Antioxidant and Anti-Inflammatory Activities of Essential Oils: A Short Review


{kind=link}
Abstract
:1. Introduction
2. How Does an Antioxidant Act?
3. Essential Oils as Antioxidants
3.1. Assays associated with lipid peroxidation
3.1.1. Peroxidation level evaluation using the ferric thiocyanate
3.1.2. Conjugated diene assay
3.1.3. β-Carotene bleaching test
3.1.4. Thiobarbituric acid reactive substances (TBARS)
3.1.5. Aldehyde/carboxylic acid assay
3.1.6. Formic acid measurement
3.2. Free radical scavenging ability
3.2.1. 2,2-Diphenyl-1-picrylhydrazyl (DPPH) assay
3.2.2. Trolox® equivalent antioxidant capacity (TEAC) or ABTS [2,2-Azinobis(3-ethylbenzo-thiazoline-6 sulphonic acid)]
3.2.3. Ferric reducing/antioxidant power (FRAP) assay
3.2.4. Reducing power
3.2.5. Chelating activity
3.2.6. Hydroxyl radical scavenging
3.2.7. Superoxide anion scavenging activity
4. Essential Oils as Anti-Inflammatory Agents
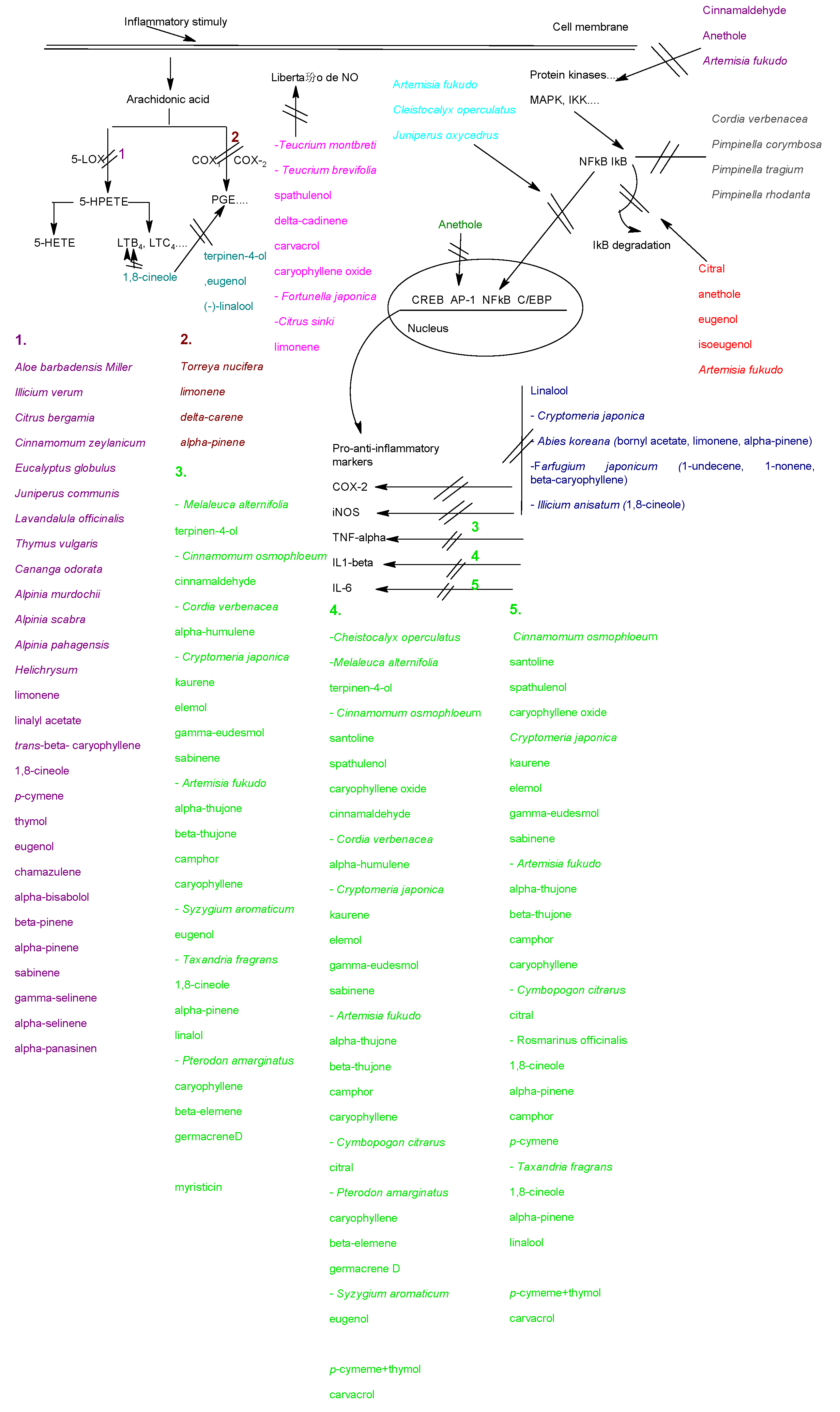
4.1. Effects on the arachidonic metabolism
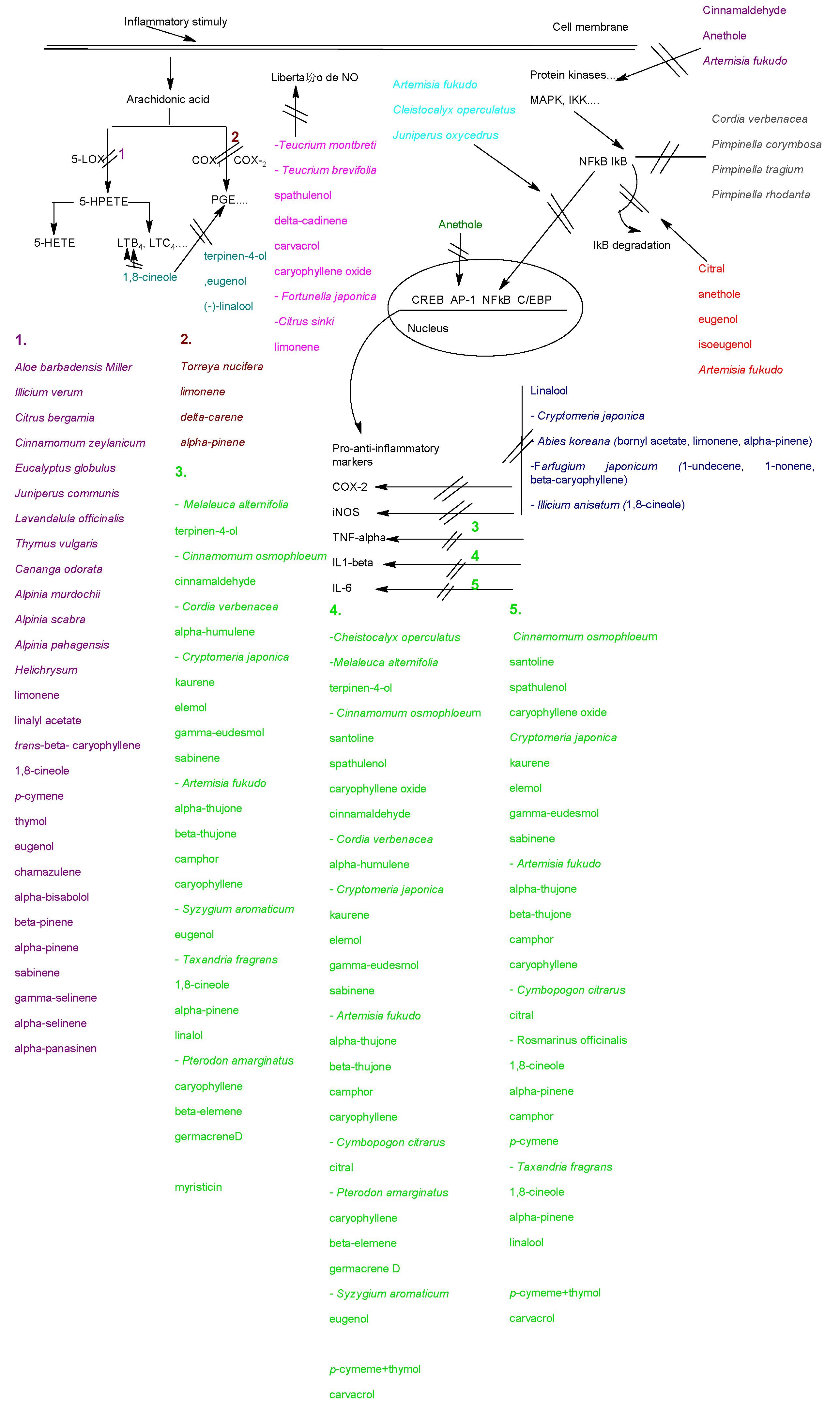
4.2. Effects on cytokines production
4.3. Modulation of pro-inflammatory gene expression
5. Conclusions
References
- Rubiolo, P.; Sgorbini, B.; Liberto, E.; Cordero, C.; Bicchi, C. Essential oils and volatiles: sample preparation and analysis. Flavour Fragr. J. 2010, 25, 282–290. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: their antibacterial properties and potential applications in foods – a review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M.M. Biological effects of essential oils- a review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Pourmortazavi, S.M.; Hajimirsadeghi, S.S. Supercritical fluid extraction in plant essential and volatile oil analysis. J. Chromatogr. A 2007, 1163, 2–24. [Google Scholar] [CrossRef]
- Miguel, M.G. Antioxidant activity of medicinal and aromatic plants. Flavour Fragr. J. 2010, 25, 291–312. [Google Scholar] [CrossRef]
- Sangwan, N.S.; Farooqui, A.H.A.; Shabih, F.; Sangwan, R.S. Regulation of essential oil production in plants. Plant Growth Regul. 2001, 34, 3–21. [Google Scholar] [CrossRef]
- Cavaleiro, C.M.F. Óleos essenciais de Juniperus de Portugal. PhD Thesis, Universidade de Coimbra, Faculdade de Farmácia, Coimbra, Portugal, 2001. [Google Scholar]
- Bicchi, C.; Liberto, E.; Matteodo, M.; Sgorbini, B.; Mondello, L.; Zellner, B.A.; Coata, R.; Rubiolo, P. Quantitative analysis of essential oils: a complex task. Flavour Fragr. J. 2008, 23, 382–391. [Google Scholar] [CrossRef]
- Dewick, P.M. The biosynthesis of C5-C-25 terpenoid components. Nat. Prod. Rep. 2002, 19, 181–222. [Google Scholar] [CrossRef]
- Litchenthaler, H.K. The 1-deoxy-D-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 47–65. [Google Scholar] [CrossRef]
- Gounaris, Y. Biotechnology for the production of essential oils, flavours and volatile isolates. Flavour Fragr. J. 2010. [Google Scholar] [CrossRef]
- Asaph, A.; Jongsma, M.A.; Kim, T.-Y.; Ri, M.-B.; Giri, A.P.; Verstappen, W.A.; Schwab, W.; Bouwmeester, H.J. Metabolic engineering of terpenoid biosynthesis in plants. Phytochem. Rev 2006, 5, 49–58. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Kainulainen, P.; Aflatuni, A.; Tiilikkala, K.; Holopainen, J.K. Insecticidal, repellent, antimicrobial activity and Phytotoxicity of essential oils: with special reference to limonene and its suitability for control of insect pest. Agr. Food Sci. Finland 2001, 10, 243–259. [Google Scholar]
- Reichling, J.; Schnitzler, P.; Suschke, U.; Saller, R. Essential oils of aromatic plants with antibacterial, antifungal, antiviral, and cytotocic properties-an overview. Forsch. Komplementmed. 2009, 16, 79–90. [Google Scholar]
- Maestri, D.M.; Nepote, V.; Lamarque, A.L.; Zygadlo, J.A. Natural products as antioxidants. In Phytochemistry: Advances in Research; Imperato, F., Ed.; Research Signopost: Kerala, India, 2006; pp. 105–135. [Google Scholar]
- Aruoma, O.I. Free radicals, oxidative stress, and antioxidants in human health and disease. J. Am. Oil Chem. Soc. 1998, 75, 199–212. [Google Scholar] [CrossRef]
- Kamatou, G.P.P.; Viljoen, A.M. A review of the application and pharmacological properties of α-bisabolol and α-bisabolol-rich oils. J. Am. Oil Chem. Soc. 2010, 87, 1–7. [Google Scholar] [CrossRef]
- Babior, B.M. NADPH oxidase. Curr. Opin. Immunol. 2004, 16, 42–47. [Google Scholar] [CrossRef]
- Quinn, M.T.; Gauss, K.A. Structure and regulation of the neutrophil respiratory burst oxidase: comparison with nonphagocyte oxidases. J. Leukocyte Biol. 2004, 76, 760–781. [Google Scholar] [CrossRef]
- Fridovich, I. Superoxide anion radical (O2•-), superoxide dismutases, and related matter. J. Biol. Chem. 1997, 272, 18515–18517. [Google Scholar] [CrossRef]
- Chen, S.-X.; Schopfer, P. Hydroxyl-radical production in physiological reaction. A novel function of peroxidase. Eur. J. Biochem. 1999, 260, 726–735. [Google Scholar] [CrossRef]
- Gomes, A.; Fernandes, E.; Lima, J.L.F.C.; Mira, L.; Corvo, M.L. Molecular mechanisms of anti-inflammatory activity mediated by flavonoids. Curr. Med. Chem. 2008, 15, 1586–1605. [Google Scholar] [CrossRef]
- Kostka, P. Free radicals (nitric oxide). Anal. Chem. 1995, 67, 411–416. [Google Scholar] [CrossRef]
- Nagano, T. Practical methods for detection of nitric oxide. Luminescence 1999, 14, 283–290. [Google Scholar] [CrossRef]
- Miyasaka, N.; Hirata, Y. Nitric oxide and inflammatory arthritides (minireview). Life Sci. 1997, 61, 2073–2081. [Google Scholar] [CrossRef]
- Halliwell, B. What nitrates tyrosine? Is nitrotyrosine specific as a biomarker of peroxynitrite formation in vivo? FEBS Lett. 1997, 411, 157–160. [Google Scholar] [CrossRef]
- Karadag, A.; Ozcelik, B.; Saner, S. Review of methods to determine antioxidant capacities. Food Anal. Methods 2009, 2, 41–60. [Google Scholar] [CrossRef]
- Singh, S.; Singh, R.P. In vitro methods of assay of antioxidants: an overview. Food Rev. Int. 2008, 24, 392–415. [Google Scholar] [CrossRef]
- Roginsky, V.; Lissi, E.A. Review of methods to determine chain-breaking antioxidant activity in food. Food Chem. 2005, 92, 235–254. [Google Scholar] [CrossRef]
- Niki, E. Assessment of antioxidant capacity in vitro and in vivo. Free Radic. Biol. Med. 2010, 9, 503–515. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agr. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Laguerre, M.; Lecomte, J.; Villeneuve, P. Evaluation of the ability of antioxidants to counteract lipid oxidation: existing methods, new trends and challenges. Prog. Lipid Res. 2007, 46, 244–282. [Google Scholar] [CrossRef]
- Frankel, E.N.; Meyer, A.S. The problems of using one dimensional methods to evaluate multifunctional food and biological antioxidants. J. Sci. Food Agr. 2000, 80, 1925–1941. [Google Scholar] [CrossRef]
- Antolovich, M.; Prenzler, P.D.; Patsalides, E.; McDonald, S.; Robards, K. Critical review: methods for testing antioxidant activity. Analyst 2002, 127, 183–198. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Sánchez-Moreno, C. Review: Methods used to evaluate the free radical scavenging activity in foods and biological systems. Food Sci. Technol. Int. 2002, 8, 121–137. [Google Scholar]
- Becker, E.M.; Nissen, L.R.; Skibsted, L.H. Antioxidant evaluation protocols: food quality or health effects. Eur. Food Res. Technol. 2004, 219, 561–571. [Google Scholar] [CrossRef]
- Decker, E.A.; Warner, K.; Richards, M.P.; Shahidi, F. Measuring antioxidant rffectiveness in food. J. Agr. Food Chem. 2005, 53, 4303–4310. [Google Scholar] [CrossRef]
- Frankel, E.N. In search of better methods to evaluate natural antioxidants and oxidative stability in food lipids. Trends Food Sci. Technol. 1993, 4, 220–225. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agr. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- MacDonalds-Wicks, L.; Wood, L.G.; Garg, M.L. Methodology for the determination of biological antioxidant capacity in vitro: a review. J. Sci. Food Agr. 2006, 86, 2046–2056. [Google Scholar] [CrossRef]
- Yang, S.-A.; Jeon, S.-K.; Lee, E.-J.; Shim, E.-H.; Lee, I.-S. Comparative study of the chemical composition and antioxidant activity of six essential oils and their components. Nat. Prod. Res. 2010, 24, 140–151. [Google Scholar] [CrossRef]
- Wei, A.; Shibamoto, T. Antioxidant/lipoxygenase inhibitory activities and chemical compositions of selected essential oils. J. Agr. Food Chem. 2010, 58, 7218–7225. [Google Scholar]
- Joshi, S.C.; Verma, A.R.; Mathela, C.S. Antioxidant and antibacterial activities of the leaf essential oils of Himalayan Lauraceae species. J. Chem. Toxicol. 2010, 48, 37–40. [Google Scholar] [CrossRef]
- Mighri, H.; Hajlaoui, H.; Akrout, A.; Najjaa, H.; Neffati, M. Antimicrobial and antioxidant activities of Artemisia herba-alba essential oil cultivated in Tunisian arid zone. C.R. Chim. 2010, 13, 380–386. [Google Scholar] [CrossRef]
- Wannes, W.A.; Mhamdi, B.; Sriti, J.; Jenia, M.B.; Ouchikh, O.; Hamdaoni, G.; Kchouk, M.E.; Marzouk, B. Antioxidant activities of the essential oils and methanol extracts from myrtle (Myrtus communis var. italica) leaf, stem and flower. Food Chem. Toxicol. 2010, 48, 1362–1370. [Google Scholar] [CrossRef]
- Ahmadi, F.; Sadeghi, S.; Modarresi, M.; Abiri, R.; Mikaeli, A. Chemical composition, in vitro antimicrobial, antifungal and antioxidant activities of the essential oil and methanolic extract of Hymenocrater longiflorus Benth., of Iran. Food Chem. Toxicol. 2010, 48, 1137–1144. [Google Scholar] [CrossRef]
- Ebrahimabadi, A.H.; Mazoochi, A.; Kashi, F.J.; Djafari-Bidgoli, Z.; Batooli, H. Essential oil composition and antioxidant and antimicrobial properties of the aerial parts of Salvia eremophila Boiss. from Iran. Food Chem. Toxicol. 2010, 48, 1371–1376. [Google Scholar] [CrossRef]
- Gholivand, M.B.; Rahimi-Nasrabadi, M.; Batooli, H.; Ebrahimabadi, A.H. Chemical composition and antioxidant activities of the essential oil and methanol extracts of Psammogeton canescens. Food Chem. Toxicol. 2010, 48, 24–28. [Google Scholar] [CrossRef]
- Goze, I.; Alim, A.; Cetinus, S.A.; Cetin, A.; Durmus, N.; Atas, A.T.; Vural, N. In vitro antimicrobial, antioxidant, and antispasmodic activities and the composition of the essential oil of Origanum acutidens (Hand.-Mazz.) letswaart. J. Med. Food 2010, 13, 705–709. [Google Scholar] [CrossRef]
- Patil, R.P.; Nimbalkar, M.S.; Jadhav, U.U.; Dawkar, V.V.; Govindwar, S.P. Antiaflatoxigenic and antioxidant activity of an essential oil from Ageratum conyzoides L. J. Sci. Food Agr. 2010, 90, 608–614. [Google Scholar]
- Viuda-Martos, M.; Navajas, Y.R.; Zapata, E.S.; Fernández-López, J.; Pérez- Alvarez, J.A. Antioxidant activity of essential oils of five spice plants widely used in a Mediterranean diet. Flavour Fragr. J. 2010, 25, 13–19. [Google Scholar] [CrossRef]
- Dandlen, S.A.; Lima, A.S.; Mendes, M.D.; Miguel, M.G.; Faleiro, M.L; Sousa, M.J.; Pedro, L.G.; Barroso, J.G.; Figueiredo, A.C. Antioxidant activity of six Portuguese thyme species essential oils. Flavour Fragr. J. 2010, 25, 150–155. [Google Scholar] [CrossRef]
- Jia, H.L.; Ji, Q.L.; Xing, S.L.; Zhang, P.H.; Zhu, G.L.; Wang, X.H. Chemical composition and antioxidant, antimicrobial activities of the essential oils of Thymus marschallianus Will. and Thymus proximus Serg. J. Food Sci 2010, 75, E59–E65. [Google Scholar] [CrossRef]
- Kulisic-Bilusic, T.; Blazevic, I.; Dejanovic, B.; Milos, M.; Pifat, G. Evaluation of the antioxidant activity of essential oils from caper (Capparis spinosa) and sea fennel (Crithmum maritimum) by different methods. J. Food Biochem. 2010, 34, 286–302. [Google Scholar] [CrossRef]
- Yang, Y.; Yue, Y.; Runwei, Y.; Guolin, Z. Cytotoxic, apoptotic and antioxidant activity of the essential oil of Amomum tsao-ko. Bioresour. Technol. 2010, 101, 4205–4211. [Google Scholar] [CrossRef]
- Miguel, M.G.; Cruz, C.; Faleiro, L.; Simões, M.T.F.; Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G. Foeniculum vulgare essential oils: chemical composition, antioxidant and antimicrobial activities. Nat. Prod. Commun. 2010, 5, 319–328. [Google Scholar]
- Suanarunsawat, T.; Ayutthaya, W.D.N.; Songsak, T.; Thirawarapan, S.; Poungshompoo, S. Antioxidant activity and lipid-lowering effect of essential oils extracted from Ocimum sanctum L. leaves in rats fed with a high cholesterol diet. J. Clin. Biochem. Nutr. 2010, 46, 52–59. [Google Scholar]
- Moon, J.-K.; Shibamoto, T. Antioxidant assays for plant and food components. J. Agr. Food Chem. 2009, 57, 1655–1666. [Google Scholar] [CrossRef]
- Grosso, C.; Figueiredo, A.C.; Burillo, J.; Mainar, A.M.; Urieta, J.S.; Barroso, J.G.; Coelho, J.A.; Palavra, A.M.F. Composition and antioxidant activity of Thymus vulgaris volatiles: comparison between supercritical fluid extraction and hydrodistillation. J. Sep. Sci. 2010, 33, 2211–2218. [Google Scholar] [CrossRef]
- Mothana, R.A.; Al-Rehaily, A.J.; Schultze, W. Chemical analysis and biological activity of the essential oils of two endemic Soqotri Commiphora species. Molecules 2010, 15, 689–698. [Google Scholar] [CrossRef]
- Barra, A.; Coroneo, V.; Dessi, S.; Cabras, P.; Angioni, A. Chemical variability, antifungal and antioxidant activity of Eucalyptus camaldulensis essential oil from Sardinia. Nat. Prod. Commun. 2010, 5, 329–335. [Google Scholar]
- Mimika-Dukić, N.; Bugarin, D.; Grebovi, S.; Mitić-Ćulafić, D.; Vuković-Gačić, D.; Jovin, E.; Couladis, M. Essential oil of Myrtus communis L. as a potential antioxidant and antimutagenic agents. Molecules 2010, 15, 2759–2770. [Google Scholar] [CrossRef]
- Alcântara, J.M.; Yamaguchi, K.K.L.; Junior, V.F.V. Composição química de óleos essenciais de espécies de Aniba e Licaria e suas actividades antioxidante e antiagregante plaquetária. Quim. Nova 2010, 33, 141–145. [Google Scholar] [CrossRef]
- Saleh, M.A.; Clark, S.; Woodard, B.; Deolu-Sobogun, A.A. Antioxidant and free radical scavenging activities of essential oils. Ethn. Dis. 2010, 20, SI78–SI82. [Google Scholar]
- Martino, L.; Feo, V.; Fratianni, F.; Nazzaro, F. Chemistry, antioxidant, antibacterial and antifungal activities of volatile oils and their components. Nat. Prod. Commun. 2010, 5, 1741–1750. [Google Scholar]
- Singh, P.; Shukla, R.; Prakash, B.; Kumar, A.; Singh, S.; Mishra, P.K.; Dubey, N.K. Chemical profile, antifungal, antiaflatoxigenic and antioxidant activity of Citrus maxima Burm. and Citrus sinensis (L.) Osbeck essential oils and their cyclic monoterpene, DL-limonene. J. Chem. Toxicol 2010, 48, 1734–1740. [Google Scholar] [CrossRef]
- Firuzi, O.; Asadollahi, M.; Gholami, M.; Javidnia, K. Composition and biological activities of essential oils from four Heracleum species. Food Chem. 2010, 122, 117–122. [Google Scholar] [CrossRef]
- Sánchez-Arana, A.; Estarrón-Espinosa, M.; Obledo,-Vázquez, E.N.; Camberos, E.P.; Silva-Vázquez, R.; Lugo-Cervantes, E. Antimicrobial and antioxidant activities of Mexican oregano essential oils (Lippia graveolens H. B. K.) with different composition when microencapsulated in b-cyclodextrin. Lett. Appl. Microbiol. 2010, 50, 585–590. [Google Scholar] [CrossRef]
- Haloui, E.; Marzouk, Z.; Marzouk, B.; Bouffira, I.; Bouraoui, A.; Fenina, N. Pharmacological activities and chemical composition of the Olea europaea L. leaf essential oils from Tunisia. J. Food Agr. Environ. 2010, 8, 204–208. [Google Scholar]
- Mechergui, K.; Coelho, J.A.; Serra, M.C.; Lamine, S.B.; Boukhchina, S.; Khouja, M.L. Essential oils of Origanum vulgare L. subsp. glandulosum (Desf.) letswaart from Tunisia: chemical composition and antioxidant activity. J. Sci. Food Agr. 2010, 90, 1745–1749. [Google Scholar] [CrossRef]
- Edziri, H.; Mastouri, M.; Chéraif, I.; Aouini, M. Chemical composition and antibacterial, antifungal and antioxidant activities of the flower oil of Retama raetam (Forssk.) Webb from Tunisia. Nat. Prod. Res. 2010, 24, 789–796. [Google Scholar] [CrossRef]
- Jabrane, A.; Jannet, H.B.; Mastouri, M.; Mighri, Z.; Casanova, J. Chemical composition and in vitro evaluation of antioxidant and antibacterial activities of the root of Ridolfa segetum (L.) Moris from Tunisia. Nat. Prod. Res. 2010, 24, 491–499. [Google Scholar] [CrossRef]
- Saei-Dehkordi, S.S.; Tajik, H.; Moradi, M.; Khalighi-Sigaroodi, F. Chemical composition of essential oils in Zataria multiflora Boiss. from different parts of Iran and their radical scavenging and antimicrobial activity. Food Chem. Toxicol. 2010, 48, 1562–1567. [Google Scholar] [CrossRef]
- Ennajar, M.; Bouajila, J.; Lebrihi, A.; Mathieu, F.; Savagnac, A.; Abderraba, M.; Raies, A.; Romdhane, M. The influence of organ, season and drying method on chemical composition and antioxidant and antimicrobial activities of Juniperus phoenica L. essential oils. essential oils. J. Sci. Food Agr. 2010, 90, 462–470. [Google Scholar]
- Ghazghazi, H.; Miguel, M.G.; Hasnaoui, B.; Sebei, H.; Ksontini, M.; Figueiredo, A.C.; Pedro, L.G.; Barroso, J.G. Phenols, essential oils, and carotenoids of Rosa canina from Tunisia and their antioxidant activities. Afr. J. Biotechnol. 2010, 9, 2709–2710. [Google Scholar]
- Jordan, M.J.; Sanchez-Gomez, P.; Jimenez, J.F.; Quilez, M.; Sotomayor, J.A. Chemical composition and antiradical activity of the essential oil from Satureja intricata, S. obovata; and their hybrid Datureja x delpozoi. Nat. Prod. Commun. 2010, 5, 629–634. [Google Scholar]
- Gourine, N.; Yousfi, M.; Bombarda, I.; Nadjemi, B.; Stocker, P.; Gaydon, E.M. Antioxidant activities and chemical composition of essential oil of Pistacia atlantica from Algeria. Ind. Crop Prod. 2010, 31, 203–208. [Google Scholar] [CrossRef]
- El-Ghorab, A.H.; Nauman, M.; Anjum, F.M.; Hussain, S.; Nadeem, M. A comparative study on chemical composition and antioxidant activity of ginger (Zingiber officinale) and cumin (Cuminum cyminum). J. Agr. Food Chem. 2010, 58, 8231–8237. [Google Scholar] [CrossRef]
- Politeo, O.; Jukic, M.; Milos, M. Comparison of chemical composition and antioxidant activity of glycosidically bound and free volatiles from clove (Eugenia caryophyllata Thumb.). J. Food Biochem. 2010, 34, 129–141. [Google Scholar] [CrossRef]
- Gourine, N.; Yousfi, M.; Bombarda, I.; Nadjemi, B.; Gaydou, E. Seasonal variation of chemical composition and antioxidant activity of essential oil from Pistacia atlantica Desf. leaves. J. Am. Oil Chem. Soc. 2010, 87, 157–166. [Google Scholar] [CrossRef]
- Ozkan, G.; Baydar, H.; Erbas, S. The influence of harvest time on essential oil composition, phenolic constituents and antioxidant properties of Turkish oregano (Origanum onites L.). J. Sci. Food Agr. 2010, 90, 205–209. [Google Scholar] [CrossRef]
- Singh, H.P.; Kaur, S.; Mittal, S.; Batish, D.R.; Kohli, R.K. In vitro screening of essential oil from young and mature leaves of Artemisia scoparia compared to its major constituents for free radical scavenging activity. Food Chem. Toxicol. 2010, 48, 1040–1044. [Google Scholar] [CrossRef]
- Tomić, S.; Božin, B.; Samojlik, I.; Milenković, M.; Mimica-Dukić, N.; Petrović, S. Effects of Athamanta turbith fruit essential oils on CCl4-induced hepatic failure in mice and their antioxidant properties. Phytother. Res. 2010, 24, 787–790. [Google Scholar]
- Stevenson, D.E.; Hurst, R.D. Polyphenolic phytochemicals-just antioxidants or much more? A review. Cell. Mol. Life Sci. 2007, 64, 2900–2916. [Google Scholar] [CrossRef]
- Darsham, S.; Doreswamug, R. Patented anti-inflammatory plant drug development from traditional medicine. Phytother. Res. 2004, 18, 343–357. [Google Scholar] [CrossRef]
- Santos, F.A.; Rao, V.S.N. Antiinflammatory and antinociceptive effects of 1,8-cineole a terpenoid oxide present in many plant essential oils. Phytother. Res. 2000, 14, 240–244. [Google Scholar] [CrossRef]
- Hajhashemi, V.; Ghannadi, A.; Jafarabadi, H. Black cumin seed essential oil, as a potent analgesic and anti-inflammatory drug. Phytother. Res. 2004, 18, 195–199. [Google Scholar] [CrossRef]
- Oyemitamn, I.A.; Iwalewa, E.O.; Akanmu, M.A.; Olugbade, T.A. Antinociceptive and anti-inflammatory effects of essential oil of Dennettia tripetala G. Baker (Annonaceae) in rodents. Afr. J. Trad. CAM 2008, 5, 355–362. [Google Scholar]
- Lino, C.S.; Gomes, P.B.; Lucetti, D.L.; Diógenes, J.P.L.; Sousa, F.C.F.; Silva, M.G.V.; Viana, G.S.B. Evaluation of antinociceptive and antiinflammatory activities of the essential oil (EO) of Ocimum micranthum Willd. from Northeastern Brazil. Phytother. Res. 2005, 19, 708–712. [Google Scholar] [CrossRef]
- Juhás, Š.; Bujňáková, D.; Rehák, P.; Cikoš, Š.; Czikková, S.; Veselá, J.; Il’ková, G.; Koppel, J. Anti-inflammatory effects of thyme essential oil in mice. Acta Vet. Brno. 2008, 77, 327–334. [Google Scholar] [CrossRef]
- Apel, M.A.; Lima, M.E.L.; Sobral, M.; Young, M.C.M.; Cordeiro, I.; Schapoval, E.E.S.; Henriques, A.T.; Moreno, P.R.H. Anti-inflammatory activity of essential oils from leaves of Myrciaria tenella and Calycorectes sellowianus. Pharm. Biol. 2010, 48, 433–438. [Google Scholar] [CrossRef]
- Mendes, S.S.; Bomfim, R.R.; Jesus, H.C.R.; Alves, P.B.; Blank, A.F.; Estevam, C.S.; Antoniolli, A.R.; Thomazzi, S.M. Evaluation of the analgesic and anti-inflammatory effects of the essential oil of Lippia gracilis leaves. J. Ethnopharmacol. 2010, 129, 391–397. [Google Scholar] [CrossRef]
- Işcan, G.; Kirimer, N.; Kürkçüoglu, M.; Arabaci, T.; Küpeli, E.; Başer, K.H.C. Biological activity and composition of the essential oils of Achillea schischkinii Sosn. and Achillea aleppica D.C. subsp. aleppica. J. Agr. Food Chem. 2006, 54, 170–173. [Google Scholar]
- Hajhashemi, V.; Ghannadi, A.; Sharif, B. Anti-inflammatory and analgesic properties of the leaf extracts and essential oil of Lavandula angustifolia Mill. J. Ethnopharmacol. 2003, 89, 67–71. [Google Scholar] [CrossRef]
- Murakami, A.; Ohigashi, H. Targeting NOX, iNOS and COX-2 in inflammatory cells: chemoprevention using food phytochemicals. Int. J. Cancer 2007, 121, 2357–2367. [Google Scholar] [CrossRef]
- González, S.B.; Houghton, P.J.; Hoult, J.R.S. The activity against leukocyte eicosanoid generation of essential oil and polar fractions of Adesmia boronioides Hook.f. Phytother. Res. 2003, 17, 290–293. [Google Scholar] [CrossRef]
- Kamatou, G.P.P.; van Zyl, R.L.; van Vuuren, S.F.; Viljoen, A.M.; Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G.; Tilney, P.M. Chemical composition, leaf trichome types and biological activities of the essential oils of four related Salvia species indigenous to Southern Africa. J. Essent. Oil Res. 2006, 18, 72–79. [Google Scholar]
- Lourens, A.C.U.; Reddy, D.; Başer, K.H.C.; Viljoen, A.M.; van Vuuren, S.F. In vitro biological activity and essential oil composition of four indigenous South African Helichrysum species. J. Ethnopharmacol. 2004, 95, 253–258. [Google Scholar] [CrossRef]
- Syamsir, D.R.B. Essential oils and biological activities of three selected wild Alpinia species. Master thesis, Institute of Biological Sciences, Faculty of Sciences, University of Malaya, Kuala Lumpur, Malaysia, 2009. [Google Scholar]
- the Yoon, W.J.; Kim, S.S.; Oh, T.H.; Lee, N.H.; Hyun, C.G. Torreya nucifera essential oil inhibits skin pathogen growth and lipopolysaccharide-induced inflammatory effects. Int. J. Pharmacol. 2009, 5, 37–43. [Google Scholar] [CrossRef]
- Juergens, U.R.; Stöber, M.; Schmidt-Schilling, L.; Kleuver, T.; Vetter, H. Antiinflammatory effects of eucalyptol (1,8-cineole) in bronchial asthma: inhibition of arachidonic acid metabolism in human blood monocytes ex vivo. Eur. J. Med. Res. 1998, 17, 407–412. [Google Scholar]
- Dung, N.T.; Bajpai, V.K.; Yoon, J.I.; Kang, S.C. Anti-inflammatory effects of essential oil isolated from the buds of Cleistocalyx operculatus (Roxb.) Merr and Perry. Food Chem. Toxicol. 2009, 47, 449–453. [Google Scholar] [CrossRef]
- Raetz, C.R.H.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef]
- Schröder, N.W.J.; Morath, S.; Alexander, C.; Hamann, L.; Hartung, T.; Zähringer, U.; Göbel, U.B.; Weber, J.R.; Schumann, R.R. Lipoteichoic acid (LTA) of Streptococcus pneumoniae and Staphylococcus aureus activated immune cells via Toll-like receptor (TLR)-2, lipopolysaccharide-binding protein (LBP), and CD14, whereas TLR-4 and MD-2 are not involved. J. Biol. Chem. 2003, 278, 15587–15594. [Google Scholar]
- Hart, P.H.; Brand, C.; Carson, C.F.; Riley, T.V.; Prager, R.H.; Finlay-Jones, J.J. Terpinen-4-ol, the main component of the essential oil of Melaleuca alternifolia (tea tree oil), suppresses inflammatory mediator production by activated human monocytes. Inflamm. Res. 2000, 9, 19–26. [Google Scholar]
- Caldefie-Chézet, F.; Fusillier, C.; Jarde, T.; Laroye, H.; Damez, M.; Vasson, M.-P.; Guillot, J. Potential anti-inflammatory effects of Melaleuca alternifolia essential oil on human peripheral blood leukocytes. Phytother. Res. 2006, 20, 364–370. [Google Scholar] [CrossRef]
- Hammer, K.A.; Carson, C.F.; Dunstan, J.A.; Hale, J.; Lehmann, H.; Robinson, C.J.; Prescott, S.L.; Riley, T.V. Antimicrobial and anti-inflammatory activities of Taxandria fragrans oils in vitro. Microbiol. Immunol. 2008, 52, 522–530. [Google Scholar] [CrossRef]
- Chao, L.K.; Hua, K.-F.; Hsu, H.-Y.; Cheng, S.-S.; Liu, J.-Y.; Chang, S.-T. Study of the anti-inflammatory activity of essential oil from leaves of Cinnamomum osmophloeum. J. Agr. Food Chem. 2005, 53, 7274–7278. [Google Scholar] [CrossRef]
- Juhás, Š.; Bukovská, A.; Cikoš, Š.; Czikková, S.; Fabian, D.; Koppel, J. Anti-inflammatory effects of Rosmarinus officinalis essential oil in mice. Acta Vet. Brno. 2009, 78, 121–127. [Google Scholar] [CrossRef]
- Chao, L.K.; Hua, K.-F.; Hsu, H.-Y.; Cheng, S.-S.; Lin, I.-F.; Chen, C.-J.; Chen, S.-T.; Chang, S.-T. Cinnamaldehyde inhibits pro-inflammatory cytokines secretion from monocytes/macrophages through suppression of intracellular signalling. Food Chem. Toxicol. 2008, 46, 220–231. [Google Scholar] [CrossRef]
- Passos, G.F.; Fernandes, E.S.; da Cunha, F.M.; Ferreira, J.; Pianowski, L.F.; Campos, M.M.; Calixto, J.B. Anti-inflammatory and anti-allergic properties of the essential oil and active compounds from Cordia verbenacea. J. Ethnopharmacol. 2007, 110, 323–333. [Google Scholar] [CrossRef]
- Medeiros, R.; Passos, G.F.; Vítor, C.E.; Koepp, J.; Mazzuco, T.L.; Pianowski, L.F.; Campos, M.M.; Calixto, J.B. Effect of two active compounds obtained from the essential oil of Cordia verbenacea on the acute inflammatory responses elicited by LPS in the rat paw. Brit. J. Pharmacol. 2007, 151, 618–627. [Google Scholar] [CrossRef]
- Fernandes, E.S.; Passos, G.F.; Medeiros, R.; da Cunha, F.M.; Ferreira, J.; Campos, M.M.; Pianowski, L.F.; Calixto, J.B. Anti-inflammatory effects of compounds alpha-humulene and (-)-trans-caryophyllene isolated from the essential oil of Cordia verbenacea. Eur. J. Pharmacol. 2007, 569, 228–236. [Google Scholar] [CrossRef]
- Yoon, W.J.; Kim, S.S.; Oh, T.H.; Lee, N.H.; Hyun, C.G. Cryptomeria japonica essential oil inhibits the growth of drug-resistant skin pathogens and LPS-induced NO and pro-inflammatory cytokine production. Polish J. Microbiol. 2009, 58, 61–68. [Google Scholar]
- Yoon, W.J.; Moon, J.Y.; Song, G.; Lee, Y.K.; Han, M.S.; Lee, J.S.; Ihm, B.S.; Lee, W.J.; Lee, N.H.; Hyun, C.G. Artemisia fukudo essential oil attenuates LPS-induced inflammation by suppressing NF-kB and MAPK activation in RAW264.7 macrophages. Food Chem. Toxicol. 2010, 48, 1222–1229. [Google Scholar] [CrossRef]
- Sforcin, J.M.; Amaral, J.T.; Fernandes, A., Jr.; Sousa, J.P.B.; Bastos, J.K. Lemongrass effects on IL-1β and IL-6 production by macrophages. Nat. Prod. Res. 2009, 23, 1151–1159. [Google Scholar] [CrossRef]
- Rodrigues, T.G.; Fernandes, A., Jr.; Sousa, J.P.B.; Bastos, J.K.; Sforcin, J.M. In vitro and in vivo effects of clove on pro-inflammatory cytokines production by macrophages. Nat. Prod. Res. 2009, 23, 319–326. [Google Scholar] [CrossRef]
- Lee, Y.-Y.; Hung, S.-L.; Pai, S.-F.; Lee, Y.-H.; Yang, S.-F. Eugenol suppressed the expression of lipopolysaccharide-induced proinflammatory mediators in human macrophages. J. Endodont. 2007, 33, 698–702. [Google Scholar] [CrossRef]
- Abe, S.; Maruyama, N.; Hayama, K.; Ishibashi, H.; Inoue, S.; Oshima, H.; Yamaguchi, H. Suppression of tumor necrosis factor-alpha-induced neutrophil adherence response by essential oils. Mediat. Inflamm. 2003, 12, 323–328. [Google Scholar] [CrossRef]
- Lin, C.-T.; Chen, C.-J.; Lin, T.-Y.; Tung, J.C.; Wang, S.-Y. Anti-inflammation activity of fruit essential oil from Cinnamomum insularimontanum Hayata. Bioresource Technol. 2008, 99, 8783–8787. [Google Scholar] [CrossRef]
- Morita, T.; Jinno, K.; Kawagishi, H.; Arimoto, Y.; Suganuma, H.; Inakuma, T.; Sugiyama, K. Hepatoprotective effect of myristicin from nutmeg (Myristica fragrans) on lipopolysaccharide/D-galactosamine-induced liver injury. J. Agr. Food Chem. 2003, 51, 1560–1565. [Google Scholar] [CrossRef]
- Dutra, R.C.; Fava, M.B.; Alves, C.S.C.; Ferreira, A.P.; Barbosa, N.R. Antiulcerogenic and anti-inflammatory activities of the essential oil from Pterodon emarginatus seeds. J. Pharm. Pharmacol. 2009, 61, 243–250. [Google Scholar]
- Burkovská, A.; Čikoš, Š.; Juhás, Š.; Il’Ková, G.; Rehák, P.; Koppel, J. Effects of a combination of thyme and oregano essential oils on TNBS-induced colitis in mice. Mediat. Inflamm 2007. doi: 10:1155/2007/23296. [Google Scholar]
- Juhás, Š.; Bujňáková, D.; Rehák, P.; Cikoš, Š.; Czikková, S.; Veselá, J.; Il’ková, G.; Koppel, J. Anti-inflammatory effects of thyme essential oil in mice. Acta Vet. Brno 2008, 77, 327–334. [Google Scholar] [CrossRef]
- Inoue, H.; Yokoyama, C.; Hara, S.; Tone, Y.; Tanabe, T. Transcriptional regulation of human prostaglandin-endoperoxide synthase-2 gene by lipopolysaccharide and phorbol ester in vascular endotelial cells. J. Biol. Chem. 1995, 270, 24965–24971. [Google Scholar] [CrossRef]
- Yoshimura, A. Signal transduction of inflammatory cytokines and tumor development. Cancer Sci. 2006, 97, 439–447. [Google Scholar] [CrossRef]
- Janssen-Heininger, Y.M.W.; Poynter, M.E.; Baeuerle, P.A. Recent advances towards understanding redox mechanisms in the activation of nuclear factor κB. Free Radic. Biol. Med. 2000, 28, 1317–1327. [Google Scholar] [CrossRef]
- Wadsworth, T.L.; Koop, D.R. Effects of Ginkgo biloba extracts and quercetin on lipopolysaccharide-induced release of nitric oxide. Chem. Biol. Interact. 2001, 137, 43–58. [Google Scholar] [CrossRef]
- Winstein, S.L.; Sanghera, J.C.; Lemke, K.; De Franco, A.L.; Pelech, S.L. Bacterial lipopolysaccacharide induces tyrosine phosphorilation and activation of mitogen-activated protein kinases in macrophages. J. Biol. Chem. 1992, 267, 14955–14962. [Google Scholar]
- De-Xing, H.; Yanagita, T.; Uto, T.; Masuzaki, S.; Fujii, M. Anthocyanidins inhibit cyclooxygenase-2 expression in LPS-evoked macrophages: structure-activity relationship and molecular mechanisms involved. Biochem. Pharmacol. 2005, 70, 417–425. [Google Scholar] [CrossRef]
- Hotta, M.; Nakata, R.; Kasukawa, M.; Hori, K.; Takahashi, S.; Inoue, H. Carvacrol, a component of thyme oil, activates PPARα and γ and suppresses COX-2 expressio. J. Lipid Res. 2010, 51, 132–139. [Google Scholar] [CrossRef]
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schütz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P.; Evans, R.M. The nuclear receptor superfamily: the second decade. Cell 1995, 83, 835–839. [Google Scholar] [CrossRef]
- Stienstra, R. The role of PPARs in inflammation and obesity. PhD thesis, Wageningen University, The Netherlands, 2007. [Google Scholar]
- Sjöholm, A.; Nystrom, T. Endothelial inflammation in insulin resistance. Lancet. 2005, 365, 610–612. [Google Scholar]
- Inoue, H.; Tanabe, T.; Unesono, K. Feedback control of cyclooxygenase-2 expression through PPARγ. J. Biol. Chem. 2000, 275, 28028–28032. [Google Scholar]
- Menichini, F.; Conforti, F.; Rigano, D.; Formisano, C.; Piozzi, F.; Senatore, F. Phytochemical composition, anti-inflammatory and antitumour activities of four Teucrium essential oils from Greece. Food Chem. 2009, 11, 670–686. [Google Scholar]
- Yang, E.-J.; Kim, S.S.; Moon, J.-Y.; Oh, T.-H.; Baik, J.S.; Lee, N.H.; Hyun, C.-G. Inhibitory effects of Fortunella japonica var. margarita and Citrus sunki essential oils on nitric oxide production and skin pathogens. Acta Microbiol. Immunol. Hung. 2010, 57, 15–27. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Menichini, F.; Conforti, F.; Tundis, R.; Bonesi, M.; Saab, A.M.; Statti, G.A.; Cindio, B.; Houghton, P.J.; Menichini, F.; Frega, N.G. Chemical analysis, antioxidant, antiiflammatory and anticholinesterase activities of Origanum ehrenbergii Boiss. and Origanum syriacum L. essential oils. Food Chem. 2009, 117, 174–180. [Google Scholar] [CrossRef]
- Yang, E.-J.; Kim, S.-S.; Oh, T.-H.; Baik, J.S.; Lee, N.H.; Hyun, C.-G. Essential oil of citrus fruit waste attenuates LPS-induced nitric oxide production and inhibits the growth of skin pathogens. Int. J. Agr. Biol. 2009, 11, 791–794. [Google Scholar]
- Tavares, A.C.; Gonçalves, M.J.; Cruz, M.T.; Cavaleiro, C.; Lopes, M.C.; Canhoto, J.; Salgueiro, L.R. Essential oils from Distichoselinum tenuifolium: chemical composition, cytotoxicity, antifungal and anti-inflammatory properties. J. Ethnopharmacol. 2010, 130, 593–598. [Google Scholar] [CrossRef]
- Paena, A.T.; Marzocco, S.; Popolo, A.; Pinto, A. (-)-Linalool inhibits in vitro NO formation. Probable involvement in the antinociceptive activity of this monoterpene compound. Life Sci. 2006, 78, 719–723. [Google Scholar] [CrossRef]
- Yoon, W.-J.; Kim, S.-S.; Oh, T.-H.; Lee, N.H.; Hyun, C.-G. Abies koreana essential oil inhibits drug-resistant skin pathogen growth and LPS-induced inflammatory effects of murine macrophage. Lipids 2009, 44, 471–476. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Oh, T.-H.; Kim, B.J.; Kim, S.-S.; Lee, N.H.; Hyun, C.-G. Chemical composition and anti-inflammatory effects of essential oil from Farfagium japonicum flower. J. Oleo Sci. 2008, 57, 623–628. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Kim, S.-S.; Oh, T.-H.; Baik, J.S.; Song, G.; Lee, N.H.; Hyun, C.-G. Chemical composition, antioxidant, anti-elastase, and anti-inflammatory activities of Illicium anisatum essential oil. Acta Pharm. 2009, 59, 289–300. [Google Scholar]
- Tabanca, N.; Ma, G.; Pasco, D.S.; Bedir, E.; Kiriner, N.; Husnu, K.; Baser, C.; Khan, I.A.; Khan, S.I. Effect of essential oils and isolated compounds from Pimpinella species on NF-kB: a target for anti-inflammatory therapy. Phytother. Res. 2007, 21, 741–745. [Google Scholar] [CrossRef]
- Katsukawa, M.; Nakata, R.; Takizawa, Y.; Hori, K.; Takahashi, S.; Inoue, H. Citral, a component of lemongrass oil, activates PPARa and g and suppresses COX-2 expression. BBA-Mol. Cell Biol. Lipids 2010, 1801, 1214–1220. [Google Scholar] [CrossRef]
- Chainy, G.B.N.; Manna, S.K.; Chaturvedi, M.M.; Aggarwal, B.B. Anethole blocks both early and late cellular responses transduced by tumor necrosis factor: effect on NF-kB, AP-1 JNK, MAPKK and apopotosis. Oncogene 2000, 19, 2943–2950. [Google Scholar] [CrossRef]
- Zhou, J.-Y.; Wang, X.-F.; Tang, F.-D.; Zhou, J.-Y.; Lu, G.-H.; Wang, Y.; Bian, R.-L. Inhibitory effect of 1,8-cineole (eucalyptol) on Erg-1 expression in lipopolysaccharide-stimulated THP-1 cells. Acta Pharmacol. Sin. 2007, 28, 908–912. [Google Scholar] [CrossRef]
- Zhou, J.-Y.; Tang, F.-D.; Mao, G.G.; Bian, R.-L. Effect of α-pinene on nuclear translocation of NF-κB in THP-1 cells. Acta Pharmacol. Sin. 2004, 25, 480–484. [Google Scholar]
- Salminen, A.; Lehtonen, M.; Suuronen, T.; Kaarniranta, K.; Huuskonen, J. Terpenoids: natural inhibitors of NF-κB signalling with anti-inflammatory and anticancer potential. Cell. Mol. Life Sci. 2008, 65, 2979–2999. [Google Scholar] [CrossRef]
- Neves, A.; Rosa, S.; Gonçalves, J.; Rufino, A.; Judas, F.; Salgueiro, L.; Lopes, M.C.; Cavaleiro, C.; Mendes, A.F. Screening of five essential oils for identification of potential inhibitors of IL-1-induced NF-κB activation and NO production in human chondrocytes: characterization of the inhibitory activity of α-pinene. Planta Med. 2010, 76, 303–308. [Google Scholar] [CrossRef]
- Cabrera, A.C.; Prieto, J.M. Application of artificial neural networks to the prediction of the antioxidant activity of essential oils in two experimental in vitro models. Food Chem. 2010, 118, 141–146. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Miguel, M.G. Antioxidant and Anti-Inflammatory Activities of Essential Oils: A Short Review. Molecules 2010, 15, 9252-9287. https://doi.org/10.3390/molecules15129252
Miguel MG. Antioxidant and Anti-Inflammatory Activities of Essential Oils: A Short Review. Molecules. 2010; 15(12):9252-9287. https://doi.org/10.3390/molecules15129252
Chicago/Turabian StyleMiguel, Maria Graça. 2010. "Antioxidant and Anti-Inflammatory Activities of Essential Oils: A Short Review" Molecules 15, no. 12: 9252-9287. https://doi.org/10.3390/molecules15129252
APA StyleMiguel, M. G. (2010). Antioxidant and Anti-Inflammatory Activities of Essential Oils: A Short Review. Molecules, 15(12), 9252-9287. https://doi.org/10.3390/molecules15129252