Plant Polyphenols and Their Anti-Cariogenic Properties: A Review

 ,
,  ,
, 
Abstract
:1. Introduction
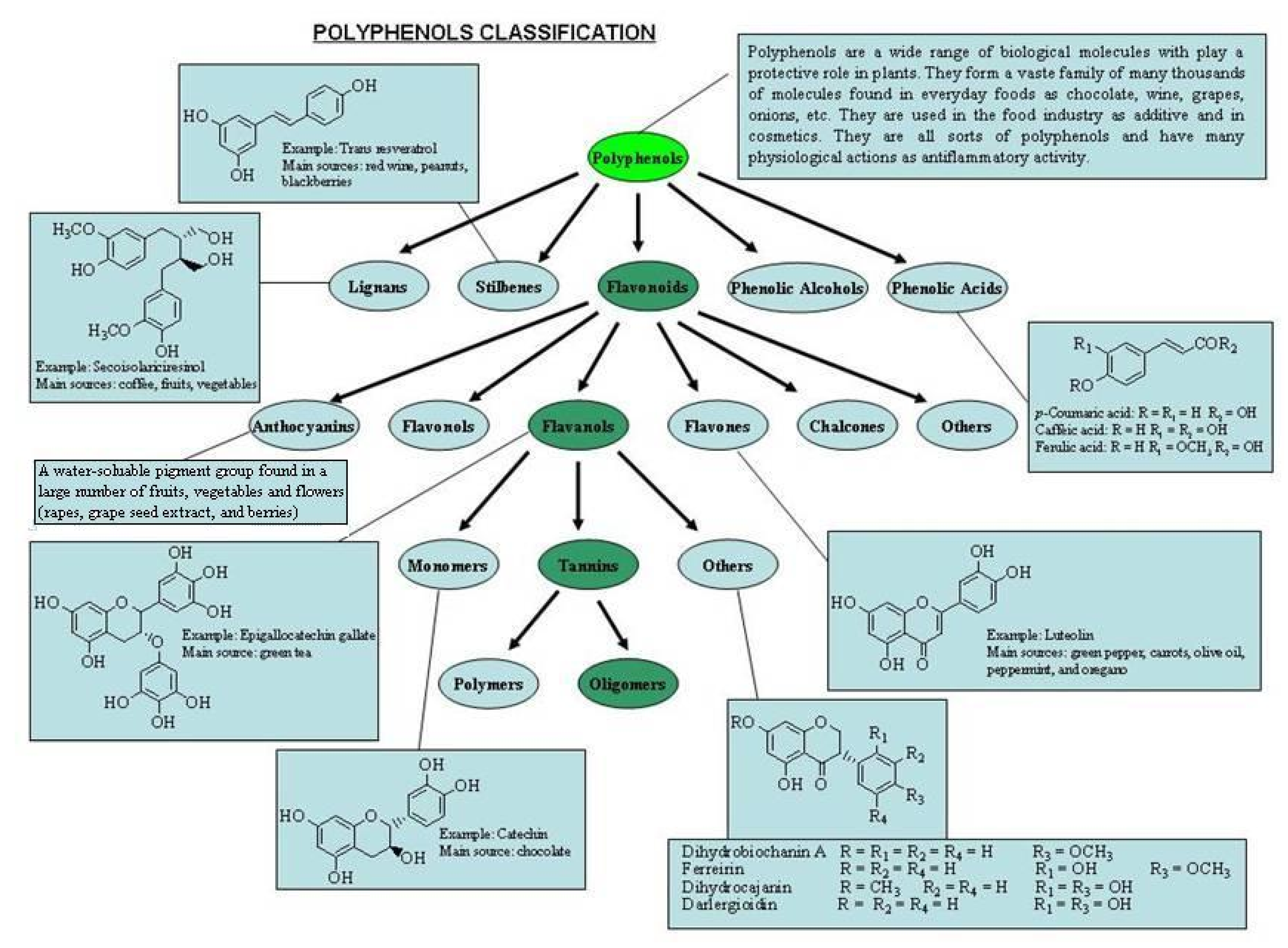
1.1. Classification of Polyphenols
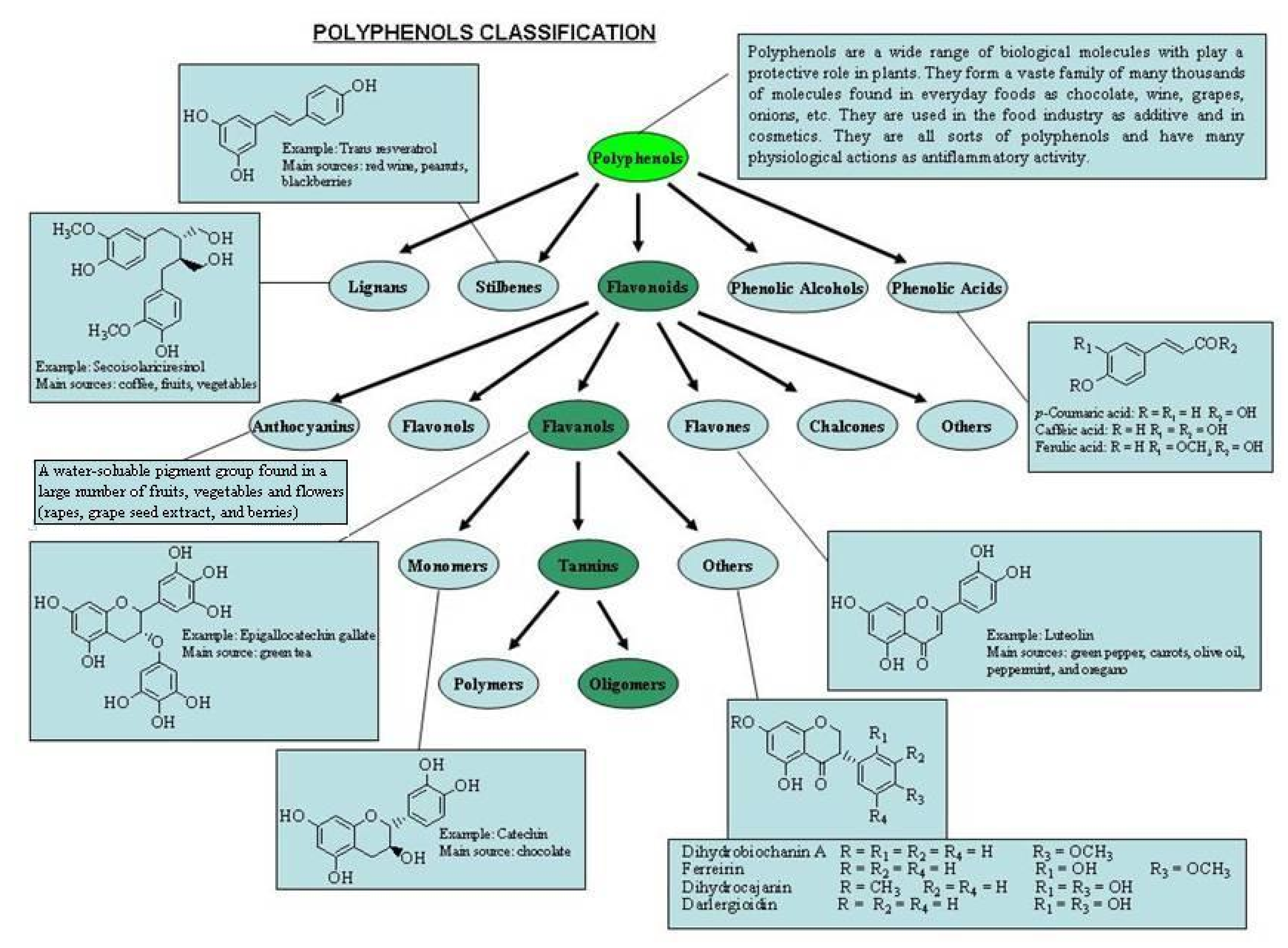
1.2. C6-C3 Phenylpropanoid Compounds
1.3. C6-C2-C6 Polyhydroxystilbenes
1.4. Lignin Derivatives
1.5. Categories of Polyphenols
2. Antibacterial Activity of Plant Polyphenols
3. Pathogenesis of Dental Caries
4. Anti-Cariogenic Action of Polyphenols
4.1. In Vitro Studies
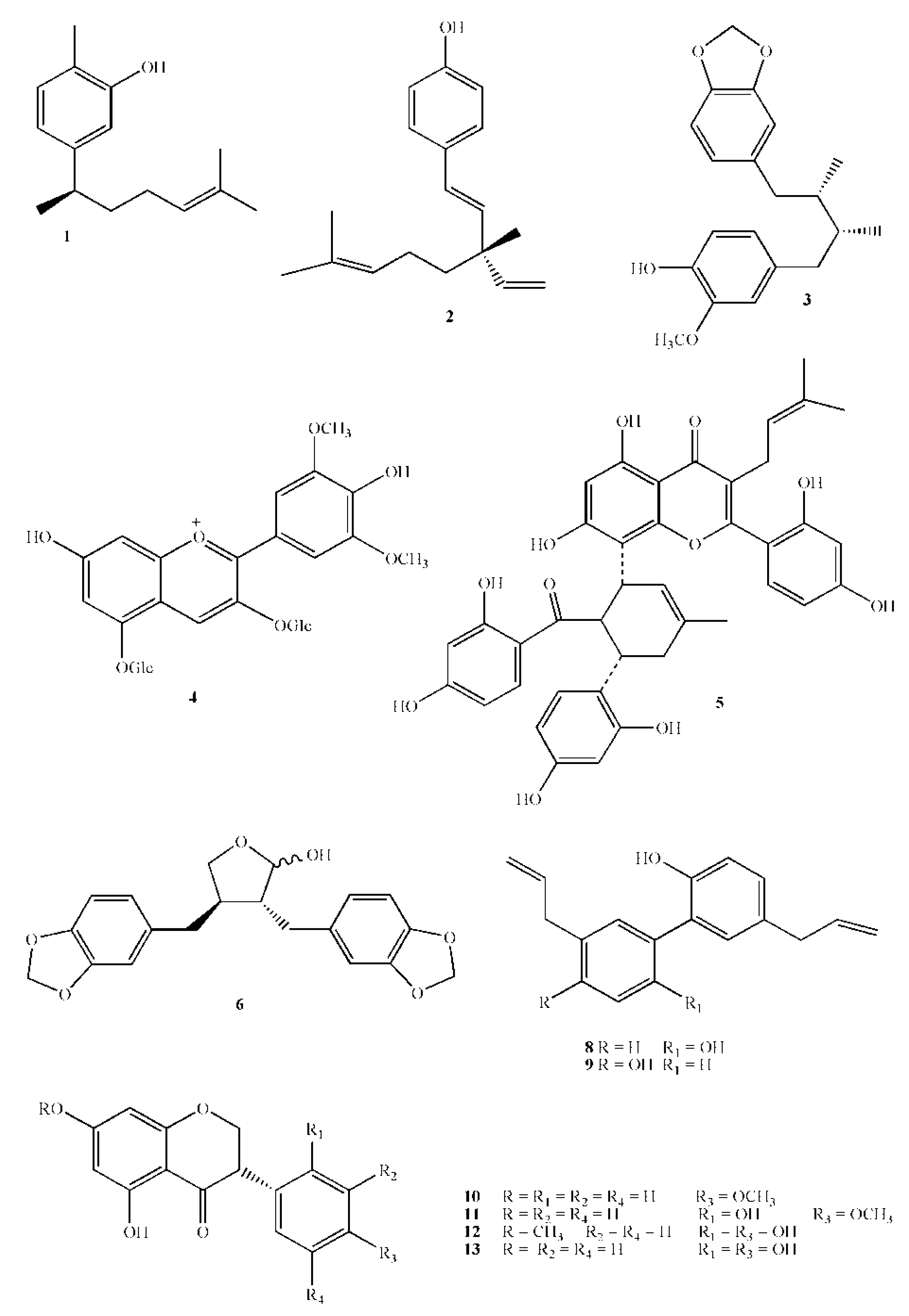
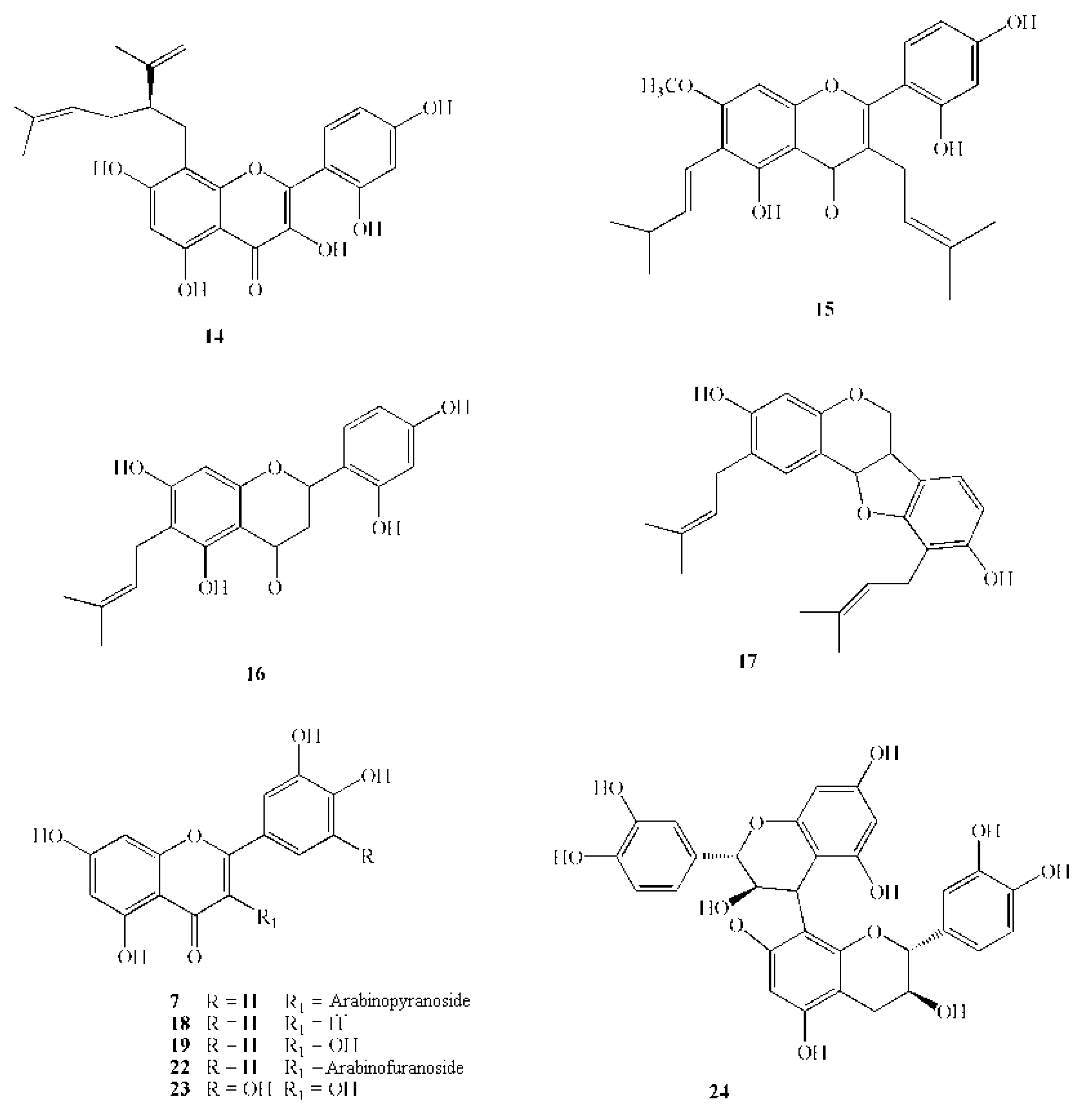
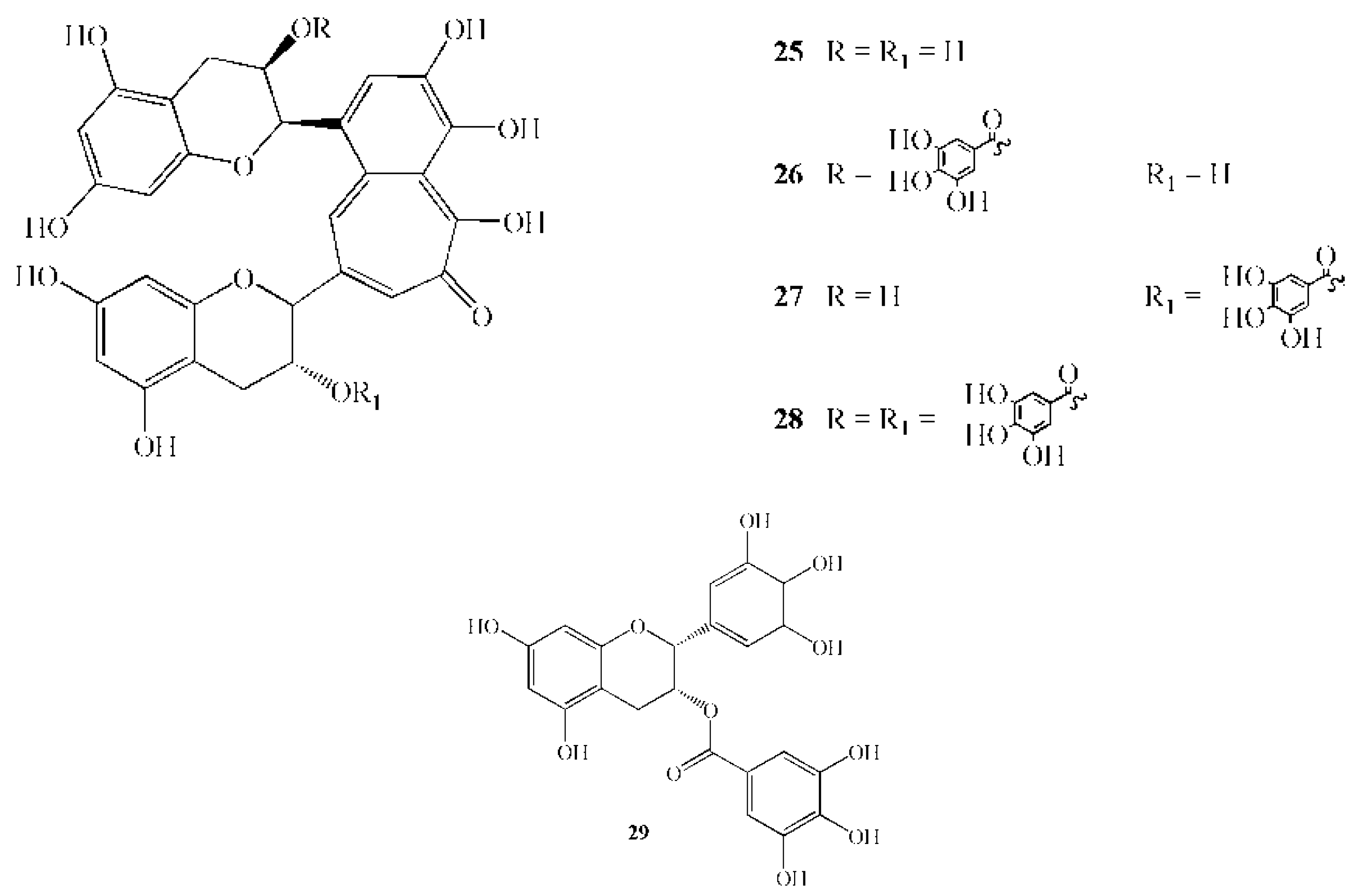

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N. | Name | Mol. weight | Plant name | Part of the plant | Activity against S. mutans | References |
|---|---|---|---|---|---|---|
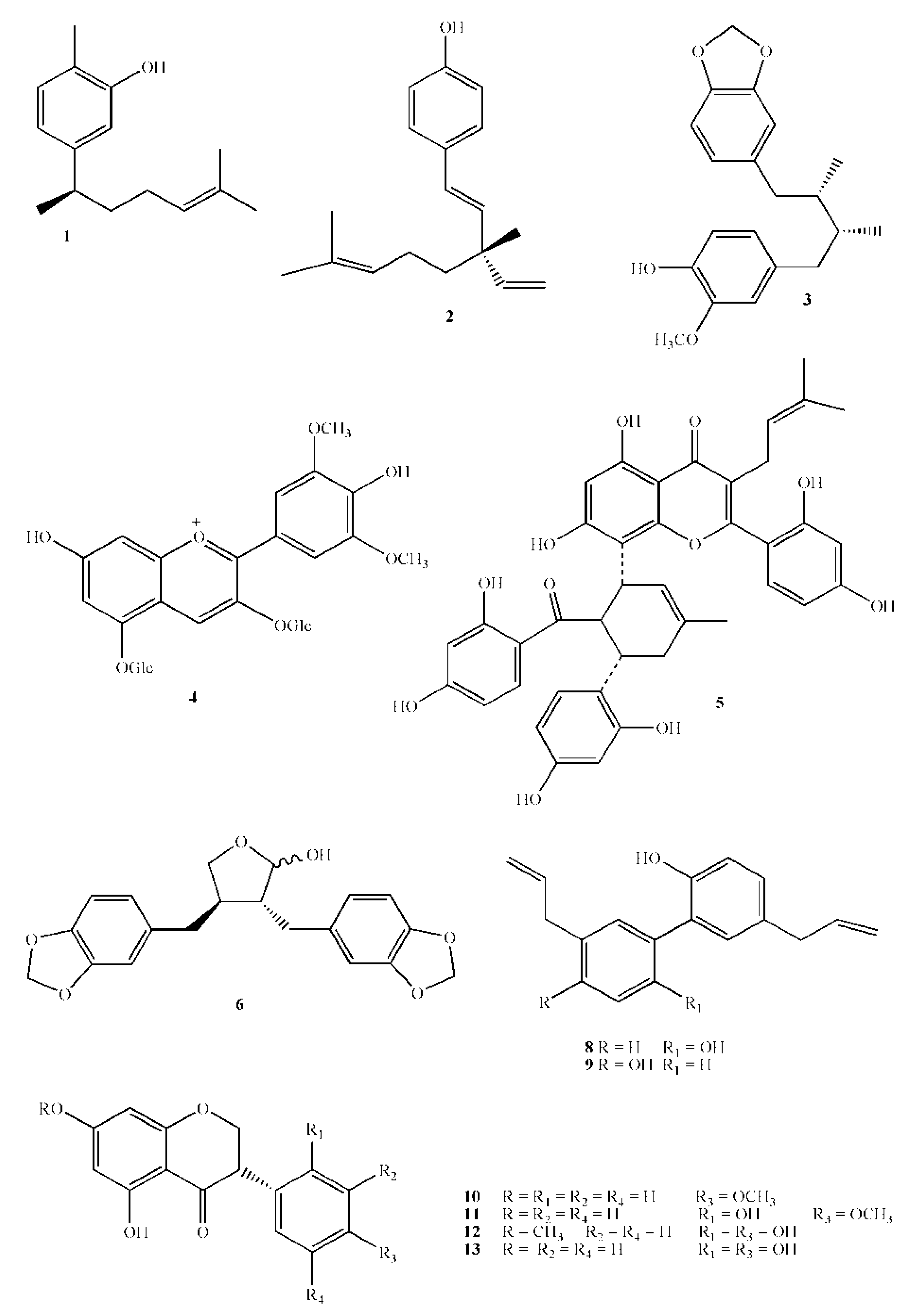
| 1 | Xanthorrhizol | 218.3 | Curcuma xanthorrihiza Roxb | rhizome | 5 mmol L−1 inhibit biofilm formation | [49] |
| 2 | Bakuchiol | 256.4 | Psoralea corylifolia L. | seeds | 20 µg/mL prevented growth | [50] |
| 3 | Macelignan | 328.4 | Myristica fragrans Houtt. | seeds | 10 μ/mL for 30’ exposure removed >50% of primary biofilm formed by S. mutans, S. sanguinis and A . viscosus | [51] |
| 4 | Malvin | 655.2 | Alcea longipedicellata I. Riedl | flowers | M.I.C. 0.16 mg/mL for S. mutans | [52] |
| 5 | Kuwanon G | 692.7 | Morus alba L. | Root bark | M.I.C. 8 μg/mL | [53] |
| 6 | (−)-Cubebin | 356.4 | Piper cubeba L. | seeds | M.I.C. 0.32 mM | [54] |
| 7 | Guaijaverin | Psidium guaiava L. | leaves | M.I.C. 4 mg/mL | [55] | |
| 8 | Magnolol | 266.3 | Magnolia officinalis | bark | 0.32 mg/mL reduced by 87.3% GTF activity | [56] |
| 9 | Honokiol | 266.3 | Magnolia officinalis | bark | 0.32 mg/mL reduced by 58.1% GTF activity | [56] |
| 10 | Dihydrobiochanin A | 286.3 | Swartzia polyphylla DC | heartwood | M.I.C. 50 µg/mL | [65] |
| 11 | Ferreirin | 302.3 | Swartzia polyphylla DC | heartwood | M.I.C. 50 µg/mL | [65] |
| 12 | Dihydrocajanin | 302.3 | Swartzia polyphylla DC | heartwood | M.I.C. 100 µg/mL | [65] |
| 13 | Dalbergioidin | 288.3 | Swartzia polyphylla DC | heartwood | M.I.C. 100 µg/mL | [65] |
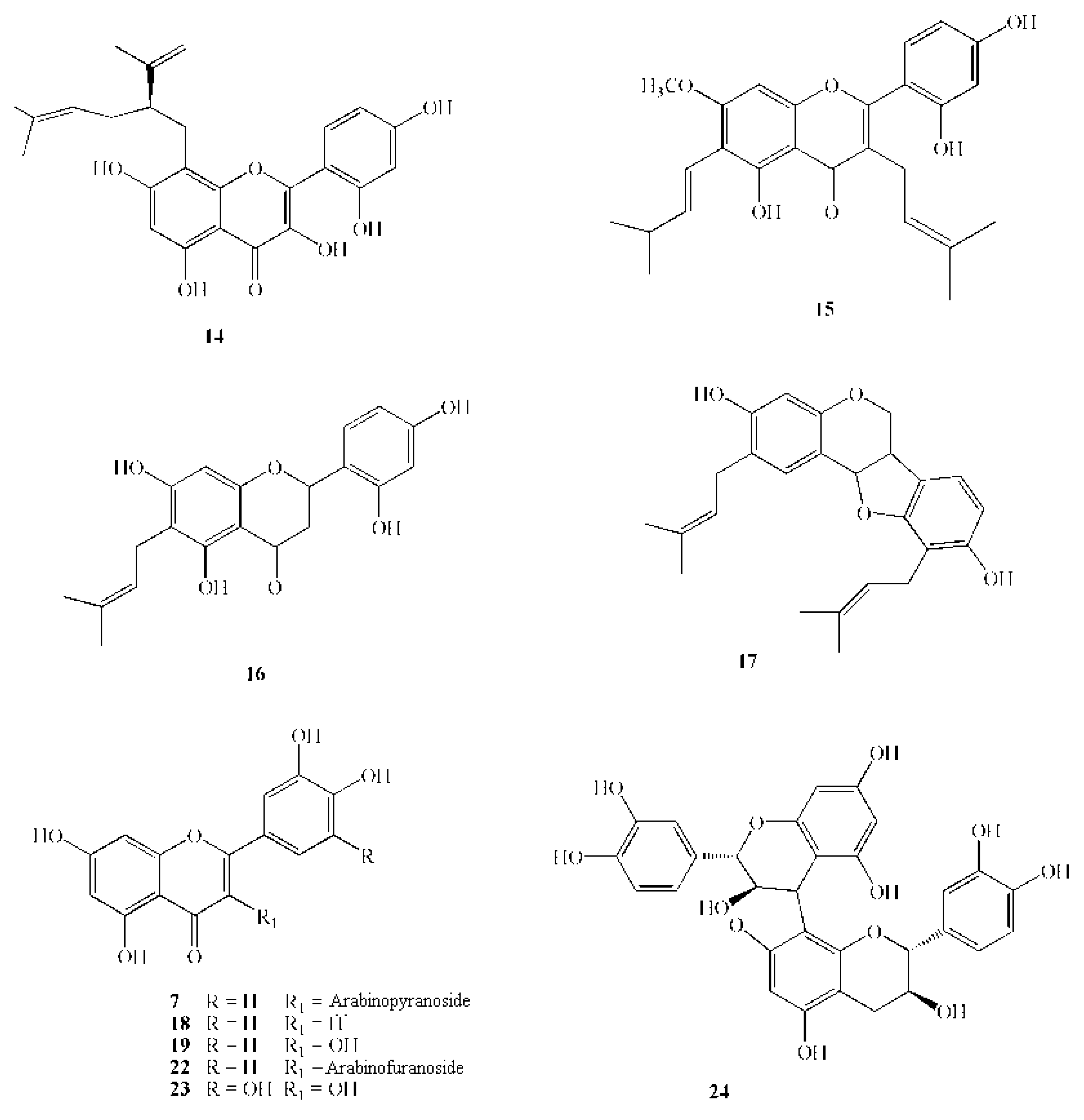
| 14 | Lavandulylflavanone | 438.5 | Sophora exigua Craigg | heartwood | Growth inhibition in the range 1.56–6.25 µg/mL | [66] |
| 15 | Artocarpin | 436.5 | Artocarpus heterophyllus Lam. | heartwood | M.I.C. 6.25 µg/mL | [67] |
| 16 | Artocarpesin | 354.4 | Artocarpus heterophyllus Lam. | heartwood | M.I.C. 6.25 µg/mL | [67] |
| 17 | Erycristagallin | 392.5 | Erythrina variegata L. | root | M.I.C. 6.25 µg/mL | [68] |
| 18 | Luteolin | 286.2 | Perilla frutescens Britton var. japonica Hara. | seeds | M.I.C. 50–100 µg/mL (on different S. mutans strains) | [69] |
| 19 | Quercetin | 302.2 | Commercial source | - | Inhibition of adhesive glucan format ion in the range 1.5–50 µg/mL | [73] |
| 20 | Proanthocyanidins | / | Humulus lupulus L. | bracts | 0.01%, Hop Bracts Polyphenols (HBP) containing 35% proanthocyanidins caused 80% inhibition of GTF | [79] |
| 21 | Tannins | / | Areca catechu L. | nut | 50% of growth inhibition at a 15% concentration | [82] |
| 22 | Quercetin-3-arabinofuranoside | 434.3 | Vaccinium macrocarpon Ait. | fruit | 21–41% Inhibition of GTF activity at500 mmol L−1 | [82] |
| 23 | Myricetin | 318.0 | Vaccinium macrocarpon Ait. | fruit | 15-28% Inhibition of GTF activity at500 mmol L−1 | [82] |
| 24 | Procyanidin A2 | 576.1 | Vaccinium macrocarpon Ait. | fruit | 21–41% Inhibition of GTF activity at500 mmol L−1 | [82] |
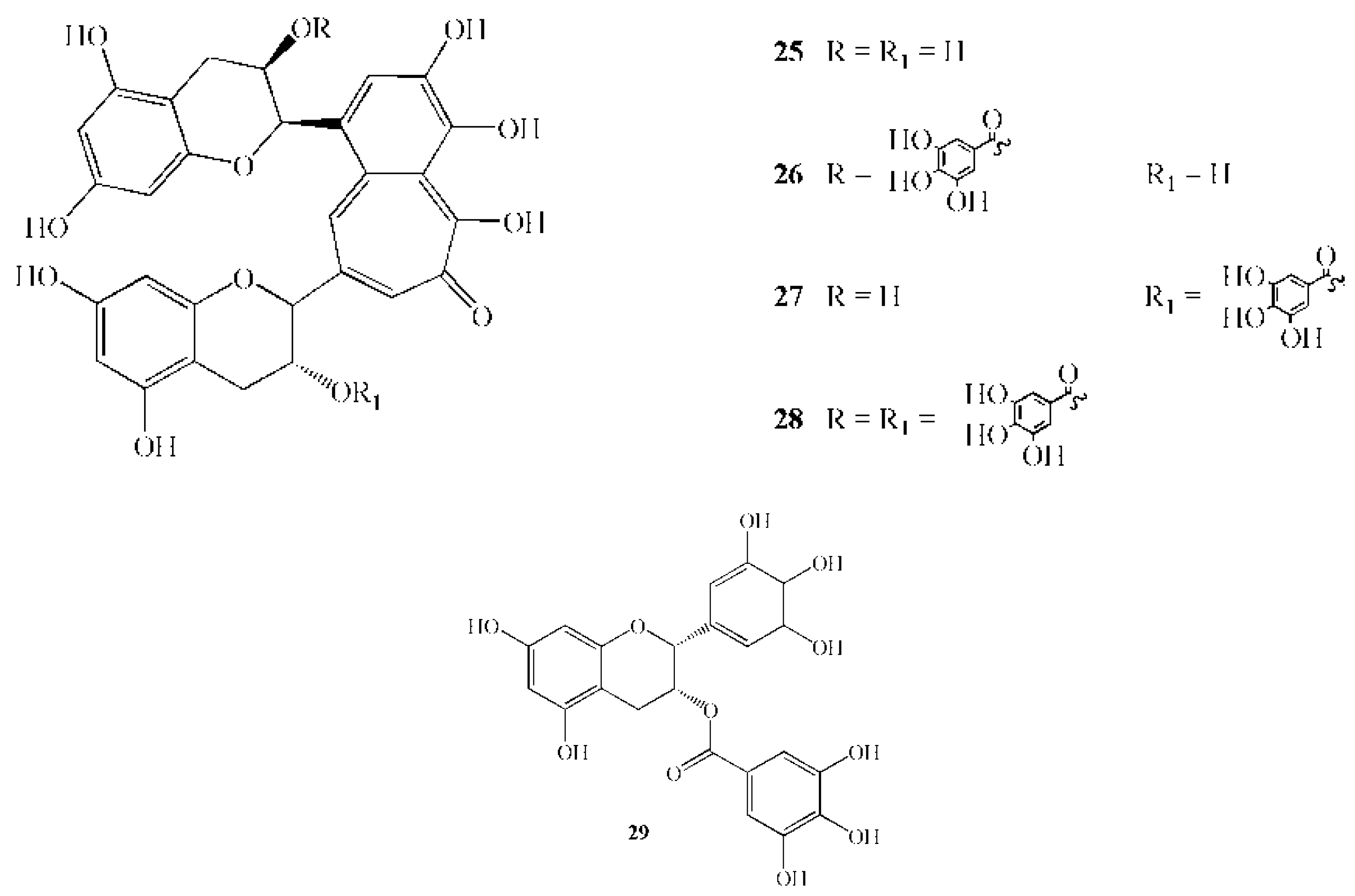
| 25 | Theaflavin | 564.1 | Camellia sinensis L. | leaves | Inhibition of GTF activity in the range 1–10 mM | [85] |
| 26 | Theaphlavin monogallate A | 716.3 | Camellia sinensis L. | leaves | Inhibition of GTF activity in the range 1–10 mM | [85] |
| 27 | Theaphlavin monogallate B | 716.3 | Camellia sinensis L. | leaves | Inhibition of GTF activity in the range 1–10 mM | [85] |
| 28 | Theaphlavin digallate | 868.1 | Camellia sinensis L. | leaves | Inhibition of GTF activity in the range 1–10 mM | [85] |
| 29 | Epigallocathechin gallate | 458.4 | Camellia sinensis L. | leaves | 167 mg/L caused 91% growth inhibition* | [85] |
4.2. Inhibition of Adherence
4.3. Inhibition of Glucosyltransferase and Amylase
4.4. In Vivo Studies
- (1) Mouthwashes with good substantivity and antibacterial spectrum with a good anti-plaque effects. To this group belong biguanides as chlorhexidine; the effect of concentrated 1% chlorhexidine gel, on oral bacteria salivary levels can be observed after a couple of applications but this use requires professional supervision [91];
- (2) Mouthwashes agents with little or no substantivity but with a good antibacterial spectrum. They have plaque inhibitory effects but lack true anti-plaque effects. In this category are included: cetyl pyridinium chloride, a quaternary ammonium compound, Listerine, which contains essential oil and phenolics (menthol, thymol, and eucalyptol), and triclosan, a trichlora-2'-hydroxydiphenyl ether;
- (3) Antiseptic mouthwashes that have be shown to have antibacterial effects in vitro but in clinical studies have been shown to have low/negligible plaque inhibitory effects. Hexetidine (Oraldene), povidone iodine, oxygenating agents and the natural product sanguinarine, a benzophenanthridine alkaloid, are members of this third group.
5. Conclusions
References
- Arakawa, H.; Maeda, M.; Okubo, S.; Shimamura, T. Role of hydrogen peroxide in bactericidal action of catechin. Biol. Pharm. Bull. 2004, 27, 277–281. [Google Scholar]
- Badria, F.A.; Zidan, O.A. Natural products for dental caries prevention. J. Med. Food 2004, 7, 381–384. [Google Scholar] [CrossRef]
- Banas, J.A.; Vickerman, M.M. Glucan-binding proteins of the oral streptococci. Crit. Rev. Oral Biol. Med. 2003, 14, 89–99. [Google Scholar]
- Banas, J.A. Virulence properties of Streptococcus mutans. Front. Biosci. 2004, 9, 1267–1277. [Google Scholar] [CrossRef]
- Bernaert, H.; Allegaert, L. Topical Skin Cosmetics Comprising a Cocoa Polyphenol Extract Combination with SUS-Rich Fat. Fat. U.S. Patent 2009/0233518 A1, 22 October 2009. [Google Scholar]
- Bhattacharya, A.; Sood, P.; Citovsky, V. The roles of plant phenolics in defence and communication during Agrobacterium and Rhizobium infection. Mol. Plant Pathol. 2010, 11, 705–719. [Google Scholar]
- Borchardt, J.R.; Wyse, D.L.; Sheaffer, C.C.; Kauppi, K.L.; Fulcher, R.G.; Ehlke, N.J.; Biesboer, D.D.; Bey, R.F. Antioxidant and antimicrobial activity of seed from plants of the Mississippi river basin. J. Med. Plants Res. 2008, 2, 81–93. [Google Scholar]
- Bowden, G.H. Controlled environment model for accumulation of biofilms of oral bacteria. Methods Enzymol. 1999, 310, 216–224. [Google Scholar]
- Bowen, W.H. Nature of plaque. Oral Sci. Rev. 1976, 9, 3–21. [Google Scholar]
- Burne, R.A. Oral streptococci products of the ir environment. J. Dent. Res. 1998, 77, 445–452. [Google Scholar]
- Cho, Y.S.; Schiller, N.L.; Kahng, H.Y.; Oh, K.H. Cellular responses and proteomic analysis of Escherichia coli exposed to green tea polyphenols. Curr. Microbiol. 2007, 55, 501–506. [Google Scholar] [CrossRef]
- Cutillo, F.; D'Abrosca, B.; DellaGreca, M.; Fiorentino, A.; Zarrelli, A. Terpenoids and phenol derivatives from Malva silvestris. Phytochemistry 2006, 67, 481–485. [Google Scholar] [CrossRef]
- Cutillo, F.; DellaGreca, M.; Gionti, M.; Previtera, L.; Zarrelli, A. Phenols and lignans from Chenopodium album. Phytochem. Analysis 2006, 17, 344–349. [Google Scholar] [CrossRef]
- D'Abrosca, B.; DellaGreca, M.; Fiorentino, A.; Monaco, P.; Zarrelli, A. Low molecular weight phenols from the bioactive aqueous fraction of Cestrum parqui. J. Agr. Food Chem. 2004, 52, 4101–4108. [Google Scholar] [CrossRef]
- DellaGreca, M.; Previtera, L.; Temussi, F.; Zarrelli, A. Low-molecular-weight components of olive oil mill waste-waters. Phytochem. Analysis 2004, 15, 184–188. [Google Scholar] [CrossRef]
- Duarte, S.; Gregoire, S.; Singh, A.P.; Vorsa, N.; Schaich, K.; Bowen, W.; Koo, H. Inhibitory effects of cranberry polyphenols on formation and acidogenicity of Streptococcus mutans biofilms. FEMS Microbiol. Lett. 2006, 257, 50–56. [Google Scholar] [CrossRef]
- Esmeelian, B.; Kamrani, Y.Y.; Amoozegar, M.A.; Rahamani, S.; Rahimi, M.; Amanlou, M. Anti-cariogenic propreties of malvidin-3,5-diglucoside isolated from Alcea longipedicellata against oral bacteria. Int. J. Pharmacol. 2007, 3, 468–474. [Google Scholar] [CrossRef]
- Fattouch, S.; Caboni, P.; Coroneo, V.; Tuberoso, C.I.G.; Angioini, A.; Dessi, S.; Marzouki, N.; Cabras, P. Antimicrobial activity of Tunisian quince (Cydonia oblonga Miller) pulp and peel polyphenolic extracts. J. Agric. Food Chem. 2007, 5, 963–969. [Google Scholar]
- Featherstone, J.D. The science and practice of caries prevention. J. Am. Dent. Assoc. 2000, 131, 887–899. [Google Scholar]
- Figueira, L. Resveratrol: Role in cardiovascular disease and cancer. Informe Medico (Caracas, Venezuela) 2010, 12, 73–83. [Google Scholar]
- Fiorentino, A.; DellaGreca, M.; D'Abrosca, B.O.P.; Golino, A.; Izzo, A.; Zarrelli, A.; Monaco, P. Lignans, neolignans and sesquilignans from Cestrum parqui l'Her. Biochem. Syst. Ecol. 2007, 35, 392–396. [Google Scholar] [CrossRef]
- Freedman, M.L.; Tanzer, J.M. Dissociation of plaque formation from glucan-induced agglutination in mutants of Streptococcus mutans. Infect. Immun. 1974, 10, 189–196. [Google Scholar]
- Furiga, A.; Lonvaud-Funel, A.; Badet, C. In vitro study of antioxidant capacity and antibacterial acitivity on oral anaerobes of a grape seed extract. Food Chem. 2009, 113, 1037–1040. [Google Scholar] [CrossRef]
- Geissman, T.A.; Hinreiner, E. Theories of the biogenesis of flavonoid compounds. Botan. Rev. 1952, 18, 77–164. [Google Scholar]
- Gregoire, S.; Singh, A.P.; Vorsa, N.; Koo, H. Influence of cranberry phenolics on glucan synthesis by glucosyltransferases and Streptococcus mutans acidogenicity. J. Appl. Microbiol. 2007, 103, 1960–1968. [Google Scholar] [CrossRef]
- Grollier, J.F.; Garnier, L.; Boussouira, B. Cosmetic treatment process based on fruit or vegetable polyphenols. PCTInt. Appl. WO 2009109946, 2009. [Google Scholar]
- Haslam, E.; Lilley, T.H.; Warminski, E.; Liao, H.; Cai, Y.; Martin, R.; Gaffney, S.H.; Goulding, P.N.; Luck, G. Polyphenol complexation. A study in molecular recognition. ACS Symp. Ser. 1992, 506, 8–50. [Google Scholar] [CrossRef]
- Hattori, M.; Kusumoto, I.T.; Namba, T.; Ishigami, T.; Hara, Y. Effect of tea polyphenols on glucan synthesis by glucosyltransferase from Streptococcus mutans. Chem. Pharm. Bull. 1990, 38, 717–720. [Google Scholar] [CrossRef]
- Hu, L.; Wang, H.; Pei, J.; Liu, Y. Research progress of antitumor effects of resveratrol and its mechanism. Shandong Yiyao 2010, 50, 111–112. [Google Scholar]
- Hubert, B.; Eberl, L.; Feucht, W.; Polster, J. Influence of polyphenols on bacterial biofilm formation and quorum-sensing. Z. Naturforsch. 2003, 58, 879–884. [Google Scholar]
- Ito, K.; Nakamura, Y.; Tokunaga, T.; Iijima, D.; Fukushima, K. Anti-cariogenic properties of a water-soluble extract from cacao. Biosci. Biotechnol. Biochem. 2003, 67, 2567–2573. [Google Scholar]
- Ito, M.; Uyeda, M.; Iwanami, T.; Nagakawa, Y. Flavonoids as a possible preventive of dental caries. Agric. Biol. Chem. 1984, 48, 2143–2145. [Google Scholar] [Green Version]
- Jiang, N. Medicinal composition of plant active components for antiaging and antianaphylaxis. Faming Zhuanli Shenqing Gongkai Shuomingshu CN 101496772, 5 August 2009. [Google Scholar]
- Juneia, R.L.; Okubo, T.; Hung, K. Catechins. In Natural Food Antimicrobial; Naidu, A.S., Ed.; 2000; pp. 381–398. [Google Scholar]
- Kashket, S.; Paolino, V.J.; Lewis, D.A.; van Houte, J. In-vitro inhibition of glucosyltransferase from the dental plaque bacterium Streptococcus mutans by common beverages and food extracts. Arch. Oral Biol. 1985, 30, 821–826. [Google Scholar] [CrossRef]
- Katsura, H.; Tsukiyama, R.I.; Suzuki, A.; Kobayashi, M. In vitro Antimicrobial Activities of Bakuchiol against Oral Microorganisms. Antimicrob. Agents Chem. 2001, 45, 3009–3013. [Google Scholar] [CrossRef]
- Kim, J.H. Anti-bacterial action of onion (Allium cepa L.) extracts against oral pathogenic bacteria. J. Nihon Univ. Sch. Dent. 1997, 9, 136–141. [Google Scholar] [CrossRef]
- Korkina, L.G.; Mikhal’chik, E.; Suprun, M.V.; Pastore, S.; Dal Toso, R. Molecular mechanisms underlying wound healing and anti-inflammatory properties of naturally occurring biotechnologically produced phenylpropanoid glycosides. Cell. Mol. Biol. (Noisy-le-grand) 2007, 53, 84–91. [Google Scholar]
- Krishnan, R.; Maru, G.B. Inhibitory effect(s) of polymeric black tea polyphenol fractions on the formation of [(3)H]-B(a)P-derived DNA adducts. J. Agric. Food Chem. 2004, 52, 4261–4269. [Google Scholar] [CrossRef]
- Kurumatani, M.; Fujita, R.; Tagashira, M.; Shoji, T.; Kanda, T.; Ikeda, M.; Shoji, A.; Yanagida, A.; Shibusawa, Y.; Shindo, H.; Ito, Y. Analysis of polyphenols from hop bract region using CCC. J. Liq. Chromatogr. 2005, 28, 1971–1983. [Google Scholar]
- Lattanzio, V. Some physiological and ecological role of plant phenolics. Polyphénols Actualités 2006, 24, 5–9. [Google Scholar]
- Lee, M.J.; Lambert, J.D.; Prabhu, S.; Meng, X.; Lu, H.; Maliakal, P.; Ho, C.T.; Yang, C.S. Delivery of tea polyphenols to the oral cavity by green tea leaves and black tea extract. Cancer Epidemiol. Biomarkers Prev. 2004, 13, 132–137. [Google Scholar]
- Li, J.Y.; Zhan, L.; Barlow, J.; Lynch, R.J.; Zhou, X.D.; Liu, T.J. Effect of tea polyphenol on the demineralization and remineralization of enamel in vitro. Sichuan Da Xue Xue Bao Yi Xue Ban 2004, 35, 364–366. [Google Scholar]
- Llorach, R.; Urpi-Sarda, M.; Rotches-Ribalta, M.; Rabassa, M.; Andres-Lacueva, C. Resveratrol: From dietary intake to promising therapeutic molecule. Agro Food Ind. Hi-Tech. 2010, 21, 42–44. [Google Scholar]
- Luczaj, W.; Skrzydlewska, E. Antioxidative properties of black tea. Prev. Med. 2005, 40, 910–918. [Google Scholar] [CrossRef]
- Manitto, P. Biosynthesis of Natural Products; Ellis Horwood Ltd.: Chichester and New York, UK, 1981. [Google Scholar]
- Matsumoto, M.; Minami, T.; Sasaki, H.; Sobue, S.; Hamada, S.; Ooshima, T. Inhibitory effects of oolong tea extract on caries-inducing properties of mutans streptococci. Caries Res. 1999, 33, 441–445. [Google Scholar] [CrossRef]
- Milgrom, P.; Riedy, C.A.; Weinstein, P.; Tanner, A.C.; Manibusan, L.; Bruss, J. Dental caries and its relationship to bacterial infection, hypoplasia, diet, and oral hygiene in 6- to 36-month-old children. Community Dent. Oral Ep idemiol. 2000, 28, 295–306. [Google Scholar]
- Murphy, C.M. Plant products as antimicrobial agents. Clin. Microbiol. 1999, 12, 564–582. [Google Scholar]
- Namba, T.; Tsumezuka, M.; Hattori, M. Dental caries by traditional Chinese medicines (part II), potent antibacterial action of Magnoliae Cortex extracts against Streptococcus mutans. Planta Med. 1982, 44, 100–106. [Google Scholar] [CrossRef]
- Nikitina, V.S.; Kuz’mina, Y.L.; Melent’ev, A.I.; Shendel, G.V. Antibacterial activity of polyphenolic compounds isolated from plants of Geraniaceae and Rosaceae families. Appl. Biochem. Microbiol. 2007, 43, 629–634. [Google Scholar]
- Ooshima, T.; Minami, T.; Aono, W.; Izumitani, A.; Sobue, S.; Fujiwara, T.; Kawabata, S.; Hamada, S. Oolong tea polyphenols inhibit experimental dental caries in SPF rats infected with mutans streptococci. Caries Res. 1993, 27, 124–129. [Google Scholar] [CrossRef]
- Ooshima, T.; Minami, T.; Matsumoto, M; Fujiwara, T.; Sobue, S.; Hamada, S. Comparison of the cariostatic effects between regimens to administer oolong tea polyphenols in SPF rats. Caries Res. 1998, 32, 75–80. [Google Scholar] [CrossRef]
- Ooshima, T.; Osaka, Y.; Sasaki, H.; Osawa, K.; Yasuda, H.; Matsumura, M.; Sobue, S.; Matsumoto, M. Caries inhibitory activity of cacao bean husk extract in in-vitro and animal experiments. Arch. Oral Biol. 2000, 45, 639–645. [Google Scholar] [CrossRef]
- Osawa, K.; Yasuda, H.; Maruyama, T.; Morita, H.; Takeya, K.; Itokawa, H. Isoflavanones from the heartwood of Swartzia polyphylla and their antibacterial activity against cariogenic bacteria. Chem. Pharm. Bull. 1992, 40, 2970–2974. [Google Scholar] [CrossRef]
- Osbourn, A.E. Preformed antimicrobial compounds and plant defense against fungal attack. Plant Cell 1996, 10, 1821–1831. [Google Scholar]
- Parfitt, G.J. The speed of development of the carious cavity. Br. Dent. J. 1956, 100, 204–207. [Google Scholar]
- Park, K.M.; You, J.S.; Lee, H.Y.; Baek, N.I., Hwang. Kuwanon G: an antibacterial agent from the root bark of Morus alba against oral pathogens. J. Ethnopharmacol. 2003, 84, 181–185. [Google Scholar] [CrossRef]
- Percival, R.S.; Devine, D.A.; Duggal, M.S.; Chartron, S.; Marsh, P.D. The effect of cocoa polyphenols on the growth, metabolism, and biofilm formation by Streptococcus mutans and Streptococcus sanguinis. Eur. J. Oral. Sci. 2006, 114, 343–348. [Google Scholar]
- Prabu, G.R.; Gnanamani, A.; Sadulla, S. Guaijaverin—a plant flavonoid as potential antiplaque agent against Streptococcus mutans. J. Appl. Microbiol. 2006, 101, 487–495. [Google Scholar] [CrossRef]
- Rao, S.; Gruber, J.V.; Brooks, G.J. Personal care composition containing yeast/ polyphenol ferment extract. US Pat. Appl. Pub. US 20100021532 A1, 28 January 2010. [Google Scholar]
- Reynolds, E.C.; Black, C.L. Cariogenicity of a confection supplemented with sodium caseinate at a palatable level. Caries Res. 1989, 23, 368–370. [Google Scholar]
- Rukayadi, Y.; Hwang, J.K. In vitro activity of xanthorrhizol against Streptococcus mutans biofilms. Lett. Appl. Microbiol. 2006, 42, 400–404. [Google Scholar] [CrossRef]
- Saito, N. Anti-caries effects of polyphenol compound from Camellia sinensis. Nichidai Koko Kagaku 1990, 16, 154–163. [Google Scholar]
- Sakagami, H.; Oi, T.; Satoh, K. Prevention of oral diseases by polyphenols. In vivo 1999, 13, 155–171. [Google Scholar]
- Sampaio, F.C.; Pereira, M.S.; Dias, C.S.; Costa, V.C.; Conde, N.C.; Buzalaf, M.A. In vitro antimicrobial activity of Caesalpinia ferrea Martius fruits against oral pathogens. J. Ethnopharmacol. 2009, 15, 289–294. [Google Scholar]
- Sang, S.; Lambert, J.D.; Tian, S.; Hong, J.; Hou, Z.; Ryu, J.H.; Stark, R.E.; Rosen, R.T.; Huang, M.T.; Yang, C.S.; Ho, C.T. Enzymatic synthesis of tea theaflavin derivatives and their anti-inflammatory and cytotoxic activities. Bioorg. Med. Chem. 2004, 12, 459–467. [Google Scholar]
- Sato, M.; Fujiwara, S.; Tsuchiya, H.; Fujii, T.; Tinuma, M.; Tosa, H.; Ohkawa, Y. Flavones with antibacterial activity against cariogenic bacteria. J. Ethnopharmacol. 1996, 54, 171–176. [Google Scholar]
- Sato, M.; Tanaka, H.; Fujiwara, S.; Hirata, M.; Yamaguchi, R.; Etoh, H.; Tokuda, C. Antibacterial property of isoflavonoids isolated from Erythrina variegata against cariogenic oral bacteria. Phytomedicine 2003, 10, 427–433. [Google Scholar] [CrossRef]
- Signoretto, C.; Burlacchini, G.; Bianchi, F.; Cavalleri, G.; Canepari, P. Differences in microbiological composition of saliva and dental plaque in subjects with different drinking habits. New Microbiol. 2006, 29, 293–302. [Google Scholar]
- Silva, M.L.A.; Coimbra, H.S.; Pereira, A.C., Almeida; Lima, T.C.; Costa, E.S.; Vinholis, A.H.C.; Royo, V.A.; Silva, R.; Filho, A.A.S.; Cunha, W.R.; Furtado, N.A.J.C.; Martins, C.H.G.; Carvalho, T.C.; Bastos, J.K. Evaluation of Piper cubeba extract, (−)-cubebin and its semi-synthetic derivatives against oral pathogens. Phytother. Res. 2007, 21, 420–422. [Google Scholar] [CrossRef]
- Slimestad, R.; Fossen, T.; Vagen, I.M. Onions: a source of unique dietary flavonoids. J. Agric. Food Chem. 2007, 5, 10067–10080. [Google Scholar]
- Smullen, J.; Koutsou, G.A.; Foster, H.A.; Zumbé, A.; Storey, D.M. The antibacterial activity of plant extracts containing polyphenols against Streptococcus mutans. Caries Res. 2007, 41, 342–349. [Google Scholar] [CrossRef]
- Stauder, M.; Papetti, A.; Daglia, M.; Vezzulli, L.; Gazzani, G.; Varaldo, P.E.; Pruzzo, C. Inhibitory activity by barley coffee components towards Streptococcus mutans biofilm. Curr. Microbiol. 2010, 55, 1–5. [Google Scholar]
- Surarit, R., Koontongkaew. Inhibitory effect of betel-nut constituents on acid production of oral Streptococcus mutans. In Conference on Science and Technology of Thailand, Chulalongkorn University, Bangkok, Thailand, 1988; pp. 378–379.
- Tagashira, M.; Uchiyama, K.; Yoshimura, T.; Shirota, M.; Uemitsu, N. Inhibition by hop bract polyphenols of cellular adherence and waterinsoluble glucan synthesis of mutans streptococci. Biosci. Biotech. Biochem. 1997, 61, 332–335. [Google Scholar]
- Taguri, T.; Tanaka, T.; Kouno, I. Antimicrobial activity of 10 different plant polyphenols against bacteria causing food-borne disease. Biol. Pharm. Bull. 2004, 27, 1965–1969. [Google Scholar] [CrossRef]
- Tamba, Y.; Ohba, S.; Kubota, M.; Yoshioka, H.; Yoshioka, H.; Yamazaki, M. Single GUV method reveals interaction of tea catechin (−)-epigallocatechin gallate with lipid membranes. Biophys. J. 2007, 92, 3178–3194. [Google Scholar]
- Tanzer, J.M.; Livingston, J.; Thompson, A.M. The microbiology of primary dental caries in humans. J. Dent. Educ. 2001, 65, 1028–1037. [Google Scholar]
- Thimothe, J.; Bonsi, I.A.; Padilla-Zakour, O.I.; Koo, H. Chemical characterization of red wine grape (Vitis vinifera and Vitis interspecific hybrids) and pomace phenolic extracts and their biological activity against Streptococcus mutans. J. Agric. Food. Chem. 2007, 55, 10200–10207. [Google Scholar] [CrossRef]
- Tsuchiya, H.; Sato, M.; Tinuma, M.; Yokoyama, J.; Ohyama, M.; Tanaka, T.; Takase, I.; Namikawa, I. Inhibition of the growth of cariogenic bacteria in vitro by plant flavanones. Experientia 1994, 50, 846–849. [Google Scholar] [CrossRef]
- Vercauteren, J. Compositions of stilbenic polyphenolic derivatives, their preparation, and their use in the treatment of disease and aging. Fr. Demande 2923717 A1, 2009. [Google Scholar]
- Xie, Q.; Li, J.Y.; Zuo, Y.L.; Zhou, X.D. The effect of galla chinensis on the growth of cariogenic bacteria in vitro. Hua Xi Kou Qiang Yi Xue Za Zhi 2005, 23, 82–84. [Google Scholar]
- Yaegaki, K.; Tanaka, T.; Sato, T.; Murata, T.; Imai, T.; Tagashira, M.; Akazome, Y.; Hirai, N.; Ohtake, Y. Hop polyphenols suppress production of water-insoluble glucan by Streptococcus mutans and dental plaque growth in vivo. J. Clin. Dent. 2008, 19, 74–78. [Google Scholar]
- Yamammoto, H.; Ogawa, T. Antimicrobial activity of Perilla seed polyphenols against oral pathogenic bacteria. Biosci. Biotechnol. Biochem. 2002, 66, 921–924. [Google Scholar] [CrossRef]
- Yamanaka-Okada, A.; Sato, E.; Kouchi, T.; Kimizuka, R.; Kato, T. Inhibitory effect of cranberry polyphenol on cariogenic bacteria. Bull. Tokyo Dental Coll. 2008, 49, 107–112. [Google Scholar]
- Yanagida, A.; Kanda, T.; Oliveira Cordeiro, J.G. Inhibitory effects of apple polyphenols and related compounds on cariogenic factors of mutans streptococci. J. Agric. Food. Chem. 2000, 48, 5666–5671. [Google Scholar]
- Yanti; Rukayadi, Y.; Kim, K.H.; Hwang, J.K. In vitro anti-biofilm activity of macelignan isolated from Myristica fragrans Houtt. against oral primary colonizer bacteria. Phytother. Res. 2008, 22, 308–312. [Google Scholar] [CrossRef]
- Gunsolley, J.C. A meta-analysis of six-month studies of antiplaque and antigingivitis agents. J. Am. Dent. Assoc. 2006, 137, 1649–1657. [Google Scholar]
- Eley, B.M. Antibacterial agents in the control of supragingival plaque—a review. Br. Dental J. 1999, 186, 286–296. [Google Scholar]
- Decker, E.M.; Maier, G., Axmann; Brecx, M.; von Ohle, C. Effect of xylitol/chlorhexidine versus xylitol or chlorhexidine as single rinses on initial biofilm formation of cariogenic streptococci. Quintessence Int. 2008, 39, 17–22. [Google Scholar]
- Linke, H.A.; LeGeros, R.Z. Black tea extract and dental caries formation in hamsters. Int. J. Food Sci. Nutr. 2003, 54, 89–95. [Google Scholar]
- Shinada, K.; Tagashira, M.; Watanabe, H.; Sopapornamorn, P.; Kanayama, A.; Kanda, T.; Ikeda, M.; Kawaguchi, Y. Hop bract polyphenols reduced three-day dental plaque regrowth. J. Dent. Res. 2007, 86, 848–851. [Google Scholar]
- Ooshima, T.; Minami, T.; Aono, W.; Tamura, Y.; Hamada, S. Reduction of dental plaque deposition in humans by oolong tea extract. Caries Res. 1994, 28, 146–149. [Google Scholar] [CrossRef]
- Jones, C.; Woods, K.; Whittle, G.; Worthington, H.; Taylor, G. Sugar, drinks, deprivation and dental caries in 14-year-old children in the north west of England in 1995. Commu. Dent. Health 1999, 16, 68–71. [Google Scholar]
- Zhang, J.; Kashket, S. Inhibition of salivary amylase by black and green teas and their effects on the intraoral hydrolysis of starch. Caries Res. 1998, 32, 233–238. [Google Scholar] [CrossRef]
- Wynn, W.; Haldi, J.; Law, M.L. Influence of the ash of the cacao bean on the cariogenicity of a high-sucrose diet. J. Dent. Res. 1960, 39, 153–157. [Google Scholar]
- Strålfors, A. Inhibition of hamster caries by substances in chocolate. Arch. Oral Biol. 1967, 12, 959–962. [Google Scholar] [CrossRef]
- Verakaki, E.; Duggal, M.S. A comparison of different kinds of European chocolates on human plaque pH. Eur. J. Paediatr. Dent. 2003, 4, 203–210. [Google Scholar]
- Srikanth, R.K.; Shashikiran, N.D.; Subba Reddy, V.V. Chocolate mouth rinse: Effect on plaque accumulation and mutans streptococci counts when used by children. J. Indian Soc. Pedod. Prev. Dent. 2008, 26, 67–70. [Google Scholar] [CrossRef]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ferrazzano, G.F.; Amato, I.; Ingenito, A.; Zarrelli, A.; Pinto, G.; Pollio, A. Plant Polyphenols and Their Anti-Cariogenic Properties: A Review. Molecules 2011, 16, 1486-1507. https://doi.org/10.3390/molecules16021486
Ferrazzano GF, Amato I, Ingenito A, Zarrelli A, Pinto G, Pollio A. Plant Polyphenols and Their Anti-Cariogenic Properties: A Review. Molecules. 2011; 16(2):1486-1507. https://doi.org/10.3390/molecules16021486
Chicago/Turabian StyleFerrazzano, Gianmaria F., Ivana Amato, Aniello Ingenito, Armando Zarrelli, Gabriele Pinto, and Antonino Pollio. 2011. "Plant Polyphenols and Their Anti-Cariogenic Properties: A Review" Molecules 16, no. 2: 1486-1507. https://doi.org/10.3390/molecules16021486
APA StyleFerrazzano, G. F., Amato, I., Ingenito, A., Zarrelli, A., Pinto, G., & Pollio, A. (2011). Plant Polyphenols and Their Anti-Cariogenic Properties: A Review. Molecules, 16(2), 1486-1507. https://doi.org/10.3390/molecules16021486