A Triterpenoid from Thalictrum fortunei Induces Apoptosis in BEL-7402 Cells Through the P53-Induced Apoptosis Pathway

Abstract
:1. Introduction
2. Results and Discussion
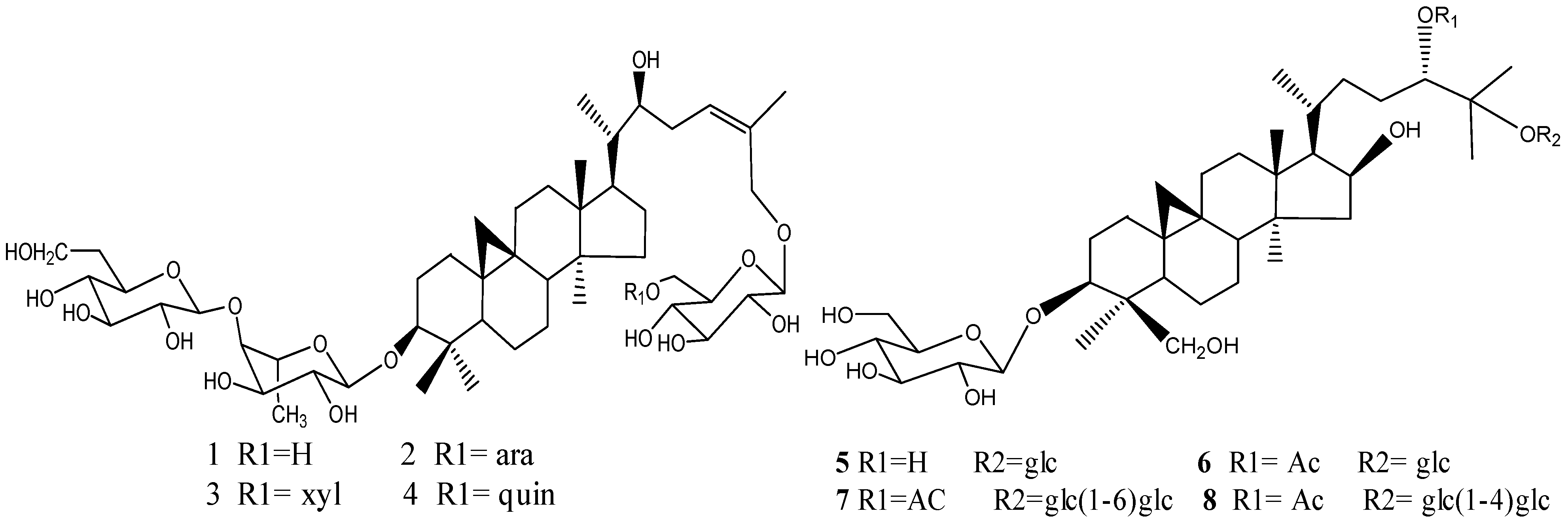
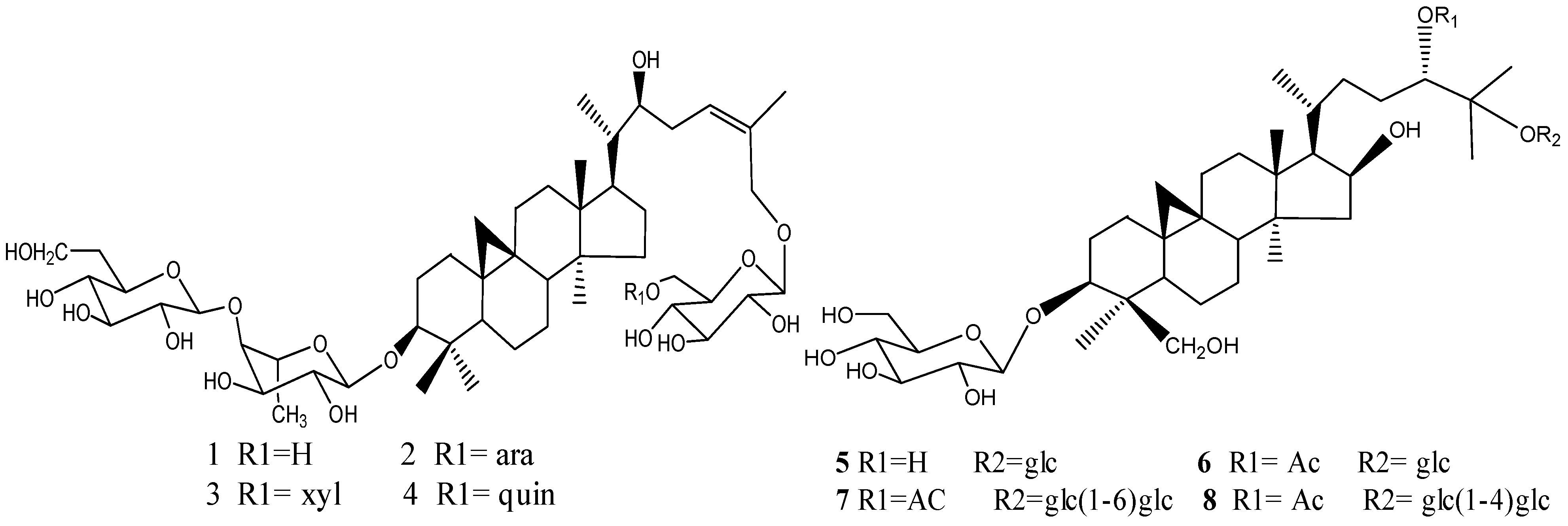
2.1. Compounds Isolated from Thalictrum fortunei
2.2. Growth Inhibitory Effect of Eight Triterpenoids on SGC-7901, Bel-7402, Lovo and NCIH-460 Cells
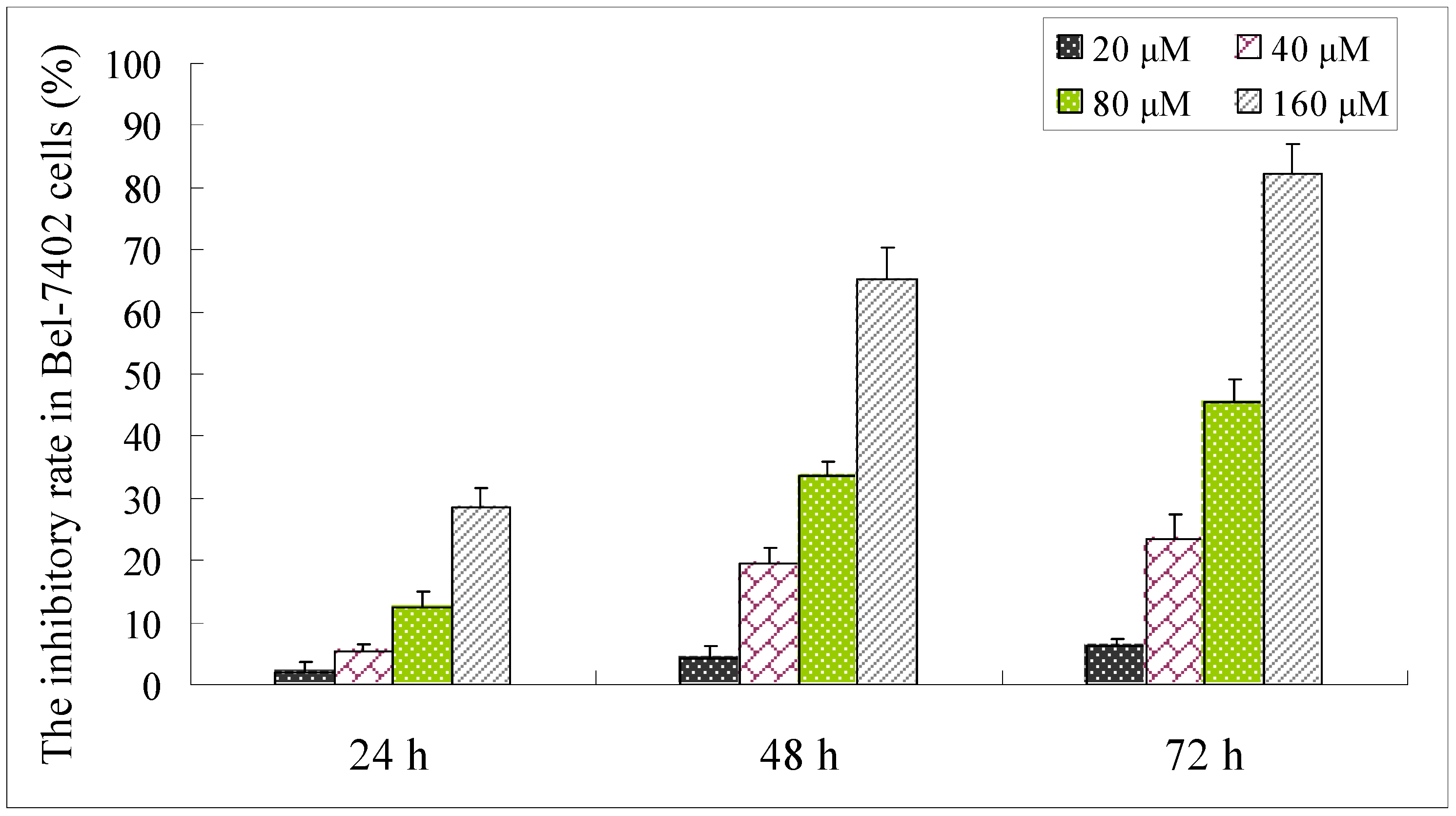
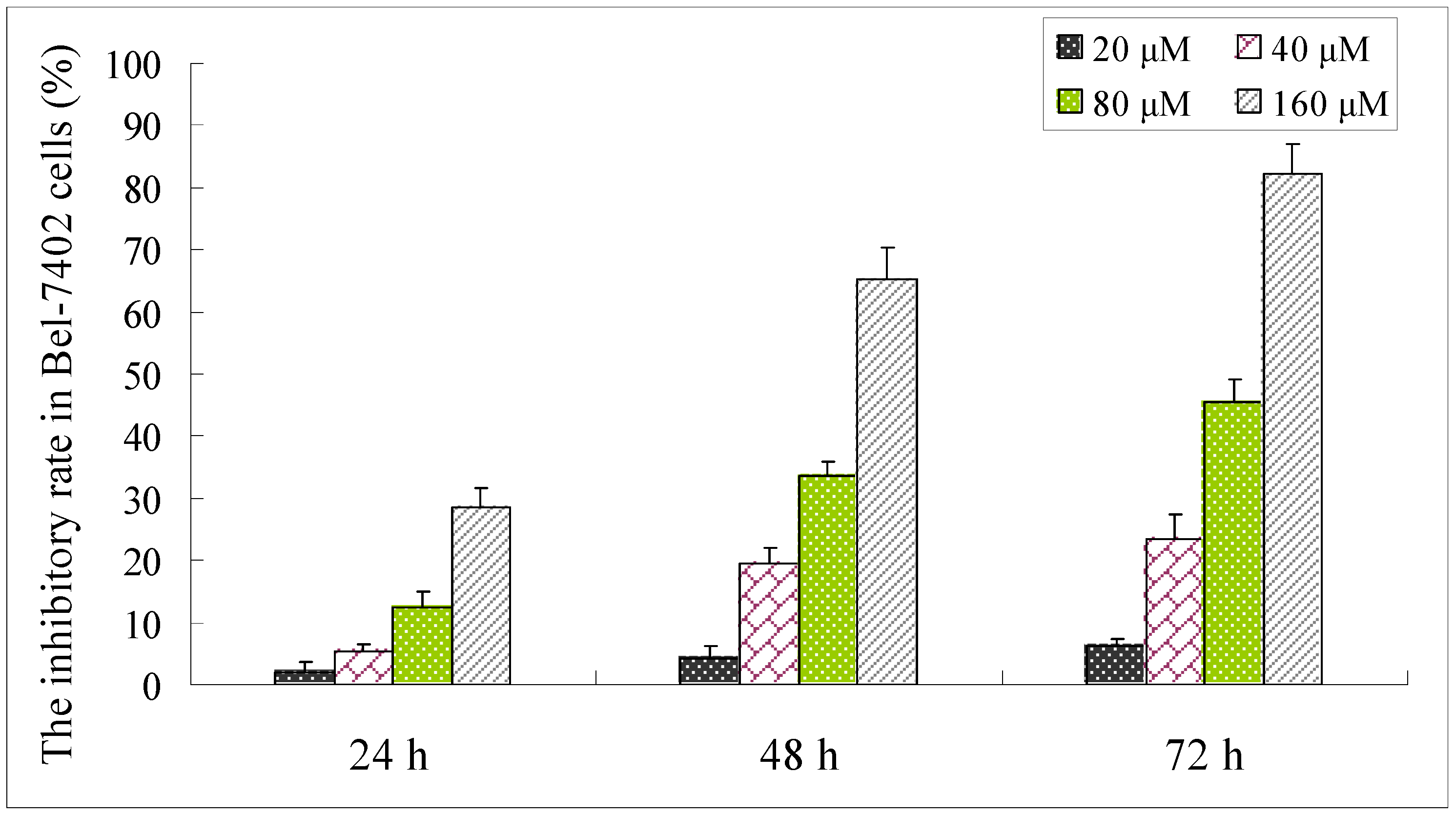
2.3. Effect of Compound 1 on the Viability of Bel-7402 Cells
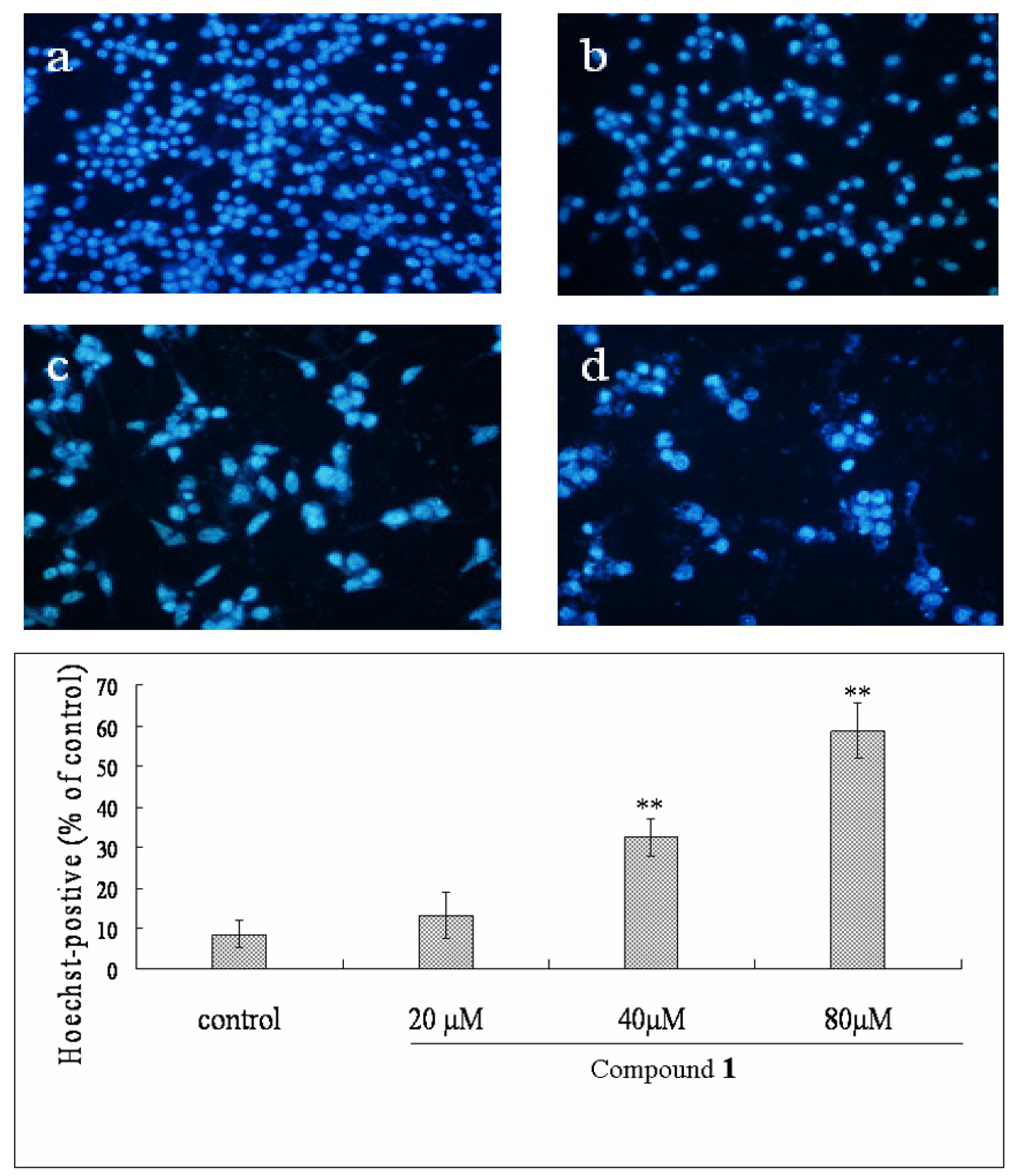
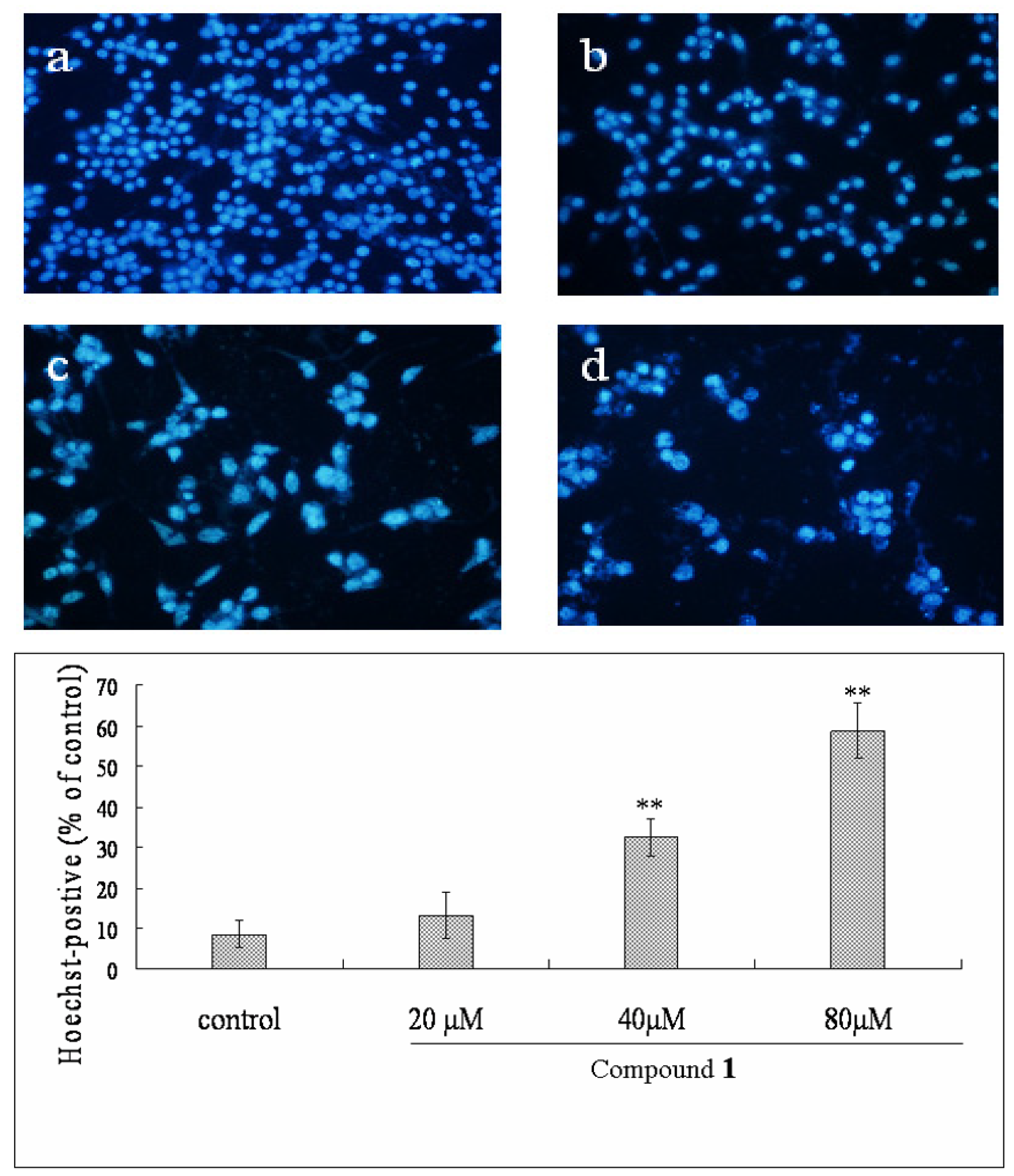
2.4. Effect of Compound 1 on the Morphological Changes of Bel-7402 Cells
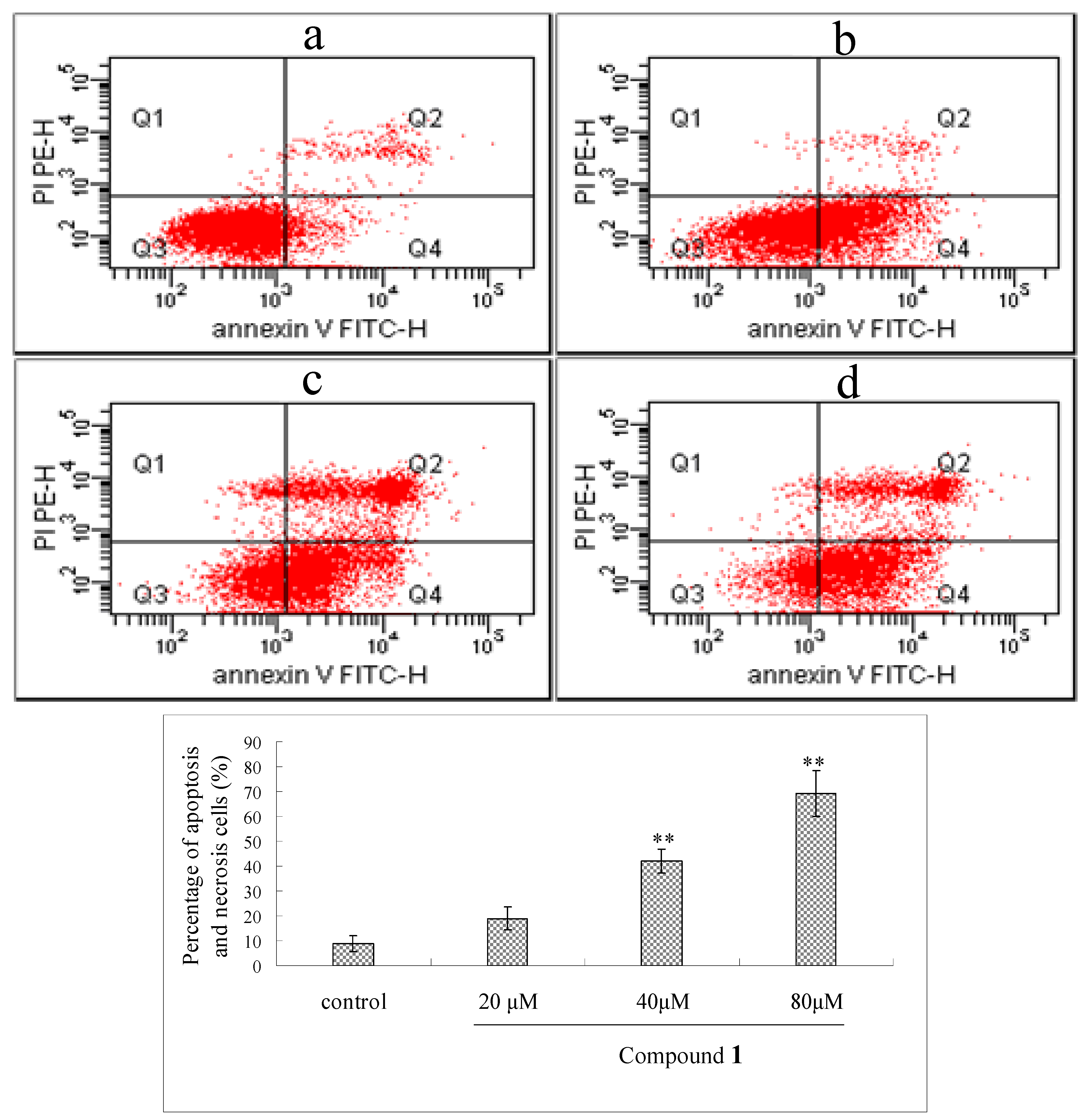
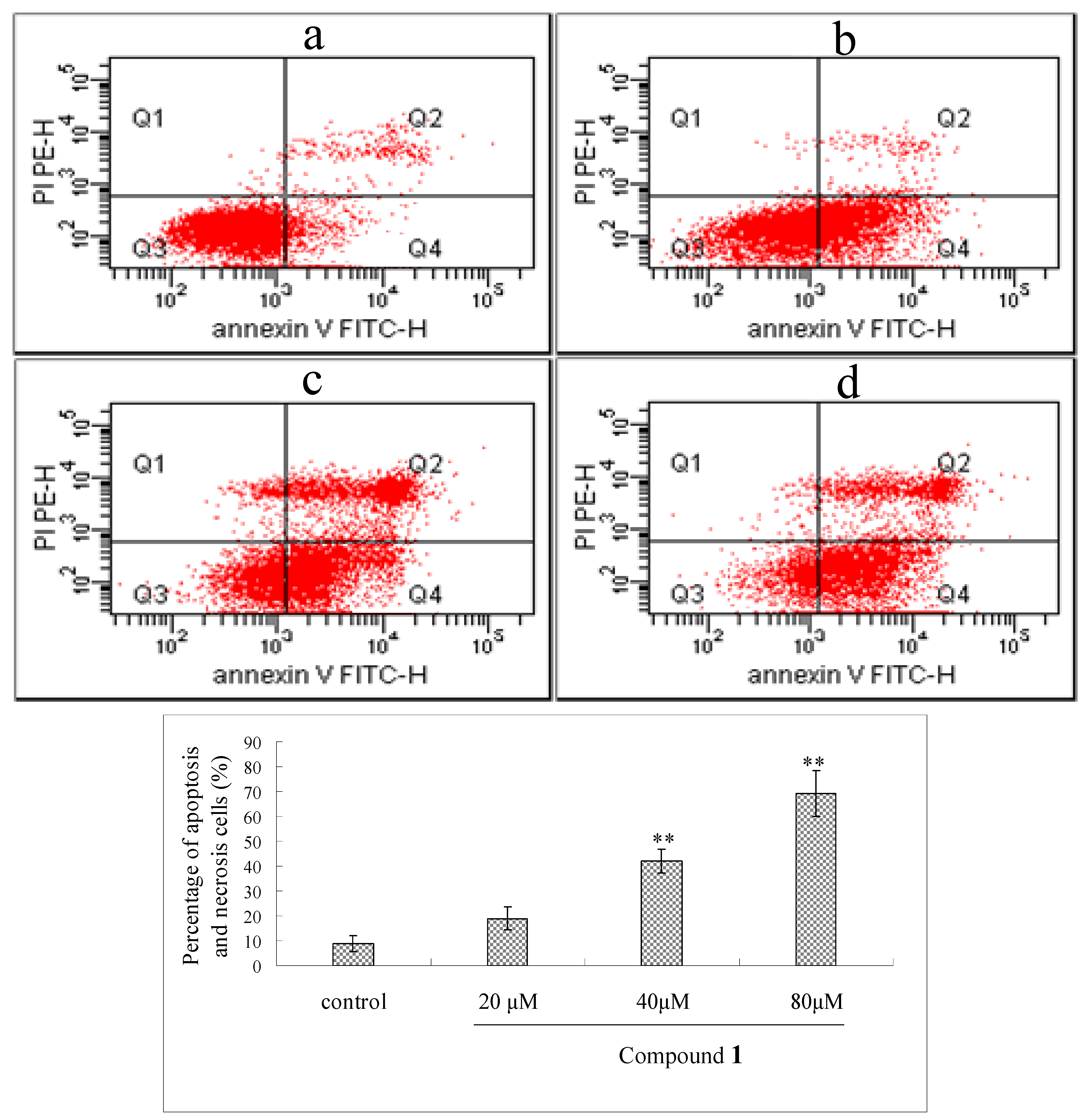
2.5. Flow Cytometry Analysis of Compound 1-Induced Bel-7402 Apoptosis
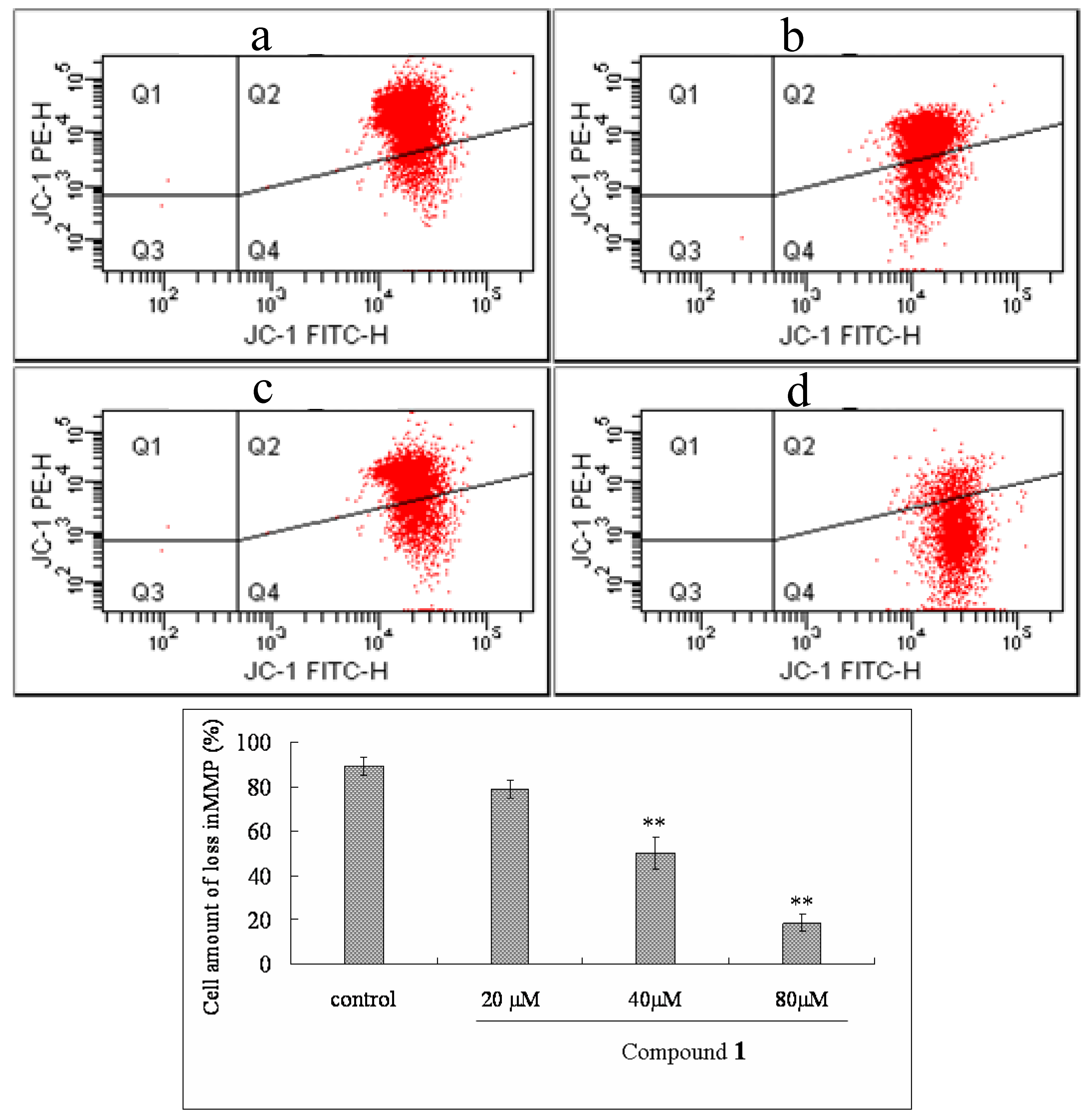
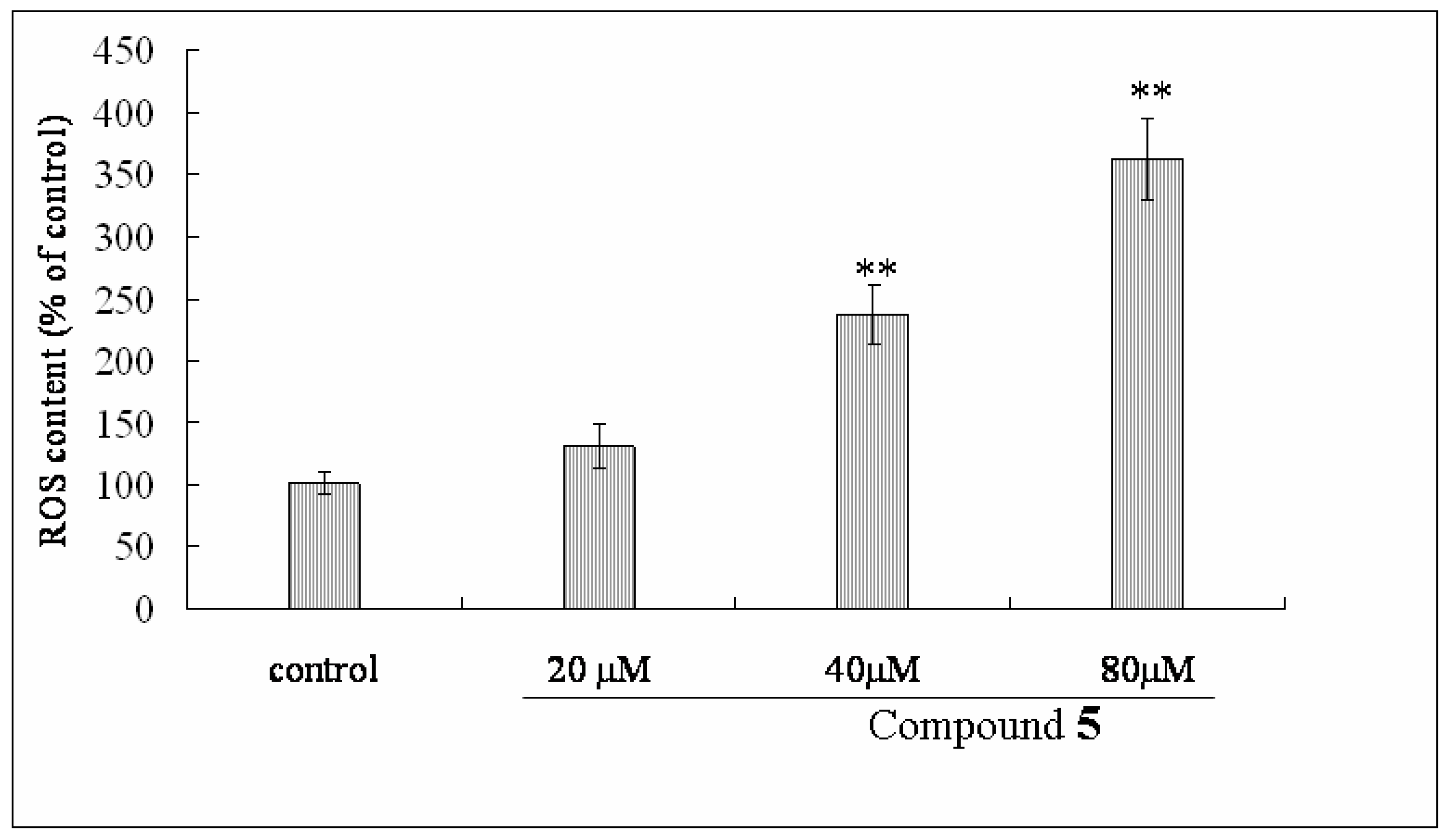
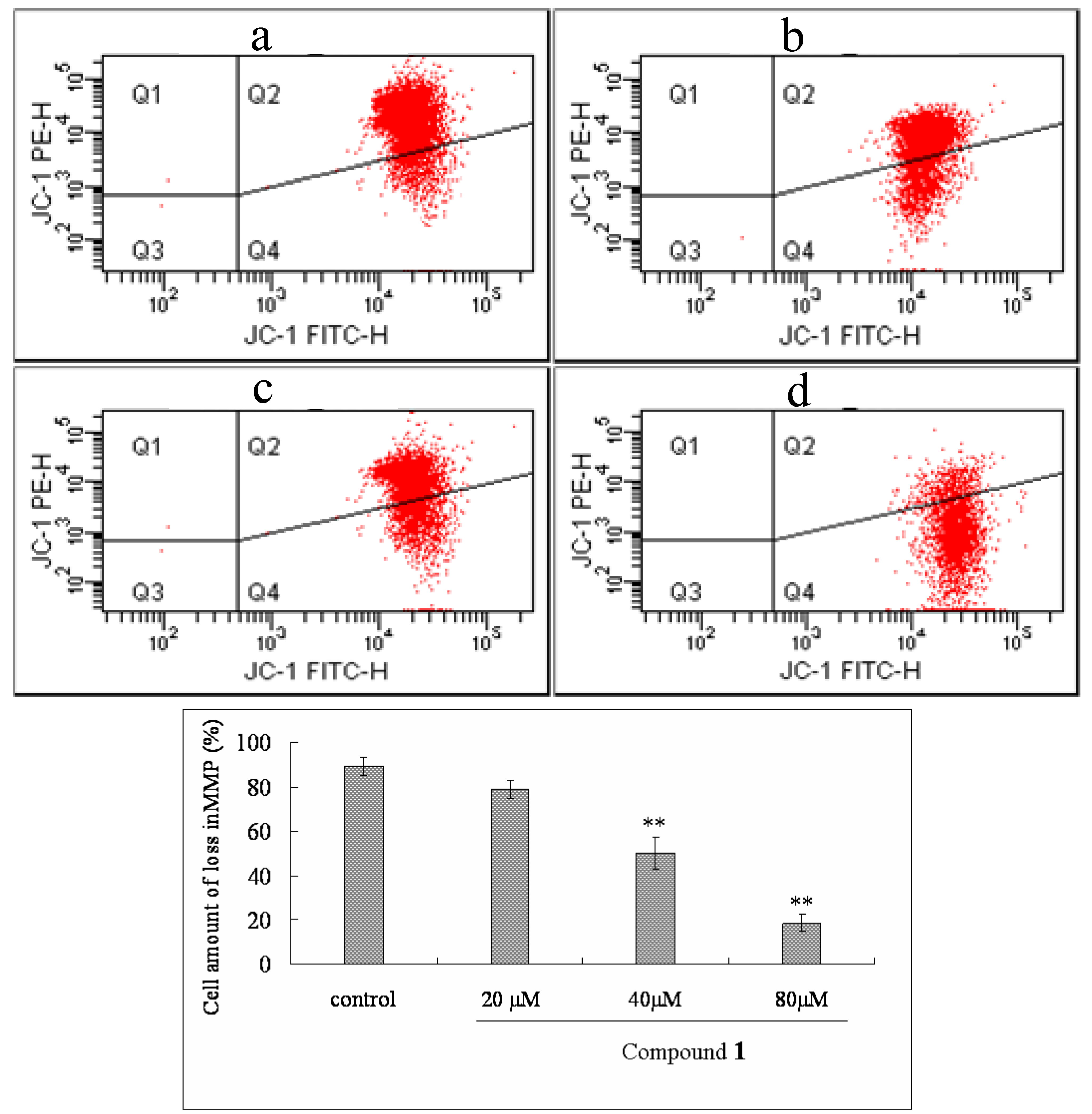
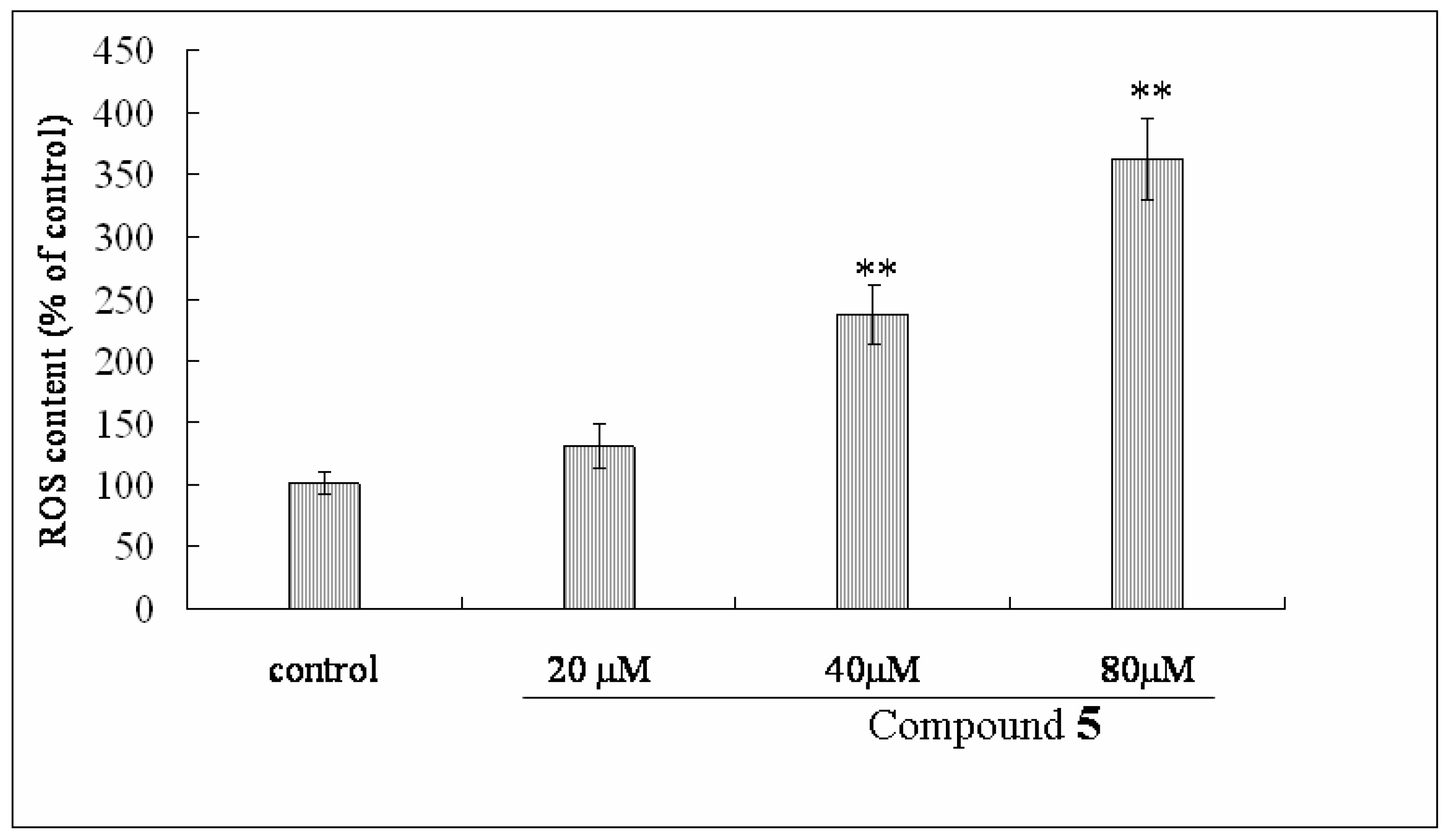
2.6. Effect of Compound 1 on MMP and ROS Levels
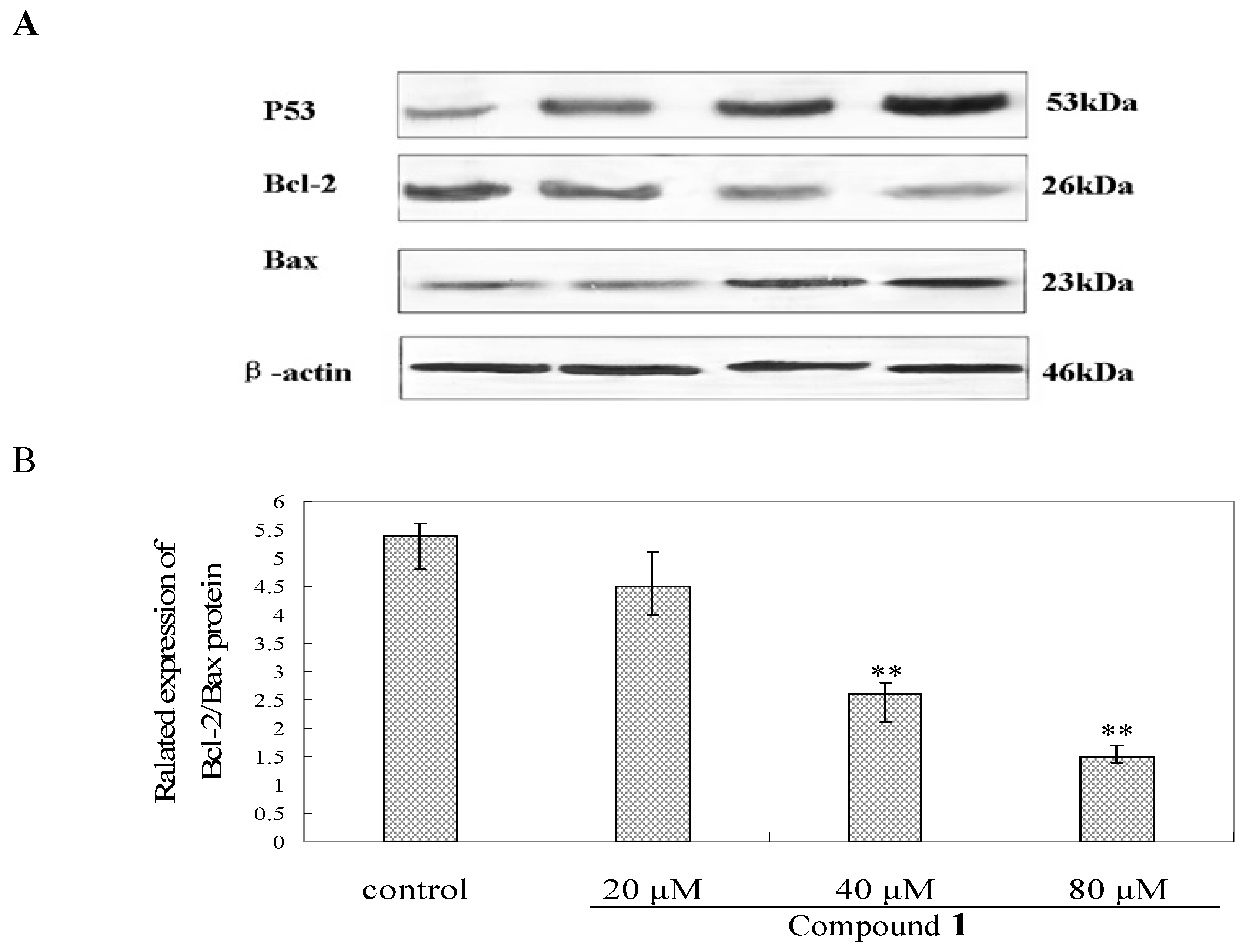
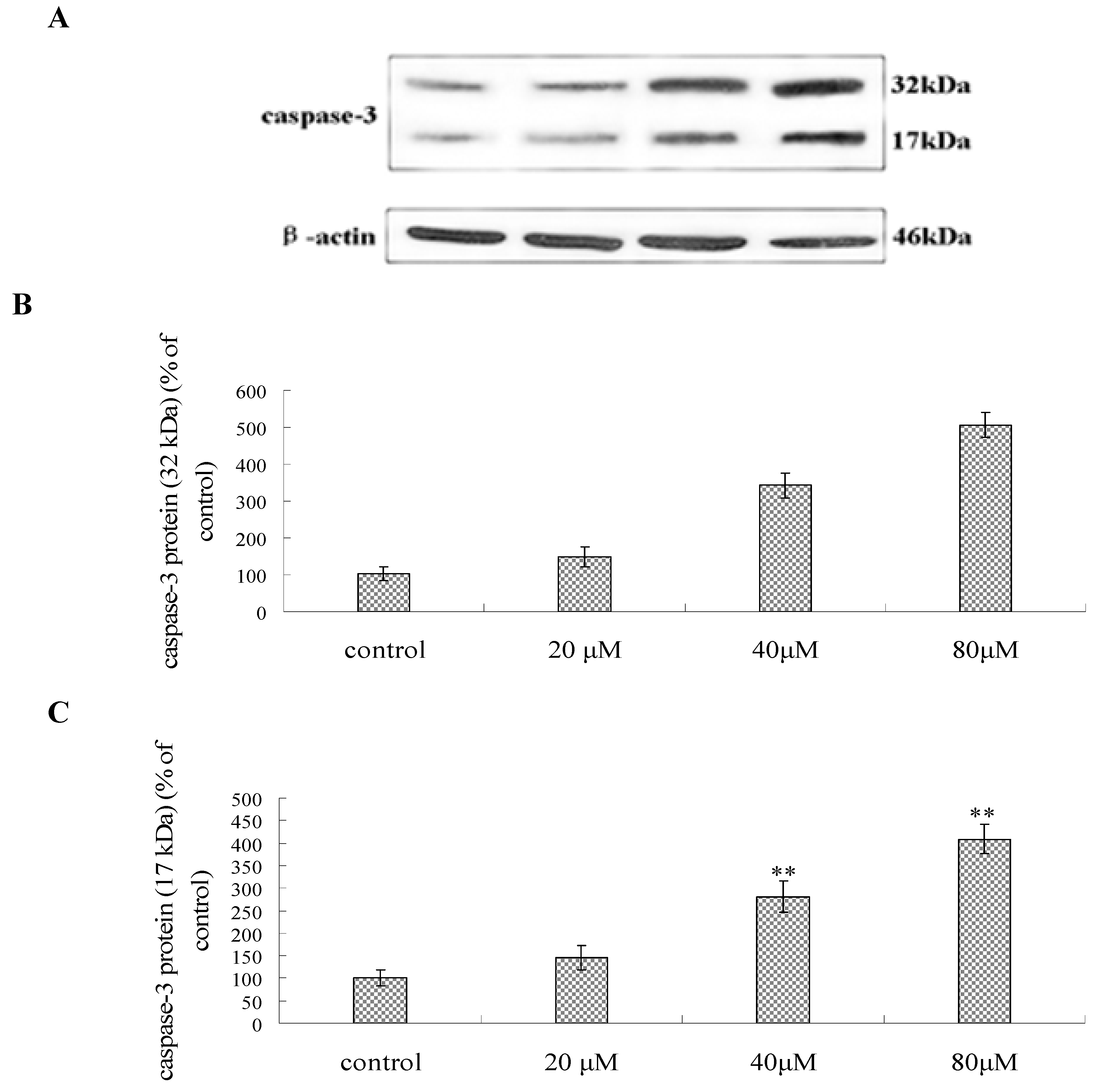
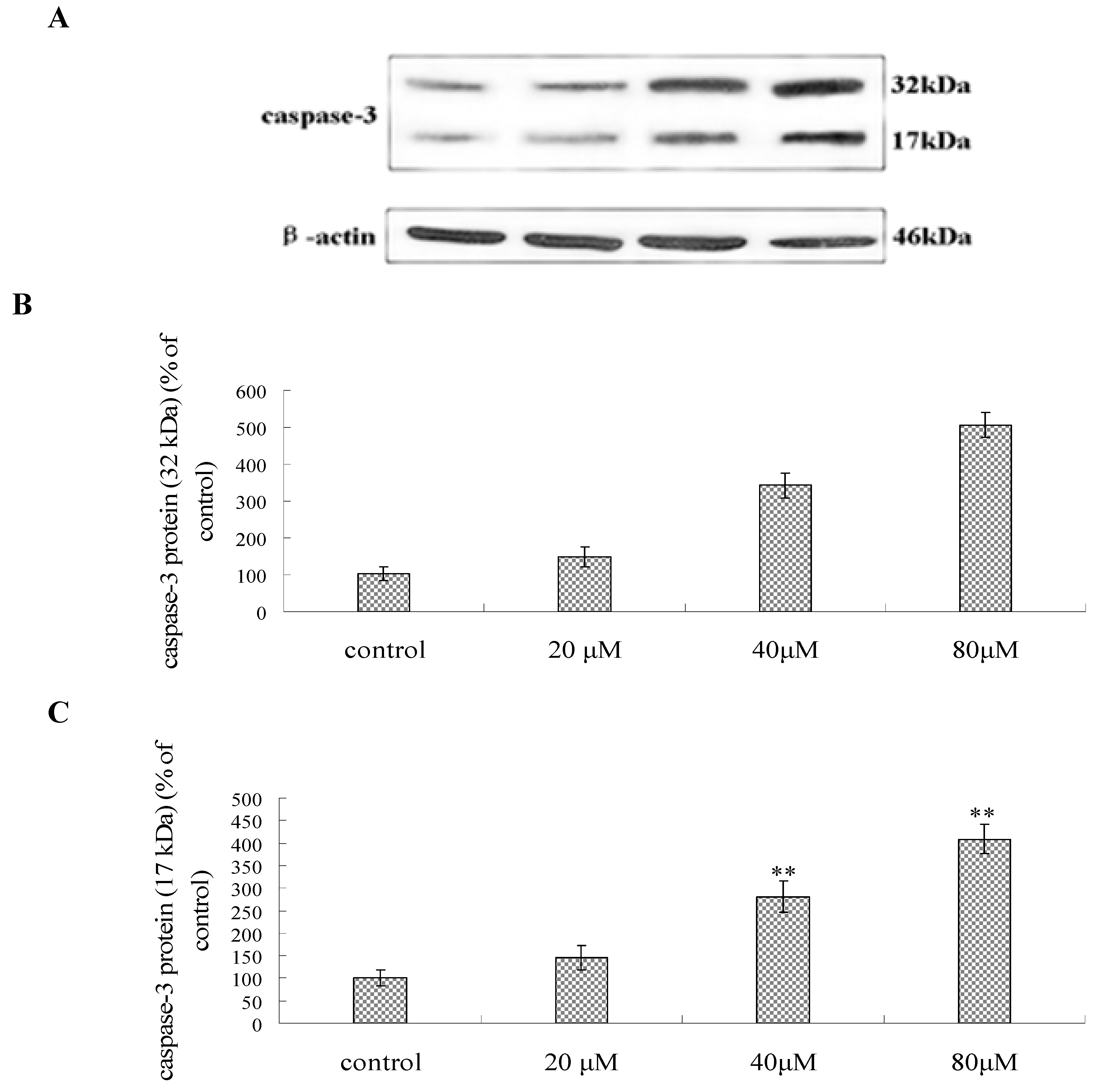
2.7. Effect of Compound 1 on the Expression of Caspase-3, P53, Bcl-2 and Bax Protein
3. Experimental
3.1. Materials
3.2. Isolation and Identification of Compounds from Thalictrum fortunei S. Moore. Rhizome
3.3. Cell Lines and Cell Culture
3.4. Assessment of Anti-Tumor Activities of Eight Triterpenoids from Thalictrum fortunei by MTT Assay
3.5. Assessment of Anti-Tumor Activity of Compound 1 on Bel-7402 Cells by MTT Assay
3.6. Apoptosis Analysis by Hoechst 33258 Staining
3.7. Determination of Cell Apoptosis by Annexin V/PI Double Staining
3.8. Measurement of MMP by Flow Cytometry
3.9. Measurement of Intracellular ROS by Fluorescent Methods
3.10. Western Blot Analysis
3.11. Statistical Analysis
4. Conclusions
Acknowledgements
References and Notes
- Parkin, D.M.; Bray, F.; Ferlay, J.; Pisani, P. Global cancer statistics, 2002. CA Cancer J. Clin. 2005, 55, 74–108. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.R.; Gores, G.J. Emerging drugs for hepatocellular carcinoma. Expert Opin. Emerg. Drugs 2006, 11, 469–487. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.G.; Luo, Y.M.; Chen, J. Survey on Research of Thalictrum Genus Plants in Chemistry and Pharmacody. J. Jiangxi Coll. Tradit. Chin. Med. 2001, 13, 93–97. [Google Scholar]
- Zhang, X.T.; Zhang, L.H.; Ye, W.C.; Zhang, X.L.; Yin, Z.Q.; Zhao, S.X.; Yao, X.S. Four new cycloartane glycosides from Thalictrum fortunei. Chem. Pharm. Bull. 2006, 54, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhang, X.-T.; Wang, L.; Zhang, X.-Q.; Wang, Y.; Chen, S.-B.; Xiao, P.-G.; Ye, W.-C. Four New Cycloartane (=9,19-Cyclolanostane) Saponins from the Aerial Parts of Thalictrum fortunei. Helv. Chim. Acta 2008, 91, 1961–1966. [Google Scholar] [CrossRef]
- Zhao, Q.; Cao, X.; Zeng, B.; Wang, C.; Yan, L.; Xu, C. Musca domestica Larva Lectin Induces Apoptosis in BEL-7402 Cells through a Mitochondria-Mediated Reactive Oxygen Species Way. Biol. Pharm. Bull. 2010, 33, 1274–1278. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.; Xu, X.; Pu, X.; Hou, Z.; Zhang, Q. Cytotoxic effects and the mechanism of three types of magnetic nanoparticles on human hepatoma BEL-7402 cells. Nanoscale Res. Lett. 2011. [Google Scholar] [CrossRef]
- Wang, L.; Chen, G.Y.; Han, C.R.; Yuan, Y.; Yang, B.; Zhang, Y.; Wang, J.; Zhong, X.Q.; Huang, X. Two novel alkaloids from the stem of Saprosma hainanense and their cytotoxic activities in vitro. Chem. Pharm. Bull. 2011, 59, 338–340. [Google Scholar] [CrossRef] [PubMed]
- Fei, H.R.; Chen, G.; Wang, J.M.; Wang, F.Z. Perifosine induces cell cycle arrest and apoptosis in human hepatocellular carcinoma cell lines by blockade of Akt phosphorylation. Cytotechnology 2010, 62, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Aiello, A.; Fattorusso, E.; Luciano, P.; Macho, A.; Menna, M.; Muñoz, E. Antitumor effects of two novel naturally occurring terpene quinones isolated from the Mediterranean ascidian Aplidium conicum. J. Med. Chem. 2005, 48, 3410–3416. [Google Scholar] [CrossRef] [PubMed]
- Park, B.H.; Lee, S.B.; Stolz, D.B.; Lee, Y.J.; Lee, B.C. Synergistic interactions between heregulin and peroxisome proliferator-activated receptor-gamma (PPARgamma) agonist in breast cancer cells. J. Biol. Chem. 2011, 286, 20087–20099. [Google Scholar] [CrossRef] [PubMed]
- Green, D.R.; Kroemer, G. The pathophysiology of mitochondrial cell death. Science 2004, 305, 626–629. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhong, Z.; Xu, Z.; Chen, L.; Wang, Y. No protective effect of curcumin on hydrogen peroxide-induced cytotoxicity in Bel-7402 cells. Pharmacol. Rep. 2011, 63, 724–732. [Google Scholar] [CrossRef]
- Yao, J.; Jiang, Z.; Duan, W.; Huang, J.; Zhang, L.; Hu, L.; He, L.; Li, F.; Xiao, Y.; Shu, B.; et al. Involvement of mitochondrial pathway in triptolide-induced cytotoxicity in human normal liver L-02 cells. Biol. Pharm. Bull. 2008, 31, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Kuribayashi, K.; El-Deiry, W.S. Regulation of programmed cell death by the p53 pathway. Adv. Exp. Med. Biol. 2008, 615, 201–221. [Google Scholar] [PubMed]
- Mihara, M.; Erster, S.; Zaika, A.; Petrenko, O.; Chittenden, T.; Pancoska, P.; Moll, U.M. p53 has a direct apoptogenic role at the mitochondria. Mol. Cell 2003, 11, 577–590. [Google Scholar] [CrossRef]
- Chipuk, J.E.; Kuwana, T.; Bouchier-Hayes, L.; Droin, N.M.; Newmeyer, D.D.; Schuler, M.; Green, D.R. Direct activation of Bax by p53 mediates mitochondrial membrane permeabilization and apoptosis. Science 2004, 303, 1010–1014. [Google Scholar] [CrossRef] [PubMed]
- Sax, J.K.; Fei, P.; Murphy, M.E.; Bernhard, E.; Korsmeyer, S.J.; El-Deiry, W.S. BID regulation by p53 contributes to chemosensitivity. Nat. Cell Biol. 2002, 4, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Bensaad, K.; Tsuruta, A.; Selak, M.A.; Vidal, M.N.; Nakano, K.; Bartrons, R.; Gottlieb, E.; Vousden, K.H. TIGAR, a p53-inducible regulator of glycolysis and apoptosis. Cell 2006, 126, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Syam, S.; Abdul, A.B.; Sukari, M.A.; Mohan, S.; Abdelwahab, S.I.; Wah, T.S. The growth suppressing effects of girinimbine on Bel-7402 involve induction of apoptosis and cell cycle arrest. Molecules 2011, 16, 7155–7170. [Google Scholar] [CrossRef] [PubMed]
- Noh, E.M.; Yi, M.S.; Youn, H.J.; Lee, B.K.; Lee, Y.R.; Han, J.H.; Yu, H.N.; Kim, J.S.; Jung, S.H. Silibinin enhances ultraviolet B-induced apoptosis in mcf-7 human breast cancer cells. J. Breast Cancer 2011, 14, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Feng, J.M. Aristolochic acid induces apoptosis of human umbilical vein endothelial cells in vitro by suppressing PI3K/Akt signaling pathway. Acta Pharmacol. Sin. 2011, 32, 1025–1030. [Google Scholar] [CrossRef] [PubMed]
- Celojevic, D.; Petersen, A.; Karlsson, J.O.; Behndig, A.; Zetterberg, M. Effects of 17β-estradiol on proliferation, cell viability and intracellular redox status in native human lens epithelial cells. Mol. Vis. 2011, 17, 1987–1996. [Google Scholar] [PubMed]
- Liu, W.B.; Zhou, J.; Qu, Y.; Li, X.; Lu, C.T.; Xie, K.L.; Sun, X.L.; Fei, Z. Neuroprotective effect of osthole on MPP+-induced cytotoxicity in PC12 cells via inhibition of mitochondrial dysfunction and ROS production. Neurochem. Int. 2010, 57, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Feng, Q.; Xu, W.; Li, X.; Kang, Z.; Ren, Y.; Du, L. Genipin induced apoptosis associated with activation of the c-Jun NH2-terminal kinase and p53 protein in HeLa cells. Biol. Pharm. Bull. 2010, 33, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors in Guangdong Research Institute of Traditional Chinese Medicine. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Cell line IC50 (μM) a | |||
|---|---|---|---|---|
| Bel-7402 | loVo | NCIH-460 | SGC-7901 | |
| 1 | 66.4 | 84.8 ** | 73.5 * | 89.6 ** |
| 2 | 89.5 | 97.5 | 81.4 | 96.3 |
| 3 | 83.7 | 107.3 | 79.3 | 112.6 |
| 4 | 83.1 | 79.4 | 89.3 | 98.6 |
| 5 | 150.7 | 123.6 | 148.9 | 166.4 |
| 6 | 162.6 | 156.8 | 147.2 | 169.1 |
| 7 | 162.7 | 145.9 | 153.5 | 157.3 |
| 8 | 149.6 | 152.1 | 142.4 | 152.5 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, X.; Zhao, M.; Chen, L.; Jiao, H.; Liu, H.; Wang, L.; Ma, S. A Triterpenoid from Thalictrum fortunei Induces Apoptosis in BEL-7402 Cells Through the P53-Induced Apoptosis Pathway. Molecules 2011, 16, 9505-9519. https://doi.org/10.3390/molecules16119505
Zhang X, Zhao M, Chen L, Jiao H, Liu H, Wang L, Ma S. A Triterpenoid from Thalictrum fortunei Induces Apoptosis in BEL-7402 Cells Through the P53-Induced Apoptosis Pathway. Molecules. 2011; 16(11):9505-9519. https://doi.org/10.3390/molecules16119505
Chicago/Turabian StyleZhang, Xiantao, Ming Zhao, Lvyi Chen, Haoyan Jiao, Hongxia Liu, Liyan Wang, and Shuwei Ma. 2011. "A Triterpenoid from Thalictrum fortunei Induces Apoptosis in BEL-7402 Cells Through the P53-Induced Apoptosis Pathway" Molecules 16, no. 11: 9505-9519. https://doi.org/10.3390/molecules16119505
APA StyleZhang, X., Zhao, M., Chen, L., Jiao, H., Liu, H., Wang, L., & Ma, S. (2011). A Triterpenoid from Thalictrum fortunei Induces Apoptosis in BEL-7402 Cells Through the P53-Induced Apoptosis Pathway. Molecules, 16(11), 9505-9519. https://doi.org/10.3390/molecules16119505