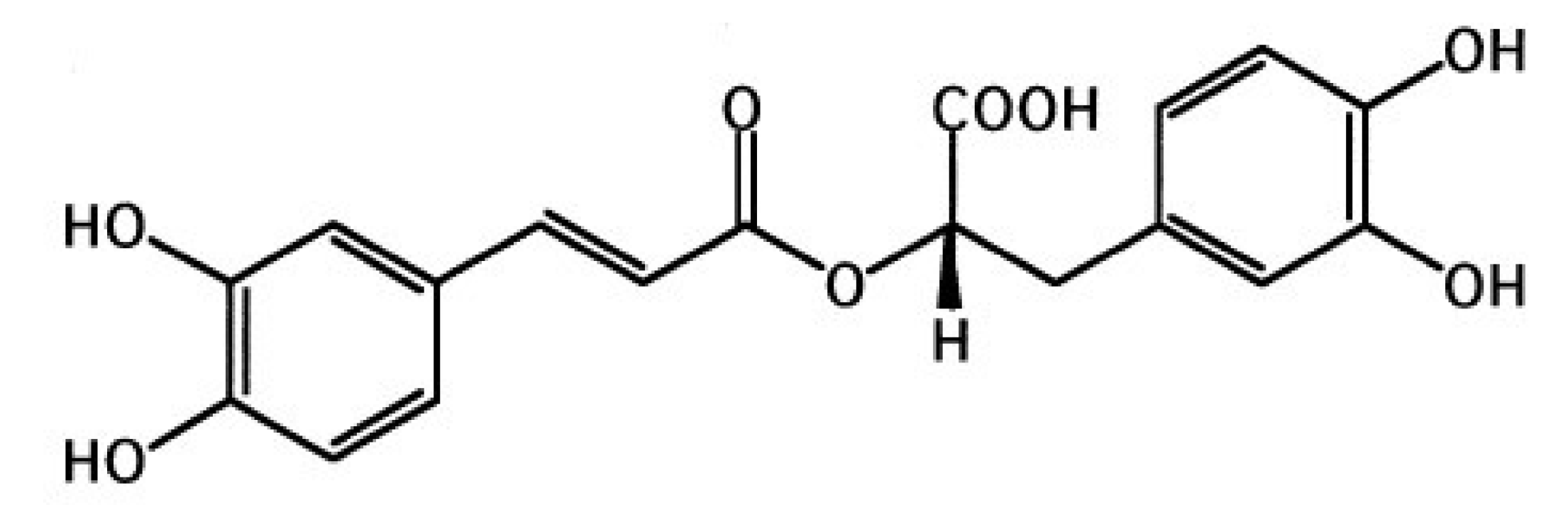
Effects of a Natural Prolyl Oligopeptidase Inhibitor, Rosmarinic Acid, on Lipopolysaccharide-Induced Acute Lung Injury in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
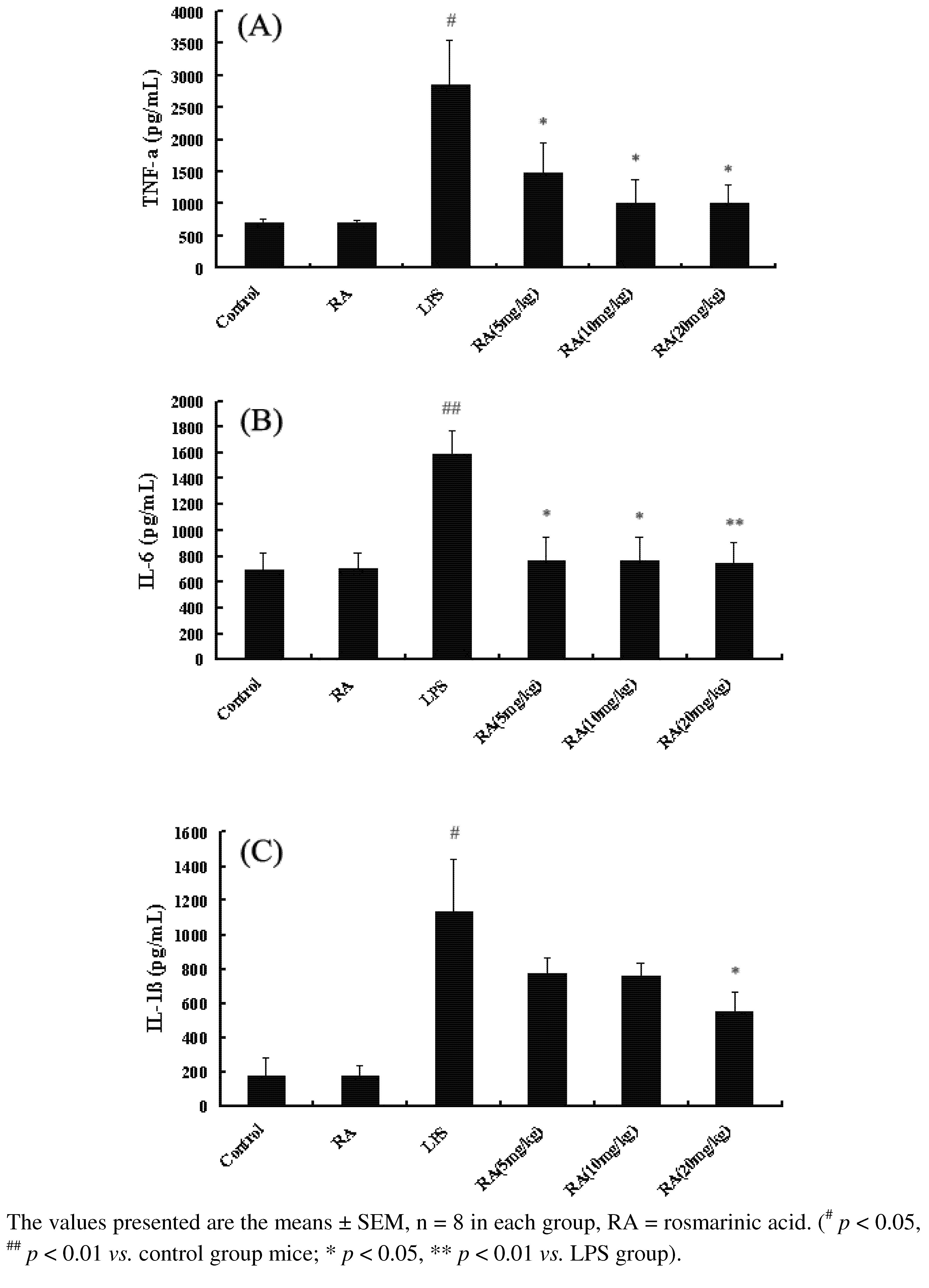
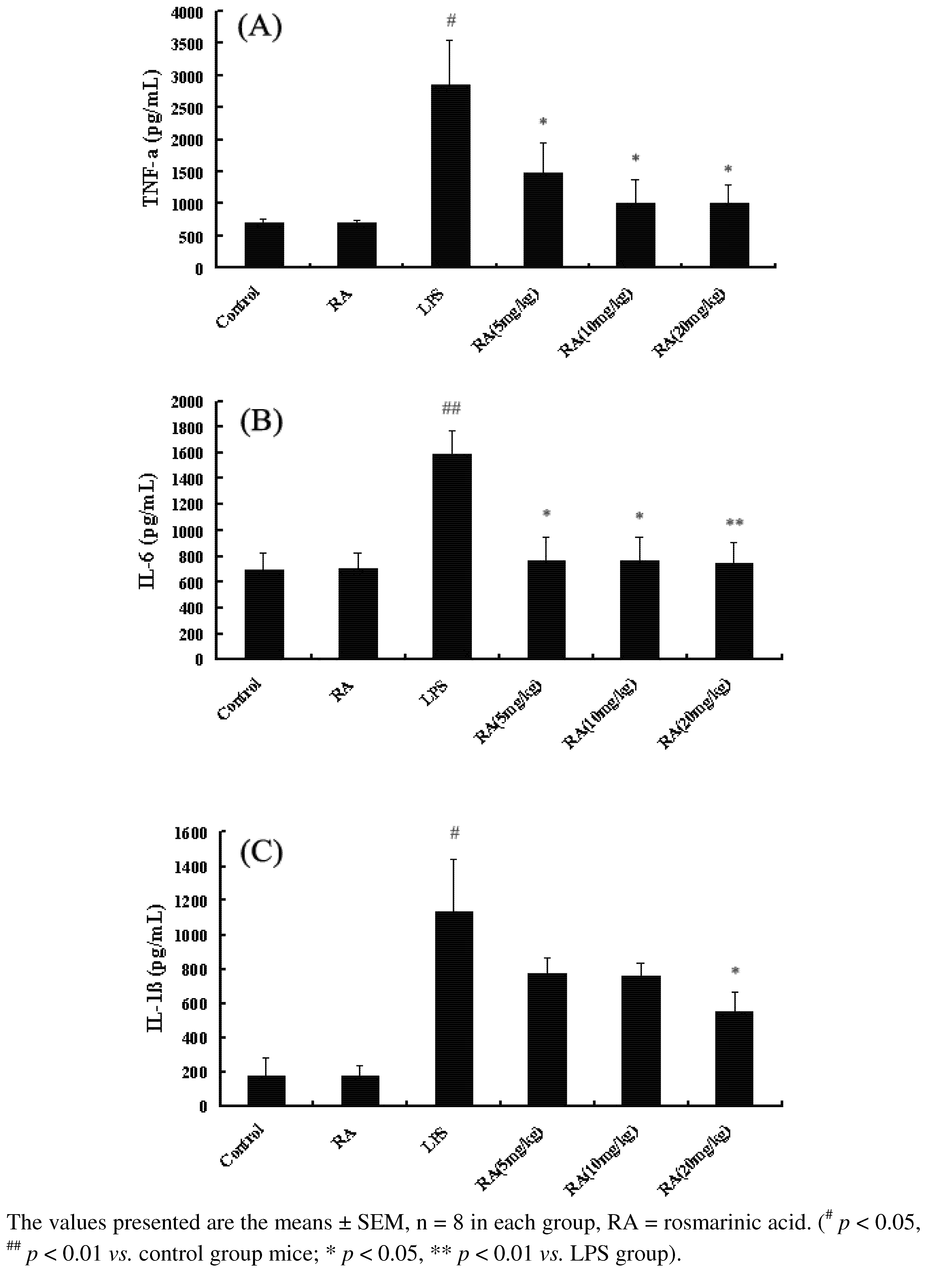
2.1. Effects of RA on Cytokine Production from Mice with ALI
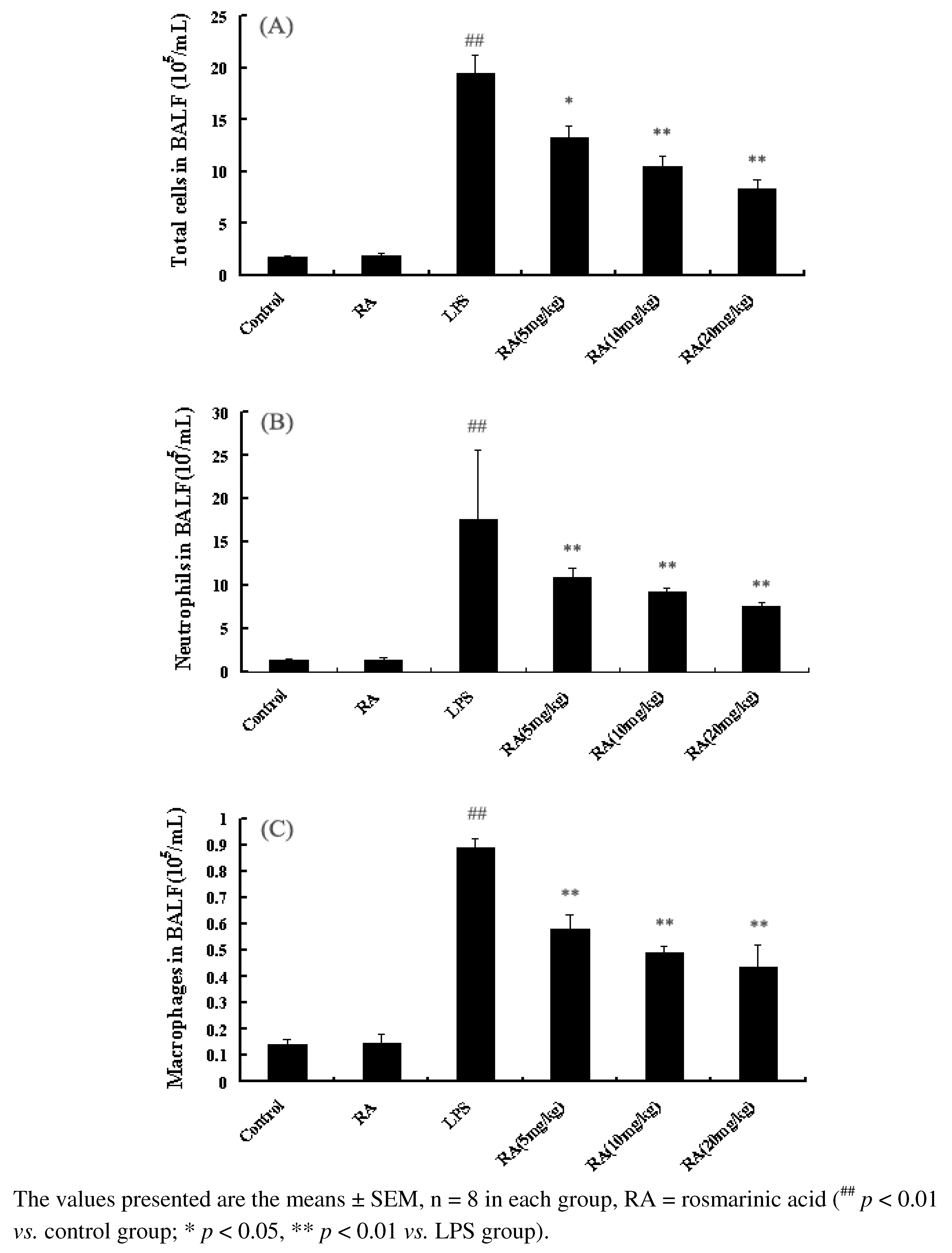
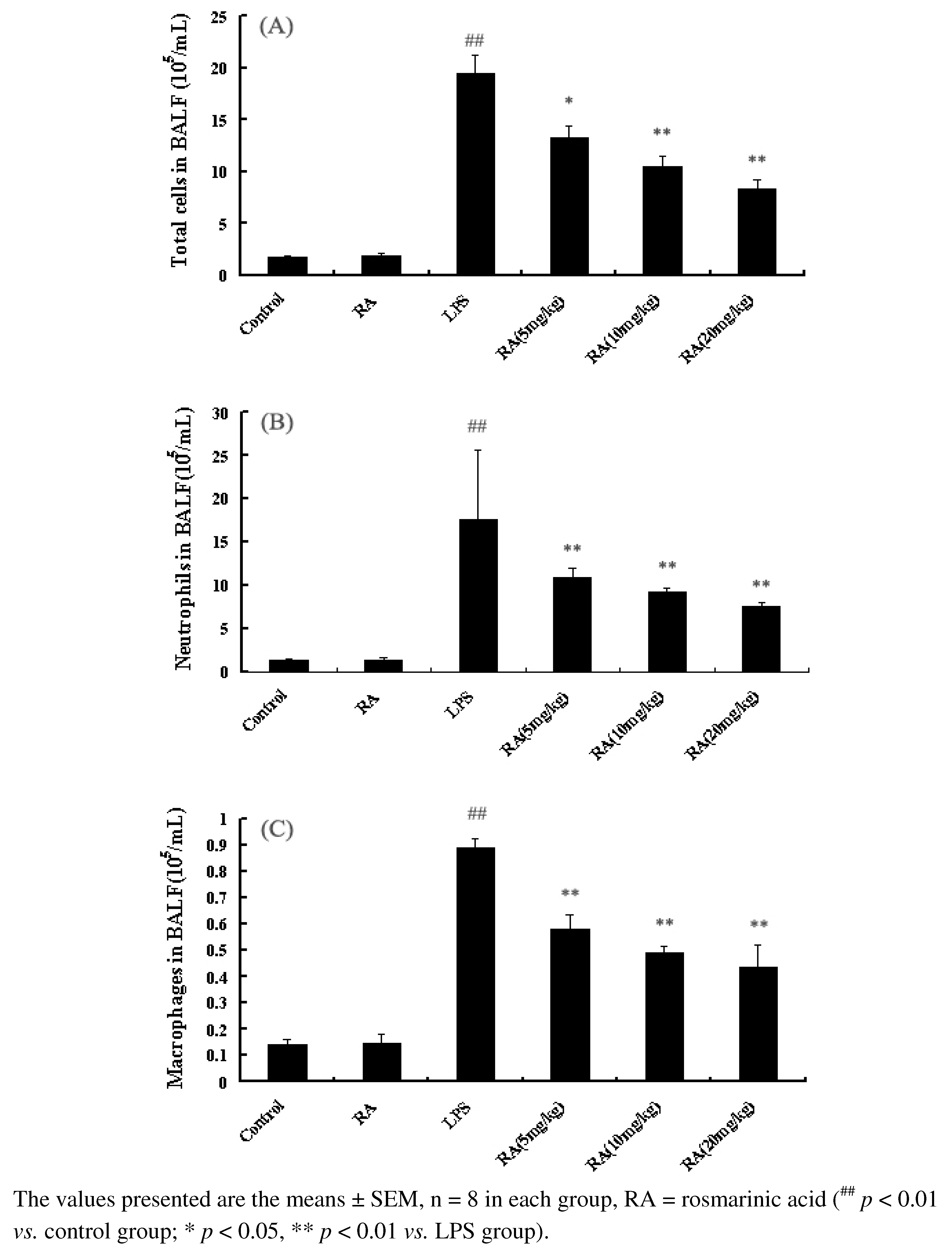
2.2. Effects of RA on Inflammatory Cell Count in the BALF from Mice with ALI
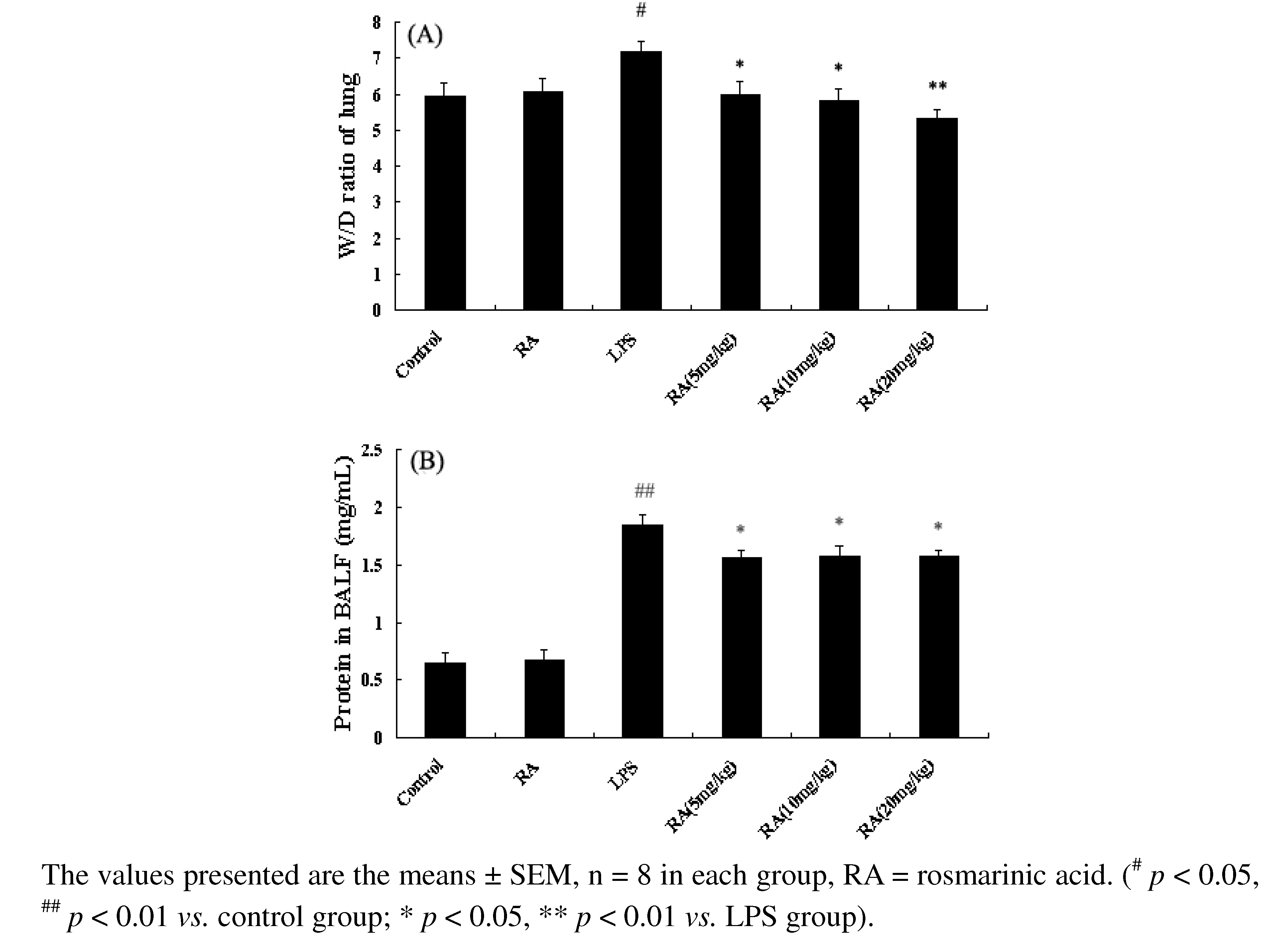
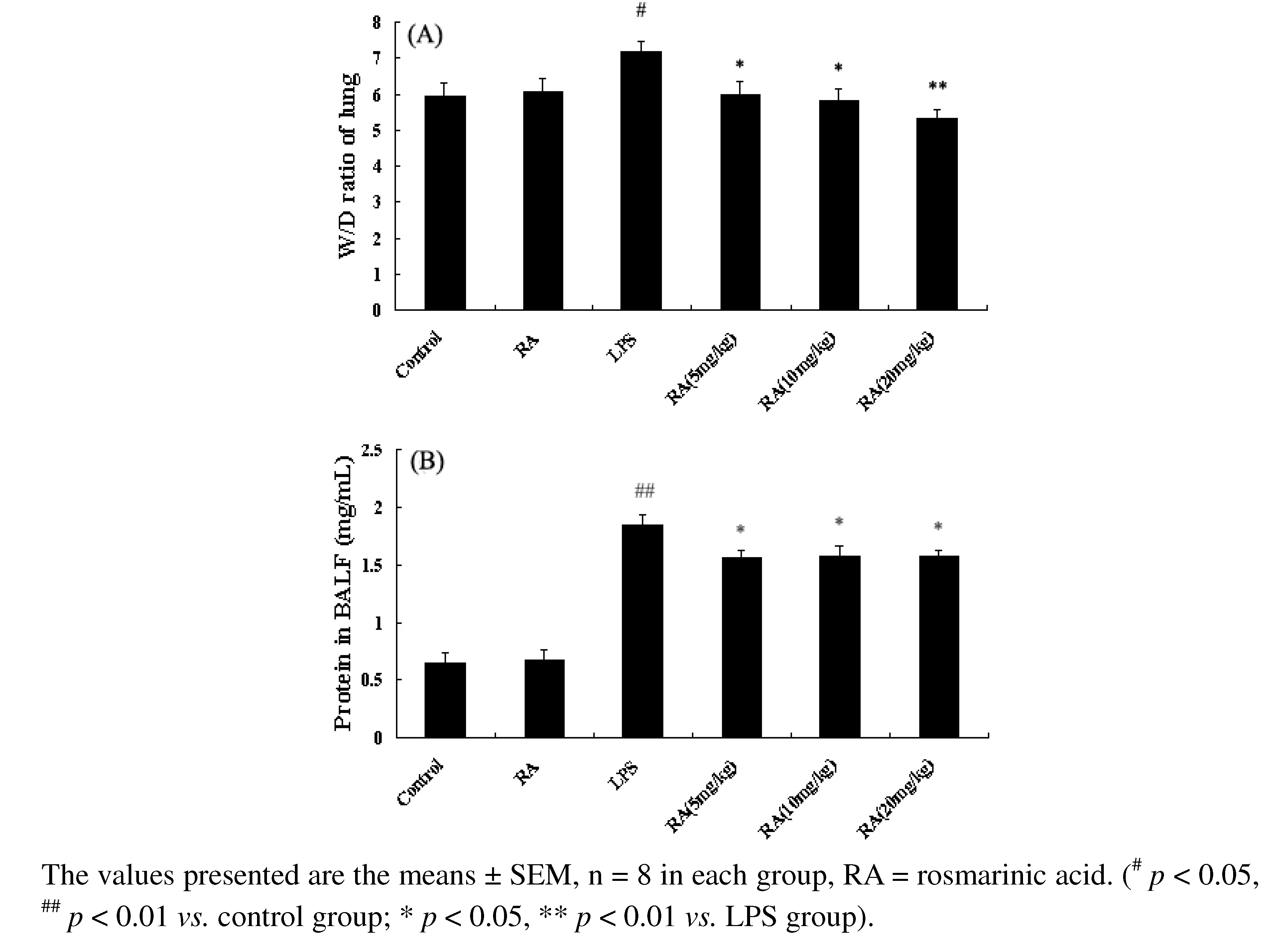
2.3. Effects of RA on Lung W/D Ratio in Mice with LPS-Induced ALI
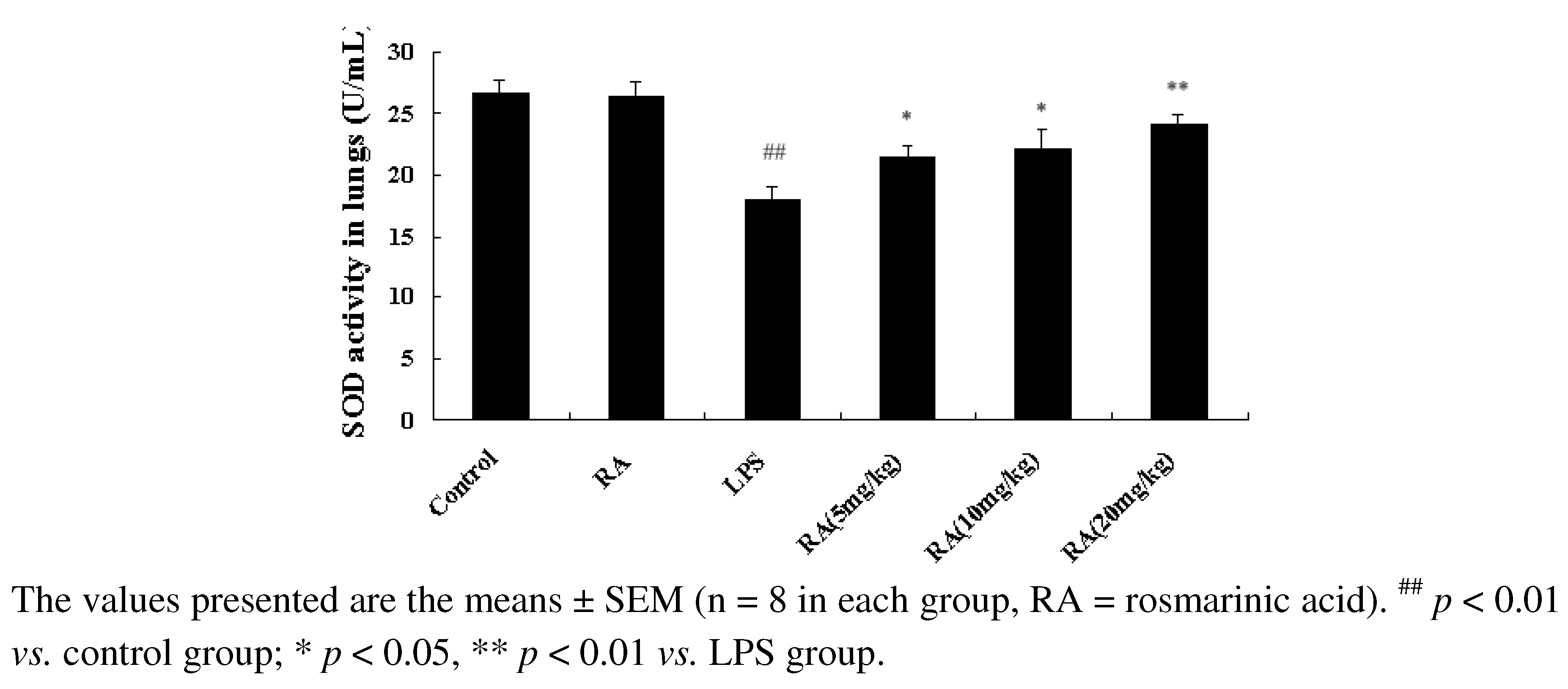
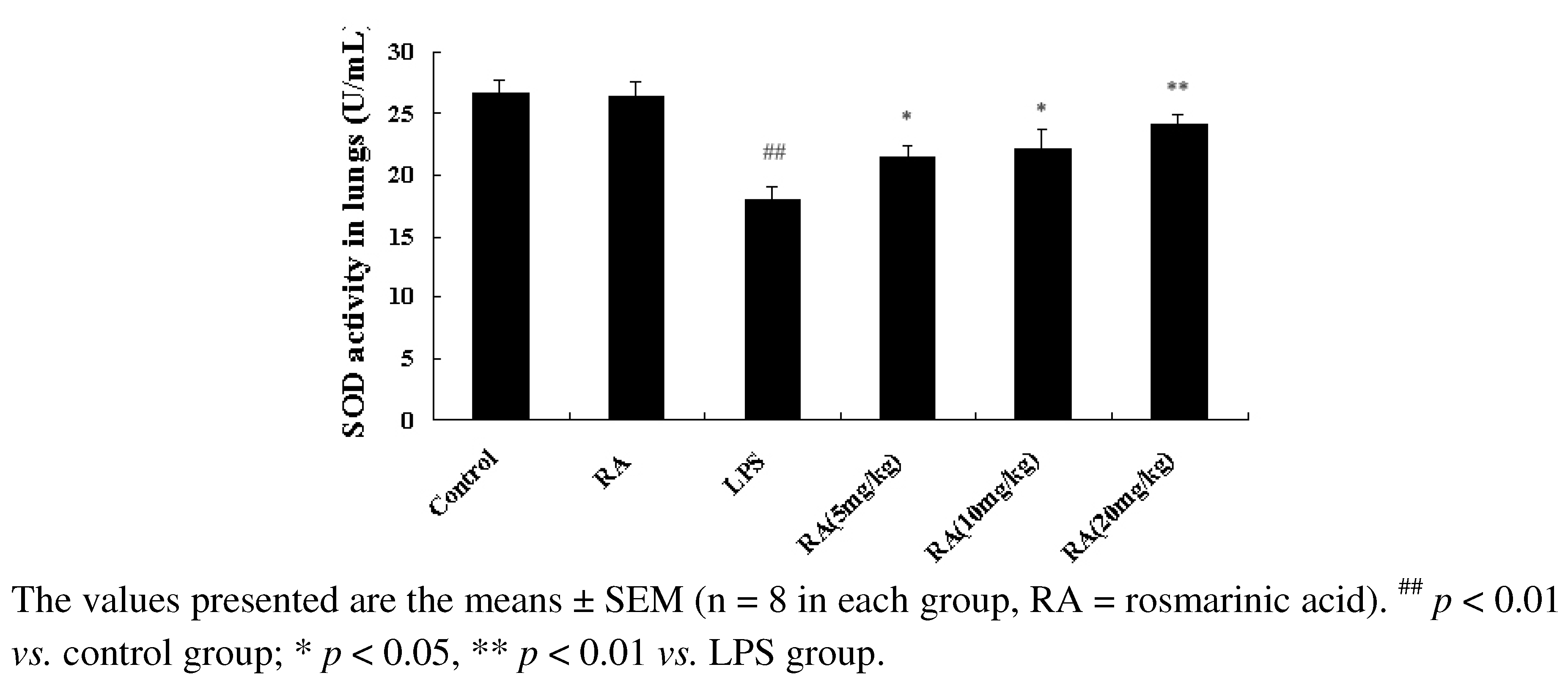
2.4. Effect of RA on SOD Activity from Mice with LPS-Induced ALI
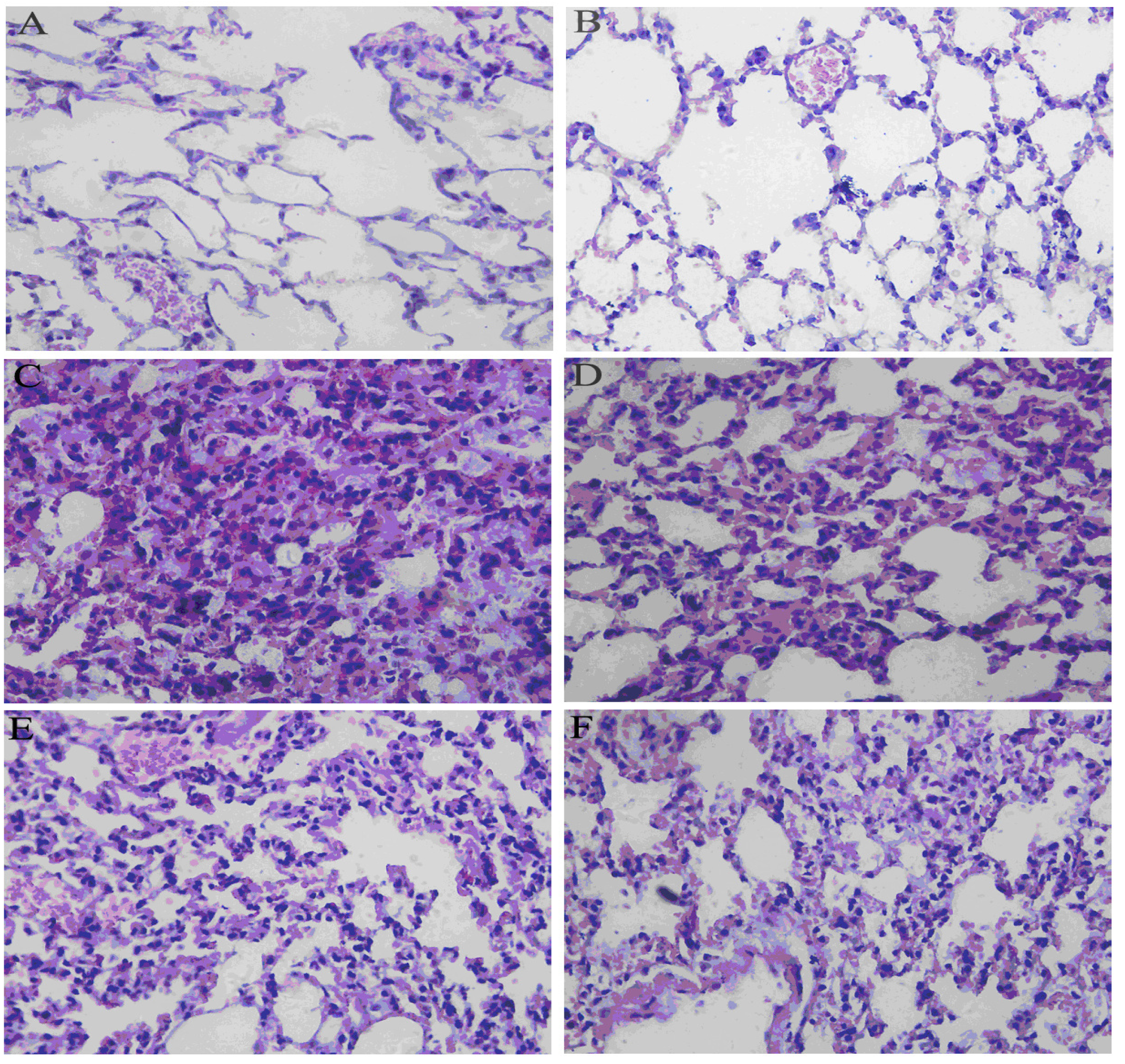
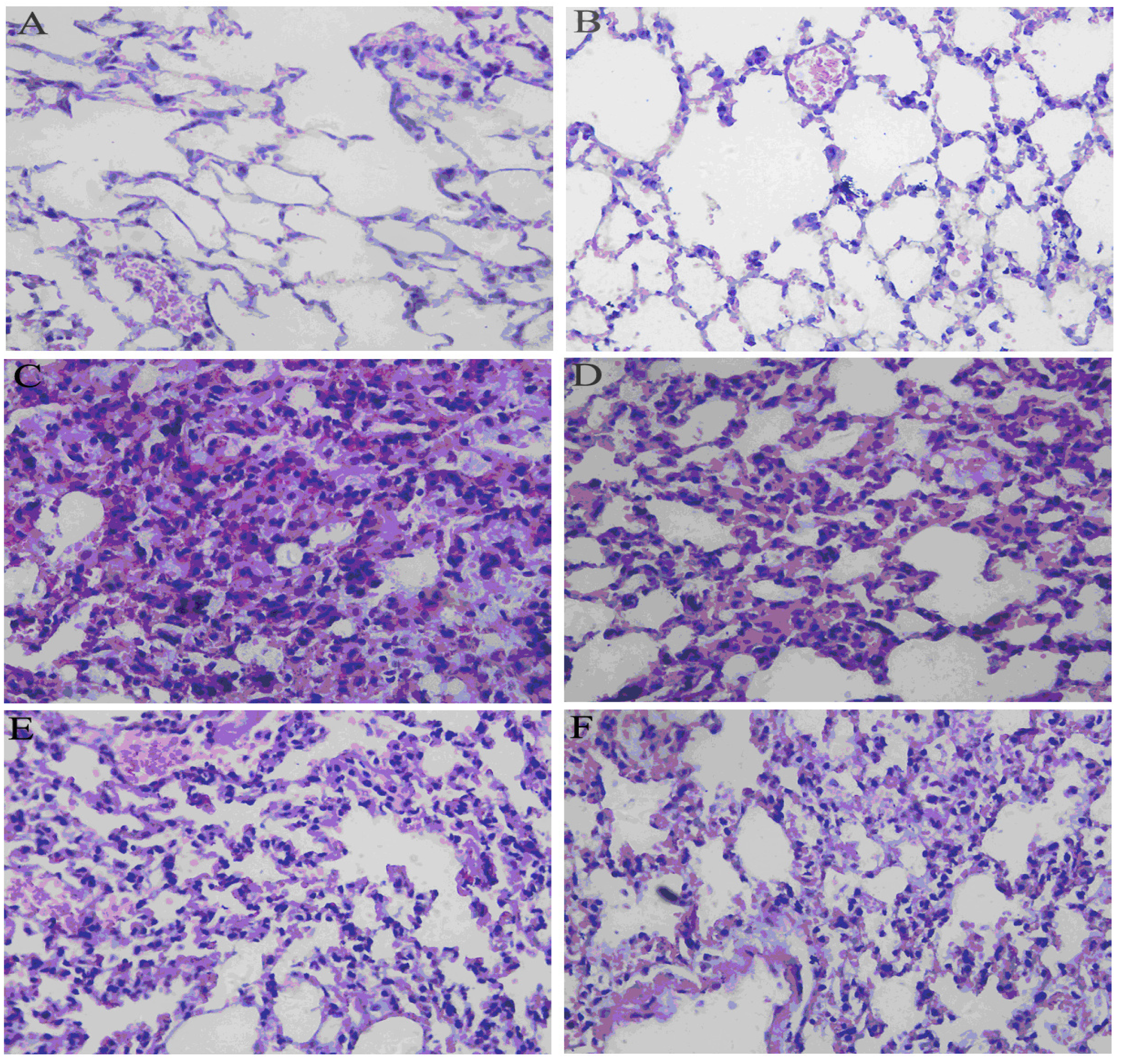
2.5. Effects of RA on Histological Changes in Lung Tissues from Mice with LPS-Induced ALI
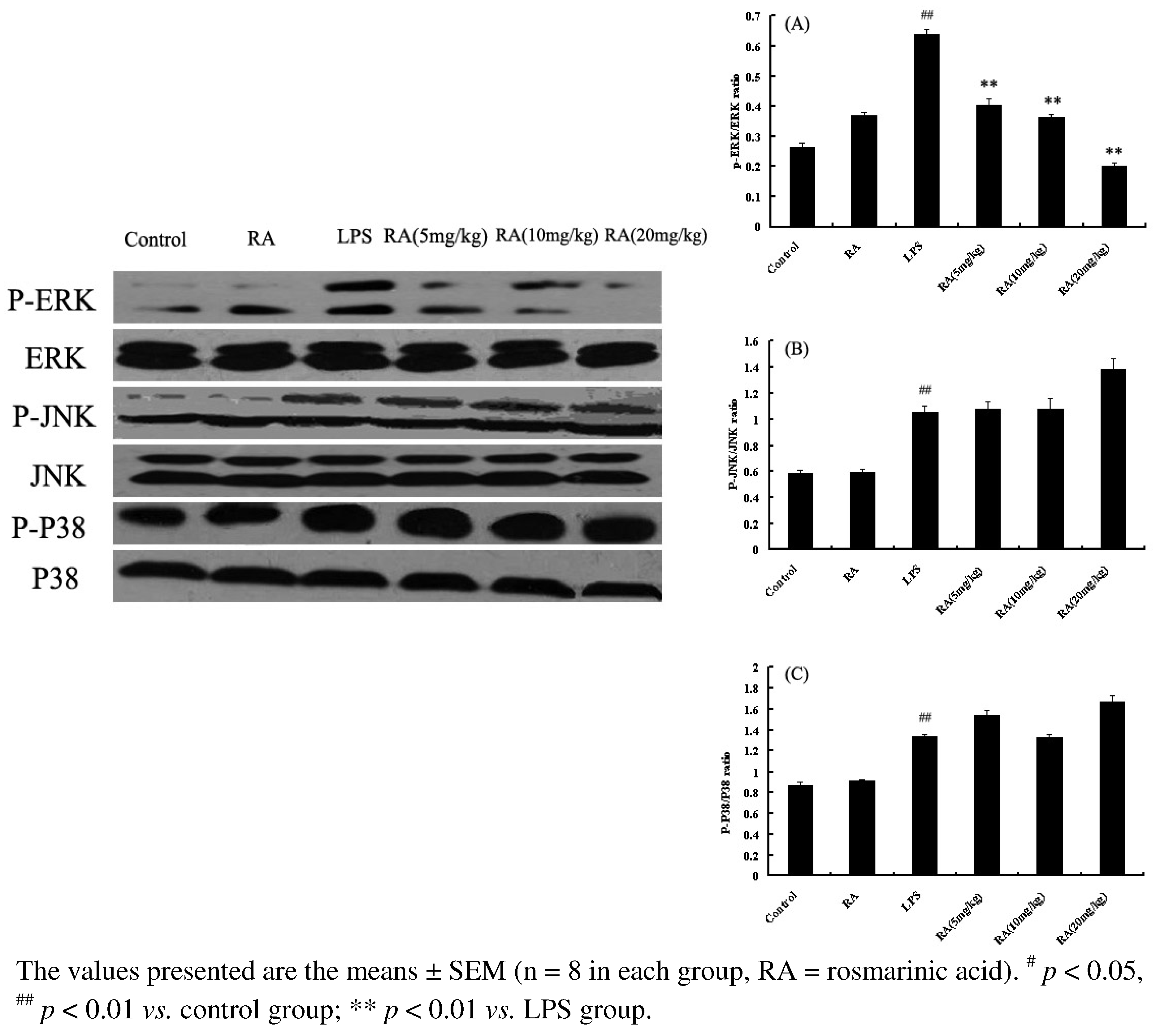
2.6. Effects of RA on MAP Kinase Phosphorylation from Mice with LPS-Induced ALI
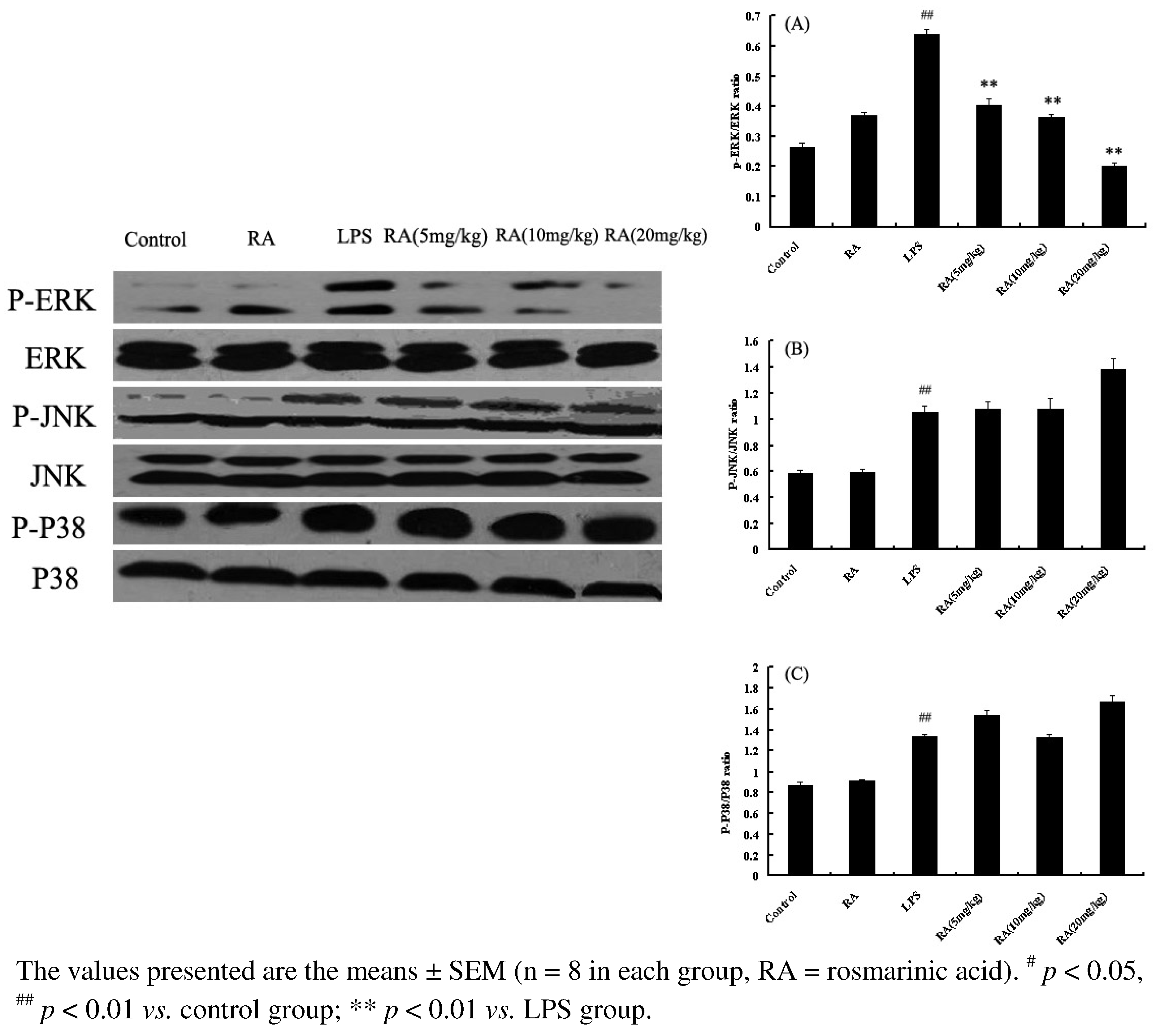
2.7. Discussion
3. Experimental
3.1. Animals
3.2. Reagents
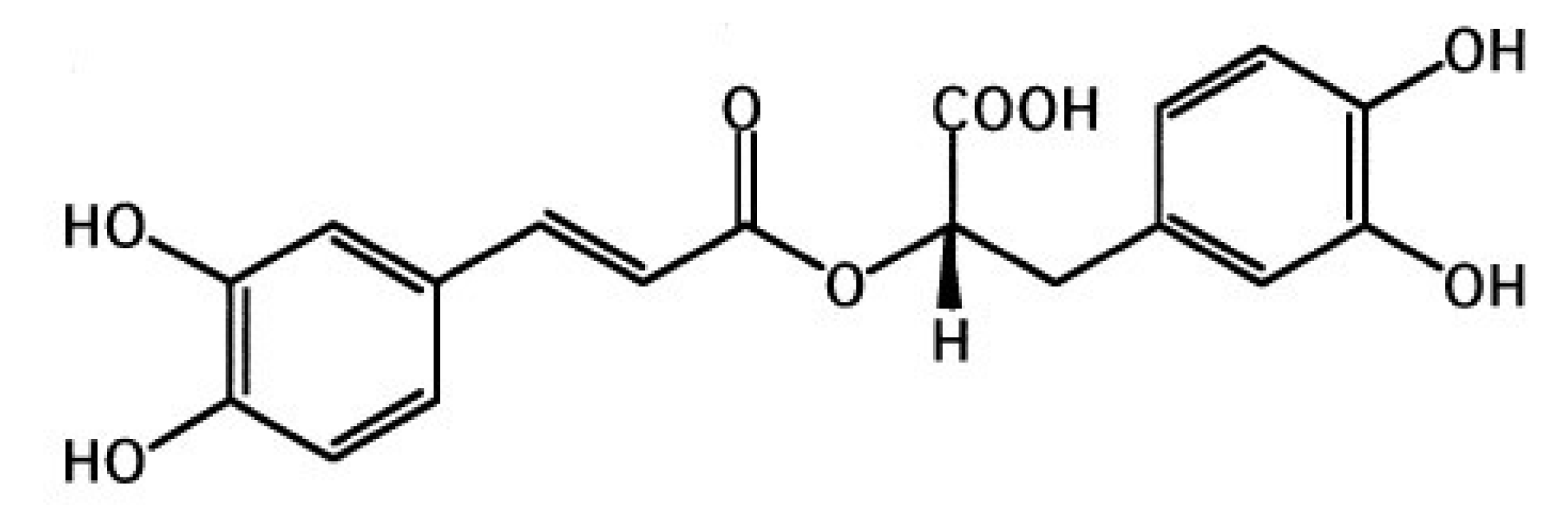
3.3. LPS-Induced ALI Model
3.4. Assays for Cytokines and SOD
3.5. Lung Wet-to-Dry Weight (W/D) Ratio
3.6. Protein Analysis
3.7. Histopathologic Evaluation
3.8. Western Blot Analysis
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
References and Notes
- Helmick, C.G.; Felson, D.T.; Lawrence, R.C.; Gabriel, S.; Hirsch, R.; Kwoh, C.K.; Liang, M.H.; Kremers, H.M.; Mayes, M.D.; Merkel, P.A.; et al. Estimates of the prevalence of arthritis and other rheumatic conditions in the United States. Part I. Arthritis Rheum. 2008, 58, 15–25. [Google Scholar]
- Garber, B.G.; Hebert, P.C.; Yelle, J.D.; Hodder, R.V.; McGowan, J. Adult respiratory distress syndrome: A systemic overview of incidence and risk factors. Crit. Care Med. 1996, 24, 687–695. [Google Scholar] [CrossRef]
- Jansson, A.H.; Eriksson, C.; Wang, X.D. Lung inflammatory responses and hyperinflation induced by an intratracheal exposure to lipopolysaccharide in rats. Lung 2004, 182, 163–171. [Google Scholar]
- Ware, L.B.; Matthay, M.A. The acute respiratory distress syndrome. N. Engl. J. Med. 2000, 342, 1334–1349. [Google Scholar] [CrossRef]
- Rahman, I. Oxidative stress, transcription factors and chromatin remodeling in lung inflammation. Biochem. Pharmacol. 2002, 64, 935–942. [Google Scholar]
- Rochelle, L.G.; Fischer, B.M.; Adler, K.B. Concurrent production of reactive oxygen and nitrogen species by airway epithelial cells in vitro. Free Radic. Biol. Med. 1998, 24, 863–868. [Google Scholar] [CrossRef]
- Psotová, J.; Kolár, M.; Sousek, J.; Svagera, Z.; Vicar, J.; Ulrichová, J. Biological activities of Prunella vulgaris extract. Phytother. Res. 2003, 17, 1082–1087. [Google Scholar] [CrossRef]
- Huang, N.; Hauck, C.; Yum, M.Y.; Rizshsky, L.; Widrlechner, M.P.; McCoy, J.A.; Murphy, P.A.; Dixon, P.M.; Nikolau, B.J.; Birt, D.F. Rosmarinic acid in Prunella vulgaris ethanol extract inhibits lipopolysaccharide-induced prostaglandin E2 and nitric oxide in RAW 264. 7 mouse macrophages. J. Agric. Food Chem. 2009, 57, 10579–10589. [Google Scholar]
- Guan, J.; Jin, D.D.; Jin, L.J.; Lu, Q. Apoptosis in organs of rats in early stage after polytrauma combined with shock. J. Trauma 2002, 52, 104–111. [Google Scholar] [CrossRef]
- Rubenfeld, G.D.; Caldwell, E.; Peabody, E.; Weaver, J.; Martin, D.P.; Neff, M.; Stern, E.J.; Hudson, L. D. Incidence and outcomes of acute lung injury. N. Engl. J. Med. 2005, 353, 1685–1693. [Google Scholar] [CrossRef]
- Brigham, K.L.; Meyrick, B. Endotoxin and lung injury. Am. Rev. Respir. Dis. 1986, 133, 913–927. [Google Scholar]
- Chu, X.; Ci, X.; He, J.; Wei, M.; Yang, X.; Cao, Q.; Li, H.; Guan, S.; Deng, Y.; Pang, D.; et al. A novel anti-inflammatory role for ginkgolide B in asthma via inhibition of the ERK/MAPK signaling pathway. Molecules 2011, 6, 7634–7648. [Google Scholar]
- Zhang, B.; Liu, Z.Y.; Li, Y.Y.; Luo, Y.; Liu, M.L.; Dong, H.Y.; Wang, Y.X.; Liu, Y.; Zhao, P.T.; Jin, F.G.; Li, Z.C. Antiinflammatory effects of matrine in LPS-induced acute lung injury in mice. Eur. J. Pharm. Sci. 2011, 44, 573–579. [Google Scholar] [CrossRef]
- Huang, N.; Hauck, C.; Yum, M.Y.; Rizshsky, L.; Widrlechner, M.P.; McCoy, J.A.; Murphy, P.A.; Dixon, P.M.; Nikolau, B.J.; Birt, D.F. Rosmarinic acid in Prunella vulgaris ethanol extract inhibits lipopolysaccharide-induced prostaglandin E2 and nitric oxide in RAW 264. 7 mouse macrophages. J. Agric. Food Chem. 2009, 25, 10579–10589. [Google Scholar]
- Bhatia, M.; Moochhala, S. Role of inflammatory mediators in the pathophysiology of acute respiratory distress syndrome. J. Pathol. 2004, 202, 145–156. [Google Scholar] [CrossRef]
- Ming, W.J.; Bersani, L.; Mantovani, A. Tumor necrosis factor is chemotactic for monocytes and polymorphonuclear leukocytes. J. Immunol. 1987, 138, 1469–1474. [Google Scholar]
- Zádor, E.; Mendler, L.; Takács, V.; de Bleecker, J.; Wuytack, F. Regenerating soleus and extensor digitorum longus muscles of the rat show elevated levels of TNF-alpha and its receptors, TNFR-60 and TNFR-80. Muscle Nerve 2001, 24, 1058–1067. [Google Scholar] [CrossRef]
- Pober, J.S.; Cotran, R.S. Cytokines and endothelial cell biology. Physiol. Rev. 1990, 70, 427–451. [Google Scholar]
- Mantovani, A.; Bussolino, F.; Dejana, E. Cytokine regulation of endothelial cell function. FASEB J. 1992, 6, 2591–2599. [Google Scholar]
- Meduri, G.U.; Kohler, G.; Headley, S.; Tolley, E.; Stentz, F.; Postlethwaite, A. Inflammatory cytokines in the BAL of patients with ARDS. Persistent elevation over time predicts poor outcome. Chest 1995, 108, 1303–1314. [Google Scholar] [CrossRef]
- Jones, S.A.; Richards, P.J.; Scheller, J.; Rose-John, S. IL-6 transsignaling: The in vivo consequences. J. Interfron Cytokine Res. 2005, 25, 241–253. [Google Scholar] [CrossRef]
- Nonas, S.A.; Miller, I.; Kawkitinarong, K.; Chatchavalvanich, S.; Gorshkova, I.; Bochkovm, V.N.; Leitinger, N.; Natarajan, V.; Garcia, J.G.; Birukov, K.G. Oxidized phospholipids reduce vascular leak and inflammation in rat model of acute lung injury. Am. J. Resp. Crit. Care. 2006, 173, 1130–1138. [Google Scholar] [CrossRef]
- Yin, H.; Jin, X.B.; Gong, Q.; Yang, H.; Hu, L.Y.; Gong, F.L.; Zhu, J.Y. Fructose-1,6-diphosphate attenuates acute lung injury induced by lipopolysaccharide in mice. Int. Immunopharmacol. 2008, 8, 1842–1847. [Google Scholar] [CrossRef]
- Bhatia, M.; Moochhala, S. Role of inflammatory mediators in the pathophysiology of acute respiratory distress syndrome. J. Pathol. 2004, 202, 145–156. [Google Scholar] [CrossRef]
- Ferrari-Baliviera, E.; Mealy, K.; Smith, R.J.; Wilmore, D.W. Tumor necrosis factor induces adult respiratory distress syndrome in rats. Arch. Surg. 1989, 124, 1400–1405. [Google Scholar] [CrossRef]
- Stephens, K.E.; Ishizaka, A.; Larrick, J.W.; Raffin, T.A. Tumor necrosis factor causes increased pulmonary permeability and edema. Comparison to septic acute lung injury. Am. Rev. Respir. Dis. 1988, 137, 1364–1370. [Google Scholar]
- Goldblum, S.E.; Hennig, B.; Jay, M.; Yoneda, K.; McClain, C.J. Tumor necrosis factor alpha-induced pulmonary vascular endothelial injury. Infect. Immun. 1989, 57, 1218–1226. [Google Scholar]
- Stephens, K.E.; Ishizaka, A.; Wu, Z.H.; Larrick, J.W.; Raffin, T.A. Granulocyte depletion prevents tumor necrosis factor-mediated acute lung injury in guinea pigs. Am. Rev. Respir. Dis. 1988, 138, 1300–1307. [Google Scholar]
- Reutershan, J.; Basit, A.; Galkina, E.V.; Ley, K. Sequential recruitment of neutrophils into lung and bronchoalveolar lavage fluid in LPS-induced acute lung injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2005, 289, L807–L815. [Google Scholar] [CrossRef]
- Kinnula, V.L.; Crapo, J.D. Superoxide dismutases in the lung and human lung diseases. Am. J. Respir. Crit. Care Med. 2003, 167, 1600–1619. [Google Scholar] [CrossRef]
- Bondeson, J. The mechanisms of action of disease-modifying antirheumatic drugs: A review with emphasis on macrophage signal transduction and the induction of proinflammatory cytokines. Gen. Pharmacol. 1997, 29, 127–150. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, H.S.; Chong, Y.H.; Kang, J.L. p38 Mitogen-activated protein kinase up-regulates LPS-induced NF-kappaB activation in the development of lung injury and RAW 264. 7 macrophages. Toxicology 2006, 225, 36–47. [Google Scholar] [CrossRef]
- Schuh, K.; Pahl, A. Inhibition of the MAP kinase ERK protects from lipopolysaccharide-induced lung injury. Biochem. Pharmacol. 2009, 77, 1827–1834. [Google Scholar]
- Szarka, R.J.; Wang, N.; Gordon, L.; Nation, P.N.; Smith, R.H. A murine model of pulmonary damage induced by lipopolysaccharide via intranasal instillation. J. Immunol. Methods 1997, 202, 49–57. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Biotechnology 1992, 24, 145–149. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chu, X.; Ci, X.; He, J.; Jiang, L.; Wei, M.; Cao, Q.; Guan, M.; Xie, X.; Deng, X.; He, J. Effects of a Natural Prolyl Oligopeptidase Inhibitor, Rosmarinic Acid, on Lipopolysaccharide-Induced Acute Lung Injury in Mice. Molecules 2012, 17, 3586-3598. https://doi.org/10.3390/molecules17033586
Chu X, Ci X, He J, Jiang L, Wei M, Cao Q, Guan M, Xie X, Deng X, He J. Effects of a Natural Prolyl Oligopeptidase Inhibitor, Rosmarinic Acid, on Lipopolysaccharide-Induced Acute Lung Injury in Mice. Molecules. 2012; 17(3):3586-3598. https://doi.org/10.3390/molecules17033586
Chicago/Turabian StyleChu, Xiao, Xinxin Ci, Jiakang He, Lanxiang Jiang, Miaomiao Wei, Qingjun Cao, Mingfeng Guan, Xianxing Xie, Xuming Deng, and Jiakang He. 2012. "Effects of a Natural Prolyl Oligopeptidase Inhibitor, Rosmarinic Acid, on Lipopolysaccharide-Induced Acute Lung Injury in Mice" Molecules 17, no. 3: 3586-3598. https://doi.org/10.3390/molecules17033586
APA StyleChu, X., Ci, X., He, J., Jiang, L., Wei, M., Cao, Q., Guan, M., Xie, X., Deng, X., & He, J. (2012). Effects of a Natural Prolyl Oligopeptidase Inhibitor, Rosmarinic Acid, on Lipopolysaccharide-Induced Acute Lung Injury in Mice. Molecules, 17(3), 3586-3598. https://doi.org/10.3390/molecules17033586