Probing the Interaction of Human Serum Albumin with Norfloxacin in the Presence of High-Frequency Electromagnetic Fields: Fluorescence Spectroscopy and Circular Dichroism Investigations

Abstract
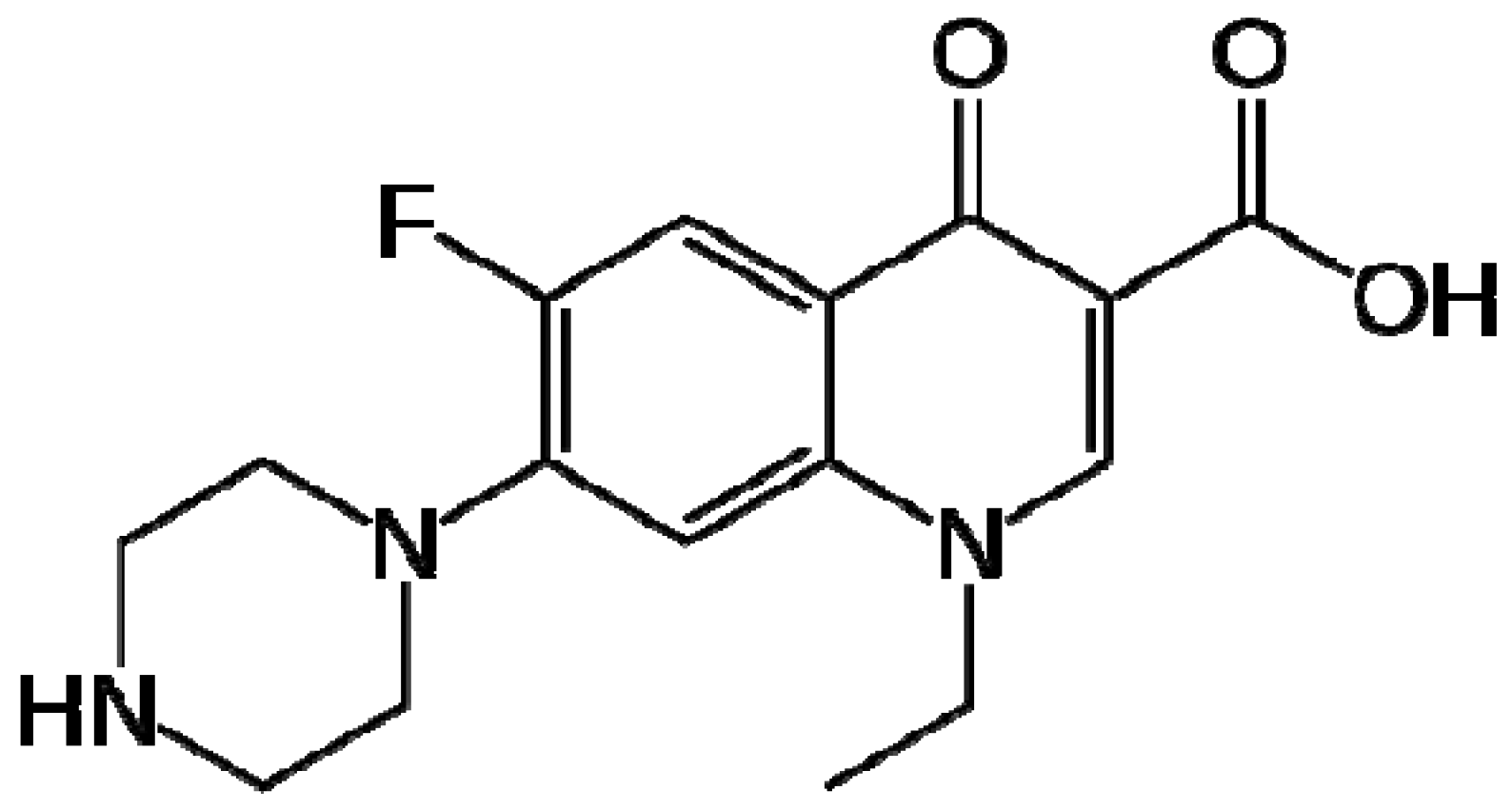
:1. Introduction
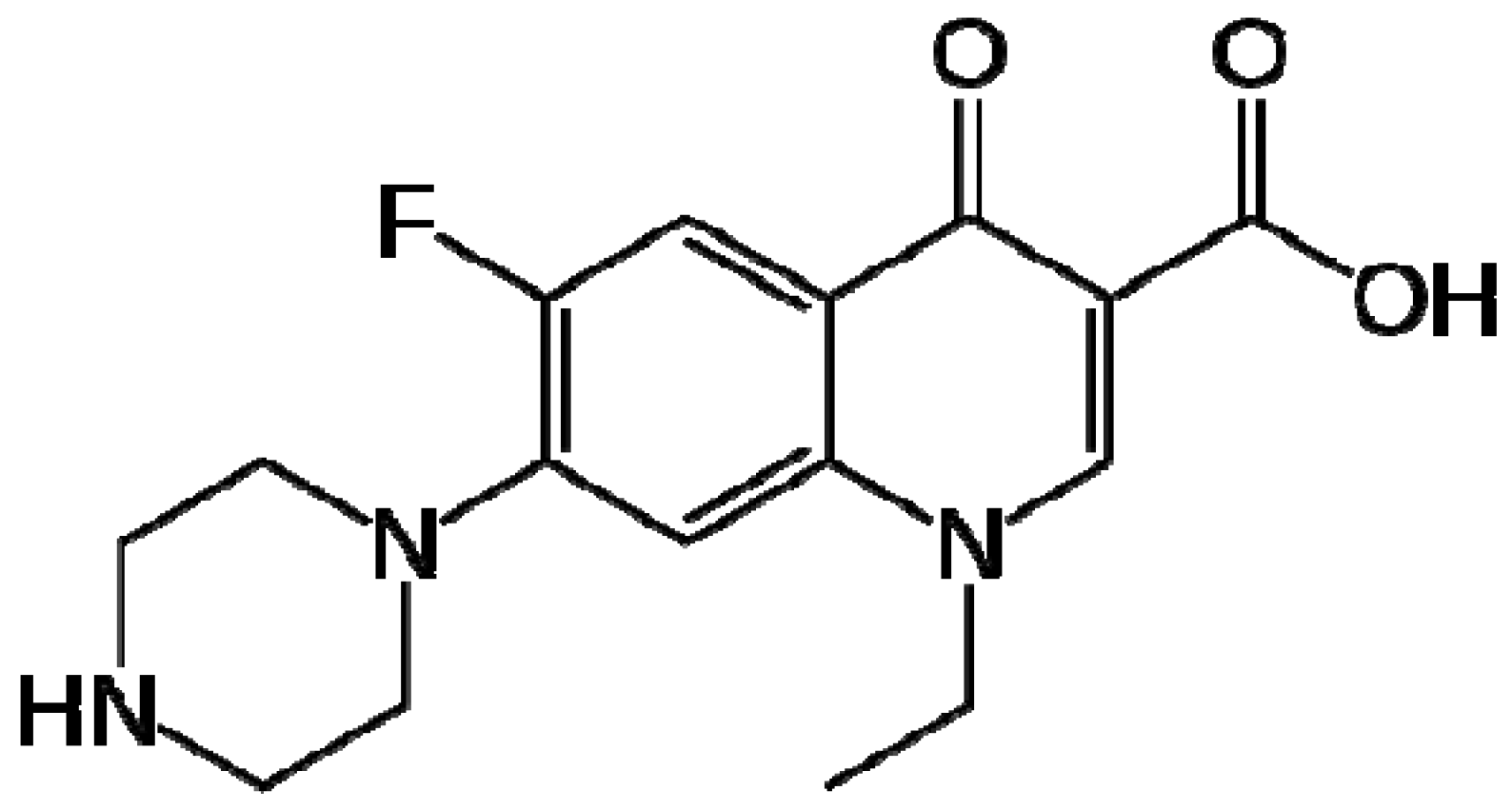
2. Results and Discussion
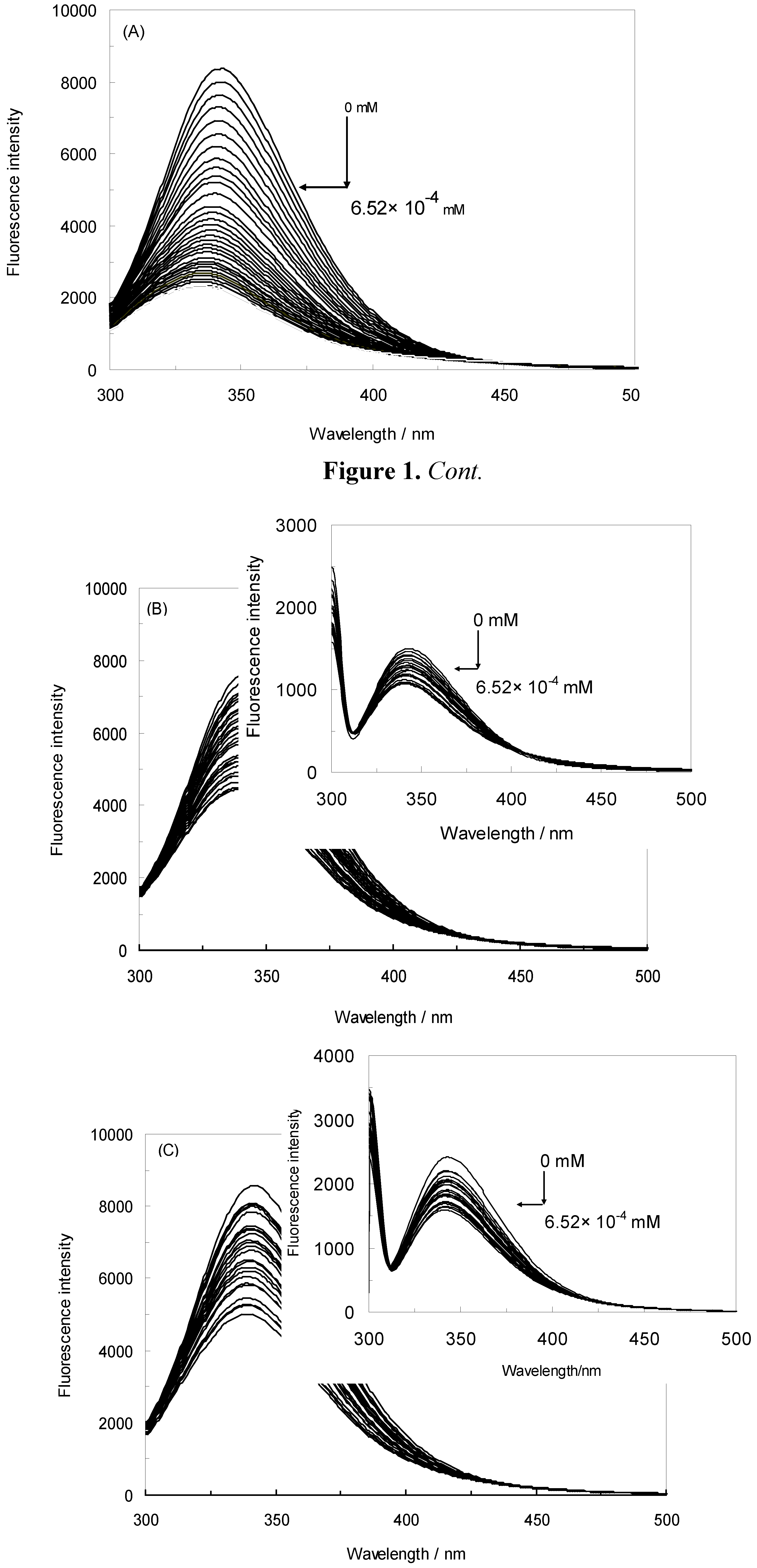
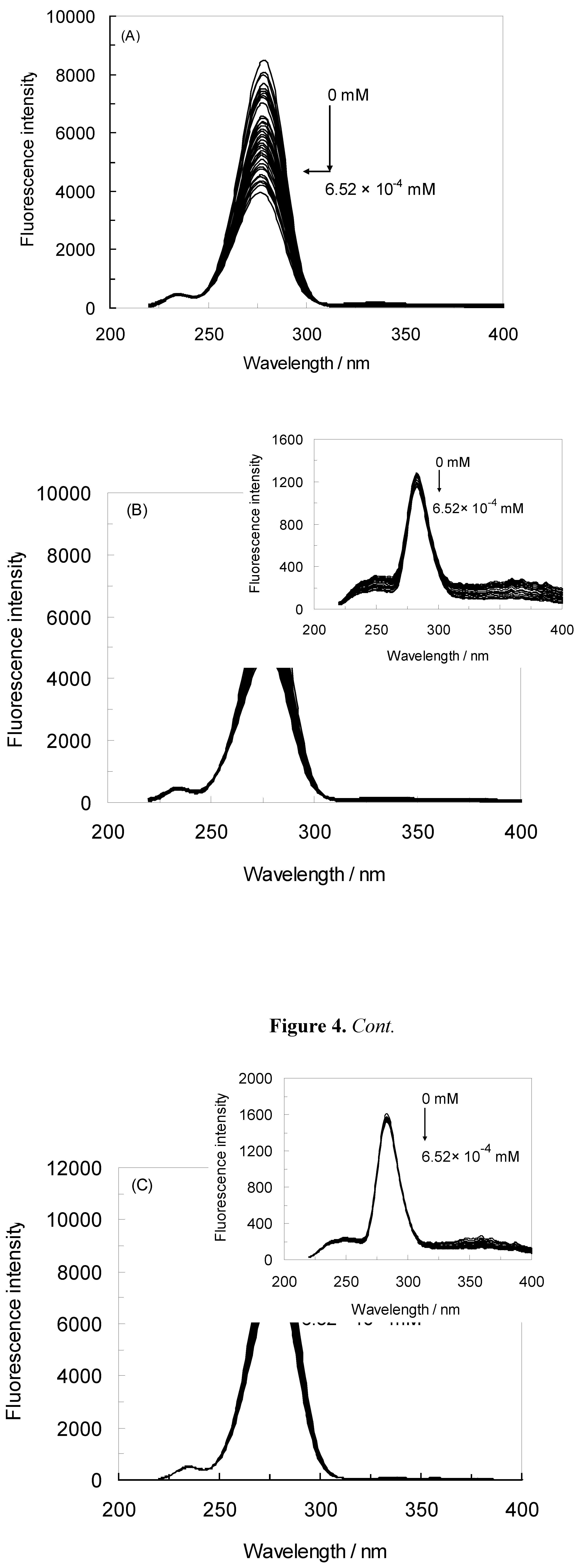
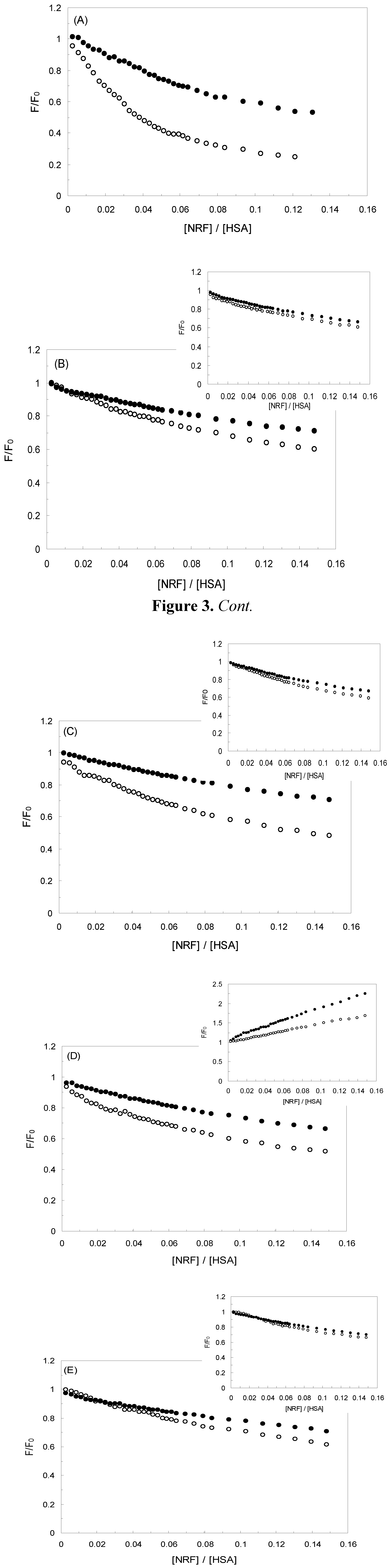
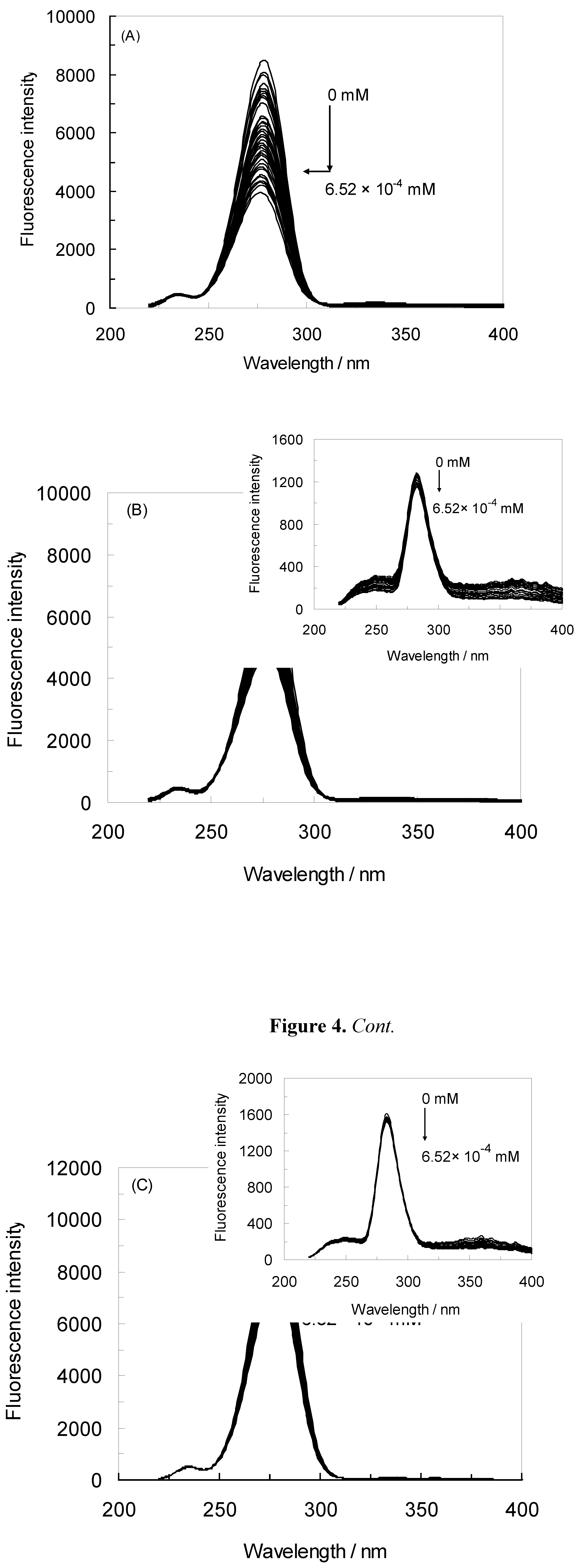
2.1. Fluorescence Quenching of HSA by NRF and Quenching Mechanism
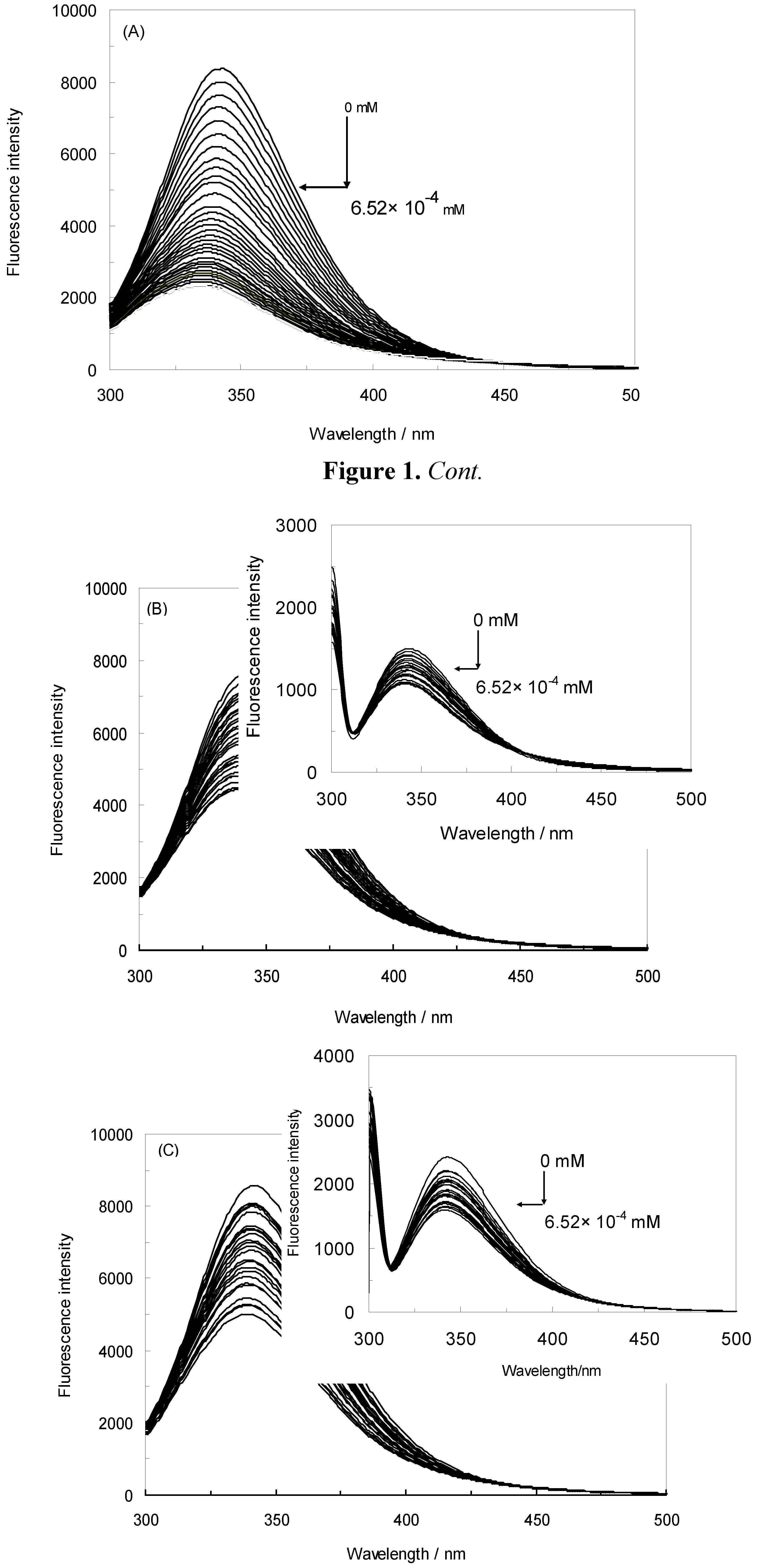

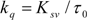
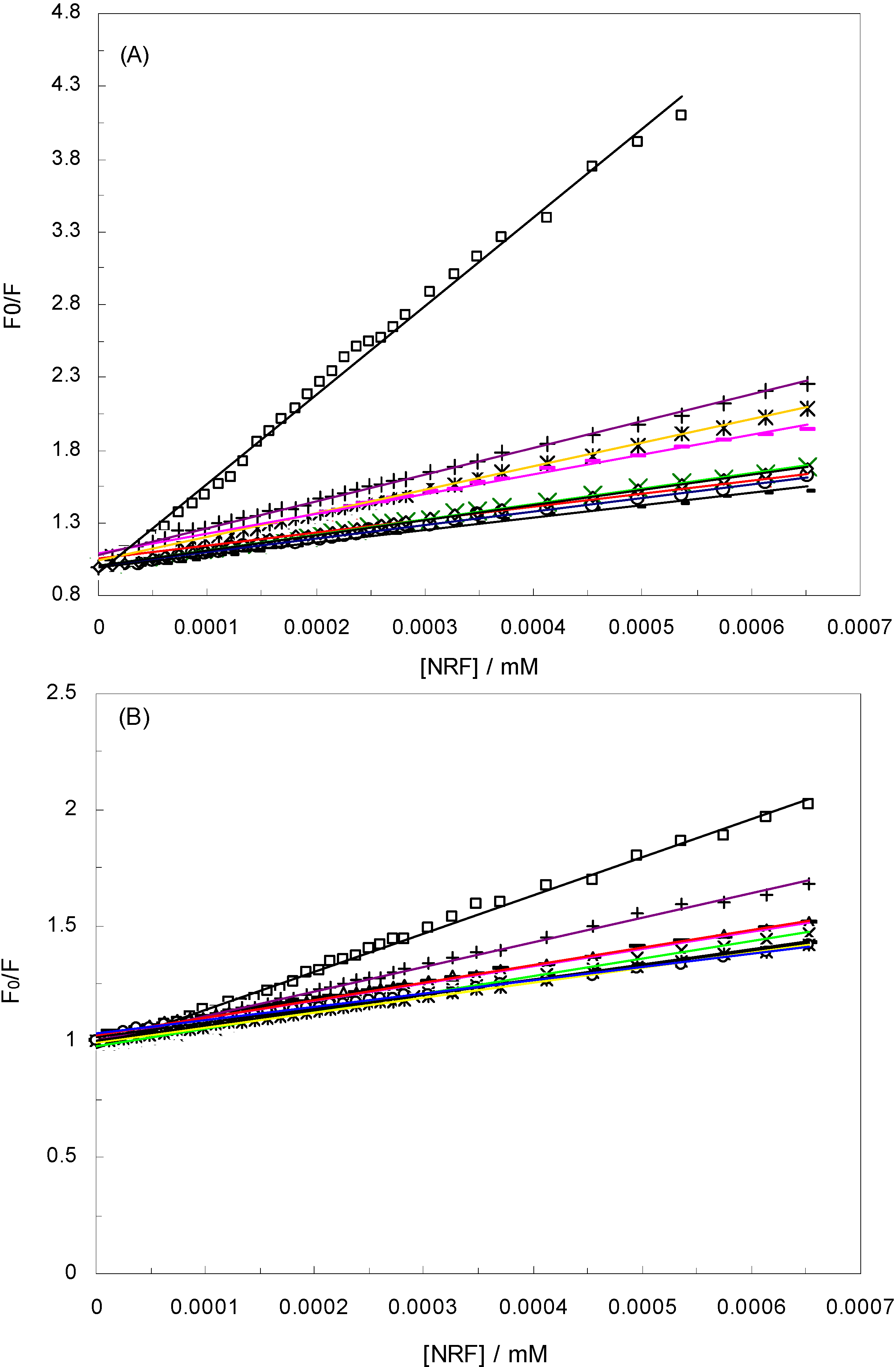
 ), 1 KHz/0.98 mT (
), 1 KHz/0.98 mT (  ), 1 KHz/1.9 mT (
), 1 KHz/1.9 mT (  ), 10 KHz/0.17 mT (-), 10 KHz/63 μT
), 10 KHz/0.17 mT (-), 10 KHz/63 μT 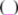 , 100 KHz/39 μT (
, 100 KHz/39 μT (  ), 1 MHz/59 μT (
), 1 MHz/59 μT (  ), (A) at λex = 280 nm, (B) at λex = 295 nm.
), 1 KHz/0.98 mT ( ), 1 KHz/1.9 mT ( ), 10 KHz/0.17 mT (-), 10 KHz/63 μT , 100 KHz/39 μT ( ), 1 MHz/59 μT ( ), (A) at λex = 280 nm, (B) at λex = 295 nm.
), (A) at λex = 280 nm, (B) at λex = 295 nm.
), 1 KHz/0.98 mT ( ), 1 KHz/1.9 mT ( ), 10 KHz/0.17 mT (-), 10 KHz/63 μT , 100 KHz/39 μT ( ), 1 MHz/59 μT ( ), (A) at λex = 280 nm, (B) at λex = 295 nm.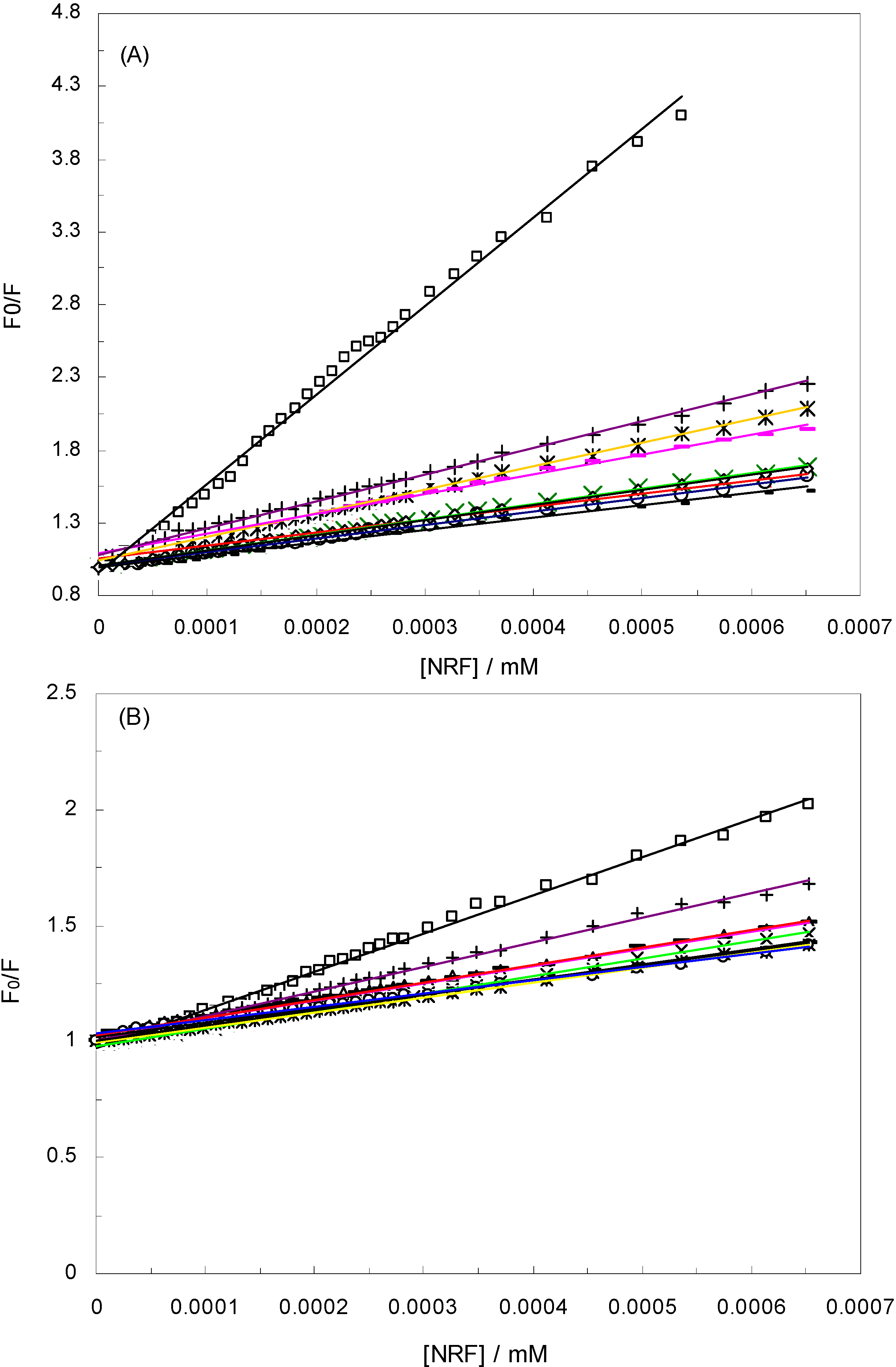

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | Ksv × 10−6/M−1 λex = 280 nm | R λex = 280 nm | Ksv × 10−6/M−1 λex = 295 nm | R λex = 295 nm | fa λex = 295 nm | fa λex = 280 nm |
|---|---|---|---|---|---|---|
| HSA-NRF | 6.12 | 0.9956 | 1.65 | 0.996 | 2.05 | 0.7 |
| HSA-NRF, 100 Hz (2.4 mT) | 1.04 | 0.9978 | 0.63 | 0.9975 | 0.038 | 0.72 |
| HSA-NRF, 100 Hz (6.4 mT) | 0.9 | 0.9914 | 0.74 | 0.9971 | 0.28 | 0.31 |
| HSA-NRF, 1 KHz (0.98 mT) | 1.05 | 0.9984 | 0.76 | 0.9981 | 0.9 | 0.34 |
| HSA-NRF, 1 KHz (1.9 mT) | 1.61 | 0.998 | 0.65 | 0.9995 | 0.37 | 0.26 |
| HAS-NRF, 10KHz (0.17 mT) | 0.86 | 0.9905 | 0.65 | 0.9992 | 0.041 | 0.37 |
| HSA-NRF, 10 KHz (63 μT) | 1.36 | 0.9917 | 0.75 | 0.9977 | 0.37 | 0.21 |
| HSA-NRF, 100 KHz (39 μT) | 0.94 | 0.9963 | 0.58 | 0.9946 | 0.067 | 0.19 |
| HSA-NRF, 1 MHz (59 μT) | 1.84 | 0.9957 | 1.07 | 0.997 | 1.63 | 0.53 |
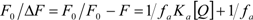
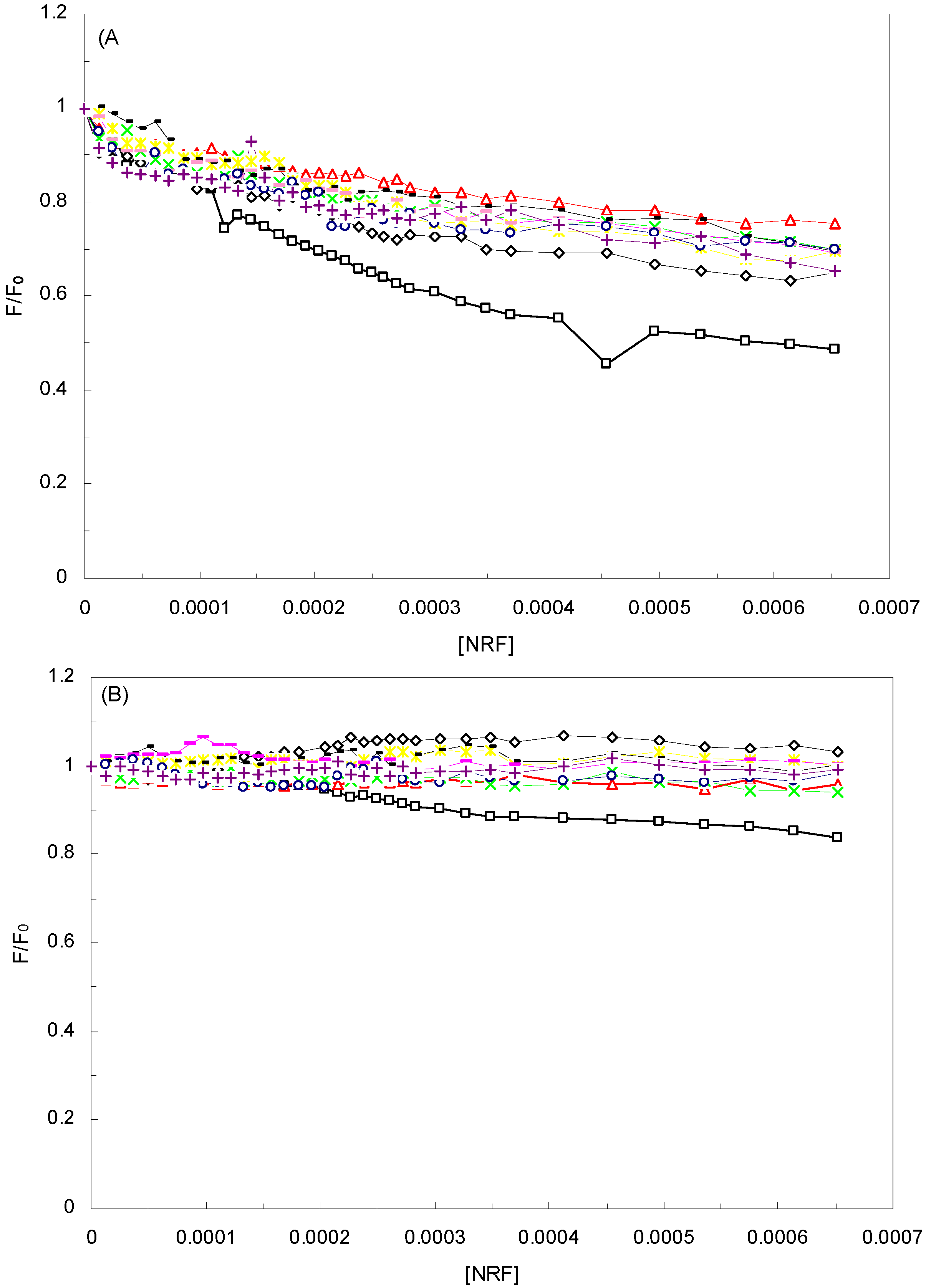
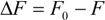 is the difference in fluorescence in the absence and presence of the quencher at concentration [Q], fa is the fraction of accessible fluorescence, and Ka is the effective quenching constant for the accessible fluorophores, which are analogous to associative binding constants for the quencher-acceptor system [26]. The plot of
is the difference in fluorescence in the absence and presence of the quencher at concentration [Q], fa is the fraction of accessible fluorescence, and Ka is the effective quenching constant for the accessible fluorophores, which are analogous to associative binding constants for the quencher-acceptor system [26]. The plot of 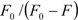 versus 1/[Q] is linear, with (1/fK) as the slope and 1/f as the intercept. The quenching constant K is a quotient of the intercept 1/f and slope (1/fK) [10]. (A plot of F0/∆F versus 1/[Q] yields fa −1 as the intercept and (fa KQ)−1 as the slope. A y– intercept of fa −1 may be understood intuitively).
versus 1/[Q] is linear, with (1/fK) as the slope and 1/f as the intercept. The quenching constant K is a quotient of the intercept 1/f and slope (1/fK) [10]. (A plot of F0/∆F versus 1/[Q] yields fa −1 as the intercept and (fa KQ)−1 as the slope. A y– intercept of fa −1 may be understood intuitively).
 versus log
versus log 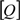 [33]. After analyzing the data with the Hill equation, the values of n and KA could be obtained and they are listed in Table 2.
[33]. After analyzing the data with the Hill equation, the values of n and KA could be obtained and they are listed in Table 2.| System | Ka × 10−6/M−1 λex = 280 nm | R λex = 280 nm | n λex = 280 nm | Ka × 10−6/M−1 λex = 295 nm | R λex = 295 nm | n λex = 295 nm |
|---|---|---|---|---|---|---|
| HSA-NRF | 22.06 | 0.9956 | 1.1589 | 8.252 | 0.996 | 1.2072 |
| HSA-NRF, 100 Hz (2.4 mT) | 6.43 | 0.9978 | 1.2154 | 0.21 | 0.9975 | 0.8561 |
| HSA-NRF, 100 Hz (6.4 mT) | 0.02 | 0.9914 | 0.5402 | 0.054 | 0.9971 | 0.6898 |
| HSA-NRF, 1 KHz (0.98 mT) | 0.66 | 0.9984 | 0.9409 | 0.42 | 0.9981 | 0.9233 |
| HSA-NRF, 1 KHz (1.9 mT) | 0.31 | 0.998 | 0.7889 | 1.307 | 0.9995 | 1.0866 |
| HAS-NRF, 10 KHz (0.17 mT) | 21.42 | 0.9905 | 1.4005 | 0.28 | 0.9992 | 0.8946 |
| HSA-NRF, 10 KHz (63 μT) | 0.11 | 0.9917 | 0.6698 | 0.086 | 0.9977 | 0.7185 |
| HSA-NRF, 100 KHz (39 μT) | 5.116 | 0.9963 | 1.2033 | 0.053 | 0.9946 | 0.6838 |
| HSA-NRF, 1 MHz (59 μT) | 0.4 | 0.9957 | 0.7908 | 0.033 | 0.997 | 0.5263 |
2.2. Conformation Investigation by Synchronous Fluorescence
 ), 1 KHz/0.98 mT( ), 1 KHz/1.9 mT( ), 10 KHz/0.17 mT (-), 10 KHz/63 μT , 100 KHz/39 μT ( ), 1 MHz/59 μT ( ), (A) in Δλ = 60. (B) Δλ = 15.
), 1 KHz/0.98 mT( ), 1 KHz/1.9 mT( ), 10 KHz/0.17 mT (-), 10 KHz/63 μT , 100 KHz/39 μT ( ), 1 MHz/59 μT ( ), (A) in Δλ = 60. (B) Δλ = 15.
), 1 KHz/0.98 mT( ), 1 KHz/1.9 mT( ), 10 KHz/0.17 mT (-), 10 KHz/63 μT , 100 KHz/39 μT ( ), 1 MHz/59 μT ( ), (A) in Δλ = 60. (B) Δλ = 15.
), 1 KHz/0.98 mT( ), 1 KHz/1.9 mT( ), 10 KHz/0.17 mT (-), 10 KHz/63 μT , 100 KHz/39 μT ( ), 1 MHz/59 μT ( ), (A) in Δλ = 60. (B) Δλ = 15.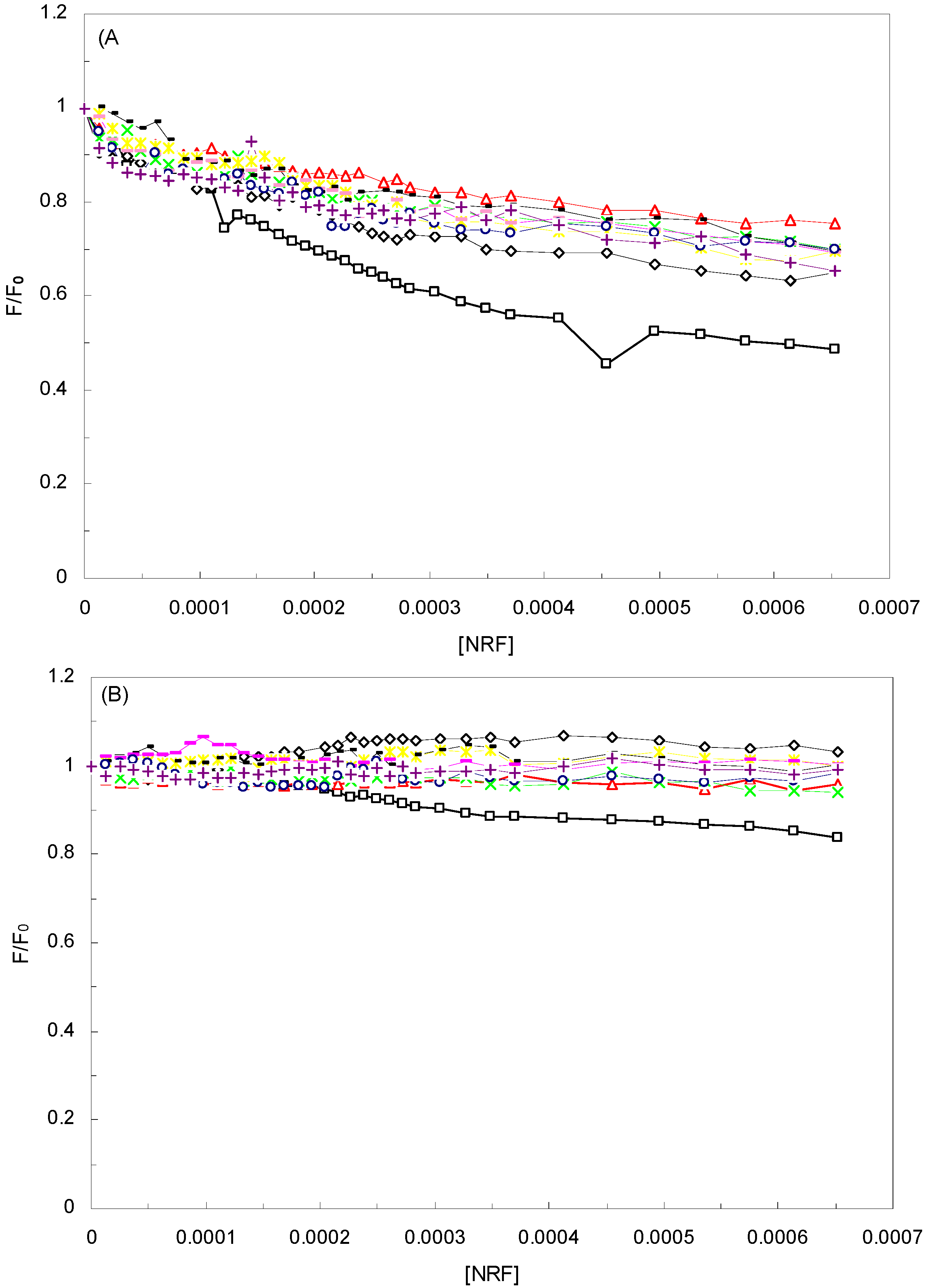
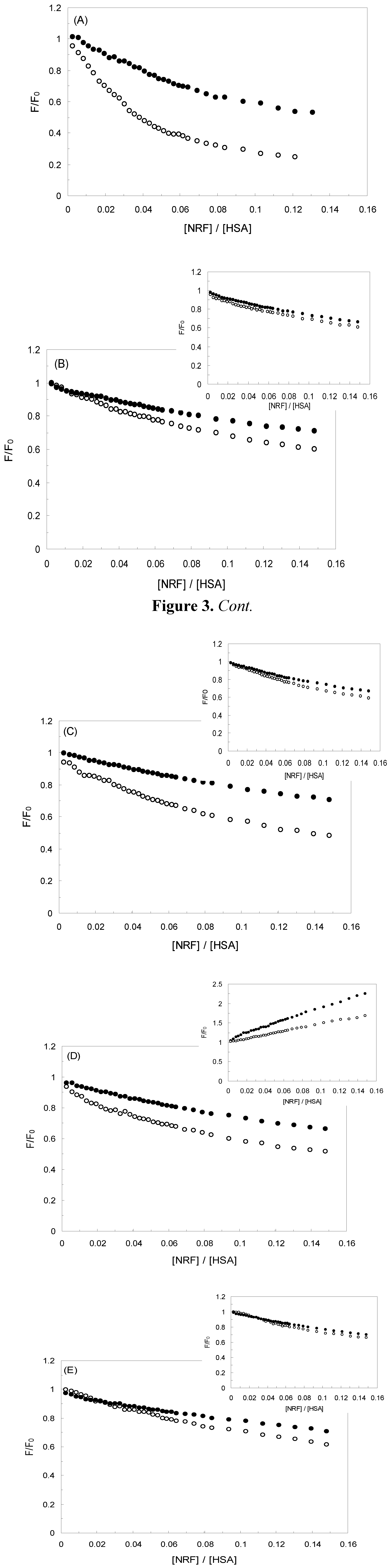
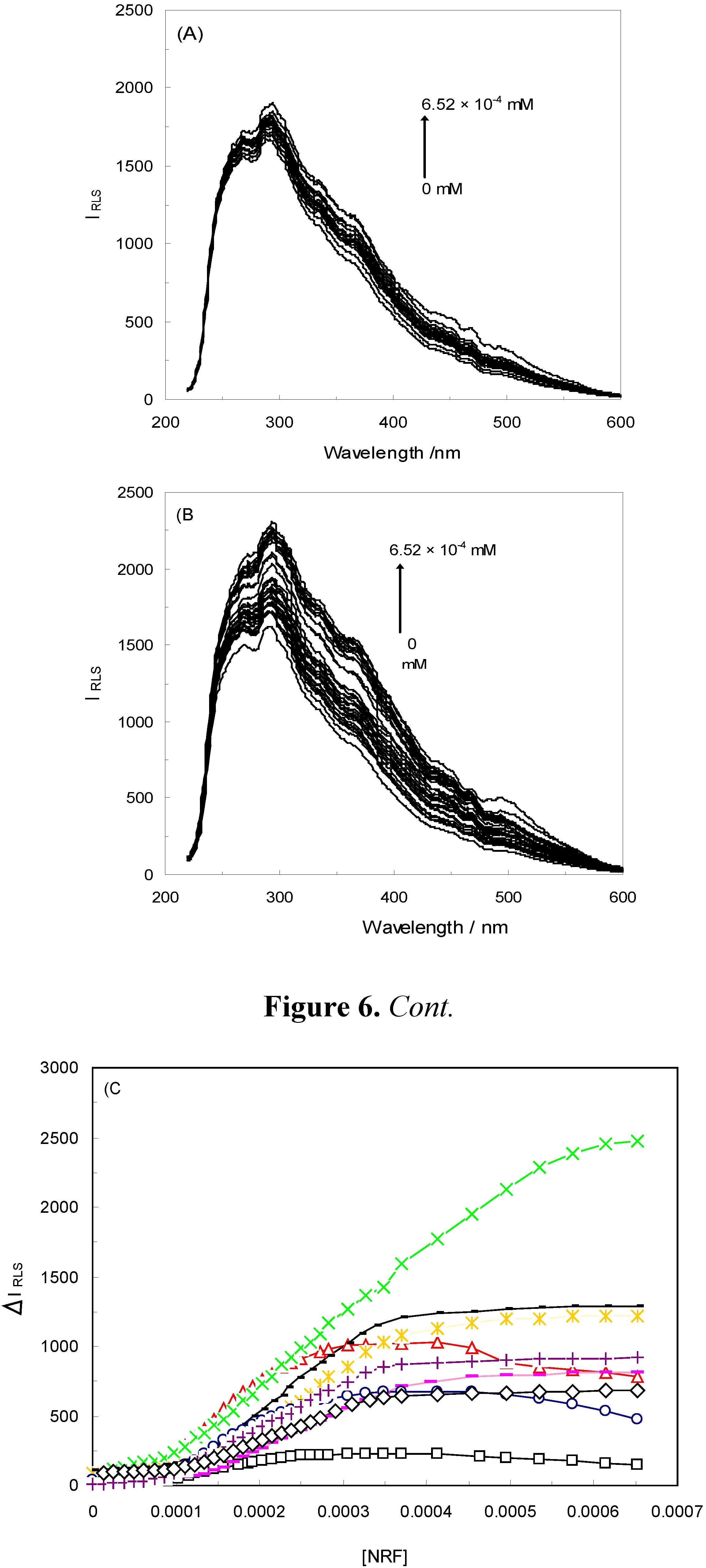
2.3. Resonance Light Scattering
), 1 KHz/0.98 mT( ), 1 KHz/1.9 mT( ), 10 KHz/0.17 mT(-), 10 KHz/63 μT , 100 KHz/39 μT 1 MHz/59 μT ( ).
), 1 KHz/0.98 mT( ), 1 KHz/1.9 mT( ), 10 KHz/0.17 mT(-), 10 KHz/63 μT , 100 KHz/39 μT 1 MHz/59 μT ( ).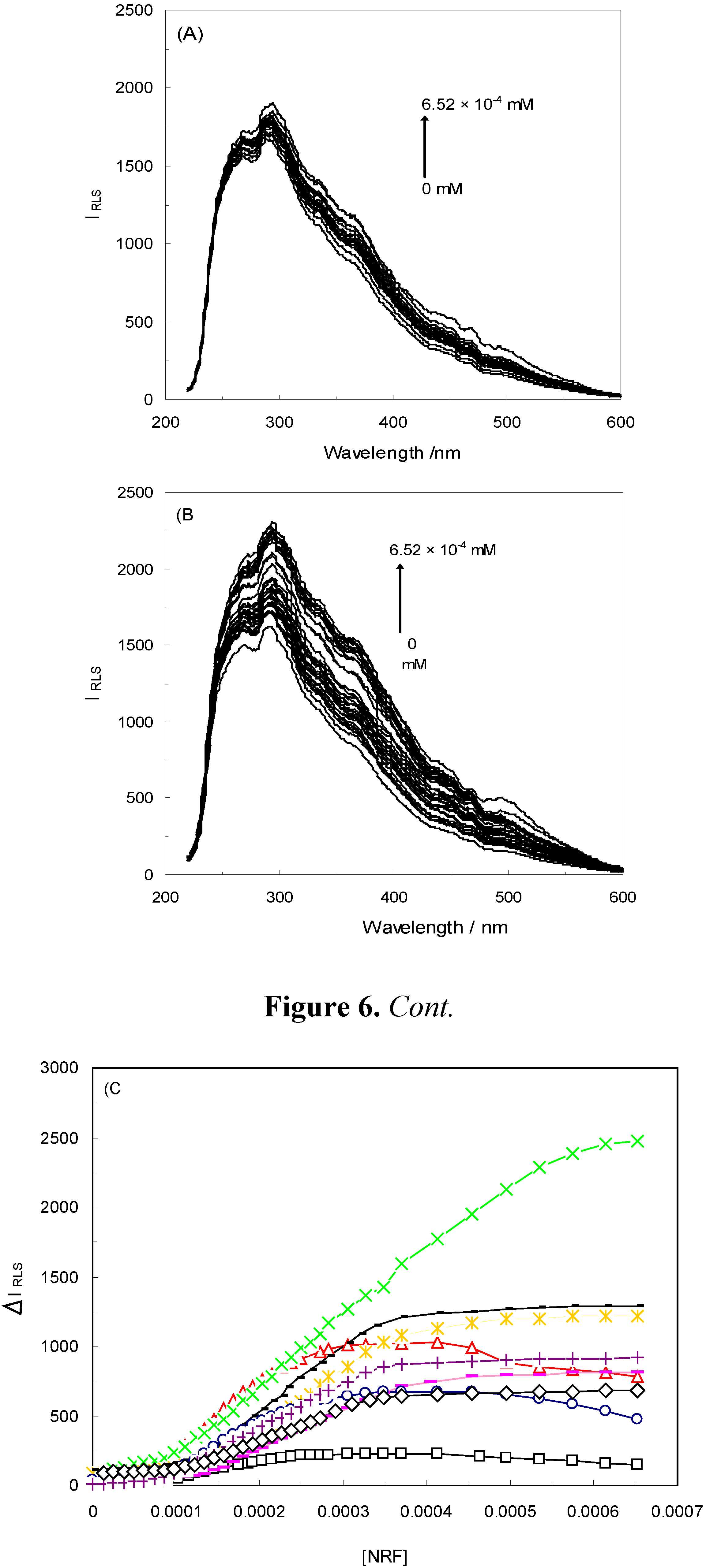
2.4. Red Edge Excitation Shift (Theory and Application)
| System | NRF-HSA / 1:10 |
|---|---|
| HSA-NRF | 7 |
| HSA-NRF, 100 Hz (2.4 mT) | 2 |
| HSA-NRF, 100 Hz (6.4 mT) | 2 |
| HSA-NRF,1 KHz (0.98 mT) | 2 |
| HSA-NRF, 1 KHz (1.9 mT) | 4 |
| HSA-NRF, 10 KHz (0.17 mT) | 4 |
| HSA-NRF, 10 KHz (63 μT) | 2 |
| HSA-NRF, 100 KHz (39 μT) | 2 |
| HSA-NRF, 1 MHz (59 μT) | 4 |
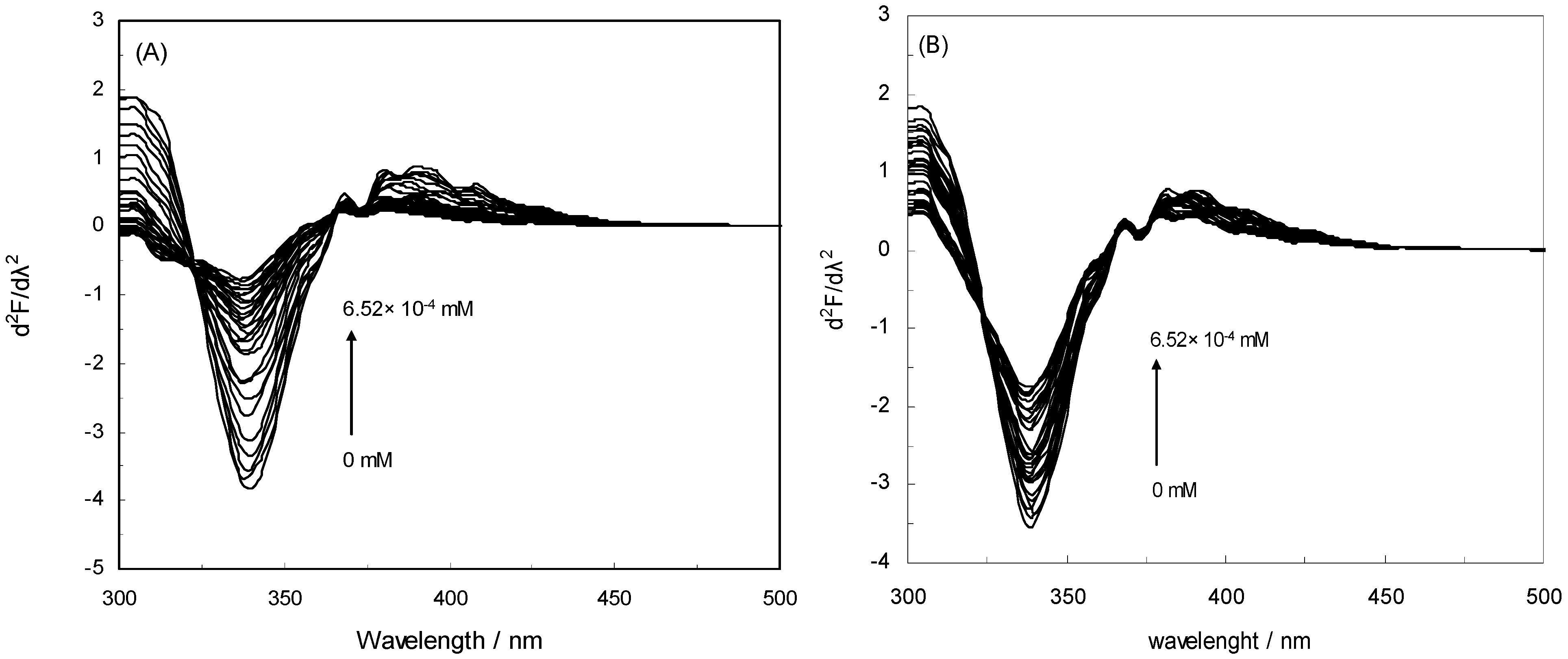
2.5. Second Derivative Fluorescence Spectroscopy
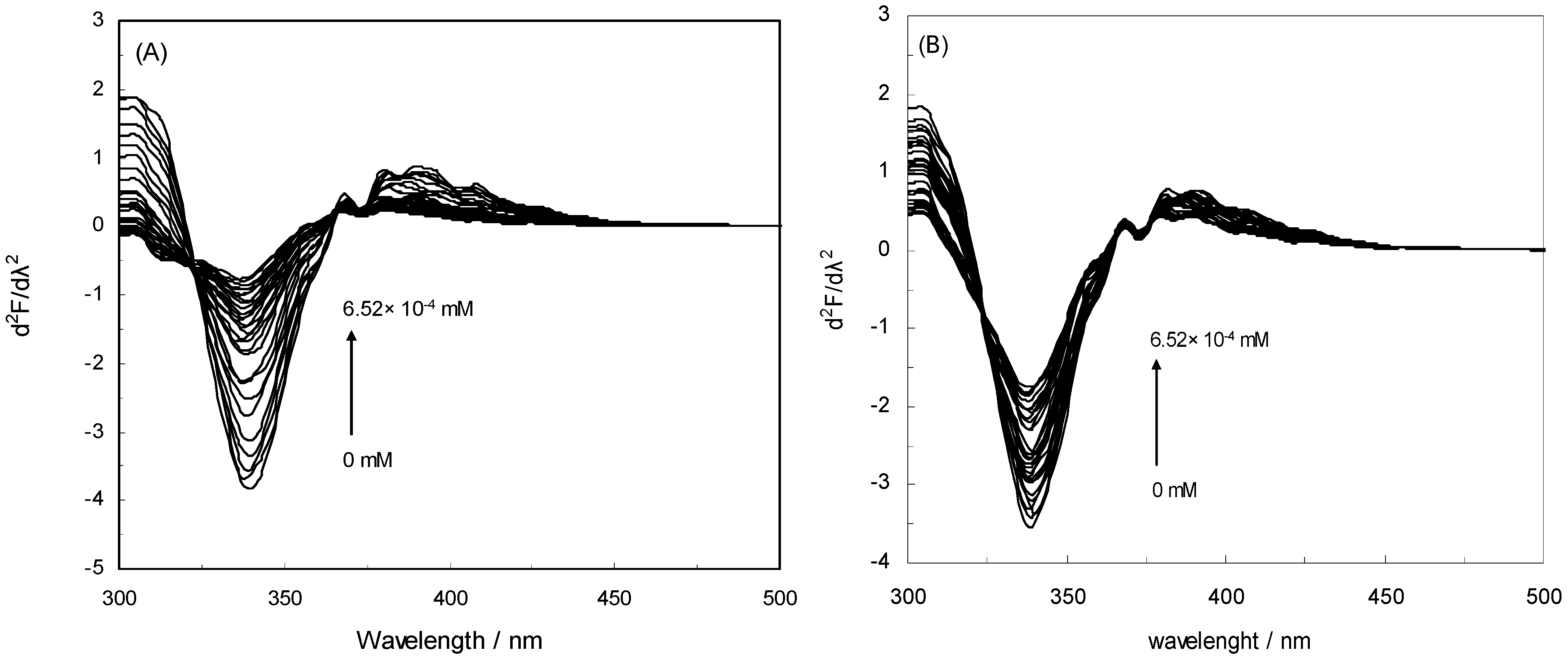
2.6. Energy Transfer from HSA to Drugs
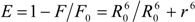

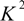 is the spatial orientation factor of the dipole, n is the refractive index of the medium,
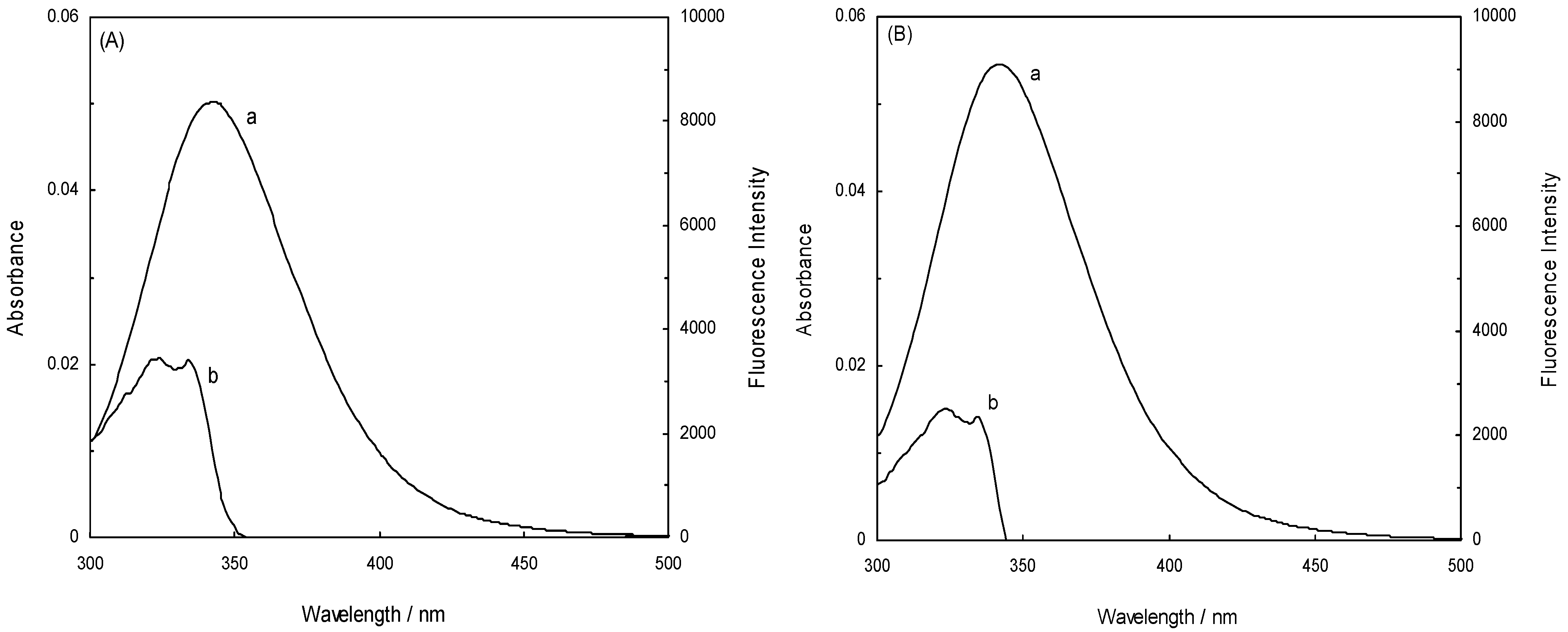
is the spatial orientation factor of the dipole, n is the refractive index of the medium, 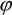 is the fluorescence quantum yield of the donor, n is the average refractive index of the medium, J is the overlap integral of the fluorescence emission spectrum of the donor and the absorption spectrum of the acceptor, (Figure 8). J is given by:
is the fluorescence quantum yield of the donor, n is the average refractive index of the medium, J is the overlap integral of the fluorescence emission spectrum of the donor and the absorption spectrum of the acceptor, (Figure 8). J is given by:

 (λ) is the molar absorption coefficient of the acceptor at wavelength λ [11,31].
(λ) is the molar absorption coefficient of the acceptor at wavelength λ [11,31].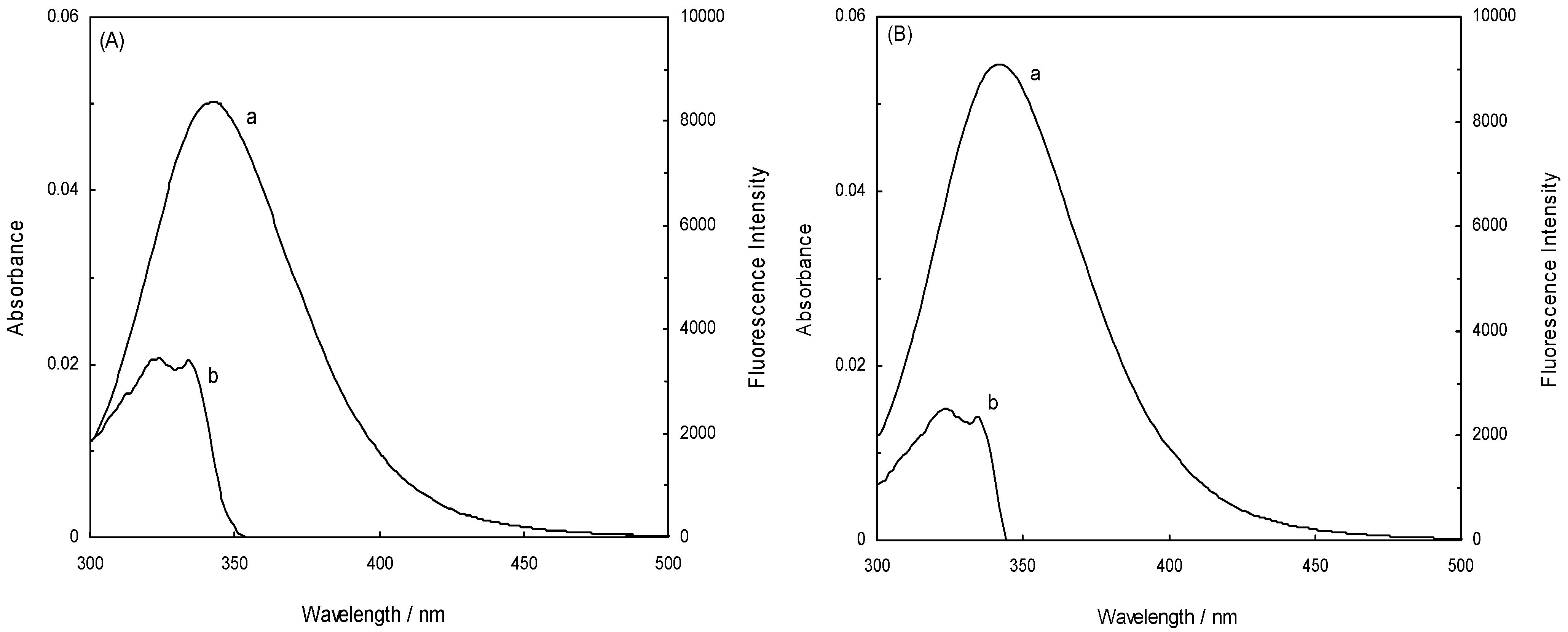
| System | H /1:10 | r/ nm |
|---|---|---|
| HSA-NRF | 7 | 1.98 |
| HSA-NRF, 100Hz (2.4 mT) | 2 | 2.03 |
| HSA-NRF, 100Hz (6.4 mT) | 2 | 2.08 |
| HSA-NRF, 1KHz (0.98 mT) | 2 | 2.27 |
| HSA-NRF, 1KHz (1.9 mT) | 4 | 2.33 |
| HAS-NRF, 10KHz (0.17 mT) | 4 | 2.51 |
| HSA-NRF, 10KHz (63 μT) | 2 | 2.57 |
| HSA-NRF, 100KHz (39 μT) | 2 | 2.62 |
| HSA-NRF, 1MHz (59 μT) | 4 | 3.11 |
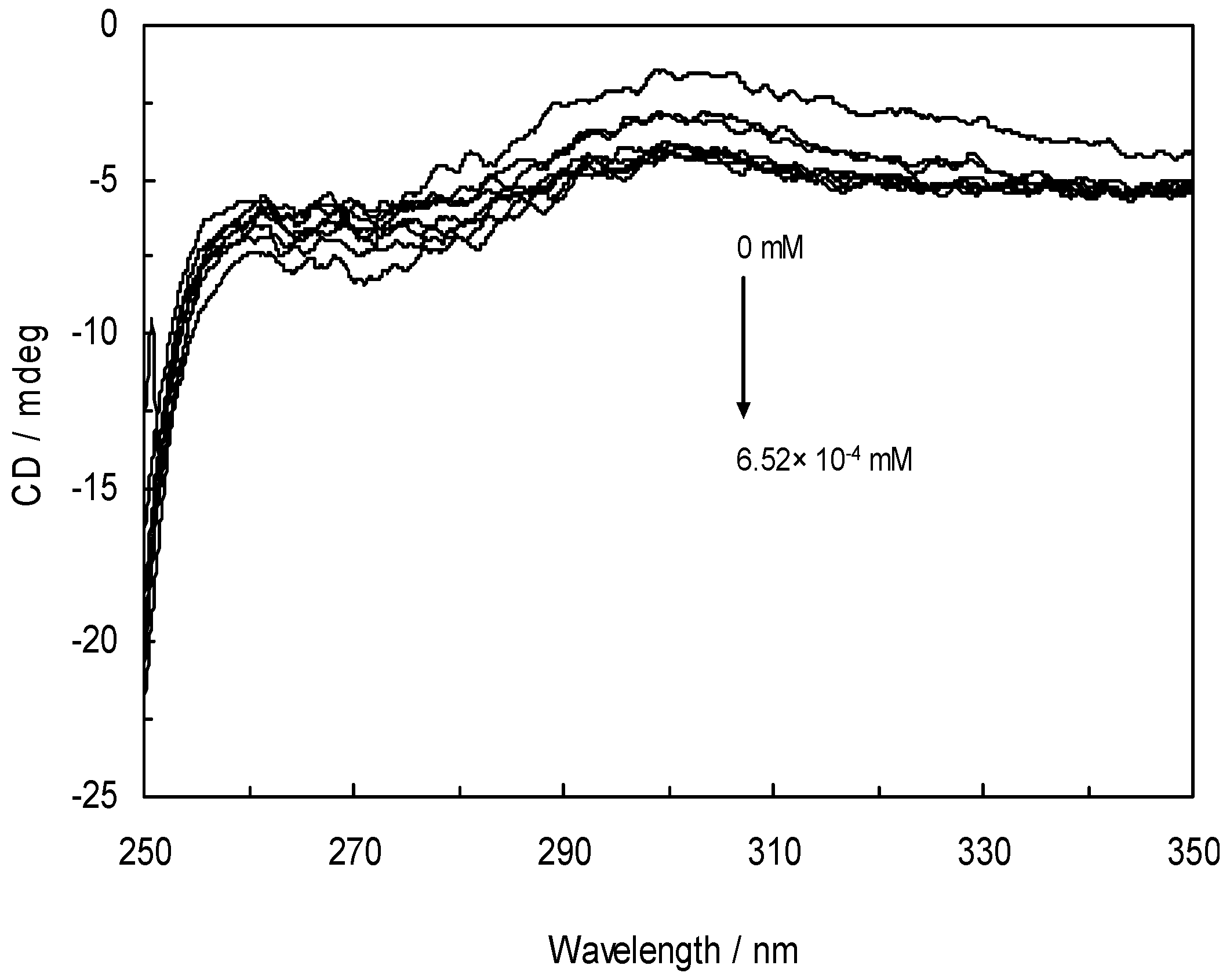
3.7. Circular Dichroism and the Conformational Analysis
| System | α-helix % | β-sheet % | Turn % | Unordered coil % |
|---|---|---|---|---|
| HSA | 53.97 | 18.31 | 13.48 | 14.25 |
| HSA-NRF | 53.17 | 17.78 | 13.22 | 15.83 |
| HSA-NRF, 100 Hz (2.4 mT) | 53.06 | 17.61 | 13.17 | 16.16 |
| HSA-NRF, 100 Hz (6.4 mT) | 53.01 | 17.57 | 13.13 | 16.29 |
| HSA-NRF, 1 KHz (0.98 mT) | 52.93 | 17.44 | 13.03 | 16.6 |
| HSA-NRF, 1 KHz (1.9 mT) | 52.88 | 17.37 | 12.81 | 16.94 |
| HAS-NRF,10 KHz (0.17 mT) | 52.78 | 17.31 | 12.75 | 17.16 |
| HSA-NRF, 10 KHz (63 μT) | 52.76 | 17.31 | 12.72 | 17.21 |
| HSA-NRF, 100 KHz (39 μT) | 52.43 | 17.11 | 12.41 | 18.05 |
| HSA-NRF, 1 MHz (59 μT) | 51.77 | 16.59 | 12.27 | 19.37 |
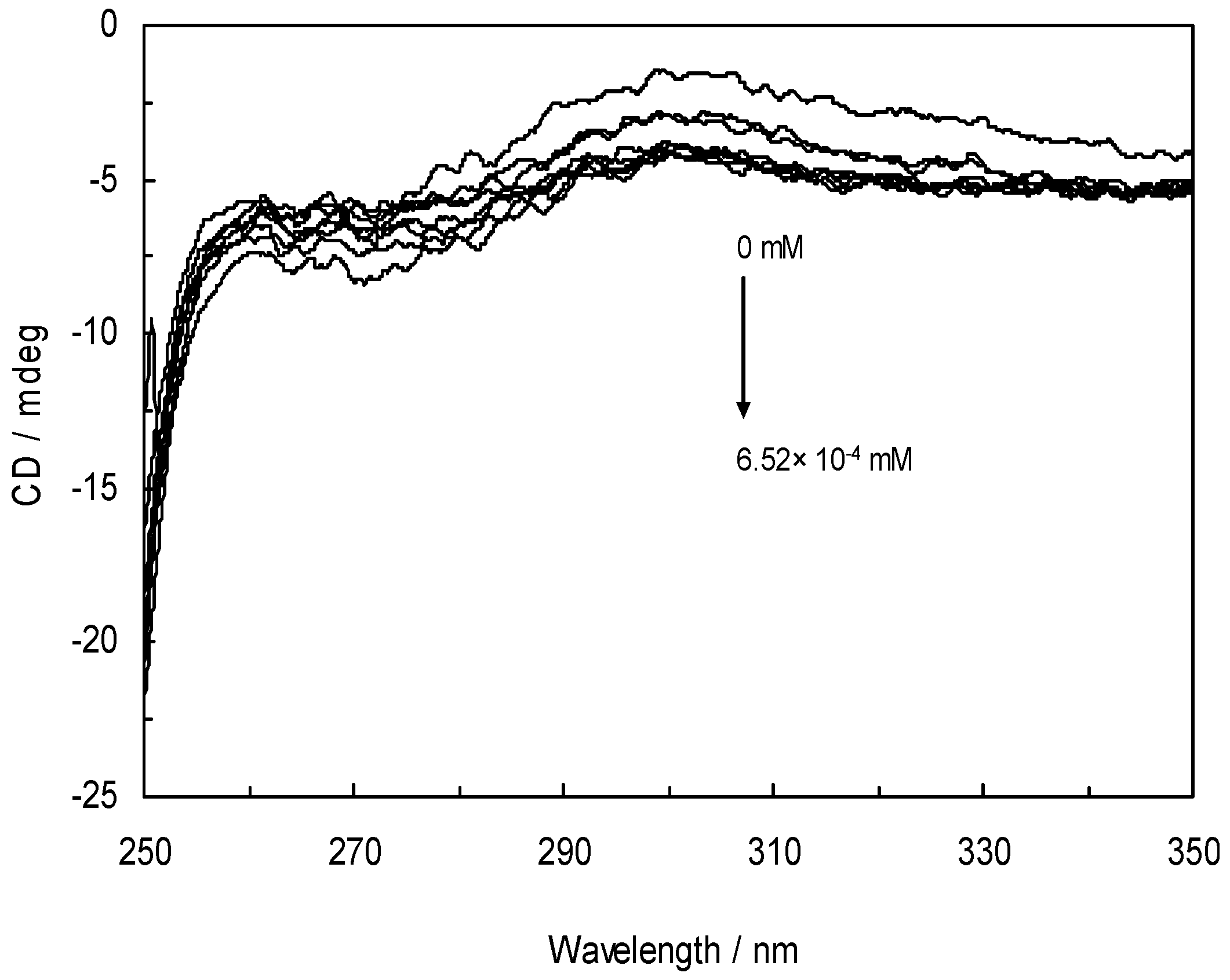
3. Experimental
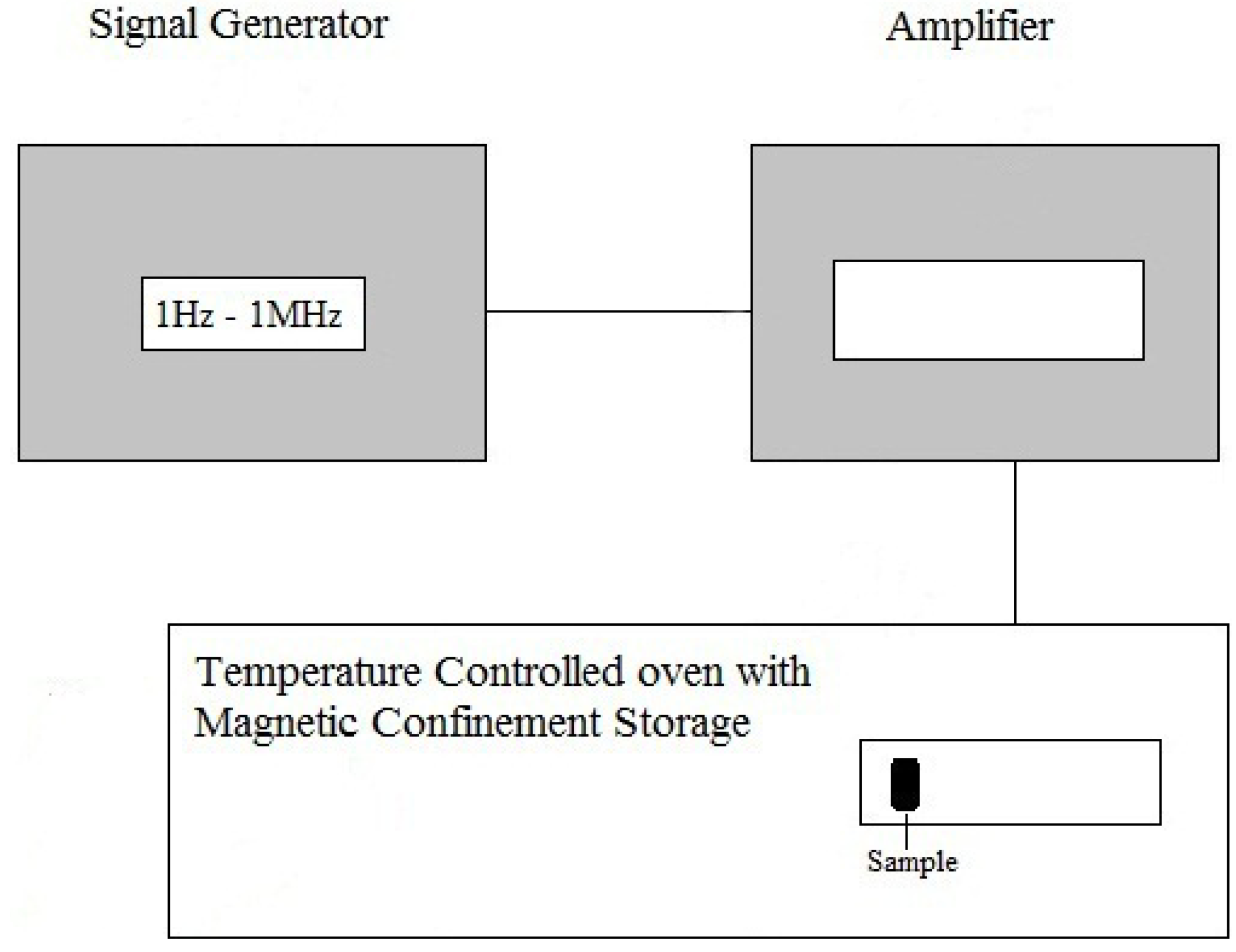
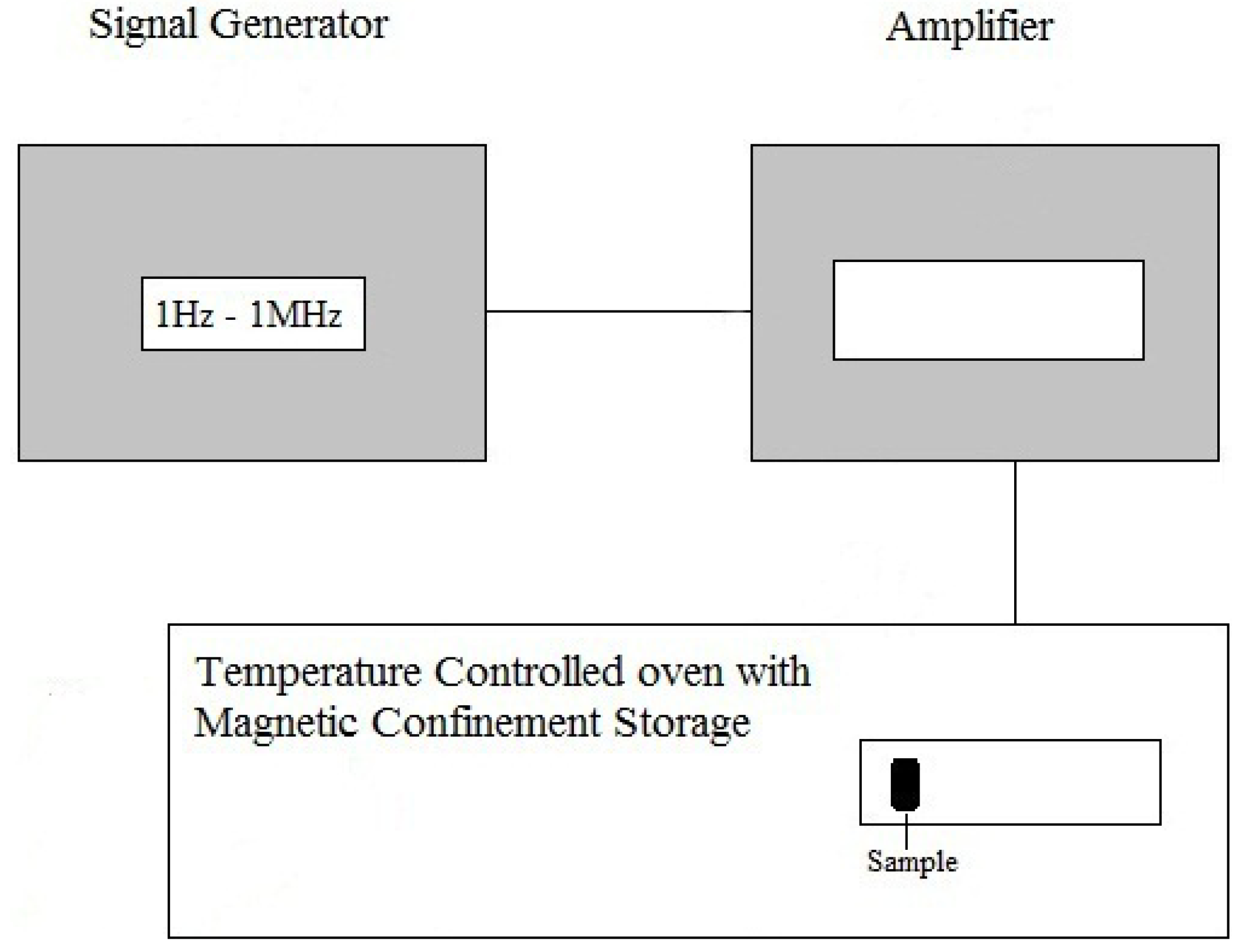
3.1. Apparatus
3.2. Reagents
3.3. Procedures
4. Conclusions
Acknowledgements
References and Notes
- Laurence, J.A.; French, P.W.; Lindner, R.A.; McKenzie, D.R. Biological effects of electromagnetic fields-mechanisms for the effects of pulsed microwave radiation on protein conformation. J. Theor. Biol. 2000, 206, 291–298. [Google Scholar] [CrossRef]
- Blank, M. Electromagn, Protein and DNA reactions stimulated by electromagnetic fields. Biol. Med. 2008, 27, 3–23. [Google Scholar]
- Mousavy, S.J.; Hossein Riazi, G.H.; Kamarei, M.; Aliakbarian, H.; Sattarahmady, N.; Sharifizadeh, A.; Safarianc, S.; Ahmad, F.; Moosavi-Movahedi, A.A. Effects of mobile phone radiofrequency on the structure and function of the normal human hemoglobin. Int. J. Biol. Macromol. 2009, 44, 278–285. [Google Scholar] [CrossRef]
- Blank, M. Do electromagnetic fields interact with electrons in the Na,K-ATPase? Bioelectromagnetics 2005, 26, 677–683. [Google Scholar] [CrossRef]
- Blank, M.; Goodman, R. Do electromagnetic fields interact directly with DNA? Bioelectromagnetics 1997, 18, 111–115. [Google Scholar] [CrossRef]
- Blank, M.; Soo, L. Enhancement of cytochrome oxidase activity in 60 Hz magnetic Fields. Bioelectrochem. Bioenerg. 1998, 45, 253–259. [Google Scholar] [CrossRef]
- Bohr, H.; Bohr, J. Microwave-enhanced folding and denaturation of globular proteins. Phys. Rev. E 2000, 61, 4310–4314. [Google Scholar] [CrossRef]
- Song, S.M.; Hou, X.L.; Wu, Y.-B.; Shuang, S.-M.; Yang, C.; Inoue, Y.; Dong, C. Study on the interaction between methyl blue and human serum albumin by fluorescence spectroscopy. J. Lumin. 2009, 129, 169–175. [Google Scholar] [CrossRef]
- Ghuman, J.; Zunszain, P.A.; Petitpas, I.; Bhattacharya, A.A.; Otagiri, M.; Curry, S. Structural basis of the drug-binding specificity of human serum albumin. J. Mol. Biol. 2005, 353, 38–52. [Google Scholar] [CrossRef]
- Tang, J.; Luan, F.; Chen, X. Binding analysis of glycyrrhetinic acid to human serumalbumin: Fluorescence spectroscopy, FTIR, and molecular modeling. Bioorg. Med. Chem. 2006, 14, 3210–3217. [Google Scholar] [CrossRef]
- Zhang, G.; Que, Q.; Pan, J.; Guo, J. Study of the interaction between icariin and human serum albuminby fluorescence spectroscopy. J. Mol. Struct. 2008, 881, 132–138. [Google Scholar] [CrossRef]
- Froehlich, E.; Mandeville, J.S.; Jennings, C.J.; Sedaghat-Herati, R.; Tajmir-Riahi, H.A. Dendrimers bind human serum albumin. J. Phys. Chem. B 2009, 113, 6986–6993. [Google Scholar]
- Mandeville, J.S.; Froehlich, E.; Tajmir-Riahi, H.A. Study of curcumin and genistein interactions with human serum albumin. J. Pharm. Biomed. Anal. 2009, 49, 468–474. [Google Scholar] [CrossRef]
- Mandeville, J.S.; Tajmir-Riahi, H.A. Complexes of dendrimers with bovine serum albumin. Biomacromolecules 2010, 11, 465–472. [Google Scholar] [CrossRef]
- Bourassa, P.; Dubeau, S.; Maharvi, G.M.; Fauq, A.H.; Thomas, T.J.; Tajmir-Riahi, H.A. Binding of anti-tumor tamoxifen and its metabolites 4-hydroxy tamoxifen and endoxifen to human serum albumin. Biochimie 2011, 93, 1089–1101. [Google Scholar] [CrossRef]
- Petitpas, I.; Grüne, T.; Bhattacharya, A.A.; Curry, S. Crystal structures of human serum albumin complexed with monounsaturated and polyunsaturated fatty acids. J. Mol. Biol. 2001, 314, 955–960. [Google Scholar] [CrossRef]
- Atmeh, R.F.; Arafa, I.M.; Al-Khateeb, M. Albumin Aggregates: Hydrodynamic Shape and Physico-Chemical Properties. Jordan J. Chem. 2007, 2, 169–182. [Google Scholar]
- Peters, T.Jr. All about Albumin, 1st ed.; Academic Press: New York, NY, USA, 1996; pp. 1–5. [Google Scholar]
- Yadav, J.; Joshi, Y.C. Synthesis and Spectral Study of Novel Norfloxacin Derivatives. Eur.-J. Chem. 2008, 5, 1154–1158. [Google Scholar]
- Zhang, L.W.; Wang, K.; Zhang, X.X. Study of the interactions between fluoroquinolones and human serum albumin by affinity capillary electrophoresis and fluorescence method. Anal. Chim. Acta 2007, 603, 101–110. [Google Scholar] [CrossRef]
- Dua, K.; Ramana, M.V.; Singh Sara, U.V.; Himaja, M.; Agrawal, A.; Garg, V.; Pabreja, K. Bentham Science Publishers Ltd. Investigation of Enhancement of Solubility of Norfloxacin β-Cyclodextrin. Curr. Drug Deliv. 2007, 4, 21–25. [Google Scholar] [CrossRef]
- Kuchekar, B.S.; Pattan, S.R.; Godge, R.K.; Laware, R.B.; Nirmal, S.A.; Parjane, S.K.; Merekar, A.N. Formulation and evaluation of Norfloxacin Dispersible tablets using Natural substances as Disintegrants. Chem. Pharm. Res. 2009, 1, 336–341. [Google Scholar]
- Sher, M.; Hussain, M.A.; Mehmood, M.H.; Hassan, M.; Bashir, N.S. Bioequivalence of norfloxacin by hplc-uv method. J. Chil. Chem. Soc. 2010, 55, 203–205. [Google Scholar]
- Harding, S.E.; Chowdhry, B.Z. The Protein-ligand Interactions: Hydrodynamics and Calorimetry; Oxford University Press: New York, NY, USA, 2001. [Google Scholar]
- Garland, C.W.; Nibler, J.W.; Shoemaker, D.P. Experiments in Physical Chemistry, 7th ed.; McGraw-Hill: Boston, MA, USA, 2003. [Google Scholar]
- Wang, Y.; Hu, Y.J.; Yang, Y.O.; Zhou, J.; Liu, Y. Characterize the interaction between naringenin and bovine serum albumin using spectroscopic approach. J. Lumin. 2010, 130, 1394–1399. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Zhang, H.M.; Zhou, Q.H. Studies on the interaction of caffeine with bovine hemoglobin. Med. Chem. 2009, 44, 2100–2105. [Google Scholar]
- Sulkowska, A.; Maciazek-Jurczyk, M.; Bojko, B.; Rownicka, J.; Zubik-skupien, I.; Temba, E.; Pentak, D.; Sulkowska, W. Competitive binding of phenylbutazone and colchicines to serum albumin in multidrug therapy: A spectroscopic study. J. Mol. Struct. 2008, 881, 97–106. [Google Scholar] [CrossRef]
- Ding, F.; Huang, J.; Lin, J.; Li, Z.Y.; Liu, F.; Jiang, Z.Q.; Sun, Y. A study of the binding of C.I. Mordant Red 3 with bovine serum albumin using fluorescence spectroscopy. Dyes Pigm. 2009, 82, 1–6. [Google Scholar] [CrossRef]
- Hamed-Akbari Tousi, S.; Saberi, M.R.; Chamani, J. Comparing the Interaction of Cyclophosphamide Monohydrate to Human Serum Albumin as Opposed to Holo-Transferrin by Spectroscopic and Molecular Modeling Methods: Evidence for Allocating the Binding Site. Protein Pept. Lett. 2010, 17, 1524–1535. [Google Scholar]
- Chamani, J.; Tafrishi, N.; Momen-Heravi, M. Characterization of the interaction between human lactoferrin and lomefloxacin at physiological condition: Multi-spectroscopic and modeling description. J. Lumin. 2010, 130, 1160–1168. [Google Scholar] [CrossRef]
- Wei, X.L.; Xiao, J.B.; Wang, Y.; Bai, Y. Which model based on fluorescence quenching is suitable to study the interaction between trans-resveratrol and BSA? Spectrochim. Acta A Mol. Biomol. Spectrosc. 2010, 75, 299–304. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 2nd ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 1999. [Google Scholar]
- Lloyd, J.B.F. Synchronized excitation of fluorescence emission spectra. Nature 1971, 23, 64–66. [Google Scholar]
- Yuan, T.; Weljie, A.M.; Vogel, H.J. Tryptophan fluorescence quenching by methionine and selenomethionine residues of calmodulin: Orientation of peptide and protein binding. Biochemistry 1998, 37, 3187–3195. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Zhang, H.M.; Zhang, G.C.; Zhou, Q.H.; Fei, Z.H.; Liu, Z.T.; Li, Z.X. Fluorescence spectroscopic investigation of the interaction between benzidine and bovine hemoglobin. J. Mol. Struct. 2008, 886, 77–84. [Google Scholar] [CrossRef]
- Yuan, J.L.; Lv, Z.; Liu, Z.G.; Hu, Z.; Zou, G.L. Study on interaction between apigenin and human serum albumin by spectroscopy and molecular modeling. J. Photochem. Photobiol. A Chem. 2007, 191, 104–113. [Google Scholar] [CrossRef]
- Yue, Y.; Chen, X.; Qin, J.; Yao, X. A study of the binding of C.I. Direct Yellow 9 to human serum albumin using optical spectroscopy and molecular modeling. Dyes Pigm. 2008, 79, 176–182. [Google Scholar] [CrossRef]
- Wu, L.; Mu, D.; Gao, D.; Deng, X.; Tian, Y.; Zhang, H.; Yu, A. Determination of protein by resonance light scattering technique usingdithiothreitol–sodium dodecylbenzene sulphonate as probe. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2009, 72, 178–181. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, J.; Han, Y. Rapid and sensitive determination of proteins by enhanced resonance light scattering spectroscopy of sodium lauroyl glutamate. Talanta 2007, 71, 1246–1251. [Google Scholar] [CrossRef]
- Gao, F.; Luo, F.; Tang, L.; Dai, L.; Wang, L. Preparation of a novel fluorescence probe of terbium–europium co-luminescence composite nanoparticles and its application in the determination of proteins. J. Lumin. 2008, 128, 462–468. [Google Scholar] [CrossRef]
- Huang, C.Z.; Li, Y.F. Resonance light scattering technique used for biochemical and pharmaceutical analysis. Anal. Chim. Acta 2003, 500, 105–117. [Google Scholar] [CrossRef]
- Lu, W.; Shang, S. A resonance light-scattering (RLS) serving for various quantitative events since 1995: a comment proposed towards how to apprehend well the meaning of RLS and its corresponding guiding role. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2009, 74, 285–291. [Google Scholar] [CrossRef]
- Huang, C.Z.; Li, Y.F.; Liu, X.D. Determination of nucleic acids at nanogram levels with safranine T by a resonance light-scattering technique. Anal. Chim. Acta 1998, 375, 89–97. [Google Scholar] [CrossRef]
- Long, X.; Zhang, C.; Cheng, J.; Bi, S. A novel method for study of the aggregation of protein induced by metal ion aluminum(III) using resonance Rayleigh scattering technique. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2008, 69, 71–77. [Google Scholar] [CrossRef]
- Chattopadhyay, A.; Mukherjee, S. Red Edge Excitation Shift of a Deeply Embedded Membrane Probe: Implications in Water Penetration in the Bilayer. J. Phys. Chem. 1999, 103, 8180–8185. [Google Scholar] [CrossRef]
- Lakowicz, J.R.; Keating-Nakamoto, S. Red-edge excitation of fluorescence and dynamic properties of proteins and membranes. Biochemistry 1984, 23, 3013–3021. [Google Scholar] [CrossRef]
- Guha, S.; Rawat, S.S.; Chattopadhyay, A.; Bhattacharyya, B. Tubulin Conformation and Dynamics: A Red Edge Excitation Shift Study. Biochemistry 1996, 35, 13426–13433. [Google Scholar] [CrossRef]
- Joshi, H.C.; Upadhyay, A.; Mishra, H.; Tripathi, H.B.; Pant, D.D. Edge excitation red shift and micro environmental effects on thephotophysics of quinine bisulphate dication. Photochem. Photobiol. A Chem. 1999, 122, 185–189. [Google Scholar] [CrossRef]
- Mozo-Villarıas, A. Second derivative fluorescence spectroscopy of tryptophan in proteins. J. Biochem. Biophys. Methods 2002, 50, 163–178. [Google Scholar] [CrossRef]
- Sarzehi, S.; Chamani, J. Investigation on the interaction between tamoxifen and human holo-transferrin: Determination of the binding mechanism by fluorescence quenching, resonance light scattering and circular dichroism methods. Int. J. Biol. Macromol. 2010, 47, 558–569. [Google Scholar] [CrossRef]
- Omidvar, Z.; Parivar, K.; Sanee, H.; Amiri-Tehranizadeh, Z.; Baratian, A.; Saberi, M.R.; Asoodeh, A.; Chamani, J. Investigations with Spectroscopy, Zeta Potential and Molecular Modeling of the Non-Cooperative Behaviour Between Cyclophosphamide Hydrochloride and Aspirin upon Interaction with Human Serum Albumin: Binary and Ternary Systems from the View Point of Multi-Drug Therapy. J. Biomol. Struct. Dyn. 2011, 29, 181–206. [Google Scholar] [CrossRef]
- Whitmore, L.; Wallace, B.A. Protein Secondary Structure Analyses from Circular Dichroism. Biopolymers 2007, 89, 392–400. [Google Scholar] [CrossRef]
- Tetin, S.Y.; Prendergast, F.G.; Yu, S. Accuracy of protein secondary structure determination from circular dichroism spectra based on immunoglobulin examples. Anal. Biochem. 2003, 321, 183–187. [Google Scholar]
- Zhu, Y.; Cheng, G.; Dong, S. The electrochemically induced conformational transition of disulfides in bovine serum albumin studied by thin layer circular dichroism spectroelectrochemistry. Biophys. Chem. 2001, 90, 1–8. [Google Scholar] [CrossRef]
- Kelly, S.M.; Jess, T.J.; Price, N.C. How to study proteins by circular dichroism. Biochim. Biophys. Acta 2005, 1751, 119–139. [Google Scholar] [CrossRef]
- Gasymov, O.K.; Abduragimov, A.R.; Yusifov, T.N.; Glasgow, J. Resolving near-ultraviolet circular dichroism spectra of single trp mutants in tear lipocalin. Anal. Biochem. 2003, 318, 300–308. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Azimi, O.; Emami, Z.; Salari, H.; Chamani, J. Probing the Interaction of Human Serum Albumin with Norfloxacin in the Presence of High-Frequency Electromagnetic Fields: Fluorescence Spectroscopy and Circular Dichroism Investigations. Molecules 2011, 16, 9792-9818. https://doi.org/10.3390/molecules16129792
Azimi O, Emami Z, Salari H, Chamani J. Probing the Interaction of Human Serum Albumin with Norfloxacin in the Presence of High-Frequency Electromagnetic Fields: Fluorescence Spectroscopy and Circular Dichroism Investigations. Molecules. 2011; 16(12):9792-9818. https://doi.org/10.3390/molecules16129792
Chicago/Turabian StyleAzimi, Olga, Zahra Emami, Hanieh Salari, and Jamshidkhan Chamani. 2011. "Probing the Interaction of Human Serum Albumin with Norfloxacin in the Presence of High-Frequency Electromagnetic Fields: Fluorescence Spectroscopy and Circular Dichroism Investigations" Molecules 16, no. 12: 9792-9818. https://doi.org/10.3390/molecules16129792
APA StyleAzimi, O., Emami, Z., Salari, H., & Chamani, J. (2011). Probing the Interaction of Human Serum Albumin with Norfloxacin in the Presence of High-Frequency Electromagnetic Fields: Fluorescence Spectroscopy and Circular Dichroism Investigations. Molecules, 16(12), 9792-9818. https://doi.org/10.3390/molecules16129792