Prebiotic Activity of Polysaccharides Extracted from Gigantochloa Levis (Buluh beting) Shoots

Abstract
:1. Introduction
2. Results and Discussion
2.1. Extraction of Crude Polysaccharides
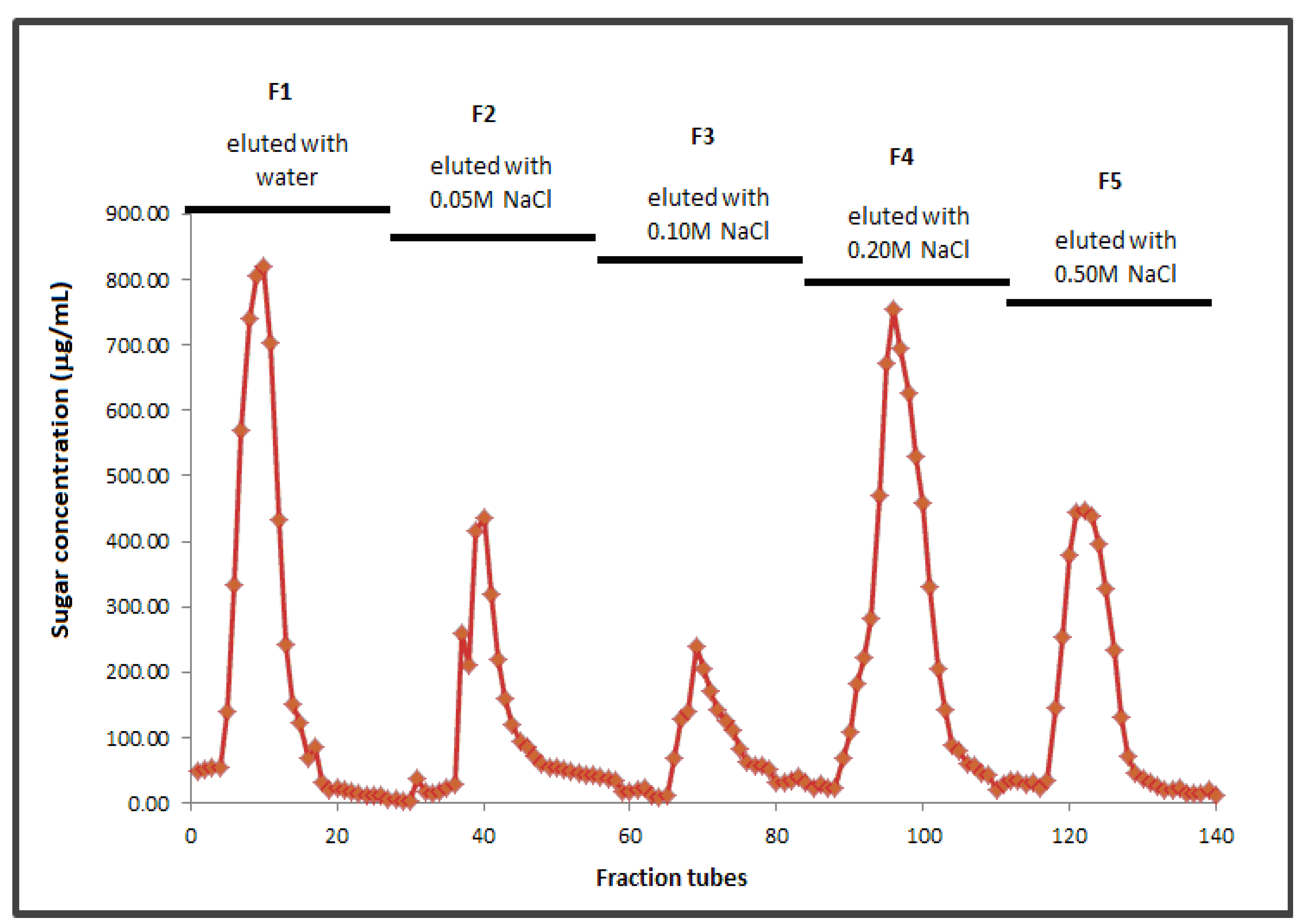
2.2. Molecular Weight and Fractionation
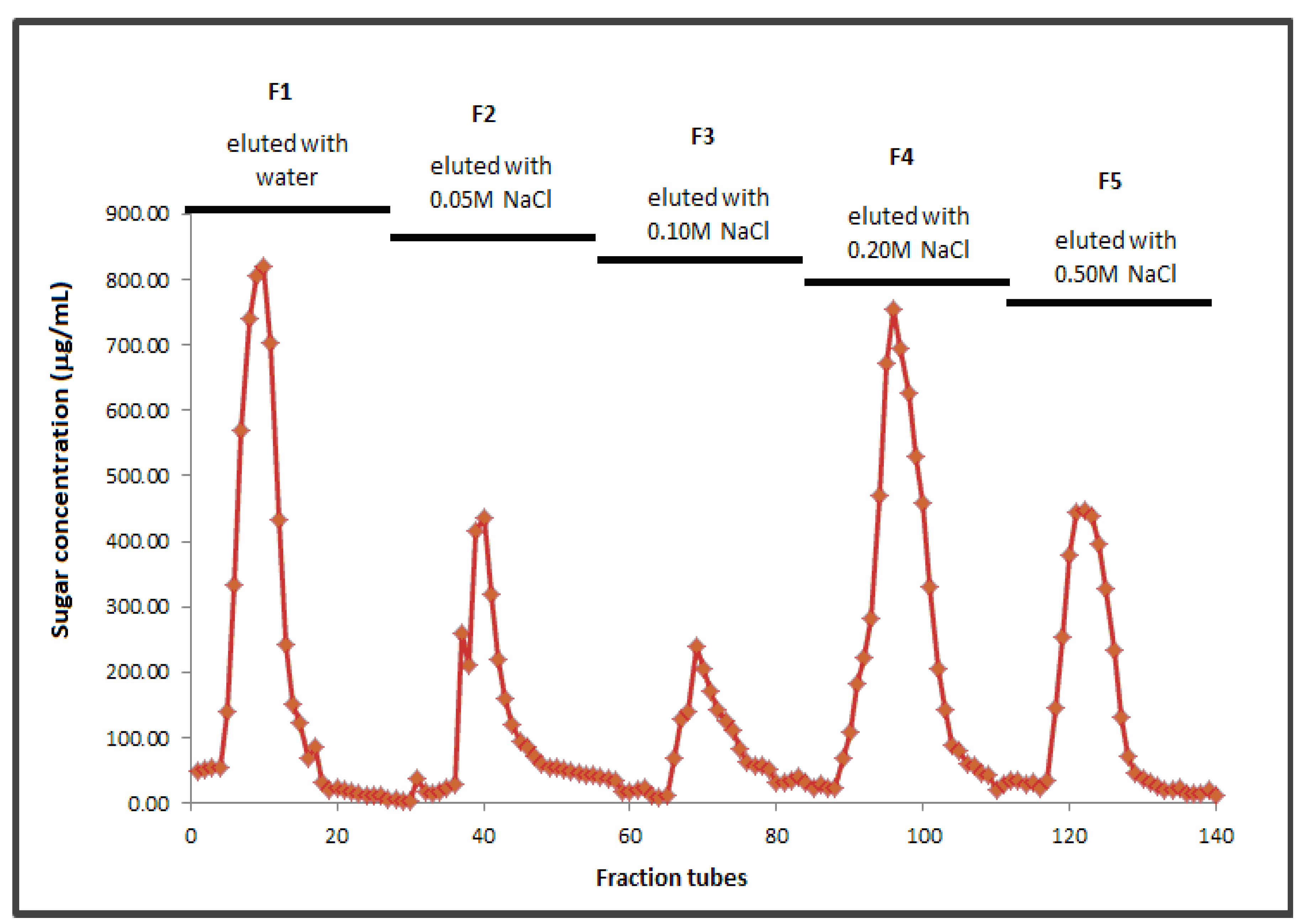
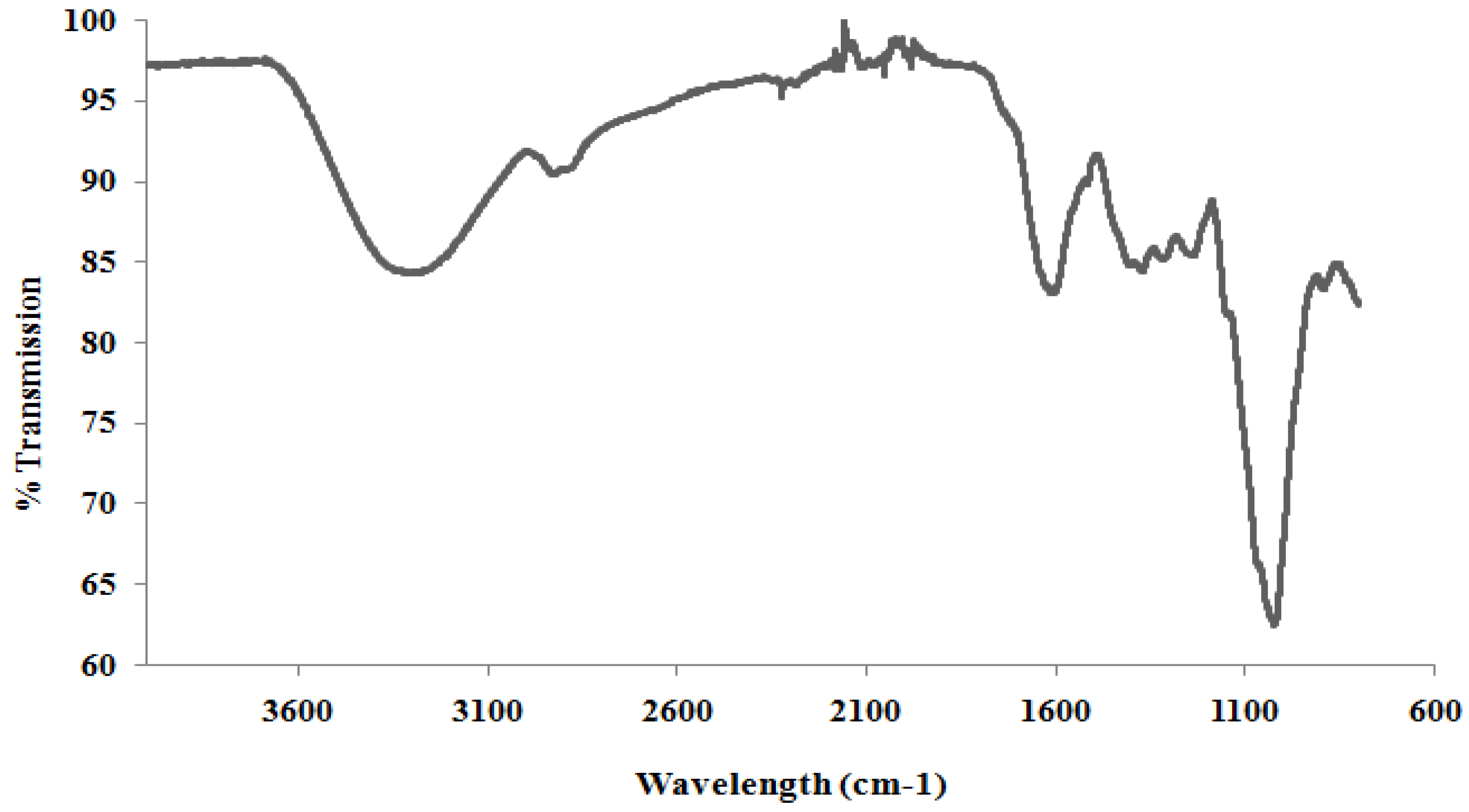
2.3. Structure of BSCP
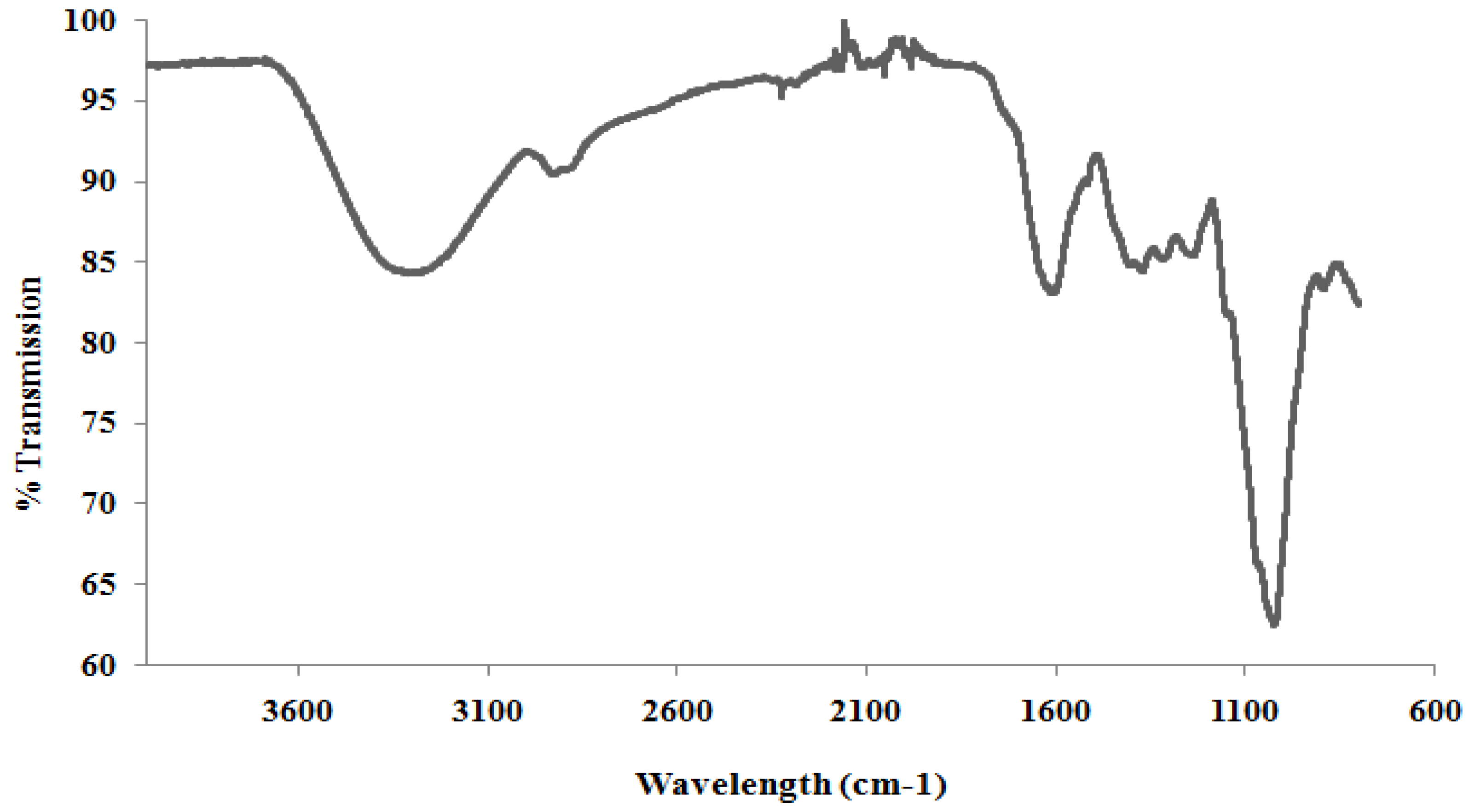
Fourier Transform Infra-Red (FTIR)
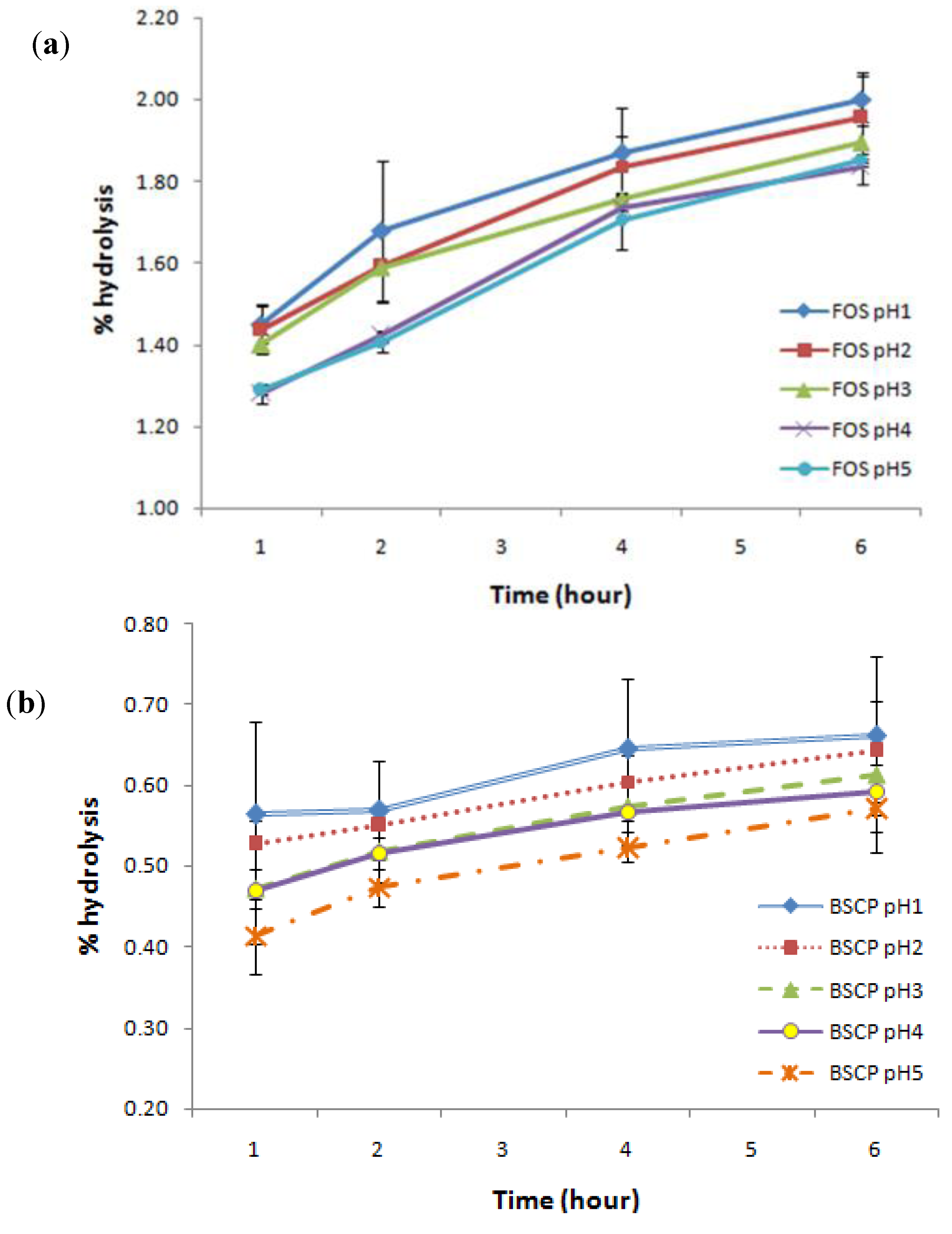
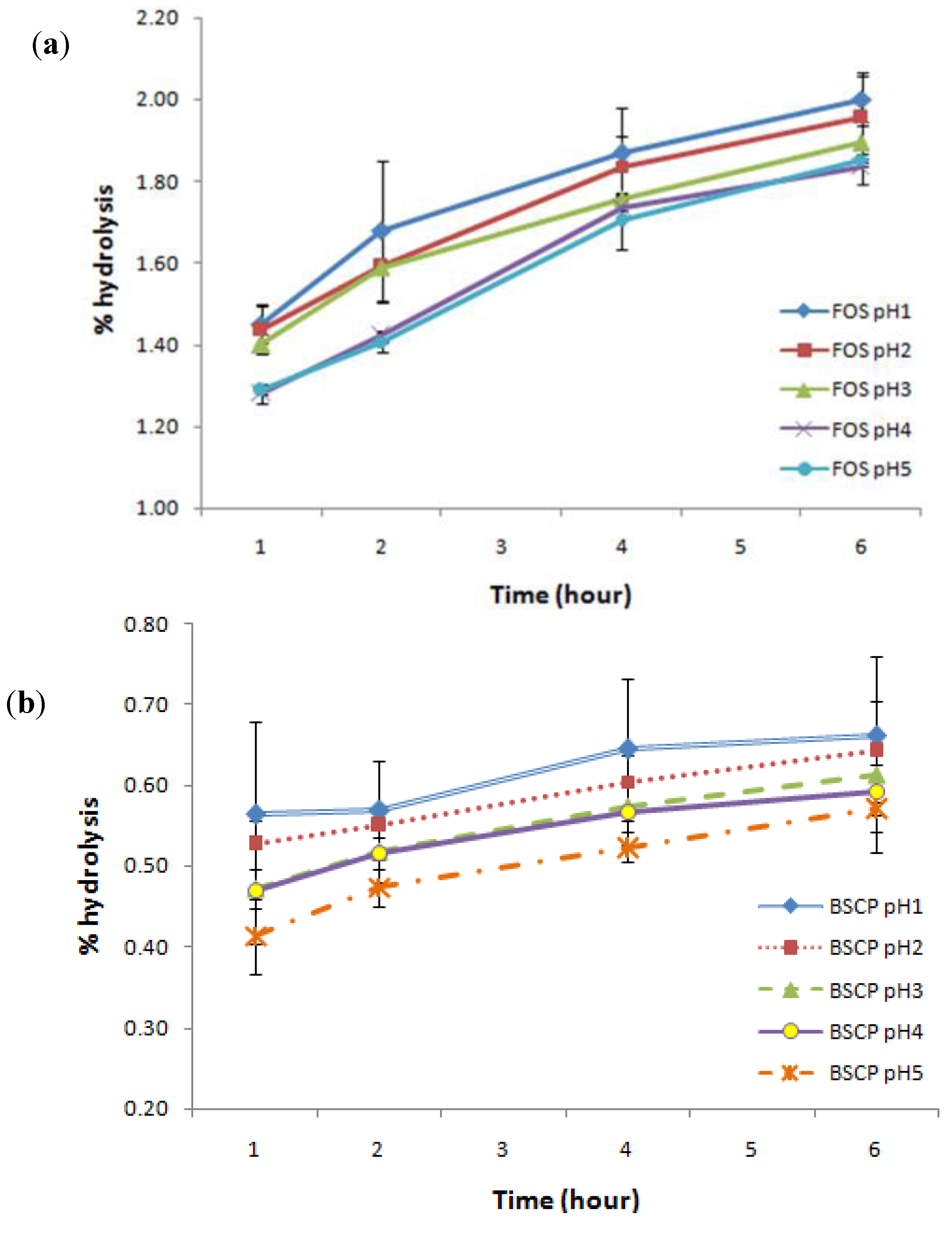
2.4. Digestibility of BSCP
2.5. Growth of Bacteria

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial strains | Carbon source | Concentration (ppm) | Fermentation time (h) | |||
|---|---|---|---|---|---|---|
| 0 | 24 | 48 | 72 | |||
| B. animalis | Control | - | 0.1163 ± 0.01 a | 0.8548 ± 0.03 e | 0.7772 ± 0.06 c | 0.5226 ± 0.12 c |
| FOS | 250 | 0.1186 ± 0.01 a | 1.3933 ± 0.05 d | 1.4844 ± 0.19 b | 1.3919 ± 0.11 a | |
| FOS | 500 | 0.1107 ± 0.02 a | 1.7201 ± 0.26 bcd | 1.6216 ± 0.08 b | 1.5814 ± 0.08 a | |
| FOS | 750 | 0.1055 ± 0.02 a | 1.6335 ± 0.14 cd | 1.5093 ± 0.03 b | 1.0356 ± 0.18 b | |
| FOS | 1000 | 0.1127 ± 0.02 a | 1.6553 ± 0.12 bcd | 1.4308 ± 0.02 b | 1.4209 ± 0.17 a | |
| BSCP | 250 | 0.1209 ± 0.03 a | 1.8616 ± 0.08 abc | 1.9139 ± 0.19 a | 1.6740 ± 0.22a | |
| BSCP | 500 | 0.1196 ± 0.03 a | 1.8667 ± 0.28 abc | 1.9268 ± 0.06 a | 1.6500 ± 0.11 a | |
| BSCP | 750 | 0.1205 ± 0.00 a | 2.0061 ± 0.27 ab | 1.9997 ± 0.25 a | 1.5276 ± 0.29 a | |
| BSCP | 1000 | 0.1227 ± 0.01 a | 2.0782 ± 0.22 a | 2.0477 ± 0.19 a | 1.7059 ± 0.10 a | |
| B. longum | Control | - | 0.1085 ± 0.01 a | 0.6053 ± 0.10 c | 0.6367 ± 0.10 b | 0.4488 ± 0.11 b |
| FOS | 250 | 0.1003 ± 0.01 a | 1.2930 ± 0.10 b | 1.4268 ± 0.05 a | 1.4867 ± 0.02 a | |
| FOS | 500 | 0.1020 ± 0.01 a | 1.2845 ± 0.06 b | 1.4655 ± 0.17 a | 1.4824 ± 0.10 a | |
| FOS | 750 | 0.1149 ± 0.02 a | 1.3171 ± 0.04 b | 1.4740 ± 0.12 a | 1.5403 ± 0.06 a | |
| FOS | 1000 | 0.1124 ± 0.02 a | 1.3381 ± 0.21 b | 1.5075 ± 0.14 a | 1.4695 ± 0.24 a | |
| BSCP | 250 | 0.1242 ± 0.01 a | 1.4576 ± 0.10 ab | 1.6077 ± 0.15 a | 1.5711 ± 0.07 a | |
| BSCP | 500 | 0.1249 ± 0.03 a | 1.4348 ± 0.22 ab | 1.6350 ± 0.18 a | 1.6075 ± 0.07 a | |
| BSCP | 750 | 0.1139 ± 0.02 a | 1.6637 ± 0.14 a | 1.6677 ± 0.05 a | 1.6142 ± 0.06 a | |
| BSCP | 1000 | 0.1166 ± 0.01 a | 1.7139 ± 0.24 a | 1.7139 ± 0.24 a | 1.5802 ± 0.17 a | |
| L. acidophilus | Control | - | 0.1223 ± 0.01 a | 1.0444 ± 0.11 c | 1.0379 ± 0.05 c | 0.7057 ± 0.15 b |
| FOS | 250 | 0.1187 ± 0.01 a | 1.5589 ± 0.10 b | 1.7159 ± 0.05 ab | 1.6722 ± 0.20 a | |
| FOS | 500 | 0.1267 ± 0.01 a | 1.5841 ± 0.01 b | 1.7705 ± 0.07 ab | 1.7743 ± 0.11 a | |
| FOS | 750 | 0.1153 ± 0.01 a | 1.5849 ± 0.09 b | 1.7459 ± 0.09 ab | 1.7240 ± 0.09 a | |
| FOS | 1000 | 0.1269 ± 0.01 a | 1.5965 ± 0.09 b | 1.6824 ± 0.05 b | 1.7611 ± 0.03 a | |
| BSCP | 250 | 0.1252 ± 0.01 a | 1.8167 ± 0.18 c | 1.9436 ± 0.23 ab | 1.8569 ± 0.07 a | |
| BSCP | 500 | 0.1294 ± 0.01 a | 1.8540 ± 0.13 c | 1.9501 ± 0.24 ab | 1.8629 ± 0.06 a | |
| BSCP | 750 | 0.1235 ± 0.01 a | 1.8733 ± 0.11 c | 1.9822 ± 0.16 a | 1.8650 ± 0.10 a | |
| BSCP | 1000 | 0.1293 ± 0.02 a | 1.8912 ± 0.19 c | 2.0053 ± 0.21 a | 1.8393 ± 0.13 a | |
| Salmonella | Control | - | 0.1162 ± 0.03 a | 0.9632 ± 0.05 c | 0.9789 ± 0.13 b | 0.8351 ± 0.11 b |
| FOS | 250 | 0.1172 ± 0.01 a | 1.3456 ± 0.06 ab | 1.4051 ± 0.22 a | 1.3943 ± 0.13 a | |
| FOS | 500 | 0.1147 ± 0.01 a | 1.1879 ± 0.10 b | 1.3427 ± 0.20 a | 1.4029 ± 0.15 a | |
| FOS | 750 | 0.1244 ± 0.01 a | 1.4754 ± 0.10 a | 1.4883 ± 0.18 a | 1.5029 ± 0.17 a | |
| FOS | 1000 | 0.1229 ± 0.01 a | 1.3560 ± 0.13 ab | 1.5154 ± 0.14 a | 1.5431 ± 0.15 a | |
| BSCP | 250 | 0.1201 ± 0.01 a | 1.4394 ± 0.15 a | 1.5205 ± 0.17 a | 1.3993 ± 0.19 a | |
| BSCP | 500 | 0.1187 ± 0.01 a | 1.4571 ± 0.09 a | 1.4273 ± 0.17 a | 1.3831 ± 0.18 a | |
| BSCP | 750 | 0.1229 ± 0.02 a | 1.4805 ± 0.14 a | 1.5354 ± 0.14 a | 1.5177 ± 0.14 a | |
| BSCP | 1000 | 0.1167 ± 0.02 a | 1.4372 ± 0.16 a | 1.5872 ± 0.21 a | 1.5849 ± 0.20 a | |
| Bacterial strains | Carbon Source | 0–24 h | 24–48 h | 48–72 h |
|---|---|---|---|---|
| B. animalis | Control | 6.35 | −0.09 | −0.33 |
| FOS 250 ppm | 10.75 | 0.07 | −0.06 | |
| FOS 500 ppm | 14.54 | −0.06 | −0.02 | |
| FOS 750 ppm | 14.48 | −0.08 | −0.31 | |
| FOS 1000 ppm | 13.69 | −0.14 | −0.01 | |
| BSCP 250 ppm | 14.40 | 0.03 | −0.13 | |
| BSCP 500 ppm | 14.61 | 0.03 | −0.14 | |
| BSCP 750 ppm | 15.65 | 0.00 | −0.24 | |
| BSCP 1000 ppm | 15.94 | −0.01 | −0.17 | |
| B. longum | Control | 4.58 | 0.05 | −0.30 |
| FOS 250 ppm | 11.89 | 0.10 | 0.04 | |
| FOS 500 ppm | 11.59 | 0.14 | 0.01 | |
| FOS 750 ppm | 10.46 | 0.12 | 0.04 | |
| FOS 1000 ppm | 10.90 | 0.13 | −0.03 | |
| BSCP 250 ppm | 10.74 | 0.10 | −0.02 | |
| BSCP 500 ppm | 10.49 | 0.14 | −0.02 | |
| BSCP 750 ppm | 13.61 | 0.00 | −0.03 | |
| BSCP 1000 ppm | 13.70 | 0.00 | −0.08 | |
| L. acidophilus | Control | 7.54 | −0.01 | −0.32 |
| FOS 250 ppm | 12.13 | 0.10 | −0.03 | |
| FOS 500 ppm | 11.50 | 0.12 | 0.00 | |
| FOS 750 ppm | 12.75 | 0.10 | −0.01 | |
| FOS 1000 ppm | 11.58 | 0.05 | 0.05 | |
| BSCP 250 ppm | 13.51 | 0.07 | −0.04 | |
| BSCP 500 ppm | 13.33 | 0.05 | −0.04 | |
| BSCP 750 ppm | 14.17 | 0.06 | −0.06 | |
| BSCP 1000 ppm | 13.63 | 0.06 | −0.08 | |
| Salmonella | Control | 7.29 | 0.02 | −0.15 |
| FOS 250 ppm | 10.48 | 0.04 | −0.01 | |
| FOS 500 ppm | 9.36 | 0.13 | 0.04 | |
| FOS 750 ppm | 10.86 | 0.01 | 0.01 | |
| FOS 1000 ppm | 10.03 | 0.12 | 0.02 | |
| BSCP 250 ppm | 10.99 | 0.06 | −0.08 | |
| BSCP 500 ppm | 11.28 | −0.02 | −0.03 | |
| BSCP 750 ppm | 11.05 | 0.04 | −0.01 | |
| BSCP 1000 ppm | 11.32 | 0.10 | 0.00 |
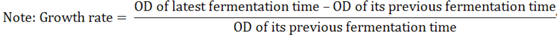
3. Experimental
3.1. Extraction of Polysaccharides
3.1.1. Bamboo Shoots
3.1.2. Chemicals
3.1.3. Extraction
3.2. Molecular Weight Estimation
3.3. Fractionation
3.4. Structural Elucidation of Polysaccharides
Fourier Transform Infra-Red (FTIR) Spectroscopy
3.5. Resistance towards Acid Digestibility
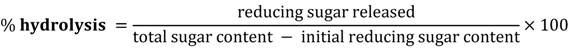
3.6. Media and Microorganisms
3.6.1. Media
3.6.2. Microorganisms
3.7. Prebiotic Activity
3.7.1. pH Measurement
3.7.2. Determination of Optical Density (OD)
3.8. Statistical Analysis
4. Conclusions
- Sample Availability: Not available.
References and Notes
- Razak, A.; Latif, A.; Liese, W.; Haron, N. Planting and Utilization of Bamboo in Peninsular Malaysia; Forest Research Institute of Malaysia: Kuala Lumpur, Malaysia, 1995. [Google Scholar]
- Panee, J. Bamboo Extract in the Prevention of Diabetes and Breast Cancer. In Complementary and Alternative Therapies in the Aging Population; Watson, R.R., Ed.; Elsevier: UK, USA, 2009; pp. 159–177. [Google Scholar]
- Roberfroid, M.B. Chicory fructooligosaccharides and the gastrointestinal tract. Nutrition 2000, 16, 677–679. [Google Scholar] [CrossRef]
- Siro, I.; Kapolna, E.; Kapolna, B.; Lugasi, A. Functional food. Product development, marketing and consumer acceptance—A review. Appetite 2008, 51, 456–467. [Google Scholar]
- Gibson, G.R.; Probert, H.M.; Rastall, R.A.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: updating the concept of prebiotics. Nutr. Res. Rev. 2004, 17, 259–275. [Google Scholar] [CrossRef]
- Carabin, I.G.; Flamm, W.G. Evaluation of safety on inulin and oligofructose as dietary fiber. Regul. Toxicol. Pharmacol. 1999, 30, 268–282. [Google Scholar] [CrossRef]
- Fuller, R. Probiotics in man and animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar] [CrossRef]
- Suvarna, V.C.; Boby, V.U. Probiotics in human health: A current assessment. Curr. Sci. 2005, 88, 1744–1745. [Google Scholar]
- Saarela, M.; Mogensen, G.; Fonden, R.; Matto, J.; Mattila-Sandholm, T. Probiotic bacteria: Safety, functional and technological properties. J. Biotechnol. 2000, 84, 197–215. [Google Scholar]
- Lee, H.W.; Park, Y.S.; Jong, J.S.; Shim, W.S. Chitosan oligosaccharides, dp 2-8, have prebiotic effect on the Bifidobacterium bifidum and Lactobacillus sp. Anaerobe 2002, 8, 319–324. [Google Scholar] [CrossRef]
- Vergara, C.M.A.C.; Honorato, T.L.; Maia, G.A.; Rodrigues, S. Prebiotic effect of fermented cashew apple (Anacardium occidentale L) juice. LWT-Food Sci. Technol. 2010, 43, 141–145. [Google Scholar]
- Rycroft, C.E.; Jones, M.R.; Gibson, G.R.; Rastall, R.A. A comparative in vitro evaluation of the fermentation properties of prebiotic oligosaccharides. J. Appl. Microbiol. 2001, 91, 878–887. [Google Scholar] [CrossRef]
- Roberfroid, M. Functional food concept and its application to prebiotics. Digest. Liver Dis. 2002, 34 (Suppl. 2), 105–110. [Google Scholar]
- Douglas, L.C.; Sanders, M.E. Probiotics and prebiotics in Dietetics Practice. J. Am. Diet. Assoc. 2008, 108, 510–521. [Google Scholar] [CrossRef]
- Synytsya, A.; Mickova, K.; Synytsya, A.; Jablonsky, I.; Spevacek, J.; Erban, V.; Kovarikova, E.; Copikova, J. Glucans from fruit bodies of cultivated mushrooms Pleurotus ostreatus and Pleurotus eryngii: Structure and potential prebiotic activity. Carbohydr. Polym. 2009, 76, 548–556. [Google Scholar] [CrossRef]
- Vidanarachchi, J.K.; Iji, P.A.; Mikkelsen, L.L.; Sims, I.; Choct, M. Isolation and characterization of water-soluble prebiotic compounds from Australian and New Zealand plants. Carbohydr. Polym. 2009, 77, 670–676. [Google Scholar] [CrossRef]
- Wichienchot, S.; Jatupornpipat, M.; Rastall, R.A. Oligosaccharides of pitaya (dragon fruit) flesh and their prebiotic properties. Food Chem. 2009, 120, 850–857. [Google Scholar]
- Vergara, C.M.A.C.; Honorato, T.L.; Maia, G.A.; Rodrigues, S. Prebiotic effect of fermented cashew apple (Anacardium occidentale L) juice. LWT-Food Sci. Technol. 2010, 43, 141–145. [Google Scholar]
- Madhukumar, M.S.; Muralikrishna, G. Structural characterisation and determination of prebiotic activity of purified xylo-oligosaccharides obtained from Bengal gram husk (Cicer arietinum L.) and wheat bran (Triticum aestivum). Food Chem. 2009, 118, 215–223. [Google Scholar]
- Fuchigami, M. Differences between bamboo shoots and vegetables in thermal disintegration of tissues and polysaccharides fractionated by successive extraction. J. Food Sci. 1990, 55, 739–745. [Google Scholar] [CrossRef]
- Humble, C.G.; Malarcher, A.M.; Tyroler, H.A. Dietary fiber and coronary heart disease in middle-aged hypercholesterolemic men. Am. J. Prev. Med. 1993, 9, 197–202. [Google Scholar]
- Wolk, A.; Manson, J.E.; Stampfer, M.J.; Colditz, G.A.; Hu, F.B.; Speizer, F.E. Long term intake of dietary fiber and decreased risk of coronary heart disease among women. JAMA-J. Am. Med. Assoc. 1999, 281, 1998–2004. [Google Scholar]
- Park, E.J.; Jhon, D.Y. Effects of bamboo shoot consumption on lipid profiles and function in healthy young women. Nutrition 2009, 25, 723–728. [Google Scholar] [CrossRef]
- Izydorczyk, M. Understanding the Chemistry of Food Carbohydrates. In Food Carbohydrates: Chemistry, Physical Properties and Applications; Cui, S.W., Ed.; Taylor & Francis: London, UK, 2005. [Google Scholar]
- Cai, W.; Gu, X.; Tang, J. Extraction, purification and characterization of the polysaccharides from Opuntia milpa alta. Carbohydr. Polym. 2008, 71, 403–410. [Google Scholar] [CrossRef]
- Staub, A.M. Removal of proteins: Sevag method. Methods Carbohydr. Chem. 1965, 5, 5–6. [Google Scholar]
- Hokputsa, S.; harding, S.E.; Inngjerdingen, K.; Jumel, K.; Michaelsen, T.E.; Heinze, T.; Koschellad, A.; Paulsen, B.S. Bioactive polysaccharides from the stems of the thai medicinal plant Acanthus ebracteatus: Their chemical and physical features. Carbohydr. Res. 2004, 339, 753–762. [Google Scholar] [CrossRef]
- Yang, C.; He, N.; Ling, X.; Ye, M.; Zhang, C.; Shao, W.; Zao, C.; Wang, Z.; Li, Q. The isolation and characterization of polysaccharides from longan pulp. Sep. Purif. Technol. 2008, 63, 226–230. [Google Scholar] [CrossRef]
- Carnel, V. Recent extraction techniques for solid matrices supercritical fluid extraction, pressurized fluid extraction and microwave-assisted extraction: their potential and pitfalls. Analyst 2001, 126, 1182–1193. [Google Scholar]
- Gullon, P.; Gonzalez-Munoz, M.J.; Parajo, J.C. Manufacture and prebiotic potential of oligosaccharides derived from industrial solid wastes. Bioresour. Technol. 2011, 102, 6112–6119. [Google Scholar] [CrossRef]
- Xu, X.; Chen, P.; Wang, Y.; Zhang, L. Chain conformation and rheological behavior of an extracellular heteropolysaccharide Erwinia gum in aqueous solution. Carbohydr. Res. 2009, 344, 113–119. [Google Scholar] [CrossRef]
- Singthong, J.; Ningsanond, S.; Cui, S.W. Extraction and physicochemical characterisation of polysaccharide gum from Yanang (Tiliacora triandra) leaves. Food Chem. 2009, 114, 1301–1307. [Google Scholar] [CrossRef]
- Ge, Q.; Zhang, A.Q.; Sun, P.L. Structural investigation of a novel water-soluble heteropolysaccharide from the fruiting bodies of Phellinus baumii Pilat. Food Chem. 2009, 114, 391–395. [Google Scholar] [CrossRef]
- Huang, Z.; Zhang, L. Chemical structuresa of water-soluble polysaccharides from Rhizoma Panacis Japonici. Carbohydr. Res. 2009, 344, 1136–1140. [Google Scholar] [CrossRef]
- Kacurakova, M.; Wilson, R.H. Developments in mid-infrared FTIR spectroscopy of selected carbohydrates. Carbohydr. Polym. 2001, 44, 291–303. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, L. Structure and chain conformation of five water-soluble derivatives of a β-D-glucan isolated from Ganoderma lucidum. Carbohydr. Res. 2009, 344, 105–112. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, L.M. Chemical structural and chain conformation characterization of some bioactive polysaccharides isolated from natural sources. Carbohydr. Polym. 2009, 76, 349–361. [Google Scholar] [CrossRef]
- Wang, Y. Prebiotics: Present and future in food science and technology. Food Res. Int. 2009, 42, 8–12. [Google Scholar] [CrossRef]
- Musatto, S.T.; Mancilha, I.M. Non-digestible oligosaccharides: A review. Carbohydr. Polym. 2007, 68, 587–597. [Google Scholar] [CrossRef]
- Nakada, T.; Nishimoto, T.; Chaen, H.; Fukuda, S. Kojioligosaccharides: Application of kojibiose, phosphorylase on the formation of various kojioligosaccharides. In Oligosaccharides in Food and Agriculture; Eggleston, G., Cote, G.L., Eds.; American Chemical Society: Washington, DC, USA, 2003; Volume 849, pp. 104–117. [Google Scholar]
- Wichienchot, S.; Prasertsan, S.; Hongpattarakere, P.; Gibson, G.R.; Rastall, R.A. In vitro fermentation of mixed linkage gluco-oligosaccharides produced by Gluconobacter oxydans NCIMB 4943, by the human colonic microflora. Curr. Issues Intest. Microbiol. 2006, 7, 7–12. [Google Scholar]
- Aida, F.M.N.A.; Shuhaimi, M.; Yazid, M.; Maaruf, A.G. Mushroom as a potential source of prebiotics: A review. Trends Food Sci. Tech. 2009, 20, 567–575. [Google Scholar] [CrossRef]
- Beards, E.; Tuohy, K.; Gibson, G. Bacterial, SCFA and gas profiles of a range of food ingredients following in vitro fermentation by human colonic microbiota. Anaerobe 2010, 16, 420–425. [Google Scholar] [CrossRef]
- Vardakou, M.; Palop, C.N.; Christakopoulos, P.; Faulds, C.B.; Gasson, M.A.; Narbad, A. Evaluation of prebiotic properties of wheat arabinoxylan fractions and induction of hydrolase activity in gut microflora. Int. J. Food Microbiol. 2008, 123, 166–170. [Google Scholar] [CrossRef]
- Kleessen, B.; Hartman, L.; Blaut, M. Oligofructose and long-chain inulin influence the gut microbial ecology of rats associated with a human faecal flora. Br. J. Nutr. 2001, 86, 291–300. [Google Scholar] [CrossRef]
- Rada, V.; Nevoral, J.; Tomankova, E.; Smehilova, M.; Killer, J. Growth of infant faecal bifidobacteria and clostridia on prebiotic oligosaccharides in in vitro conditions. Anaerobe 2008, 14, 205–208. [Google Scholar] [CrossRef]
- Teitelbaum, J.E.; Walker, W.A. Nutritional impact of pre- and probiotics as protective gastrointestinal organisms. Annu. Rev. Nutr. 2002, 22, 107–138. [Google Scholar] [CrossRef]
- Macfarlane, S.; Macfarlane, G.T.; Cumming, J.H. Review article: Prebiotics in the gastrointestinal tract. Aliment. Pharmacol. Ther. 2006, 24, 701–714. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Chen, Y.; Xie, M.Y.; Nie, S.P.; Li, C.; Wang, Y.X. Purification, composition analysis and antioxidant activity of a polysaccharide from the fruiting bodies of Ganoderma atrum. Food Chem. 2008, 107, 231–241. [Google Scholar] [CrossRef]
- Bao, X.; Liu, C.; Fang, J.; Li, X. Structural and immunological studies of a major spores of Ganoderma lucidum (Fr.) Karst. Carbohydr. Res. 2001, 332, 67–74. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hhamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Robertson, J.A.; Ryden, P.; Botham, R.L.; Reading, L.; Gibson, G.R.; Ring, S.G. Structural properties if diet-derived polysaccharides and their influence on butyrate production during fermentation. Br. J. Nutr. 2001, 81, S219–S223. [Google Scholar]
- Palframan, R.J.; Gibson, G.R.; Rastall, R.A. Effect of pH and dose on the growth of gut bacteria on prebiotic carbohydrates in vitro. Anaerobe 2002, 8, 287–292. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Azmi, A.F.M.N.; Mustafa, S.; Hashim, D.M.; Manap, Y.A. Prebiotic Activity of Polysaccharides Extracted from Gigantochloa Levis (Buluh beting) Shoots. Molecules 2012, 17, 1635-1651. https://doi.org/10.3390/molecules17021635
Azmi AFMN, Mustafa S, Hashim DM, Manap YA. Prebiotic Activity of Polysaccharides Extracted from Gigantochloa Levis (Buluh beting) Shoots. Molecules. 2012; 17(2):1635-1651. https://doi.org/10.3390/molecules17021635
Chicago/Turabian StyleAzmi, Aida Firdaus Muhammad Nurul, Shuhaimi Mustafa, Dzulkifly Md. Hashim, and Yazid Abdul Manap. 2012. "Prebiotic Activity of Polysaccharides Extracted from Gigantochloa Levis (Buluh beting) Shoots" Molecules 17, no. 2: 1635-1651. https://doi.org/10.3390/molecules17021635
APA StyleAzmi, A. F. M. N., Mustafa, S., Hashim, D. M., & Manap, Y. A. (2012). Prebiotic Activity of Polysaccharides Extracted from Gigantochloa Levis (Buluh beting) Shoots. Molecules, 17(2), 1635-1651. https://doi.org/10.3390/molecules17021635