Assessment of Worldwide Genetic Diversity of Siberian Wild Rye (Elymus sibiricus L.) Germplasm Based on Gliadin Analysis

Abstract
:1. Introduction
2. Results and Discussion

{kind=link}
{kind=link}
| Source | Accession codes |
|---|---|
| Mongolia | PI610850, PI610857, PI610860, PI610862, PI610866, PI610876, PI610886, PI628726, PI634230, PI634231 |
| Kazakhstan | PI314619, PI598773, PI598778, PI598788 |
| Alaska, US | PI372696, PI348916 |
| British Columbia, Canada | PI372541 |
| Siberia, Russia | PI326266 **, PI628699, PI598478, PI369236, PI611013, PI598800, PI634228, PI406467 **, PI611020, PI315427, PI598789, PI345600 **, PI345599 **, PI326267 **, PI315428, PI598774 |
| Xinjiang, China | Y0811, Y004, Y0860, Y0771, Y0766, Y0760, Y2027, Y0822, Y0473, Y013, Y0877, Y1914, Y2024, Y1823, Y0909, Y0486, Y005, PI628677, PI499462 |
| Inner Mongolia, China | PI499453, PI499455, PI499457 *, PI499459 * |
| Sichuan, Qinghai-Tibet Plateau, China | 205124, 204145, 204155, 205189, 205201, 205179, 204081, 204089, 204119, 205215, 205156, 205226, Chuancao No.2 **, Chuancao No.1**, 205171, 205172 |
| Gansu, Qinghai-Tibet Plateau, China | Y2906, PI499456 *, PI499458 *, PI499460 *, PI499461, PI531671, PI636676, 204451 |
| Qinghai, Qinghai-Tibet Plateau, China | PI504462, PI504463, PI531669, 204441 |
| Tibet, Qinghai-Tibet Plateau, China | 204404, 204251 |
2.1. Polymorphism of Gliadin Bands

| Zones | Total bands | Polymorphic bands | Po1ymorphism percentage (%) | Shannon index (Ho) | Simpson index (He) |
|---|---|---|---|---|---|
| α | 10 | 10 | 100 | 0.431 | 0.274 |
| β | 9 | 7 | 77.8 | 0.472 | 0.318 |
| γ | 12 | 12 | 100 | 0.557 | 0.376 |
| ω | 21 | 18 | 85.7 | 0.401 | 0.253 |
| Total | 52 | 47 | 90.4 | 0.460 | 0.302 |
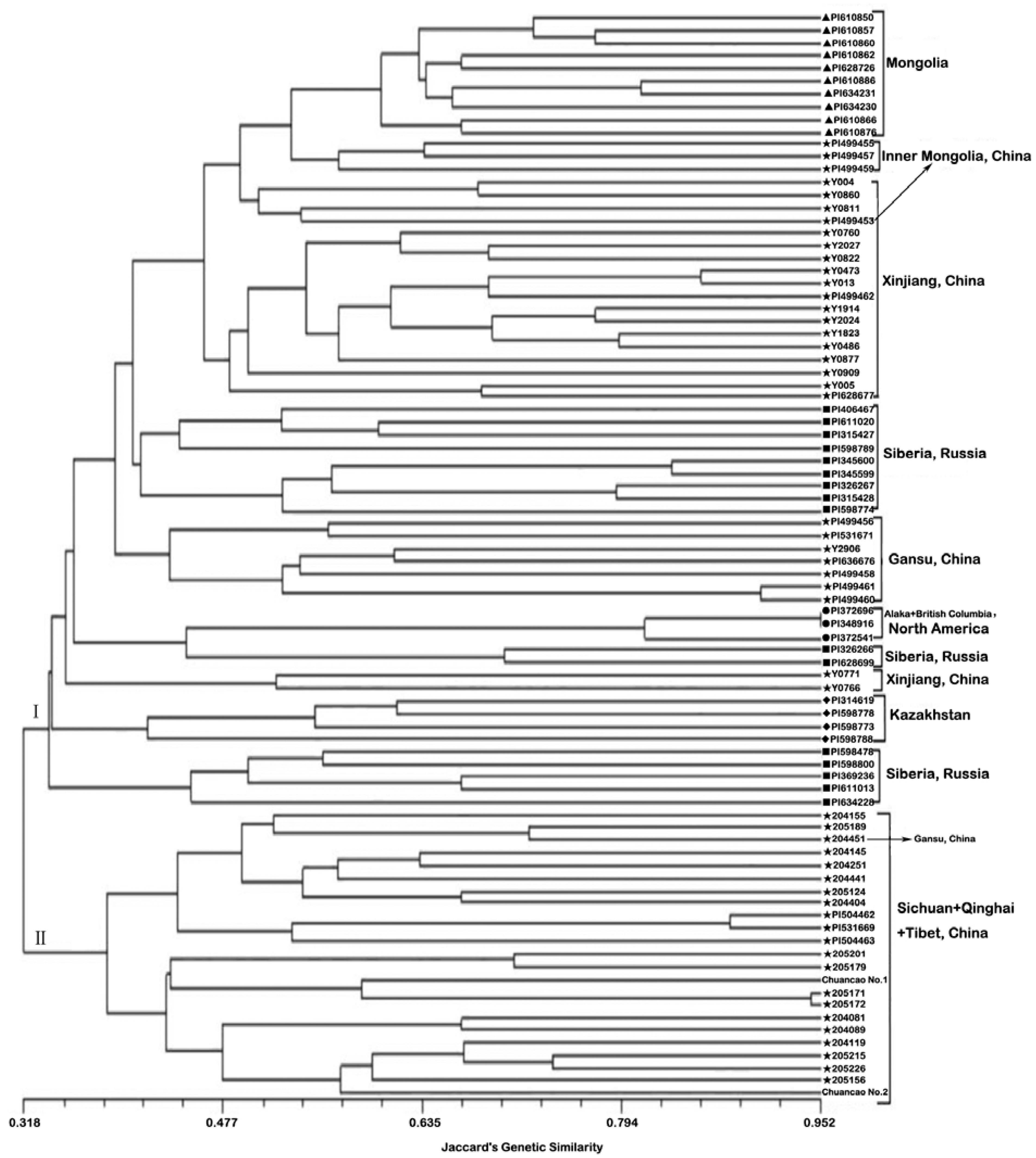
2.2. Genetic Similarities and Cluster Analysis
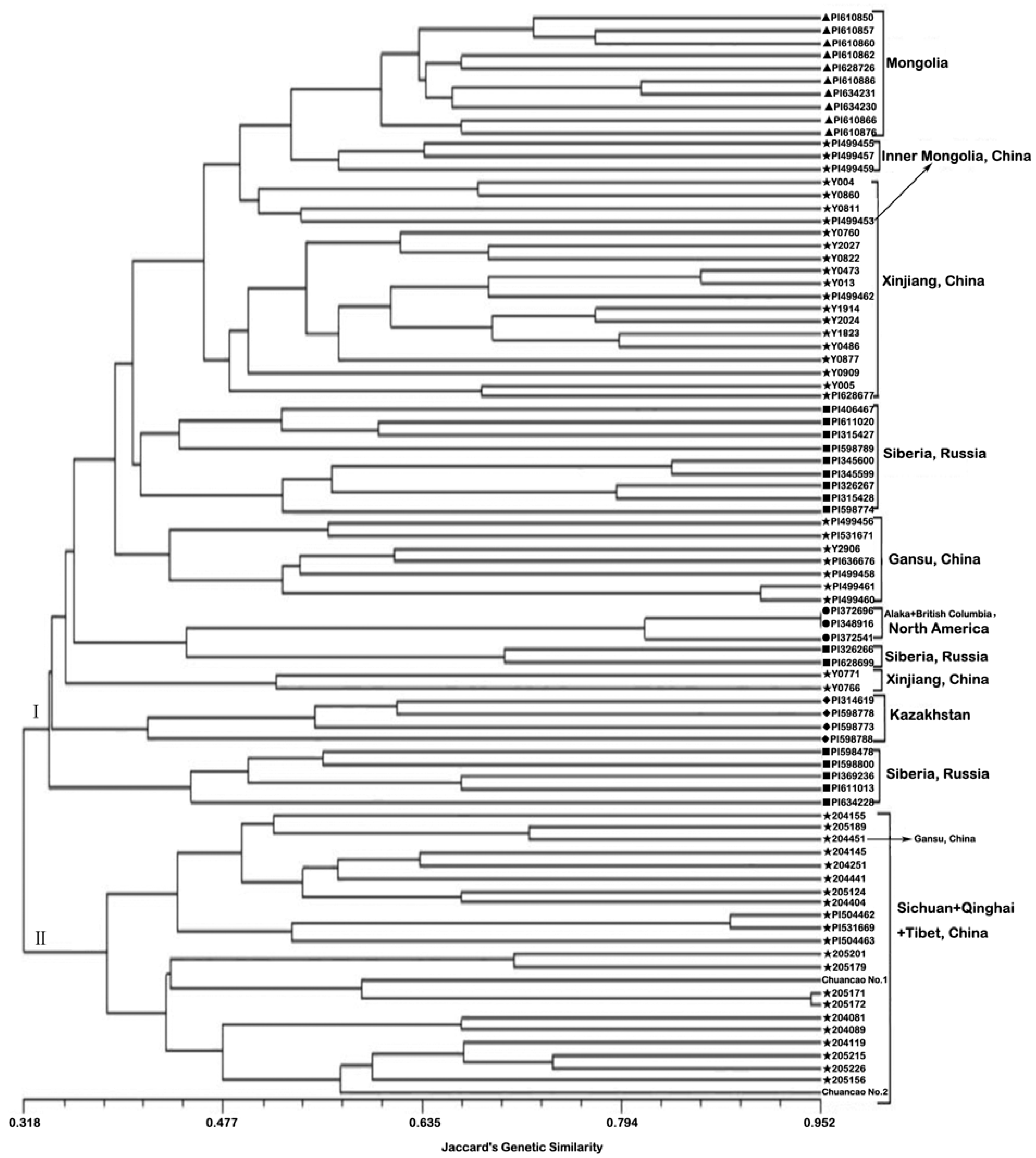
2.3. Genetic Differentiation of Geographic Groups
| Shannon Index | North America | Russia | Mongolia | Kazakhstan | China |
|---|---|---|---|---|---|
| Ho | 0.053 | 0.340 | 0.205 | 0.240 | 0.446 |
| Hpop = Σ Ho/n | 0.257 | ||||
| Hsp | 0.460 | ||||
| Hwithin = Hpop/Hsp | 0.559 | ||||
| Hbetween = (Hsp − Hpop)/Hsp | 0.441 | ||||
| Shannon Index | Xinjiang | Inner Mongolia | Gansu | Sichuan | Qinghai | Tibet |
|---|---|---|---|---|---|---|
| Ho | 0.316 | 0.201 | 0.356 | 0.374 | 0.280 | 0.173 |
| Hpop = Σ Ho/n | 0.283 | |||||
| Hsp | 0.446 | |||||
| Hwithin = Hpop/Hsp | 0.634 | |||||
| Hbetween = (Hsp − Hpop)/Hsp | 0.366 | |||||
3. Experimental
3.1. Plant Materials
3.2. Acid Ployacrylamide Gel Electrophoresis (A-PAGE)
3.3. Data Analysis
4. Conclusions
Acknowledgments
References and Notes
- Dewey, D.R. The Genomic System of Classification as a Guide to Intergeneric Hybridization with the Perennial Triticeae. In Gene Manipulation in Plant Improvement; Gustafson, J.P., Ed.; Plenum: New York, NY, USA, 1984; pp. 209–279. [Google Scholar]
- Dewey, D.R. Cytogenetics of Elymus sibiricus and its hybrids with Agropyron tauri, Elymus canadensis, and Agropyron caninu. Bot. Gaz. 1974, 135, 80–87. [Google Scholar]
- Bowden, W.M.; Cody, W.J. Recognition of Elymus sibiricus L. from Alaska and the District of Mackenzie. Bull. Torrey Bot. Club 1961, 88, 153–155. [Google Scholar] [CrossRef]
- Lu, B.R. Biosystematic Investigations of Asiatic Wheatgrasses—Elymus L. (Triticeae: Poaceae). Ph.D. Thesis, The Swedish University of Agricultural Sciences, Svalöv, Sweden, 1993. [Google Scholar]
- Yan, J.J.; Bai, S.Q.; Ma, X.; Gan, Y.M.; Zhang, J.B. Genetic diversity of Elymus sibiricus and its breeding in China (In Chinese with English abstract). Chin. Bull. Bot. 2007, 24, 226–231. [Google Scholar]
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed storage proteins: Structures and biosynthesis. Plant Cell. 1995, 7, 945–956. [Google Scholar]
- Chen, L.; Fischer, H.; Jensen, U. Accumulation of seed storage proteins and the taxonomy of Poaceae. Plant Syst. Evol. 1997, 206, 243–257. [Google Scholar]
- Alvarez, J.B.; Broccoli, A.; Martin, L.M. Variability and genetic diversity for gliadins in natural populations of Hordeum chilense Roem. et Schult. Genet. Resour. Crop Evol. 2006, 53, 1419–1425. [Google Scholar] [CrossRef]
- Jaradat, A.A. Grain protein variability among populations of wild barley (Hordeum spontaneum C. Koch.) from Jordan. Theor. Appl. Genet. 1991, 83, 164–168. [Google Scholar]
- Alvarez, J.B.; Moral, A.; Martin, L.M. Polymorphism and genetic diversity for the seed storage proteins in Spanish cultivated einkorn wheat (Triticum monococcum L. ssp. monococcum). Genet. Resour. Crop Evol. 2006, 53, 1061–1067. [Google Scholar] [CrossRef]
- Caballero, L.; Martin, L.M.; Alvarez, J.B. Variation and genetic diversity for gliadins in Spanish spelt wheat accessions. Genet. Resour. Crop Evol. 2004, 51, 679–686. [Google Scholar]
- Metakovsky, E.V.; Gemez, M.; Vazquez, J.F.; Carrillo, J.M. High genetic diversity of Spanish common wheat as judged from gliadin alleles. Plant Breed. 2000, 119, 37–42. [Google Scholar]
- Syros, T.; Yupsanis, T.; Economou, A. Fractionation and electrophoretic patterns of storage proteins of Ebenus cretica. A prelimnary survey as a tool in taxonomy. Biol. Plant. 2003, 46, 435–443. [Google Scholar] [CrossRef]
- Shewry, P.R.; Tatham, A.S.; Halford, N.G. The Prolamins of the Triticeae. In Seed Proteins; Shewry, P.R., Casey, R., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1999; pp. 35–78. [Google Scholar]
- Chen, M.Z. α-Gliadin and Wheat Breeding (In Chinese with English abstract). Master Thesis, The Shandong University, Jinan, China, 2008. [Google Scholar]
- Yan, J.J.; Bai, S.Q.; Zhang, X.Q.; Zhang, C.B.; You, M.H. Genetic diversity of gliadin in native Elymus sibiricus from Qinghai-Tibetan Plateau in China (In Chinese with English abstract). J. Agric. Biotech. 2009, 17, 891–901. [Google Scholar]
- Miao, J.M.; Zhang, X.Q.; Chen, S.Y.; Ma, X.; Chen, Z.H.; Zhong, J.C.; Bai, S.Q. Gliadin analysis of Elymus nutans Griseb. from the Qinghai-Tibetan Plateau and Xinjiang, China. Grassland Sci. 2011, 57, 127–134. [Google Scholar] [CrossRef]
- Gerus, D.E.; Agafonov, A.V. Genetic diversity in natural Elymus fibrosus (Triticeae: Poaceae) populations assessed using endosperm storage proteins. Russ. J. Genet. 2012, 2, 58–64. [Google Scholar]
- Yan, J.J.; Bai, S.Q.; Chang, D.; You, M.H.; Zhang, C.B.; Li, D.X. Genetic diversity of wild Elymus sibiricus L. germplasm from Qinghai-Tibetan Plateau in China detected by SSR markers (In Chinese with English abstract). Chin. Agric. Sci. Bull. 2010, 26, 26–33. [Google Scholar]
- Ma, X.; Zhang, X.Q.; Zhou, Y.H.; Bai, S.Q.; Liu, W. Assessing genetic diversity of Elymus sibiricus (Poaceae: Triticeae) populations from Qinghai-Tibet plateau by ISSR markers. Biochem. Syst. Ecol. 2008, 36, 514–522. [Google Scholar]
- Yan, J.J.; Bai, S.Q.; Zhang, X.Q.; You, M.H.; Zhang, C.B.; Li, D.X.; Zeng, Y. Genetic diversity of wild Elymus sibiricus germplasm from the Qinghai-Tibetan Plateau in China detected by SRAP markers (In Chinese with English abstract). Acta Prataculturae Sinica 2010, 19, 173–183. [Google Scholar]
- Barkworth, M.E.; Campbell, J.J.N.; Salomon, B. Elymus. In Flora of North America; Barkworth, M.E., Capels, K.M.C., Long, S., Anderton, L.K., Piep, M., Eds.; Oxford University Press: New York, NY, USA, 2007; Volume 24, pp. 288–343. [Google Scholar]
- Johnson, R.C.; Johnson, W.J.; Golob, C.T.; Nelson, M.C.; Soreng, R.J. Characterization of the USDA Poa pratensis collection using RAPD markers and agronomic descriptors. Genet. Resour. Crop Evol. 2002, 49, 349–361. [Google Scholar]
- Nevo, E. Genetic diversity in wild cereals: Regional and local studies and their bearing on conservation ex situ and in situ. Genet. Resour. Crop. Evol. 1998, 45, 355–370. [Google Scholar]
- Schoen, D.J.; Brown, A.H.D. Intraspecific variation in population gene diversity and effective population size correlates with the mating system in plants. Proc. Natl. Acad. Sci. USA 1991, 88, 4494–4497. [Google Scholar]
- Godt, M.J.W.; Johnson, B.R.; Hamrick, J.L. Genetic diversity and population size in four rare southern appalachian plant species. Conserv. Biol. 1996, 10, 796–805. [Google Scholar]
- Sun, M. Effects of population size, mating system, and evolutionary origin on genetic diversity in Spiranthes sinensis and S. hongkongensis. Conserv. Biol. 1996, 10, 785–795. [Google Scholar]
- Draper, S.R. ISTA variety committee report of the working group for biochemical tests for cultivar identification 1983–1986. Seed Sci. Technol. 1987, 15, 431–434. [Google Scholar]
- Bushuk, W.; Zillman, R.R. Wheat cultivar identification by gliadin electrophoregram. I. Apparatus, method, and nomenclauture. Can. J. Plant Sci. 1978, 58, 505–515. [Google Scholar] [CrossRef]
- Wachira, F.N.; Waugh, R.; Hachett, C.A.; Powell, W. Detection of genetic diversity in tea (Camellia sinesis) using RAPD markers. Genome 1995, 38, 201–210. [Google Scholar]
- Smith, J.S.C.; Chin, E.C.L.; Shu, H.; Smith, O.S.; Wall, S.J.; Senior, M.L.; Mitchell, S.E.; Kresovich, S.; Ziegle, J. An evaluation of the utility of SSR loci as molecular markers in maize (Zea mays L.): Comparisons with data from RFLPs and pedigree. Theor. Appl. Genet. 1997, 95, 163–173. [Google Scholar] [CrossRef]
- Jaccard, P. Nonvelles Recherches sur la Distribution Florale. Bull. Soc. Vaud. Sci. Nat. 1908, 44, 223–270. [Google Scholar]
- Sokal, R.R.; Sneath, P.H.A. Principles of Numerical Taxonomy; W. H. Freeman: San Francisco, CA, USA, 1963. [Google Scholar]
- Rohlf, F.J. NTSYSpc. Numerical Taxonomy and Multivariate Analysis System, Version 2.2, Exeter Software, State University of New York: New York, NY, USA, 2010.
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar]
- Sample Availability: Samples of the 86 of E. sibiricus accessions are available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ma, X.; Chen, S.; Zhang, X.; Bai, S.; Zhang, C. Assessment of Worldwide Genetic Diversity of Siberian Wild Rye (Elymus sibiricus L.) Germplasm Based on Gliadin Analysis. Molecules 2012, 17, 4424-4434. https://doi.org/10.3390/molecules17044424
Ma X, Chen S, Zhang X, Bai S, Zhang C. Assessment of Worldwide Genetic Diversity of Siberian Wild Rye (Elymus sibiricus L.) Germplasm Based on Gliadin Analysis. Molecules. 2012; 17(4):4424-4434. https://doi.org/10.3390/molecules17044424
Chicago/Turabian StyleMa, Xiao, Shiyong Chen, Xinquan Zhang, Shiqie Bai, and Changbing Zhang. 2012. "Assessment of Worldwide Genetic Diversity of Siberian Wild Rye (Elymus sibiricus L.) Germplasm Based on Gliadin Analysis" Molecules 17, no. 4: 4424-4434. https://doi.org/10.3390/molecules17044424
APA StyleMa, X., Chen, S., Zhang, X., Bai, S., & Zhang, C. (2012). Assessment of Worldwide Genetic Diversity of Siberian Wild Rye (Elymus sibiricus L.) Germplasm Based on Gliadin Analysis. Molecules, 17(4), 4424-4434. https://doi.org/10.3390/molecules17044424