Ligand Binding Affinities of Arctigenin and Its Demethylated Metabolites to Estrogen Receptor Alpha

Abstract
:1. Introduction
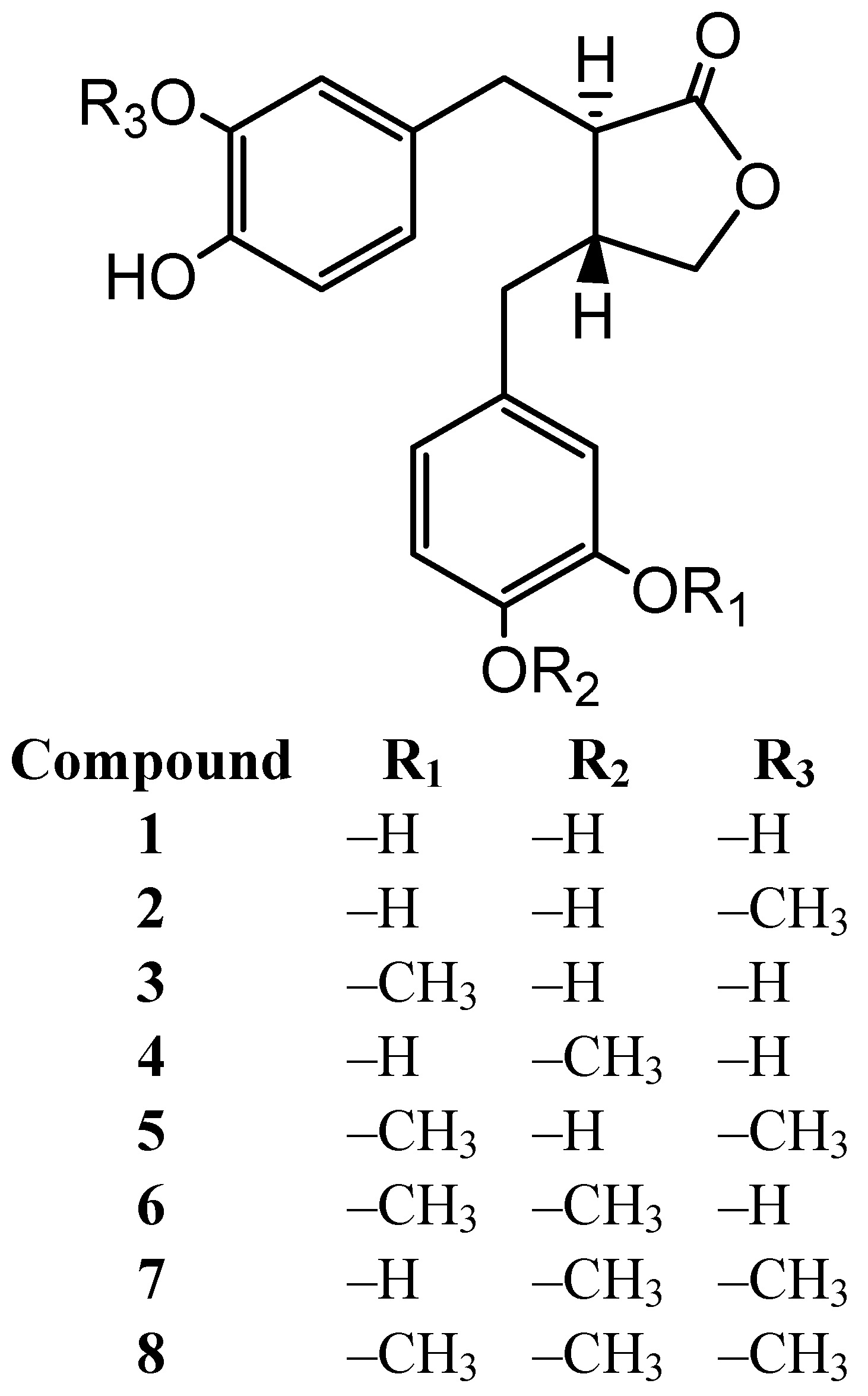
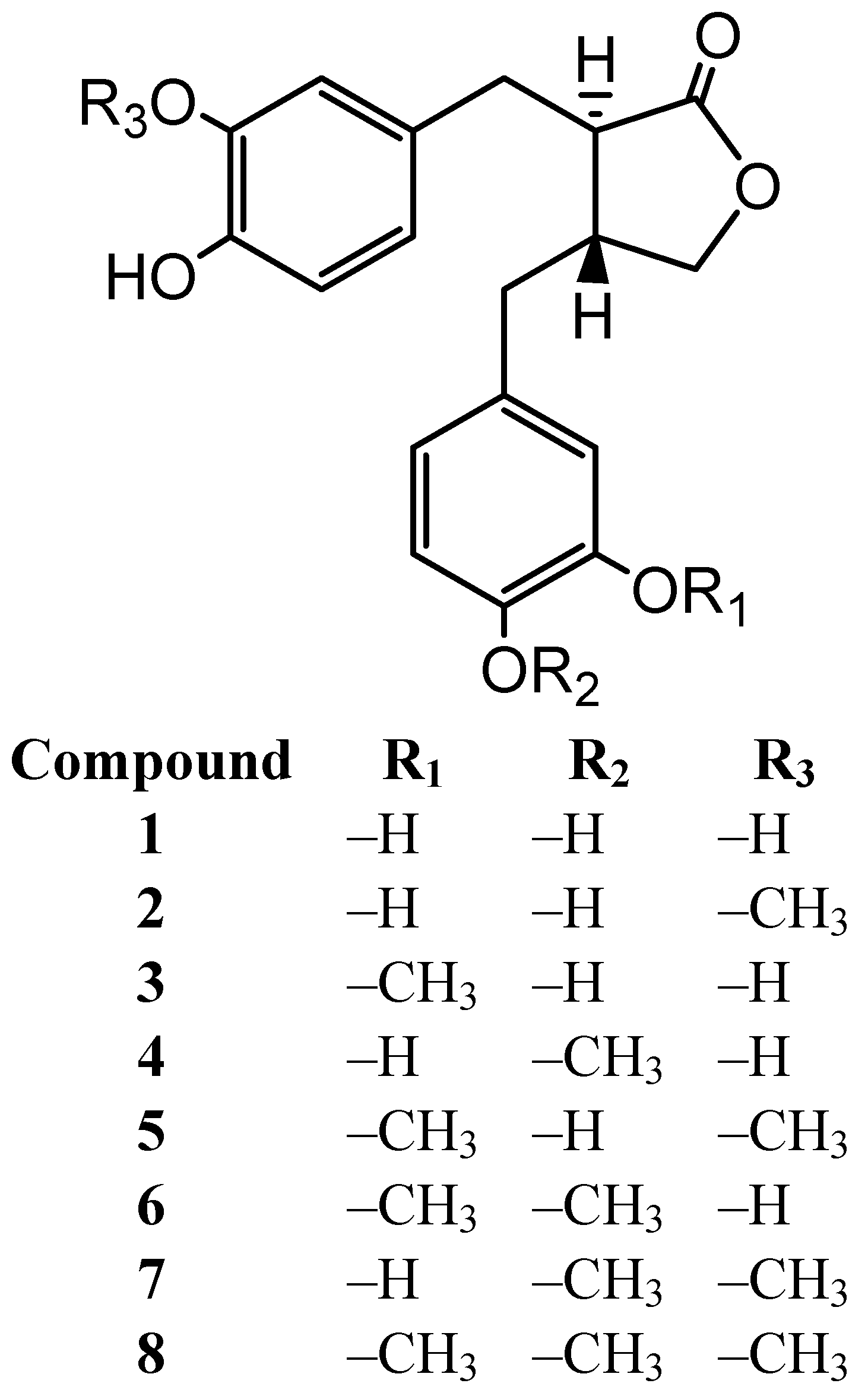
2. Results and Discussion

{kind=link}
| Compound | % Binding to ERα | IC50 | RBA | |||
|---|---|---|---|---|---|---|
| 10−5 M | 10−4 M | 5 × 10−4 M | 5 × 10−3 M | |||
| 1 | 5.6 ± 3.3 * | 13.7 ± 0.7 ** | 34.8 ± 1.3 ** | 65.3 ± 0.2 ** | 1.6 × 10−3 M | 1.2 × 10−3 |
| 2 | 1.4 ± 1.2 | 13.5 ± 2.0 ** | 44.4 ± 0.9 ** | 67.7 ± 0.2 ** | 7.9 × 10−4 M | 2.5 × 10−3 |
| 3 | 2.2 ± 1.6 | 14.1 ± 1.3 ** | 25.8 ± 0.8 ** | 67.5 ± 0.2 ** | 1.9 × 10−3 M | 1.0 × 10−3 |
| 4 | 1.0 ± 1.4 | −0.5 ± 0.7 | 19.6 ± 2.3 ** | 63.2 ± 0.3 ** | 2.5 × 10−3 M | 7.9 × 10−4 |
| 5 | 0.4 ± 0.9 | 18.1 ± 9.0 ** | 0.1 ± 2.6 | 17.6 ± 1.1 ** | >5 × 10−3 M | – |
| 6 | −2.3 ± 0.9 | 1.4 ± 2.4 | 16.6 ± 0.9 ** | 64.5 ± 0.2 ** | 2.5 × 10−3 M | 7.9 × 10−4 |
| 7 | 3.6 ± 0.6 | 1.2 ± 1.7 | −4.2 ± 0.8 | 30.1 ± 1.7 ** | >5 × 10−3 M | – |
| 8 | −1.4 ± 0.8 | −4.3 ± 0.8 | −4.1 ± 3.5 | 19.4 ± 1.5 ** | >5 × 10−3 M | – |
3. Experimental
3.1. Chemicals
3.2. Ligand Binding Screen
3.3. Statistical Analysis
4. Conclusions
Conflicts of Interest
- Sample Availability: Samples of the compounds were isolated by authors. Not all the compounds are available.
References
- Bingham, S.A.; Atkinson, C.; Liggins, J.; Bluck, L.; Coward, A. Phyto-oestrogens: Where are we now? Br. J. Nutr. 1998, 79, 393–406. [Google Scholar] [CrossRef]
- Rowland, I.; Faughnan, M.; Hoey, L.; Wahala, K.; Williamson, G.; Cassidy, A. Bioavailability of phyto-oestrogens. Br. J. Nutr. 2003, 89, S45–S58. [Google Scholar]
- Jin, J.S.; Tobo, T.; Chung, M.H.; Ma, C.M.; Hattori, M. Transformation of trachelogenin, an aglycone of tracheloside from safflower seeds, to phytoestrogenic (−)-enterolactone by human intestinal bacteria. Food Chem. 2012, 134, 74–80. [Google Scholar] [CrossRef]
- Adlercreutz, H.; Mousavi, Y.; Clark, J.; Hockerstedt, K.; Hamalainen, E.; Wahala, K.; Makela, T.; Hase, T. Dietary phytoestrogens and cancer: In vitro and in vivo studies. J. Steroid. Biochem. Mol. Biol. 1992, 41, 331–337. [Google Scholar] [CrossRef]
- Mueller, S.O.; Simon, S.; Chae, K.; Metzler, M.; Korach, K.S. Phytoestrogens and their human metabolites show distinct agonistic and antagonistic properties on estrogen receptor alpha (ERalpha) and ERbeta in human cells. Toxicol. Sci. 2004, 80, 14–25. [Google Scholar] [CrossRef]
- Schottner, M.; Gansser, D.; Spiteller, G. Lignans from the roots of Urtica dioica and their metabolites bind to human sex hormone binding globulin (SHBG). Planta Med. 1997, 63, 529–532. [Google Scholar] [CrossRef]
- Xie, L.H.; Akao, T.; Hamasaki, K.; Deyama, T.; Hattori, M. Biotransformation of pinoresinol diglucoside to mammalian lignans by human intestinal microflora, and isolation of Enterococcus faecalis strain PDG-1 responsible for the transformation of (+)-pinoresinol to (+)-lariciresinol. Chem. Pharm. Bull. 2003, 51, 508–515. [Google Scholar] [CrossRef]
- Wang, L.Q.; Meselhy, M.R.; Li, Y.; Qin, G.W.; Hattori, M. Human intestinal bacteria capable of transforming secoisolariciresinol diglucoside to mammalian lignans, enterodiol and enterolactone. Chem. Pharm. Bull. 2000, 48, 1606–1610. [Google Scholar] [CrossRef]
- Penalvo, J.L.; Heinonen, S.M.; Aura, A.M.; Adlercreutz, H. Dietary sesamin is converted to enterolactone in humans. J. Nutr. 2005, 135, 1056–1062. [Google Scholar]
- Xie, L.H.; Ahn, E.M.; Akao, T.; Abdel-Hafez, A.A.M.; Nakamura, N.; Hattori, M. Transformation of arctiin to estrogenic and antiestrogenic substances by human intestinal bacteria. Chem. Pharm. Bull. 2003, 51, 378–384. [Google Scholar] [CrossRef]
- Jin, J.S.; Hattori, M. A new mammalian lignan precursor, asarinin. Food Chem. 2011, 124, 895–899. [Google Scholar] [CrossRef]
- Jin, J.S.; Hattori, M. Human intestinal bacterium, strain END-2 is responsible for demethylation as well as lactonization during plant lignan metabolism. Biol. Pharm. Bull. 2010, 33, 1443–1447. [Google Scholar] [CrossRef]
- Jin, J.S.; Hattori, M. Further studies on a human intestinal bacterium Ruminococcus sp. END-1 for transformation of plant lignans to mammalian lignans. J. Agric. Food Chem. 2009, 57, 7537–7542. [Google Scholar] [CrossRef]
- Clavel, T.; Henderson, G.; Alpert, C.A.; Philippe, C.; Rigottier-Gois, L.; Doré, J.; Blaut, M. Intestinal bacterial communities that produce active estrogen-like compounds enterodiol and enterolactone in humans. Appl. Environ. Microbiol. 2005, 71, 6077–6085. [Google Scholar] [CrossRef]
- Clavel, T.; Henderson, G.; Engst, W.; Doré, J.; Blaut, M. Phylogeny of human intestinal bacteria that activate the dietary lignan secoisolariciresinol diglucoside. FEMS Microbiol. Ecol. 2006, 55, 471–478. [Google Scholar] [CrossRef]
- Clavel, T.; Borrmann, D.; Braune, A.; Doré, J.; Blaut, M. Occurrence and activity of human intestinal bacteria involved in the conversion of dietary lignans. Anaerobe 2006, 12, 140–147. [Google Scholar] [CrossRef]
- Clavel, T.; Lippman, R.; Gavini, F.; Doré, J.; Blaut, M. Clostridium saccharogumia sp. nov. and Lactonifactor longoviformis gen. nov., sp. nov., two novel human faecal bacteria involved in the conversion of the dietary phytoestrogen secoisolariciresinol diglucoside. Syst. Appl. Microbiol. 2007, 30, 16–26. [Google Scholar] [CrossRef]
- Jin, J.S.; Kakiuchi, N.; Hattori, M. Enantioselective oxidation of enterodiol to enterolactone by human intestinal bacteria. Biol. Pharm. Bull. 2007, 30, 2204–2206. [Google Scholar] [CrossRef]
- Jin, J.S.; Zhao, Y.F.; Nakamura, N.; Akao, T.; Kakiuchi, N.; Min, B.S.; Hattori, M. Enantioselective dehydroxylation of enterodiol and enterolactone precursors by human intestinal bacteria. Biol. Pharm. Bull. 2007, 30, 2113–2119. [Google Scholar] [CrossRef]
- Jin, J.S.; Zhao, Y.F.; Nakamura, N.; Akao, T.; Kakiuchi, N.; Hattori, M. Isolation and characterization of a human intestinal bacterium, Eubacterium sp. ARC-2, capable of demethylating arctigenin, in the essential metabolic process to enterolactone. Biol. Pharm. Bull. 2007, 30, 904–911. [Google Scholar] [CrossRef]
- Nose, M.; Fujimoto, T.; Takeda, T.; Nishibe, S.; Ogihara, Y. Structural transformation of lignan compounds in rat gastrointestinal tract. Planta Med. 1992, 58, 520–523. [Google Scholar] [CrossRef]
- Pfitscher, A.; Reiter, E.; Jungbauer, A. Receptor binding and transactivation activities of red clover isoflavones and their metabolites. J. Steroid Biochem. Mol. Biol. 2008, 112, 87–94. [Google Scholar] [CrossRef]
- Klopman, G.; Chakravarti, S.K. Structure-activity relationship study of a diverse set of estrogen receptor ligands (I) using MultiCASE expert system. Chemosphere 2003, 51, 445–459. [Google Scholar] [CrossRef]
- Fang, H.; Tong, W.; Shi, L.M.; Blair, R.; Perkins, R.; Branham, W.; Hass, B.S.; Xie, Q.; Dial, S.L.; Moland, C.L.; Sheehan, D.M. Structure-activity relationships for a large diverse set of natural, synthetic, and environmental estrogens. Chem. Res. Toxicol. 2001, 14, 280–294. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jin, J.-S.; Lee, J.-H.; Hattori, M. Ligand Binding Affinities of Arctigenin and Its Demethylated Metabolites to Estrogen Receptor Alpha. Molecules 2013, 18, 1122-1127. https://doi.org/10.3390/molecules18011122
Jin J-S, Lee J-H, Hattori M. Ligand Binding Affinities of Arctigenin and Its Demethylated Metabolites to Estrogen Receptor Alpha. Molecules. 2013; 18(1):1122-1127. https://doi.org/10.3390/molecules18011122
Chicago/Turabian StyleJin, Jong-Sik, Jong-Hyun Lee, and Masao Hattori. 2013. "Ligand Binding Affinities of Arctigenin and Its Demethylated Metabolites to Estrogen Receptor Alpha" Molecules 18, no. 1: 1122-1127. https://doi.org/10.3390/molecules18011122
APA StyleJin, J.-S., Lee, J.-H., & Hattori, M. (2013). Ligand Binding Affinities of Arctigenin and Its Demethylated Metabolites to Estrogen Receptor Alpha. Molecules, 18(1), 1122-1127. https://doi.org/10.3390/molecules18011122