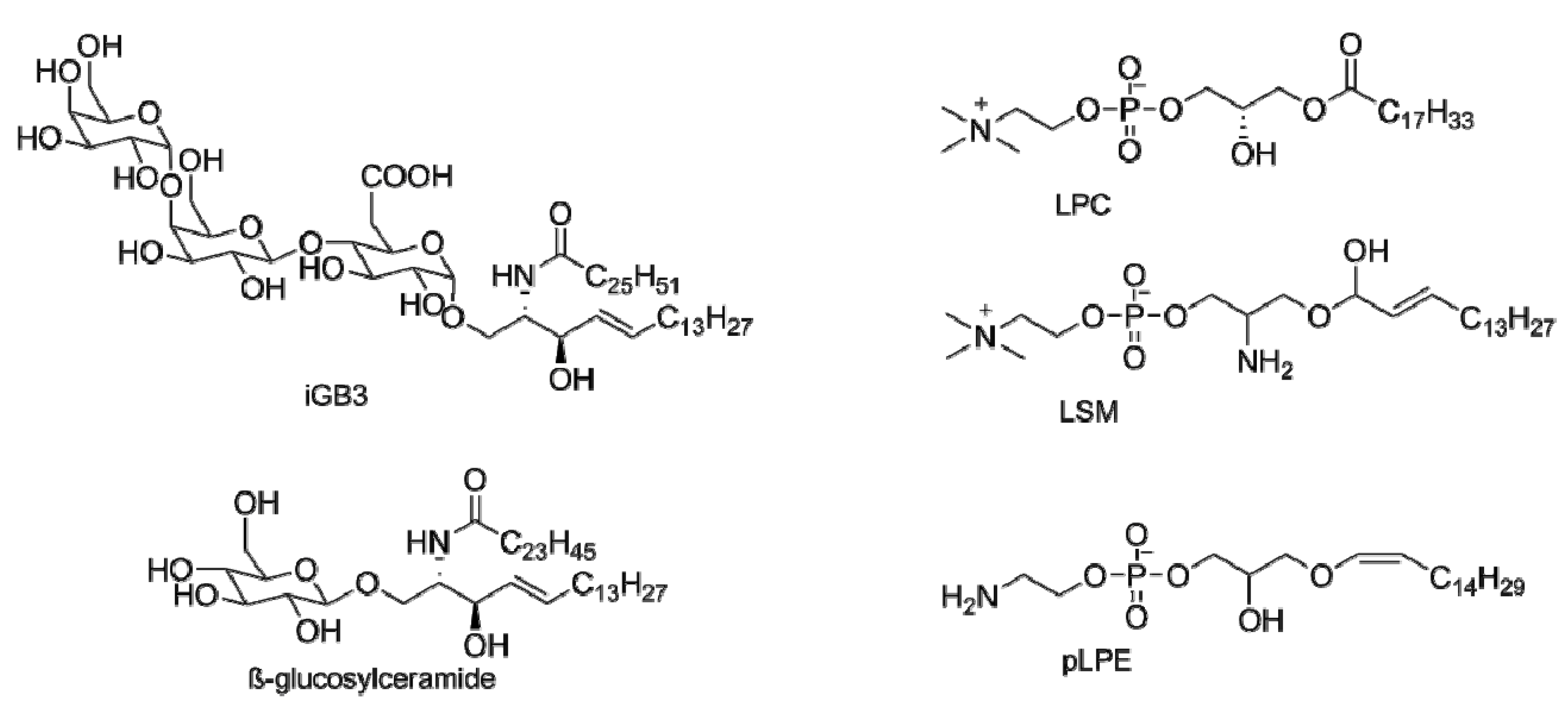
3. Endogenous Antigens for iNKT Cells
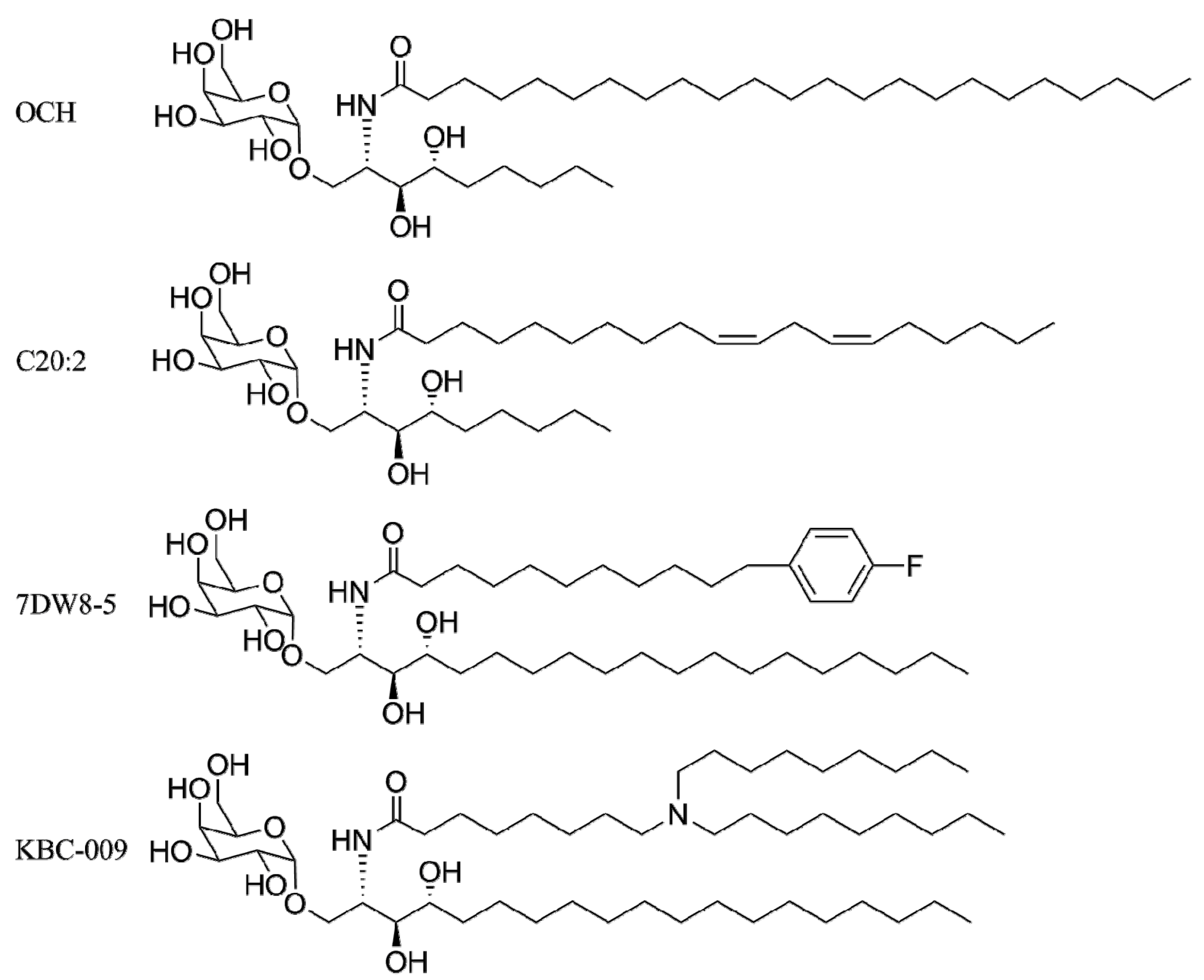
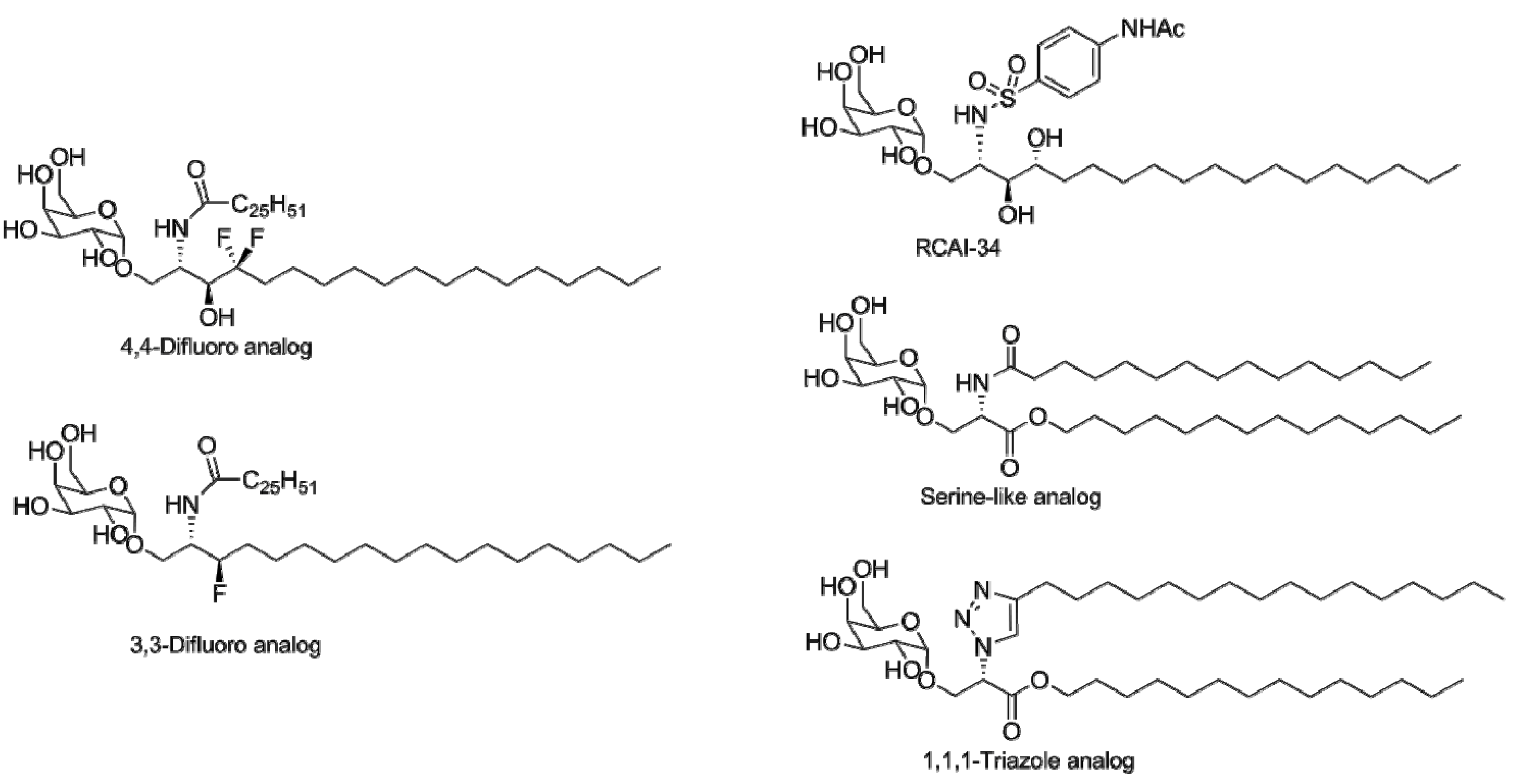
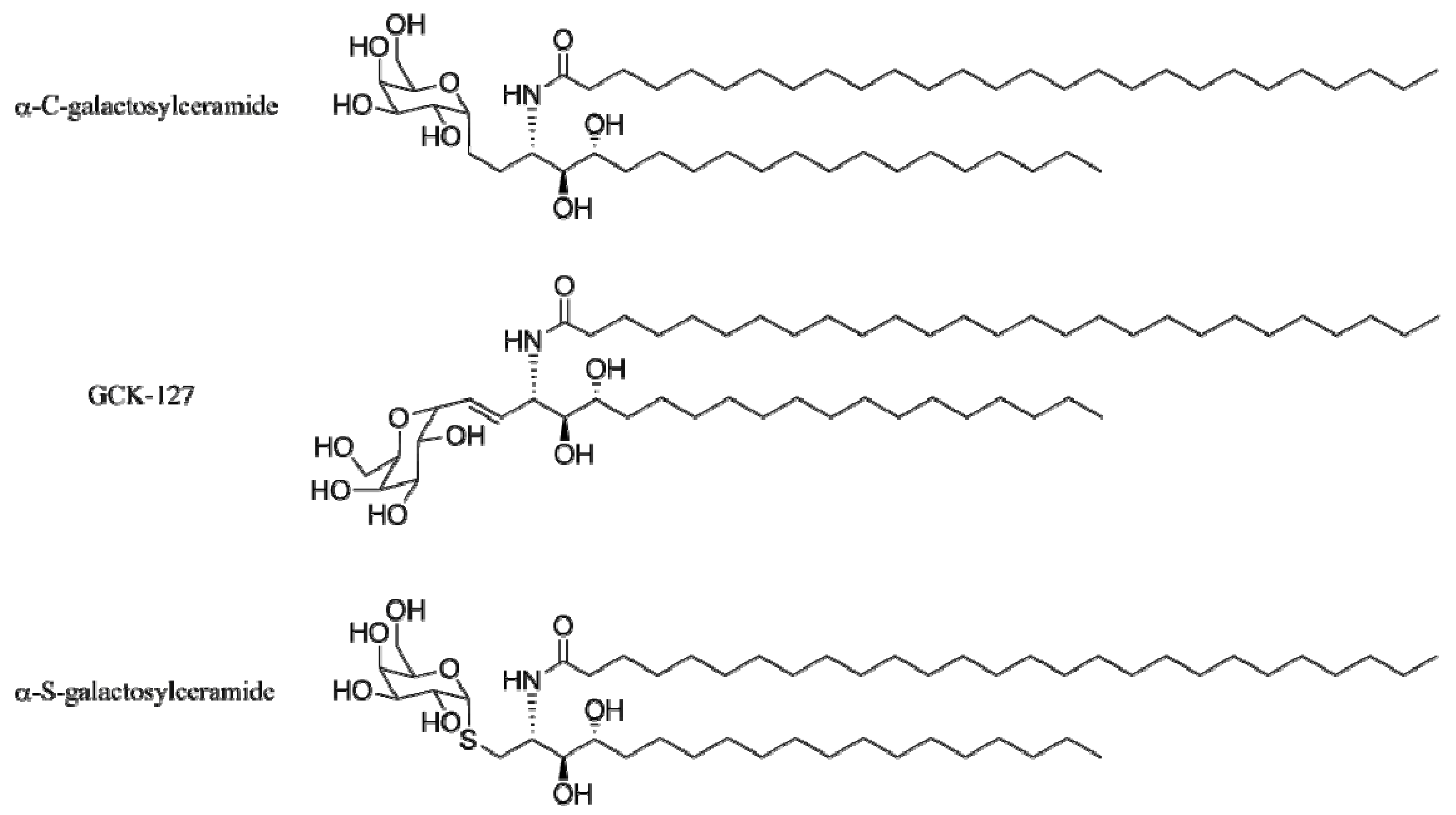
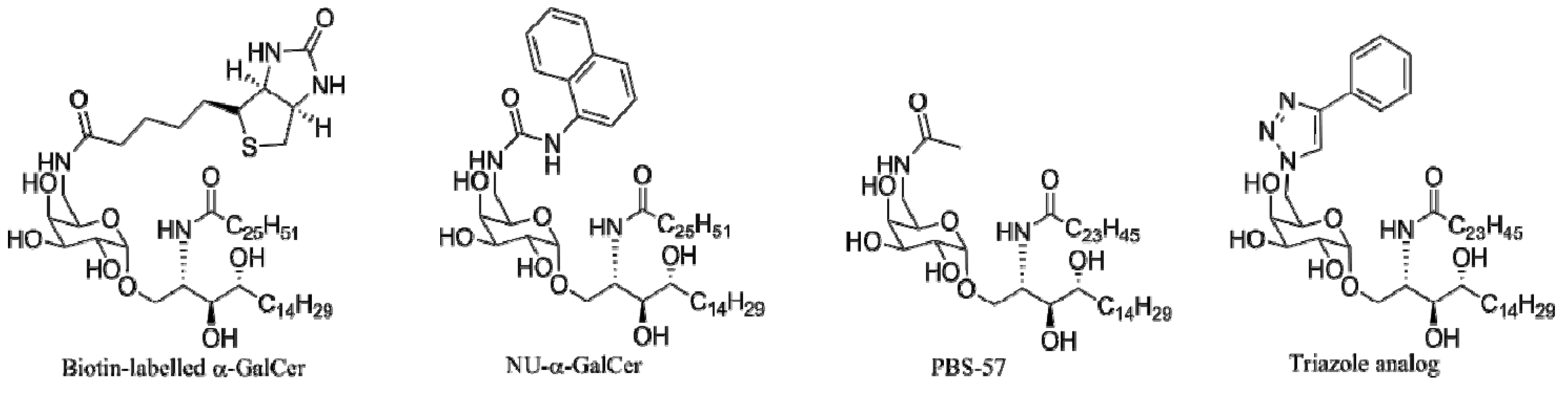
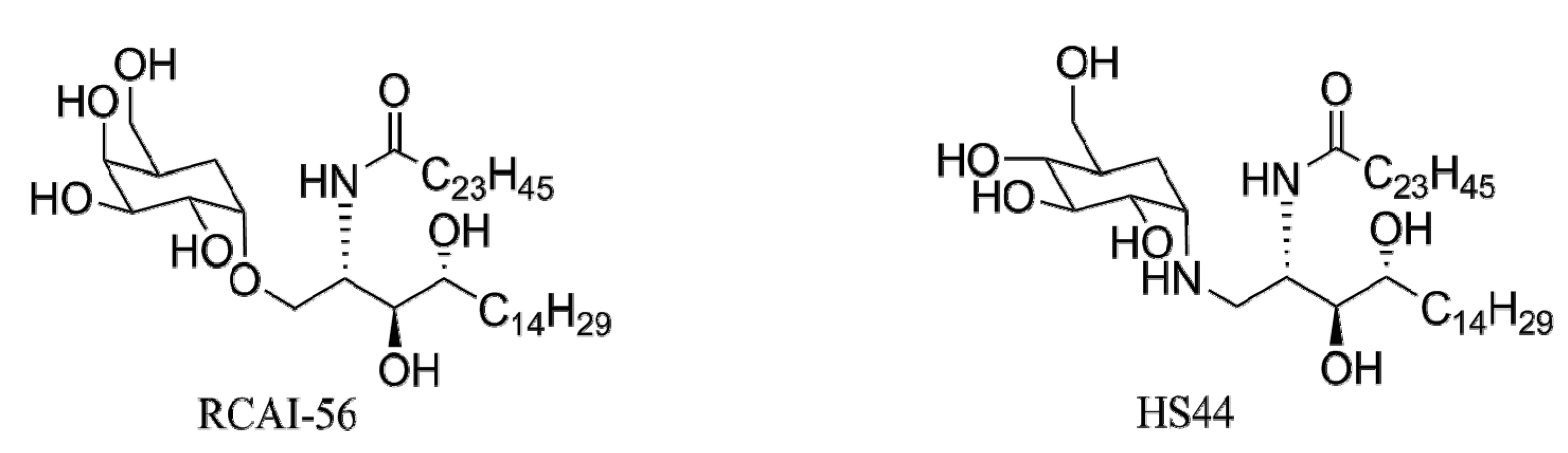
4. α-GalCer Analogs
4.1. Modification of the Lipid Chains
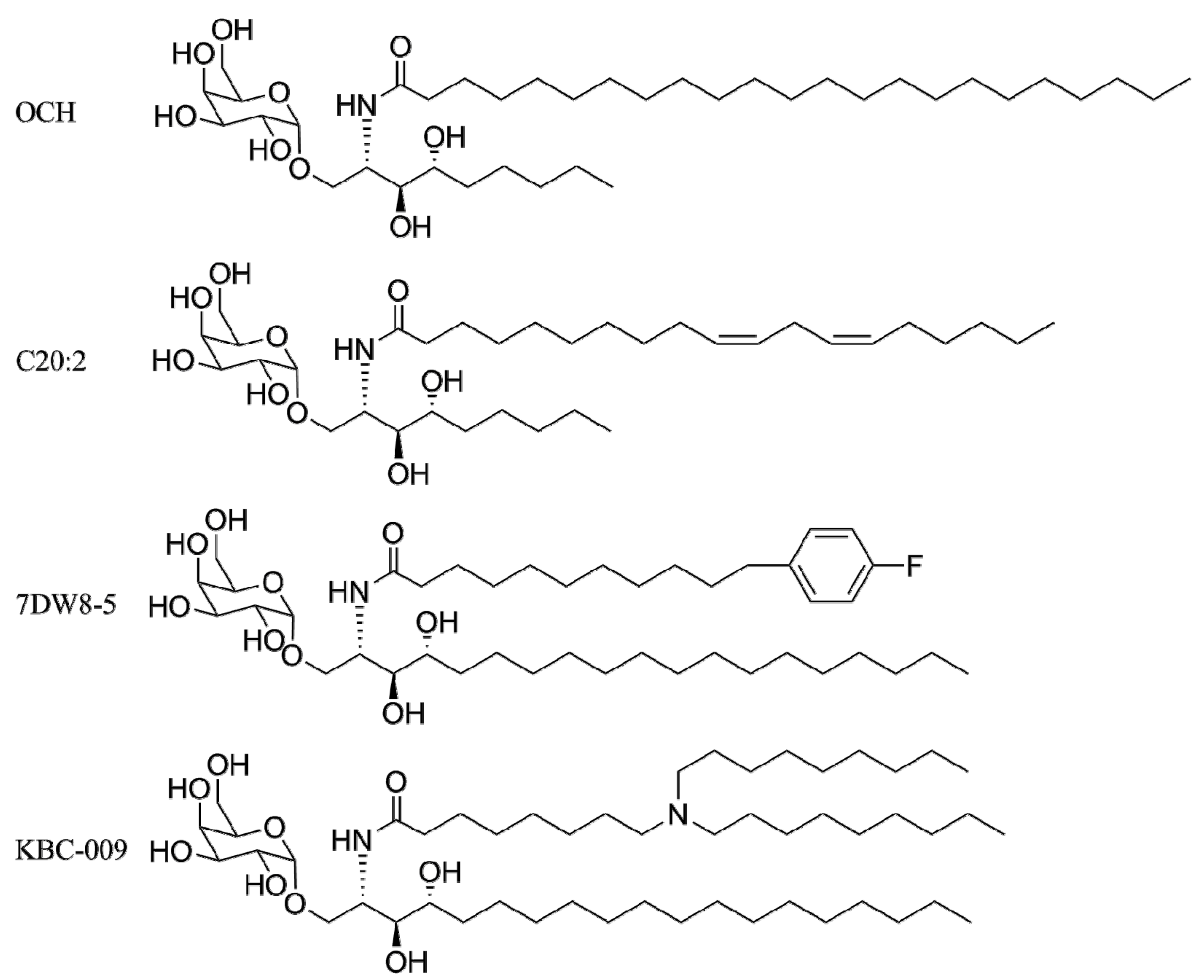
4.2. Modifications of the Sphingosine Base
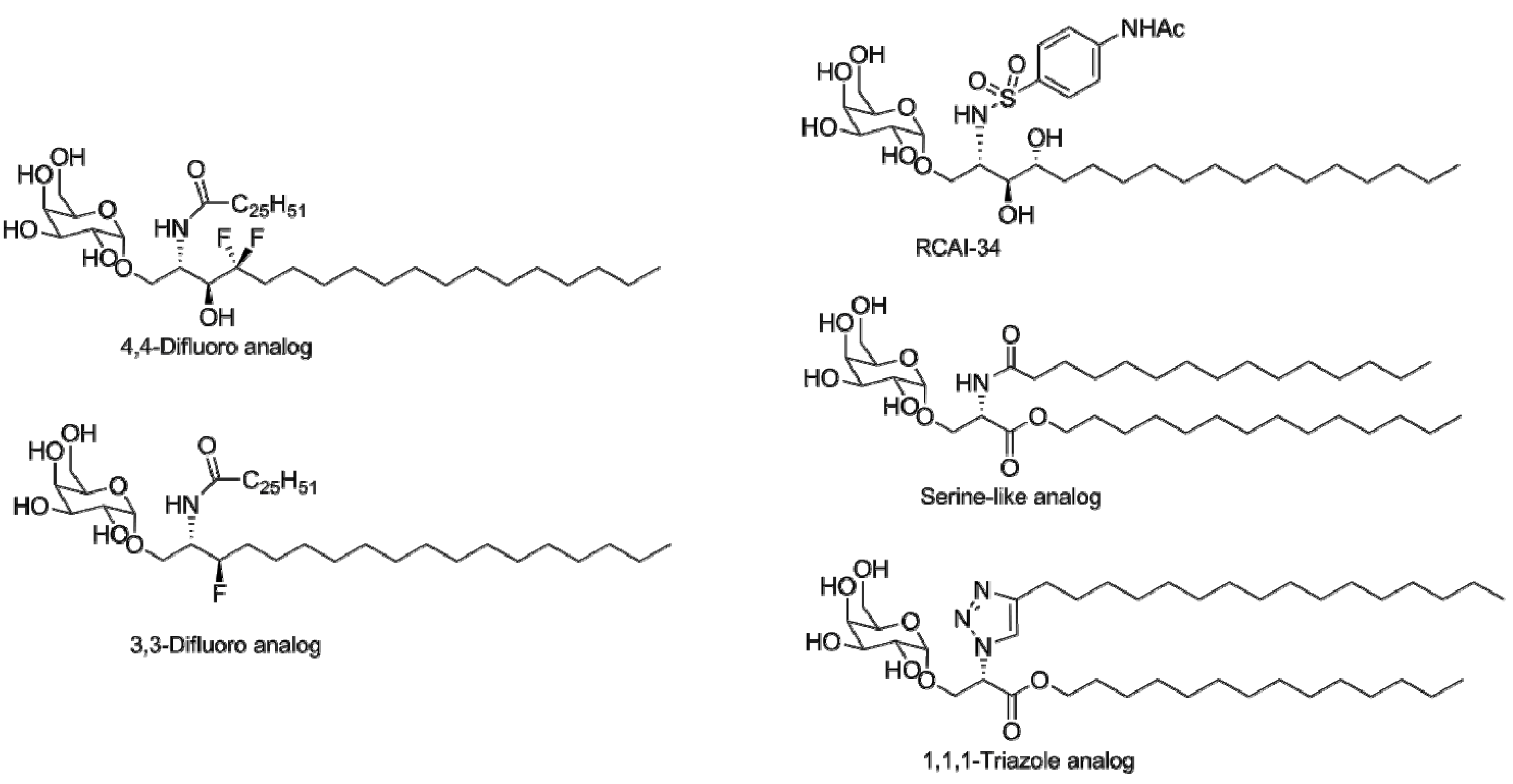
4.3. Modifications of the Glycosidic Linkage
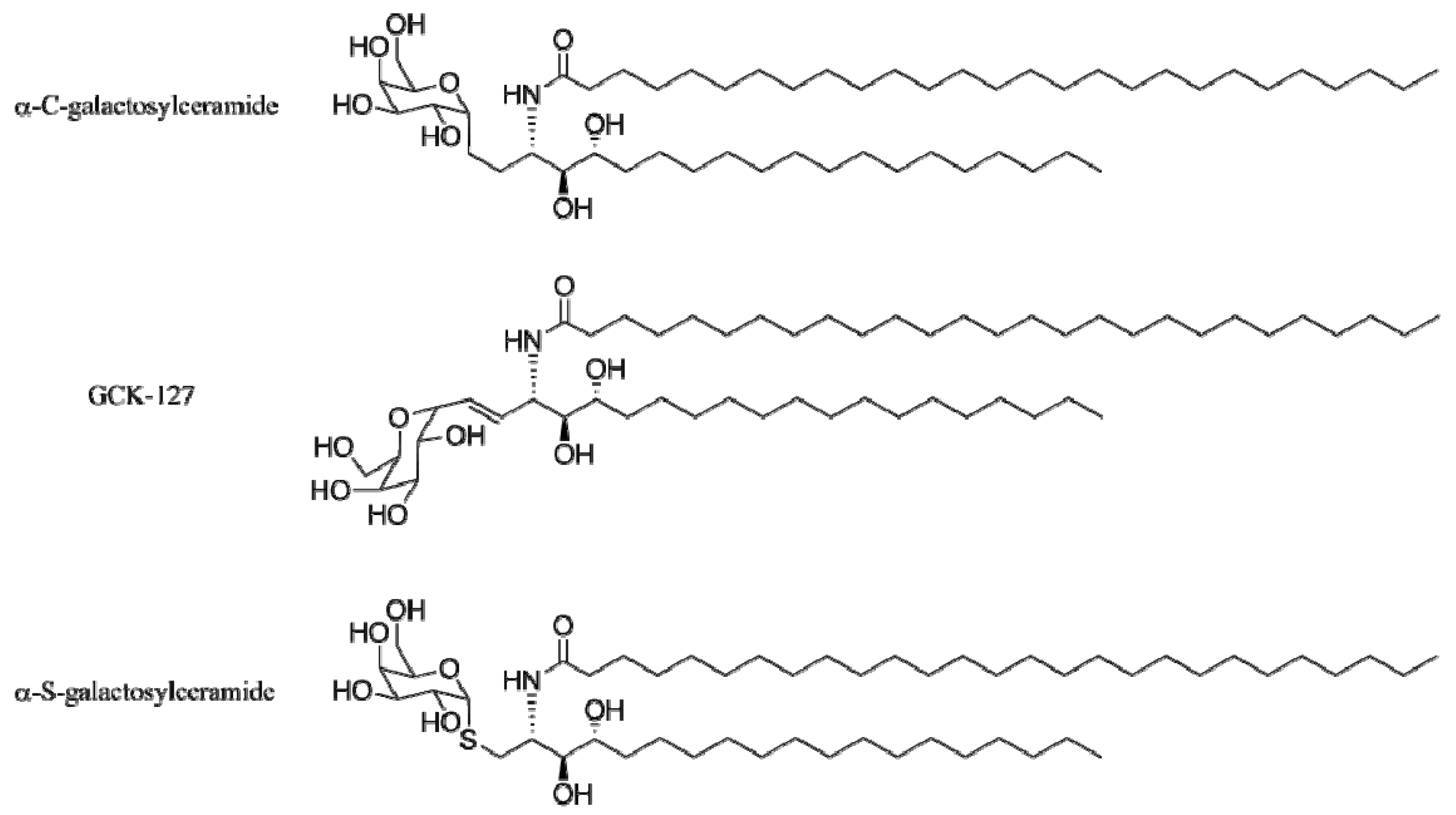
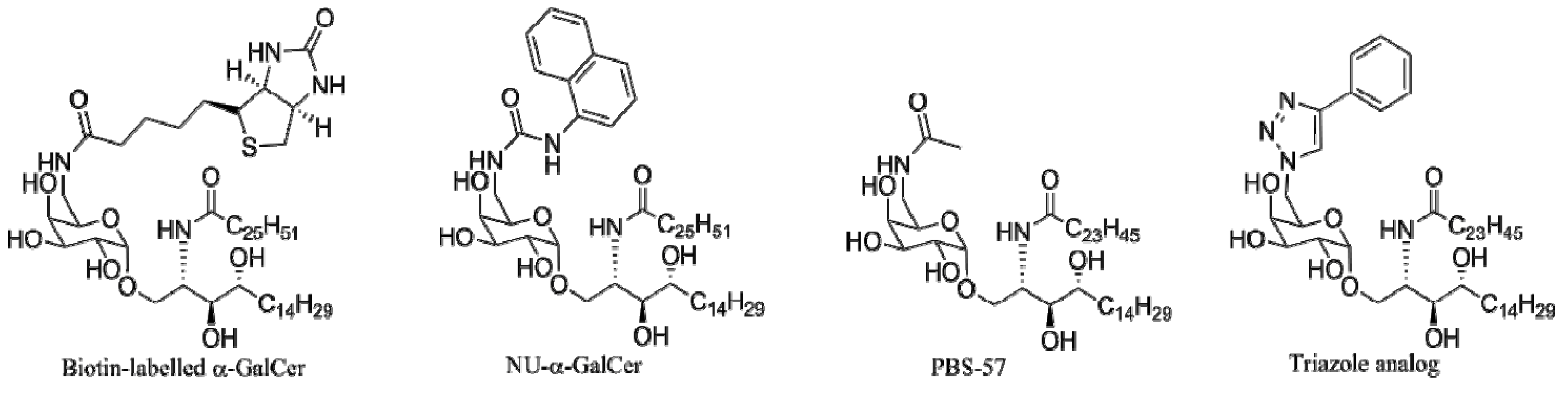
4.4. Modifications of the Saccharide
4.4.1. 6"-α-GalCer Analogs
4.4.2. Carbocyclic Analogs of α-GalCer
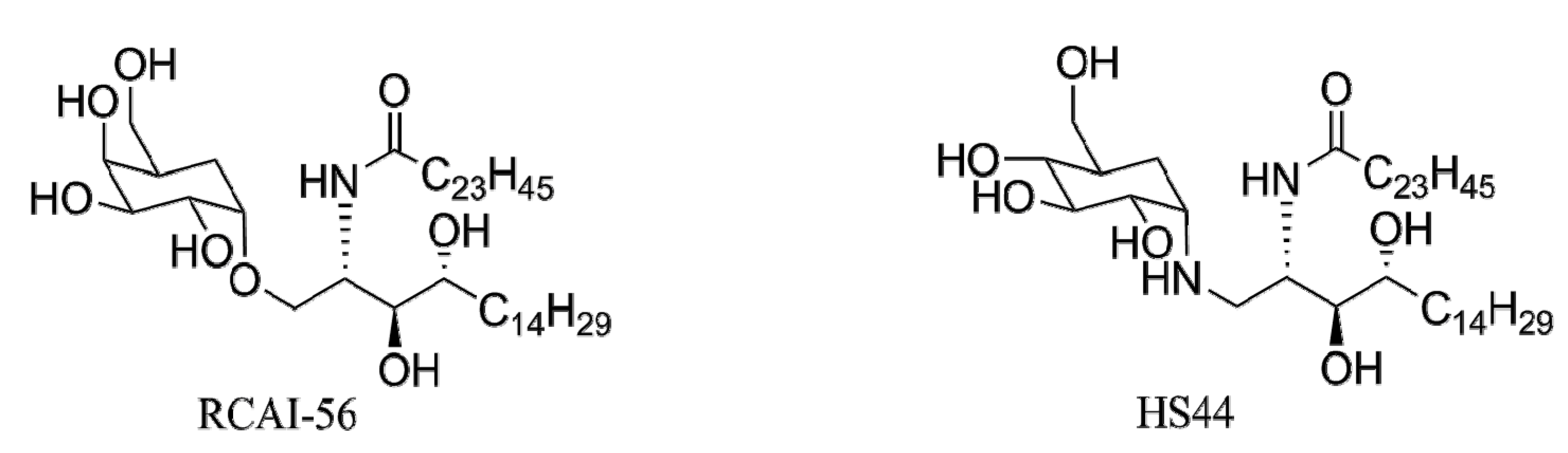
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Bendelac, A.; Savage, P.B.; Teyton, L. The biology of NKT cells. Annu. Rev. Immunol. 2007, 25, 297–336. [Google Scholar] [CrossRef]
- Savage, P.B.; Teyton, L.; Bendelac, A. Glycolipids for natural killer T cells. Chem. Soc. Rev. 2006, 35, 771–779. [Google Scholar] [CrossRef]
- Gapin, L.; Godfrey, D.I.; Rossjohn, J. Natural killer T cell obsession with self-antigens. Curr. Opin. Immunol. 2013, 25, 168–173. [Google Scholar] [CrossRef]
- Pellicci, D.G.; Patel, O.; Kjer-Nielsen, L.; Pang, S.S.; Sullivan, L.C.; Kyparissoudis, K.; Brooks, A.G.; Reid, H.H.; Gras, S.; Lucet, I.S.;et al. Differential recognition of CD1d-alpha-galactosyl ceramide by the v beta 8.2 and v beta 7 semi-invariant NKT T cell receptors. Immunity 2009, 31, 47–59. [Google Scholar] [CrossRef]
- Zajonc, D.M.; Kronenberg, M. Cd1 mediated T cell recognition of glycolipids. Curr. Opin. Struct. Biol. 2007, 17, 521–529. [Google Scholar] [CrossRef]
- Cerundolo, V.; Barral, P.; Batista, F.D. Synthetic iNKT cell-agonists as vaccine adjuvants--finding the balance. Curr. Opin. Immunol. 2010, 22, 417–424. [Google Scholar] [CrossRef]
- Hong, S.; Scherer, D.C.; Singh, N.; Mendiratta, S.K.; Serizawa, I.; Koezuka, Y.; van Kaer, L. Lipid antigen presentation in the immune system: Lessons learned from CD1d knockout mice. Immunol. Rev. 1999, 169, 31–44. [Google Scholar] [CrossRef]
- Berkers, C.R.; Ovaa, H. Immunotherapeutic potential for ceramide-based activators of iNKT cells. Trends Pharmacol. Sci. 2005, 26, 252–257. [Google Scholar] [CrossRef]
- Wu, D.; Fujio, M.; Wong, C.H. Glycolipids as immunostimulating agents. Bioorg. Med. Chem. 2008, 16, 1073–1083. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Kinjo, Y.; Mac, T.T.; Gibson, D.; Painter, G.F.; Kronenberg, M.; Zajonc, D.M. Lipid binding orientation within CD1d affects recognition of borrelia burgorferi antigens by NKT cells. Proc. Natl. Acad. Sci. USA 2010, 107, 1535–1540. [Google Scholar]
- Cernadas, M.; Cavallari, M.; Watts, G.; Mori, L.; de Libero, G.; Brenner, M.B. Early recycling compartment trafficking of CD1a is essential for its intersection and presentation of lipid antigens. J. Immunol. 2010, 184, 1235–1241. [Google Scholar]
- Cerundolo, V.; Silk, J.D.; Masri, S.H.; Salio, M. Harnessing invariant NKT cells in vaccination strategies. Nat. Rev. Immunol. 2009, 9, 28–38. [Google Scholar] [CrossRef]
- Godfrey, D.I.; Pellicci, D.G.; Patel, O.; Kjer-Nielsen, L.; McCluskey, J.; Rossjohn, J. Antigen recognition by CD1d-restricted NKT T cell receptors. Semin Immunol. 2010, 22, 61–67. [Google Scholar] [CrossRef]
- Zajonc, D.M.; Maricic, I.; Wu, D.; Halder, R.; Roy, K.; Wong, C.H.; Kumar, V.; Wilson, I.A. Structural basis for CD1d presentation of a sulfatide derived from myelin and its implications for autoimmunity. J. Exp. Med. 2005, 202, 1517–1526. [Google Scholar] [CrossRef]
- Kronenberg, M. Toward an understanding of NKT cell biology: Progress and paradoxes. Annu. Rev. Immunol. 2005, 23, 877–900. [Google Scholar] [CrossRef]
- Lawson, V. Turned on by danger: Activation of CD1d-restricted invariant natural killer T cells. Immunology 2012, 137, 20–27. [Google Scholar] [CrossRef]
- Matsuda, J.L.; Mallevaey, T.; Scott-Browne, J.; Gapin, L. Cd1d-restricted iNKT cells, the ‘swiss-army knife’ of the immune system. Curr. Opin. Immunol. 2008, 20, 358–368. [Google Scholar] [CrossRef]
- Van Kaer, L. Alpha-galactosylceramide therapy for autoimmune diseases: Prospects and obstacles. Nat. Rev. Immunol. 2005, 5, 31–42. [Google Scholar] [CrossRef]
- Kinjo, Y.; Ueno, K. iNKT cells in microbial immunity: Recognition of microbial glycolipids. Microbiol. Immunol. 2011, 55, 472–482. [Google Scholar] [CrossRef]
- Schneiders, F.L.; Scheper, R.J.; von Blomberg, B.M.E.; Woltman, A.M.; Janssen, H.L.A.; van den Eertwegh, A.J.M.; Verheul, H.M.W.; de Gruijl, T.D.; van der Vliet, H.J. Clinical experience with alpha-galactosylceramide (krn7000) in patients with advanced cancer and chronic hepatitis b/c infection. Clin. Immunol. 2011, 140, 130–141. [Google Scholar] [CrossRef]
- Tsuji, M. Glycolipids and phospholipids as natural CD1d-binding NKT cell ligands. Cell. Mol. Life Sci. 2006, 63, 1889–1898. [Google Scholar] [CrossRef]
- Natori, T.; Koezuka, Y.; Higa, T. Agelasphins, novel alpha-galactosylceramides from the marine sponge agelas-mauritianus. Tetrahedron Lett. 1993, 34, 5591–5592. [Google Scholar] [CrossRef]
- Natori, T.; Morita, M.; Akimoto, K.; Koezuka, Y. Agelasphins, novel antitumor and immunostimulatory cerebrosides from the marine sponge agelas-mauritianus. Tetrahedron 1994, 50, 2771–2784. [Google Scholar] [CrossRef]
- Kobayashi, E.; Motoki, K.; Yamaguchi, Y.; Uchida, T.; Fukushima, H.; Koezuka, Y. Enhancing effects of alpha-,beta-monoglycosylceramides on natural killer cell activity. Bioorg. Med. Chem. 1996, 4, 615–619. [Google Scholar] [CrossRef]
- Morita, M.; Motoki, K.; Akimoto, K.; Natori, T.; Sakai, T.; Sawa, E.; Yamaji, K.; Koezuka, Y.; Kobayashi, E.; Fukushima, H. Structure-activity relationship of alpha-galactosylceramides against b16-bearing mice. J. Med. Chem. 1995, 38, 2176–2187. [Google Scholar] [CrossRef]
- Morita, M.; Natori, T.; Akimoto, K.; Osawa, T.; Fukushima, H.; Koezuka, Y. Syntheses of alpha-monoglycosylceramides, beta-monoglycosylceramides and 4 diastereomers of an alpha-galactosylceramide. Bioorg. Med. Chem. Lett. 1995, 5, 699–704. [Google Scholar] [CrossRef]
- Kawano, T.; Cui, J.Q.; Koezuka, Y.; Toura, I.; Kaneko, Y.; Motoki, K.; Ueno, H.; Nakagawa, R.; Sato, H.; Kondo, E.; et al. Cd1d-restricted and TCR-mediated activation of v(alpha)14 NKT cells by glycosylceramides. Science 1997, 278, 1626–1629. [Google Scholar] [CrossRef]
- Yu, K.O.A.; Porcelli, S.A. The diverse functions of CD1d-restricted NKT cells and their potential for immunotherapy. Immunol. Lett. 2005, 100, 42–55. [Google Scholar] [CrossRef]
- Matsuda, J.L.; Gapin, L.; Baron, J.L.; Sidobre, S.; Stetson, D.B.; Mohrs, M.; Locksley, R.M.; Kronenberg, M. Mouse v alpha 14i natural killer T cells are resistant to cytokine polarization in vivo. Proc. Natl. Acad. Sci. USA 2003, 100, 8395–8400. [Google Scholar] [CrossRef]
- Uldrich, A.P.; Crowe, N.Y.; Kyparissoudis, K.; Pellicci, D.G.; Zhan, Y.F.; Lew, A.M.; Bouillet, P.; Strasser, A.; Smyth, M.J.; Godfrey, D.I. NkT cell stimulation with glycolipid antigen in vivo: Costimulation-dependent expansion, bim-dependent contraction, and hyporesponsiveness to further antigenic challenge. J. Immunol. 2005, 175, 3092–3101. [Google Scholar]
- Parekh, V.V.; Wilson, M.T.; Olivares-Villagomez, D.; Singh, A.K.; Wu, L.; Wang, C.R.; Joyce, S.; van Kaer, L. Glycolipid antigen induces long-term natural killer T cell anergy in mice. J. Clin. Invest. 2005, 115, 2572–2583. [Google Scholar]
- Mattner, J.; DeBord, K.L.; Ismail, N.; Goff, R.D.; Cantu, C.; Zhou, D.P.; Saint-Mezard, P.; Wang, V.; Gao, Y.; Yin, N.; et al. Exogenous and endogenous glycolipid antigens activate NKT cells during microbial infections. Nature 2005, 434, 525–529. [Google Scholar] [CrossRef]
- Kinjo, Y.; Wu, D.; Kim, G.S.; Xing, G.W.; Poles, M.A.; Ho, D.D.; Tsuji, M.; Kawahara, K.; Wong, C.H.; Kronenberg, M. Recognition of bacterial glycosphingolipids by natural killer T cells. Nature 2005, 434, 520–525. [Google Scholar] [CrossRef]
- Sriram, V.; Du, W.J.; Gervay-Hague, J.; Brutkiewicz, R.R. Cell wall glycosphingolipids sphingomonas paucimobilis are CD1d-specific ligands for NKT cells. Eur. J. Immunol. 2005, 35, 1692–1701. [Google Scholar] [CrossRef]
- Long, X.T.; Deng, S.; Mattner, J.C.; Zang, Z.; Zhou, D.; McNary, N.; Goff, R.D.; Teyton, L.; Bendelac, A.; Savage, P.B. Synthesis and evaluation of stimulatory properties of sphingomonadaceae glycolipids. Nat. Chem. Biol. 2007, 3, 559–564. [Google Scholar] [CrossRef]
- Wu, D.; Zajonc, D.M.; Fujio, M.; Sullivan, B.A.; Kinjo, Y.; Kronenberg, M.; Wilson, I.A.; Wong, C.H. Design of natural killer T cell activators: Structure and function of a microbial glycosphingolipid bound to mouse CD1d. Proc. Natl. Acad. Sci. USA 2006, 103, 3972–3977. [Google Scholar] [CrossRef]
- Kinjo, Y.; Pei, B.; Bufali, S.; Raju, R.; Richardson, S.K.; Imamura, M.; Fujio, M.; Wu, D.; Khurana, A.; Kawahara, K.; et al. Natural sphingomonas glycolipids vary greatly in their ability to activate natural killer T cells. Chem. Biol. 2008, 15, 654–664. [Google Scholar] [CrossRef]
- Fischer, K.; Scotet, E.; Niemeyer, M.; Koebernick, H.; Zerrahn, J.; Maillet, S.; Hurwitz, R.; Kursar, M.; Bonneville, M.; Kaufmann, S.H.E.; et al. Mycobacterial phosphatidylinositol mannoside is a natural antigen for CD1d-restricted T cells. Proc. Natl. Acad. Sci. USA 2004, 101, 10685–10690. [Google Scholar] [CrossRef]
- Kinjo, Y.; Tupin, E.; Wu, D.; Fujio, M.; Garcia-Navarro, R.; Benhnia, M.R.; Zajonc, D.M.; Ben-Menachem, G.; Ainge, G.D.; Painter, G.F.; et al. Natural killer T cells recognize diacylglycerol antigens from pathogenic bacteria. Nat. Immunol. 2006, 7, 978–986. [Google Scholar] [CrossRef]
- Centers for disease control and prevention. Summary of Notifiable Diseases – United States 2011. Morb. Mortal. Wkly. Rep. 2013, 60, 1–117. [Google Scholar]
- Kinjo, Y.; Illarionov, P.; Vela, J.L.; Pei, B.; Girardi, E.; Li, X.M.; Li, Y.L.; Imamura, M.; Kaneko, Y.; Okawara, A.; et al. Invariant natural killer T cells recognize glycolipids from pathogenic gram-positive bacteria. Nat. Immunol. 2011, 12, U966–U972. [Google Scholar] [CrossRef]
- Lotter, H.; Gonzalez-Roldan, N.; Lindner, B.; Winau, F.; Isibasi, A.; Moreno-Lafont, M.; Ulmer, A.J.; Holst, O.; Tannich, E.; Jacobs, T. Natural killer T cells activated by a lipopeptidophosphoglycan from entamoeba histolytica are critically important to control amebic liver abscess. PLoS Pathog. 2009, 5, e1000434. [Google Scholar] [CrossRef]
- Amoebiasis. In Weekly Epidemiology Record; World Health Organization: Geneva, Switzerland, 1997; pp. 97–99.
- Lotter, H.; Jacobs, T.; Gaworski, I.; Tannich, E. Sexual dimorphism in the control of amebic liver abscess in a mouse model of disease. Infect. Immun. 2006, 74, 118–124. [Google Scholar] [CrossRef]
- Albacker, L.A.; Chaudhary, V.; Chang, Y.J.; Kim, H.Y.; Chuang, Y.T.; Pichavant, M.; Dekruyff, R.H.; Savage, P.B.; Umetsu, D.T. Invariant natural killer T cells recognize a fungal glycosphingolipid that can induce airway hyperreactivity. Nat. Med. 2013, 19, 1297–1304. [Google Scholar] [CrossRef]
- Shelton, B.G.; Kirkland, K.H.; Flanders, W.D.; Morris, G.K. Profiles of airborne fungi in buildings and outdoor environments in the united states. Appl. Environ. Microbiol. 2002, 68, 1743–1753. [Google Scholar] [CrossRef]
- O’Connor, G.T.; Walter, M.; Mitchell, H.; Kattan, M.; Morgan, W.J.; Gruchalla, R.S.; Pongracic, J.A.; Smartt, E.; Stout, J.W.; Evans, R.; et al. Airborne fungi in the homes of children with asthma in low-income urban communities: The inner-city asthma study. J. Allergy Clin. Immunol. 2004, 114, 599–606. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, S.; Li, X.M.; Cui, C.M.; Feng, C.; Wang, B.G. New sphingolipids with a previously unreported 9-methyl-c20-sphingosine moiety from a marine algous endophytic fungus Aspergillus niger en-13. Lipids 2007, 42, 759–764. [Google Scholar] [CrossRef]
- Chaudhary, V.; Albacker, L.A.; Deng, S.; Chuang, Y.T.; Li, Y.; Umetsu, D.T.; Savage, P.B. Synthesis of fungal glycolipid asperamide B and investigation of its ability to stimulate natural killer T cells. Org. Lett. 2013, 15, 5242–5245. [Google Scholar] [CrossRef]
- Reibman, J.; Marmor, M.; Filner, J.; Fernandez-Beros, M.E.; Rogers, L.; Perez-Perez, G.I.; Blaser, M.J. Asthma is inversely associated with helicobacter pylori status in an urban population. PLoS One 2008, 3, e4060. [Google Scholar] [CrossRef]
- Chang, Y.J.; Kim, H.Y.; Albacker, L.A.; Lee, H.H.; Baumgarth, N.; Akira, S.; Savage, P.B.; Endo, S.; Yamamura, T.; Maaskant, J.; et al. Influenza infection in suckling mice expands an NKT cell subset that protects against airway hyperreactivity. J. Clin. Invest. 2011, 121, 57–69. [Google Scholar] [CrossRef]
- Stritesky, G.L.; Jameson, S.C.; Hogquist, K.A. Selection of self-reactive T cells in the thymus. Annu. Rev. Immunol. 2012, 30, 95–114. [Google Scholar] [CrossRef]
- Pei, B.; Speak, A.O.; Shepherd, D.; Butters, T.; Cerundolo, V.; Platt, F.M.; Kronenberg, M. Diverse endogenous antigens for mouse NKT cells: Self-antigens that are not glycosphingolipids. J. Immunol. 2011, 186, 1348–1360. [Google Scholar] [CrossRef]
- Zhou, D.; Mattner, J.; Cantu, C., 3rd; Schrantz, N.; Yin, N.; Gao, Y.; Sagiv, Y.; Hudspeth, K.; Wu, Y.P.; Yamashita, T.; et al. Lysosomal glycosphingolipid recognition by NKT cells. Science 2004, 306, 1786–1789. [Google Scholar] [CrossRef]
- Christiansen, D.; Milland, J.; Mouhtouris, E.; Vaughan, H.; Pellicci, D.G.; McConville, M.J.; Godfrey, D.I.; Sandrin, M.S. Humans lack iGb3 due to the absence of functional iGb3-synthase: Implications for NKT cell development and transplantation. PLoS Biol. 2008, 6, e172. [Google Scholar] [CrossRef]
- Sanderson, J.P.; Brennan, P.J.; Mansour, S.; Matulis, G.; Patel, O.; Lissin, N.; Godfrey, D.I.; Kawahara, K.; Zahringer, U.; Rossjohn, J.; et al. Cd1d protein structure determines species-selective antigenicity of isoglobotrihexosylceramide (iGb3) to invariant NKT cells. Eur. J. Immunol. 2013, 43, 815–825. [Google Scholar] [CrossRef]
- Porubsky, S.; Speak, A.O.; Luckow, B.; Cerundolo, V.; Platt, F.M.; Grone, H.J. Normal development and function of invariant natural killer T cells in mice with isoglobotrihexosylceramide (iGb3) deficiency. Proc. Natl. Acad. Sci. USA 2007, 104, 5977–5982. [Google Scholar]
- Porubsky, S.; Speak, A.O.; Salio, M.; Jennemann, R.; Bonrouhi, M.; Zafarulla, R.; Singh, Y.; Dyson, J.; Luckow, B.; Lehuen, A.; et al. Globosides but not isoglobosides can impact the development of invariant NKT cells and their interaction with dendritic cells. J. Immunol. 2012, 189, 3007–3017. [Google Scholar] [CrossRef]
- Cox, D.; Fox, L.; Tian, R.Y.; Bardet, W.; Skaley, M.; Mojsilovic, D.; Gumperz, J.; Hildebrand, W. Determination of cellular lipids bound to human CD1d molecules. PLoS One 2009, 4, e5325. [Google Scholar]
- Fox, L.M.; Cox, D.G.; Lockridge, J.L.; Wang, X.H.; Chen, X.X.; Scharf, L.; Trott, D.L.; Ndonye, R.M.; Veerapen, N.; Besra, G.S.; et al. Recognition of lyso-phospholipids by human natural killer t lymphocytes. PLoS Biol. 2009, 7, e1000228. [Google Scholar] [CrossRef]
- Chang, D.H.; Deng, H.; Matthews, P.; Krasovsky, J.; Ragupathi, G.; Spisek, R.; Mazumder, A.; Vesole, D.H.; Jagannath, S.; Dhodapkar, M.V. Inflammation-associated lysophospholipids as ligands for CD1d-restricted T cells in human cancer. Blood 2008, 112, 1308–1316. [Google Scholar] [CrossRef]
- Lopez-Sagaseta, J.; Sibener, L.V.; Kung, J.E.; Gumperz, J.; Adams, E.J. Lysophospholipid presentation by CD1d and recognition by a human natural killer t-cell receptor. EMBO J. 2012, 31, 2047–2059. [Google Scholar] [CrossRef]
- Facciotti, F.; Ramanjaneyulu, G.S.; Lepore, M.; Sansano, S.; Cavallari, M.; Kistowska, M.; Forss-Petter, S.; Ni, G.H.; Colone, A.; Singhal, A.; et al. Peroxisome-derived lipids are self antigens that stimulate invariant natural killer T cells in the thymus. Nat. Immunol. 2012, 13, U474–U473. [Google Scholar] [CrossRef]
- Brennan, P.J.; Tatituri, R.V.V.; Brigl, M.; Kim, E.Y.; Tuli, A.; Sanderson, J.P.; Gadola, S.D.; Hsu, F.F.; Besra, G.S.; Brenner, M.B. Invariant natural killer T cells recognize lipid self antigen induced by microbial danger signals. Nat. Immunol. 2011, 12, U1202–U1297. [Google Scholar] [CrossRef]
- Stanic, A.K.; de Silva, A.D.; Park, J.J.; Sriram, V.; Ichikawa, S.; Hirabyashi, Y.; Hayakawa, K.; Van Kaer, L.; Brutkiewicz, R.R.; Joyce, S. Defective presentation of the CD1d1-restricted natural va14ja18 NKT lymphocyte antigen caused by beta-d-glucosylceramide synthase deficiency. Proc. Natl. Acad. Sci. USA 2003, 100, 1849–1854. [Google Scholar] [CrossRef]
- Miyamoto, K.; Miyake, S.; Yamamura, T. A synthetic glycolipid prevents autoimmune encephalomyelitis by inducing T(h)2 bias of natural killer T cells. Nature 2001, 413, 531–534. [Google Scholar] [CrossRef]
- Yu, K.O.A.; Im, J.S.; Molano, A.; Dutronc, Y.; Illarionov, P.A.; Forestier, C.; Fujiwara, N.; Arias, I.; Miyake, S.; Yamamura, T.; et al. Modulation of CD1d-restricted NKT cell responses by using n-acyl variants of alpha-galactosylceramides. Proc. Natl. Acad. Sci. USA 2005, 102, 3383–3388. [Google Scholar] [CrossRef]
- Forestier, C.; Takaki, T.; Molano, A.; Im, J.S.; Baine, I.; Jerud, E.S.; Illarionov, P.; Ndonye, R.; Howell, A.R.; Santamaria, P.; et al. Improved outcomes in nod mice treated with a novel th2 cytokine-biasing NKT cell activator. J. Immunol. 2007, 178, 1415–1425. [Google Scholar]
- Velmourougane, G.; Raju, R.; Bricard, G.; Im, J.S.; Besra, G.S.; Porcelli, S.A.; Howell, A.R. Synthesis and evaluation of an acyl-chain unsaturated analog of the th2 biasing, immunostimulatory glycolipid, och. Bioorg. Med. Chem. Lett. 2009, 19, 3386–3388. [Google Scholar] [CrossRef]
- Goff, R.D.; Gao, Y.; Mattner, J.; Zhou, D.P.; Yin, N.; Cantu, C.; Teyton, L.; Bendelac, A.; Savage, P.B. Effects of lipid chain lengths in alpha-galactosylceramides on cytokine release by natural killer T cells. J. Am. Chem. Soc. 2004, 126, 13602–13603. [Google Scholar] [CrossRef]
- Oki, S.; Chiba, A.; Yamamura, T.; Miyake, S. The clinical implication and molecular mechanism of preferential IL-4 production by modified glycolipid-stimulated NKT cells. J. Clin. Inv. 2004, 113, 1631–1640. [Google Scholar]
- Fujio, M.; Wu, D.G.; Garcia-Navarro, R.; Ho, D.D.; Tsuji, M.; Wong, C.H. Structure-based discovery of glycolipids for CD1d-mediated NKT cell activation: Tuning the adjuvant versus immunosuppression activity. J. Am. Chem Soc. 2006, 128, 9022–9023. [Google Scholar] [CrossRef]
- Chang, Y.J.; Huang, J.R.; Tsai, Y.C.; Hung, J.T.; Wu, D.; Fujio, M.; Wong, C.H.; Yu, A.L. Potent immune-modulating and anticancer effects of NKT cell stimulatory glycolipids. Proc. Natl. Acad. Sci. USA 2007, 104, 10299–10304. [Google Scholar]
- Schiefner, A.; Fujio, M.; Wu, D.; Wong, C.H.; Wilson, I.A. Structural evaluation of potent NKT cell agonists: Implications for design of novel stimulatory ligands. J. Mol. Biol. 2009, 394, 71–82. [Google Scholar] [CrossRef]
- Li, X.M.; Fujio, M.; Imamura, M.; Wu, D.; Vasan, S.; Wong, C.H.; Ho, D.D.; Tsuji, M. Design of a potent CD1d-binding NKT cell ligand as a vaccine adjuvant. Proc. Natl. Acad. Sci. USA 2010, 107, 13010–13015. [Google Scholar]
- Michieletti, M.; Bracci, A.; Compostella, F.; de Libero, G.; Mori, L.; Fallarini, S.; Lombardi, G.; Panza, L. Synthesis of alpha-galactosyl ceramide (krn7000) and analogues thereof via a common precursor and their preliminary biological assessment. J. Org. Chem. 2008, 73, 9192–9195. [Google Scholar] [CrossRef]
- Toba, T.; Murata, K.; Futamura, J.; Nakanishi, K.; Takahashi, B.; Takemoto, N.; Tomino, M.; Nakatsuka, T.; Imajo, S.; Goto, M.; et al. Synthesis and biological evaluation of truncated alpha-galactosylceramide derivatives focusing on cytokine induction profile. Bioorg. Med. Chem. 2012, 20, 2850–2859. [Google Scholar] [CrossRef]
- Leung, L.; Tomassi, C.; Van Beneden, K.; Decruy, T.; Trappeniers, M.; Elewaut, D.; Gao, Y.; Elliott, T.; Al-Shamkhani, A.; Ottensmeier, C.; et al. The synthesis and in vivo evaluation of 2 ',2'-difluoro krn7000. ChemMedChem 2009, 4, 329–334. [Google Scholar] [CrossRef]
- Bricard, G.; Venkataswamy, M.M.; Yu, K.O.A.; Im, J.S.; Ndonye, R.M.; Howell, A.R.; Veerapen, N.; Illarionov, P.A.; Besra, G.S.; Li, Q.A.; et al. Alpha-galactosylceramide analogs with weak agonist activity for human iNKT cells define new candidate anti-inflammatory agents. PLoS One 2010, 5, e14374. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lee, K.A.; Lee, J.Y.; Kang, M.H.; Song, Y.C.; Baek, D.J.; Kim, S.; Kang, C.Y. An alpha-galcer analogue with branched acyl chain enhances protective immune responses in a nasal influenza vaccine. Vaccine 2011, 29, 417–425. [Google Scholar] [CrossRef]
- Borg, N.A.; Wun, K.S.; Kjer-Nielsen, L.; Wilce, M.C.J.; Pellicci, D.G.; Koh, R.; Besra, G.S.; Bharadwaj, M.; Godfrey, D.I.; McCluskey, J.; et al. Cd1d-lipid-antigen recognition by the semi-invariant NKT t-cell receptor. Nature 2007, 448, 44–49. [Google Scholar] [CrossRef]
- Trappeniers, M.; Goormans, S.; van Beneden, K.; Decruy, T.; Linclau, B.; Al-Shamkhani, A.; Elliott, T.; Ottensmeier, C.; Werner, J.M.; Elewaut, D.; et al. Synthesis and in vitro evaluation of alpha-galcer epimers. ChemMedChem 2008, 3, 1061–1070. [Google Scholar] [CrossRef]
- Ndonye, R.M.; Izmirian, D.P.; Dunn, M.F.; Yu, K.O.; Porcelli, S.A.; Khurana, A.; Kronenberg, M.; Richardson, S.K.; Howell, A.R. Synthesis and evaluation of sphinganine analogues of KRN7000 and OCH. J. Org. Chem. 2005, 70, 10260–10270. [Google Scholar] [CrossRef]
- Leung, L.; Tomassi, C.; van Beneden, K.; Decruy, T.; Elewaut, D.; Elliott, T.; Al-Shamkhani, A.; Ottensmeier, C.; van Calenbergh, S.; Werner, J.; et al. Synthesis and in vivo evaluation of 4-deoxy-4,4-difluoro-krn7000. Org. Lett. 2008, 10, 4433–4436. [Google Scholar] [CrossRef]
- Park, J.J.; Lee, J.H.; Ghosh, S.C.; Bricard, G.; Venkataswamy, M.M.; Porcelli, S.A.; Chung, S.K. Synthesis of all stereoisomers of krn7000, the CD1d-binding NKT cell ligand. Bioorg. Med. Chem. Lett. 2008, 18, 3906–3909. [Google Scholar] [CrossRef]
- Dangerfield, E.M.; Cheng, J.M.H.; Knight, D.A.; Weinkove, R.; Dunbar, P.R.; Hermans, I.F.; Timmer, M.S.M.; Stocker, B.L. Species-specific activity of glycolipid ligands for invariant NKT cells. Chembiochem 2012, 13, 1349–1356. [Google Scholar] [CrossRef]
- Hunault, J.; Diswall, M.; Frison, J.C.; Blot, V.; Rocher, J.; Marionneau-Lambot, S.; Oullier, T.; Douillard, J.Y.; Guillarme, S.; Saluzzo, C.; et al. 3-fluoro- and 3,3-difluoro-3,4-dideoxy-krn7000 analogues as new potent immunostimulator agents: Total synthesis and biological evaluation in human invariant natural killer T cells and mice. J. Med. Chem. 2012, 55, 1227–1241. [Google Scholar] [CrossRef]
- Lee, T.; Cho, M.; Ko, S.Y.; Youn, H.J.; Baek, D.J.; Cho, W.J.; Kang, C.Y.; Kim, S. Synthesis and evaluation of 1,2,3-triazole containing analogues of the immunostimulant alpha-galcer. J. Med. Chem. 2007, 50, 585–589. [Google Scholar] [CrossRef]
- Shiozaki, M.; Tashiro, T.; Koshino, H.; Nakagawa, R.; Inoue, S.; Shigeura, T.; Watarai, H.; Taniguchi, M.; Mori, K. Synthesis and biological activity of ester and ether analogues of alpha-galactosylceramide (KRN7000). Carbohydr. Res. 2010, 345, 1663–1684. [Google Scholar] [CrossRef]
- Sun, M.; Wang, Y.H.; Ye, X.S. Design and synthesis of new krn7000 analogues. Tetrahedron 2013, 69, 7438–7447. [Google Scholar] [CrossRef]
- Tashiro, T.; Hongo, N.; Nakagawa, R.; Seino, K.; Watarai, H.; Ishii, Y.; Taniguchi, M.; Mori, K. Rcai-17, 22, 24–26, 29, 31, 34–36, 38–40, and 88, the analogs of krn7000 with a sulfonamide linkage: Their synthesis and bioactivity for mouse natural killer T cells to produce th2-biased cytokines. Bioorg. Med. Chem. 2008, 16, 8896–8906. [Google Scholar] [CrossRef]
- Fan, G.T.; Pan, Y.S.; Lu, K.C.; Cheng, Y.P.; Lin, W.C.; Lin, S.; Lin, C.H.; Wong, C.H.; Fang, J.M.; Lin, C.C. Synthesis of alpha-galactosyl ceramide and the related glycolipids for evaluation of their activities on mouse splenocytes. Tetrahedron 2005, 61, 1855–1862. [Google Scholar] [CrossRef]
- Schmieg, J.; Yang, G.L.; Franck, R.W.; Tsuji, M. Superior protection against malaria and melanoma metastases by a C-glycoside analogue of the natural killer T cell ligand alpha-galactosylceramide. J. Exp. Med. 2003, 198, 1631–1641. [Google Scholar] [CrossRef]
- Tashiro, T. Structure-activity relationship studies of novel glycosphingolipids that stimulate natural killer t-cells. Biosci. Biotech. Bioch. 2012, 76, 1055–1067. [Google Scholar] [CrossRef]
- Kopecky-Bromberg, S.A.; Fraser, K.A.; Pica, N.; Carnero, E.; Moran, T.M.; Franck, R.W.; Tsuji, M.; Palese, P. Alpha-C-galactosylceramide as an adjuvant for a live attenuated influenza virus vaccine. Vaccine 2009, 27, 3766–3774. [Google Scholar] [CrossRef]
- Zajonc, D.M.; Cantu, C.; Mattner, J.; Zhou, D.P.; Savage, P.B.; Bendelac, A.; Wilson, I.A.; Teyton, L. Structure and function of a potent agonist for the semi-invariant natural killer T cell receptor. Nat. Immunol. 2005, 6, 810–818. [Google Scholar] [CrossRef]
- Chen, G.; Chien, M.; Tsuji, M.; Franck, R.W. E and Z alpha-C-galactosylceramides by julia-lythgoe-kocienski chemistry: A test of the receptor-binding model for glycolipid immunostimulants. Chembiochem 2006, 7, 1017–1022. [Google Scholar] [CrossRef]
- Li, X.M.; Chen, G.W.; Garcia-Navarro, R.; Franck, R.W.; Tsuji, M. Identification of C-glycoside analogues that display a potent biological activity against murine and human invariant natural killer T cells. Immunology 2009, 127, 216–225. [Google Scholar] [CrossRef]
- Sullivan, B.A.; Nagarajan, N.A.; Wingender, G.; Wang, J.; Scott, I.; Tsuji, M.; Franck, R.W.; Porcelli, S.A.; Zajonc, D.M.; Kronenberg, M. Mechanisms for glycolipid antigen-driven cytokine polarization by valpha14i NKT cells. J. Immunol. 2010, 184, 141–153. [Google Scholar] [CrossRef]
- Patel, O.; Cameron, G.; Pellicci, D.G.; Liu, Z.; Byun, H.S.; Beddoe, T.; McCluskey, J.; Franck, R.W.; Castano, A.R.; Harrak, Y.; et al. NKT cell recognition of CD1d-alpha-C-galactosylceramide. J. Immunol. 2011, 187, 4705–4713. [Google Scholar] [CrossRef]
- Blauvelt, M.L.; Khalili, M.; Jaung, W.; Paulsen, J.; Anderson, A.C.; Brian Wilson, S.; Howell, A.R. Alpha-s-galcer: Synthesis and evaluation for iNKT cell stimulation. Bioorg. Med. Chem. Lett. 2008, 18, 6374–6376. [Google Scholar] [CrossRef]
- Dere, R.T.; Zhu, X. The first synthesis of a thioglycoside analogue of the immunostimulant krn7000. Org. Lett. 2008, 10, 4641–4644. [Google Scholar] [CrossRef]
- Hogan, A.E.; O’Reilly, V.; Dunne, M.R.; Dere, R.T.; Zeng, S.G.; O’Brien, C.; Amu, S.; Fallon, P.G.; Exley, M.A.; O’Farrelly, C.; et al. Activation of human invariant natural killer T cells with a thioglycoside analogue of alpha-galactosylceramide. Clin. Immunol. 2011, 140, 196–207. [Google Scholar] [CrossRef]
- Xing, G.W.; Wu, D.; Poles, M.A.; Horowitz, A.; Tsuji, M.; Ho, D.D.; Wong, C.H. Synthesis and human NKT cell stimulating properties of 3-o-sulfo-alpha/beta-galactosylceramides. Bioorg. Med. Chem. 2005, 13, 2907–2916. [Google Scholar] [CrossRef]
- Barbieri, L.; Costantino, V.; Fattorusso, E.; Mangoni, A.; Aru, E.; Parapini, S.; Taramelli, D. Immunomodulatory α-galactoglycosphingolipids: Synthesis of a 2'-o-methyl-α-gal-gsl and evaluation of its immunostimulating capacity. Eur. J. Org. Chem. 2004, 2004, 468–473. [Google Scholar]
- Veerapen, N.; Leadbetter, E.A.; Brenner, M.B.; Cox, L.R.; Besra, G.S. Synthesis of a novel alpha-galactosyl ceramide haptenated-lipid antigen, a useful tool in demonstrating the involvement of iNKT cells in the production of antilipid antibodies. Bioconjugate Chem. 2010, 21, 741–747. [Google Scholar] [CrossRef]
- Wu, D.; Xing, G.W.; Poles, M.A.; Horowitz, A.; Kinjo, Y.; Sullivan, B.; Bodmer-Narkevitch, V.; Plettenburg, O.; Kronenberg, M.; Tsuji, M.; et al. Bacterial glycolipids and analogs as antigens for CD1d-restricted NKT cells. Proc. Natl. Acad. Sci. USA 2005, 102, 1351–1356. [Google Scholar] [CrossRef]
- Raju, R.; Castillo, B.F.; Richardson, S.K.; Thakur, M.; Severins, R.; Kronenberg, M.; Howell, A.R. Synthesis and evaluation of 3''- and 4''-deoxy and -fluoro analogs of the immunostimulatory glycolipid, krn7000. Bioorg. Med. Chem. Lett. 2009, 19, 4122–4125. [Google Scholar] [CrossRef]
- Xia, C.; Zhang, W.; Zhang, Y.; Chen, W.; Nadas, J.; Severin, R.; Woodward, R.; Wang, B.; Wang, X.; Kronenberg, M.; et al. The roles of 3' and 4' hydroxy groups in alpha-galactosylceramide stimulation of invariant natural killer T cells. ChemMedChem 2009, 4, 1810–1815. [Google Scholar] [CrossRef]
- Zhang, W.; Xia, C.; Nadas, J.; Chen, W.; Gu, L.; Wang, P.G. Introduction of aromatic group on 4'-oh of alpha-galcer manipulated NKT cell cytokine production. Bioorg. Med. Chem. 2011, 19, 2767–2776. [Google Scholar] [CrossRef]
- Silk, J.D.; Salio, M.; Reddy, B.G.; Shepherd, D.; Gileadi, U.; Brown, J.; Masri, S.H.; Polzella, P.; Ritter, G.; Besra, G.S.; et al. Cutting edge: Nonglycosidic CD1d lipid ligands activate human and murine invariant NKT cells. J. Immunol. 2008, 180, 6452–6456. [Google Scholar]
- Prigozy, T.I.; Naidenko, O.; Qasba, P.; Elewaut, D.; Brossay, L.; Khurana, A.; Natori, T.; Koezuka, Y.; Kulkarni, A.; Kronenberg, M. Glycolipid antigen processing for presentation by CD1d molecules. Science 2001, 291, 664–667. [Google Scholar] [CrossRef]
- Zhou, X.T.; Forestier, C.; Goff, R.D.; Li, C.; Teyton, L.; Bendelac, A.; Savage, P.B. Synthesis and NKT cell stimulating properties of fluorophore- and biotin-appended 6"-amino-6"-deoxy-galactosylceramides. Org. Lett. 2002, 4, 1267–1270. [Google Scholar] [CrossRef]
- Xia, C.; Zhang, W.; Zhang, Y.; Woodward, R.L.; Wang, J.; Wang, P.G. Facile synthesis of biotin-labelled α-galactosylceramide as antigen for invariant natural killer T cells. Tetrahedron 2009, 65, 6390–6395. [Google Scholar] [CrossRef]
- Cheng, J.M.; Chee, S.H.; Knight, D.A.; Acha-Orbea, H.; Hermans, I.F.; Timmer, M.S.; Stocker, B.L. An improved synthesis of dansylated alpha-galactosylceramide and its use as a fluorescent probe for the monitoring of glycolipid uptake by cells. Carbohydr. Res. 2011, 346, 914–926. [Google Scholar] [CrossRef]
- Liu, Y.; Deng, S.; Bai, L.; Freigang, S.; Mattner, J.; Teyton, L.; Bendelac, A.; Savage, P.B. Synthesis of diglycosylceramides and evaluation of their iNKT cell stimulatory properties. Bioorg. Med. Chem. Lett. 2008, 18, 3052–3055. [Google Scholar] [CrossRef]
- Liu, Y.; Goff, R.D.; Zhou, D.; Mattner, J.; Sullivan, B.A.; Khurana, A.; Cantu, C., 3rd; Ravkov, E.V.; Ibegbu, C.C.; Altman, J.D.; et al. A modified alpha-galactosyl ceramide for staining and stimulating natural killer T cells. J. Immunol. Methods 2006, 312, 34–39. [Google Scholar] [CrossRef]
- Trappeniers, M.; van Beneden, K.; Decruy, T.; Hillaert, U.; Linclau, B.; Elewaut, D.; van Calenbergh, S. 6'-derivatised alpha-galcer analogues capable of inducing strong CD1d-mediated th1-biased NKT cell responses in mice. J. Am. Chem. Soc. 2008, 130, 16468–16469. [Google Scholar] [CrossRef]
- Aspeslagh, S.; Li, Y.; Yu, E.D.; Pauwels, N.; Trappeniers, M.; Girardi, E.; Decruy, T.; van Beneden, K.; Venken, K.; Drennan, M.; et al. Galactose-modified iNKT cell agonists stabilized by an induced fit of CD1d prevent tumour metastasis. EMBO J. 2011, 30, 2294–2305. [Google Scholar] [CrossRef]
- Jervis, P.J.; Graham, L.M.; Foster, E.L.; Cox, L.R.; Porcelli, S.A.; Besra, G.S. New CD1d agonists: Synthesis and biological activity of 6''-triazole-substituted alpha-galactosyl ceramides. Bioorg. Med. Chem. Lett. 2012, 22, 4348–4352. [Google Scholar] [CrossRef]
- Jervis, P.J.; Moulis, M.; Jukes, J.P.; Ghadbane, H.; Cox, L.R.; Cerundolo, V.; Besra, G.S. Towards multivalent CD1d ligands: Synthesis and biological activity of homodimeric alpha-galactosyl ceramide analogues. Carbohydr. Res. 2012, 356, 152–162. [Google Scholar] [CrossRef]
- Tashiro, T.; Nakagawa, R.; Hirokawa, T.; Inoue, S.; Watarai, H.; Taniguchi, M.; Mori, K. Rcai-56, a carbocyclic analogue of krn7000: Its synthesis and potent activity for natural killer (nk) T cells to preferentially produce interferon-γ. Tetrahedron Lett. 2007, 48, 3343–3347. [Google Scholar] [CrossRef]
- Tashiro, T.; Sekine-Kondo, E.; Shigeura, T.; Nakagawa, R.; Inoue, S.; Omori-Miyake, M.; Chiba, T.; Hongo, N.; Fujii, S.; Shimizu, K.; et al. Induction of th1-biased cytokine production by alpha-carba-galcer, a neoglycolipid ligand for NKT cells. Int. Immunol. 2010, 22, 319–328. [Google Scholar] [CrossRef]
- Yoshiga, Y.; Goto, D.; Segawa, S.; Horikoshi, M.; Hayashi, T.; Matsumoto, I.; Ito, S.; Taniguchi, M.; Sumida, T. Activation of natural killer T cells by alpha-carba-galcer (rcai-56), a novel synthetic glycolipid ligand, suppresses murine collagen-induced arthritis. Clin. Exp. Immunol. 2011, 164, 236–247. [Google Scholar] [CrossRef]
- Harrak, Y.; Barra, C.M.; Bedia, C.; Delgado, A.; Castano, A.R.; Llebaria, A. Aminocyclitol-substituted phytoceramides and their effects on iNKT cell stimulation. ChemMedChem 2009, 4, 1608–1613. [Google Scholar] [CrossRef]
- Kerzerho, J.; Yu, E.D.; Barra, C.M.; Alari-Pahissa, E.; Girardi, E.; Harrak, Y.; Lauzurica, P.; Llebaria, A.; Zajonc, D.M.; Akbari, O.; et al. Structural and functional characterization of a novel nonglycosidic type i NKT agonist with immunomodulatory properties. J. Immunol. 2012, 188, 2254–2265. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Anderson, B.L.; Teyton, L.; Bendelac, A.; Savage, P.B. Stimulation of Natural Killer T Cells by Glycolipids. Molecules 2013, 18, 15662-15688. https://doi.org/10.3390/molecules181215662
Anderson BL, Teyton L, Bendelac A, Savage PB. Stimulation of Natural Killer T Cells by Glycolipids. Molecules. 2013; 18(12):15662-15688. https://doi.org/10.3390/molecules181215662
Chicago/Turabian StyleAnderson, Brian L., Luc Teyton, Albert Bendelac, and Paul B. Savage. 2013. "Stimulation of Natural Killer T Cells by Glycolipids" Molecules 18, no. 12: 15662-15688. https://doi.org/10.3390/molecules181215662
APA StyleAnderson, B. L., Teyton, L., Bendelac, A., & Savage, P. B. (2013). Stimulation of Natural Killer T Cells by Glycolipids. Molecules, 18(12), 15662-15688. https://doi.org/10.3390/molecules181215662