Vitamin B6: A Molecule for Human Health?

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Suggested Reaction Mechanisms of VitB6 for Amino Acid Metabolism
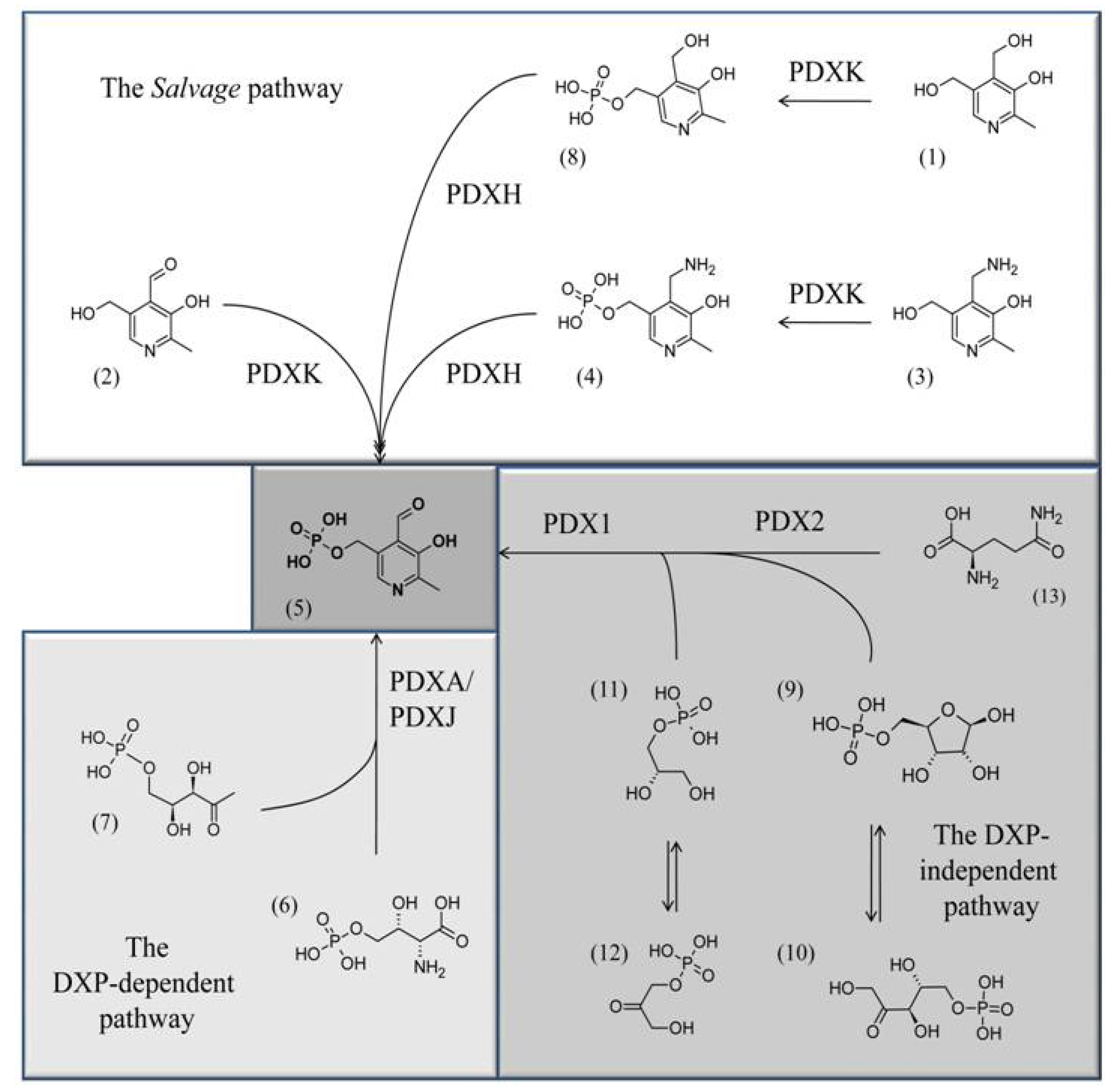
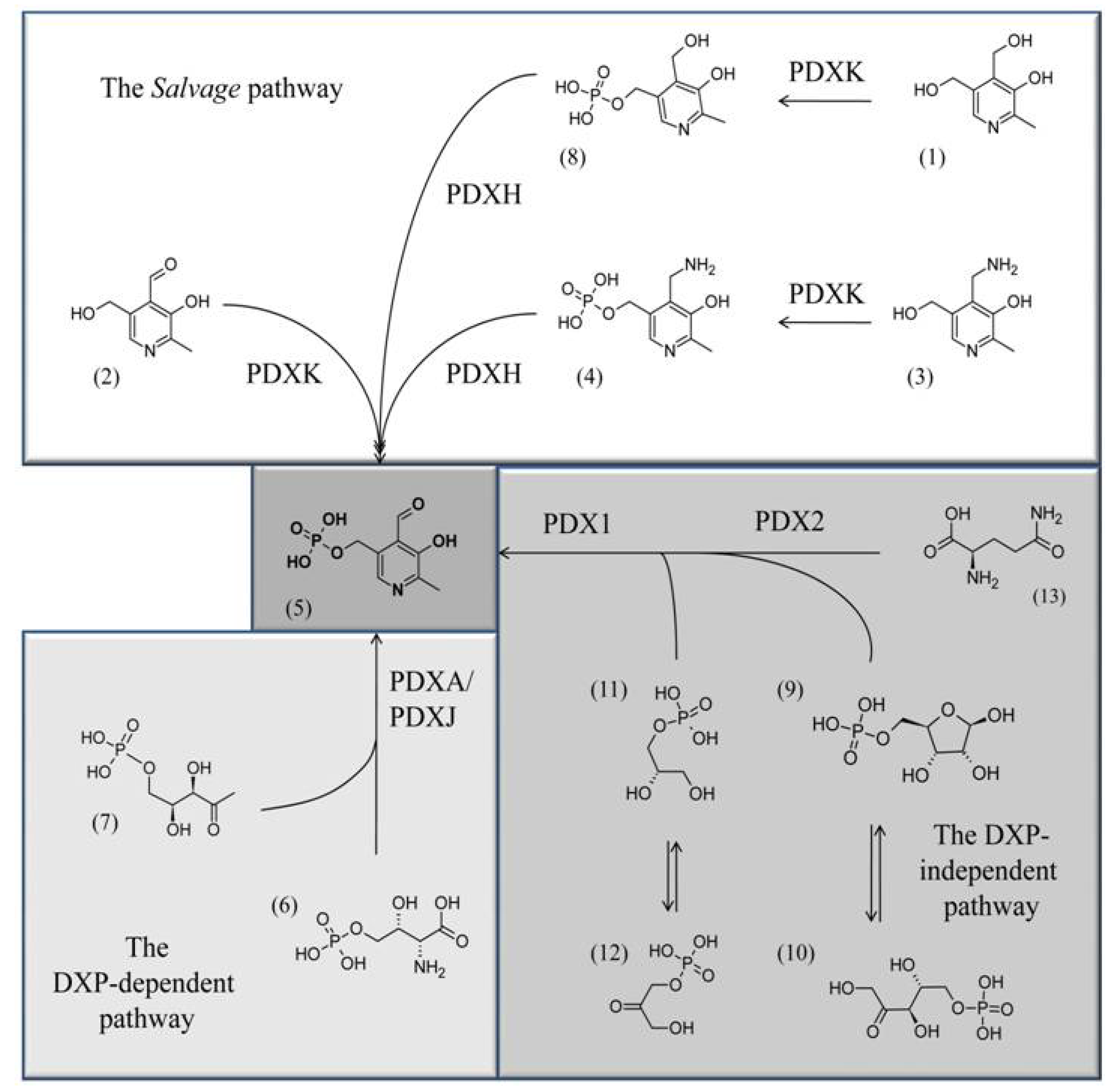
3. Three Different Biosynthetic Pathways for VitB6 Are Known
4. VitB6 and Its Healthy Face
4.1. Therapeutic applications by using drugs against PLP-dependent enzymes
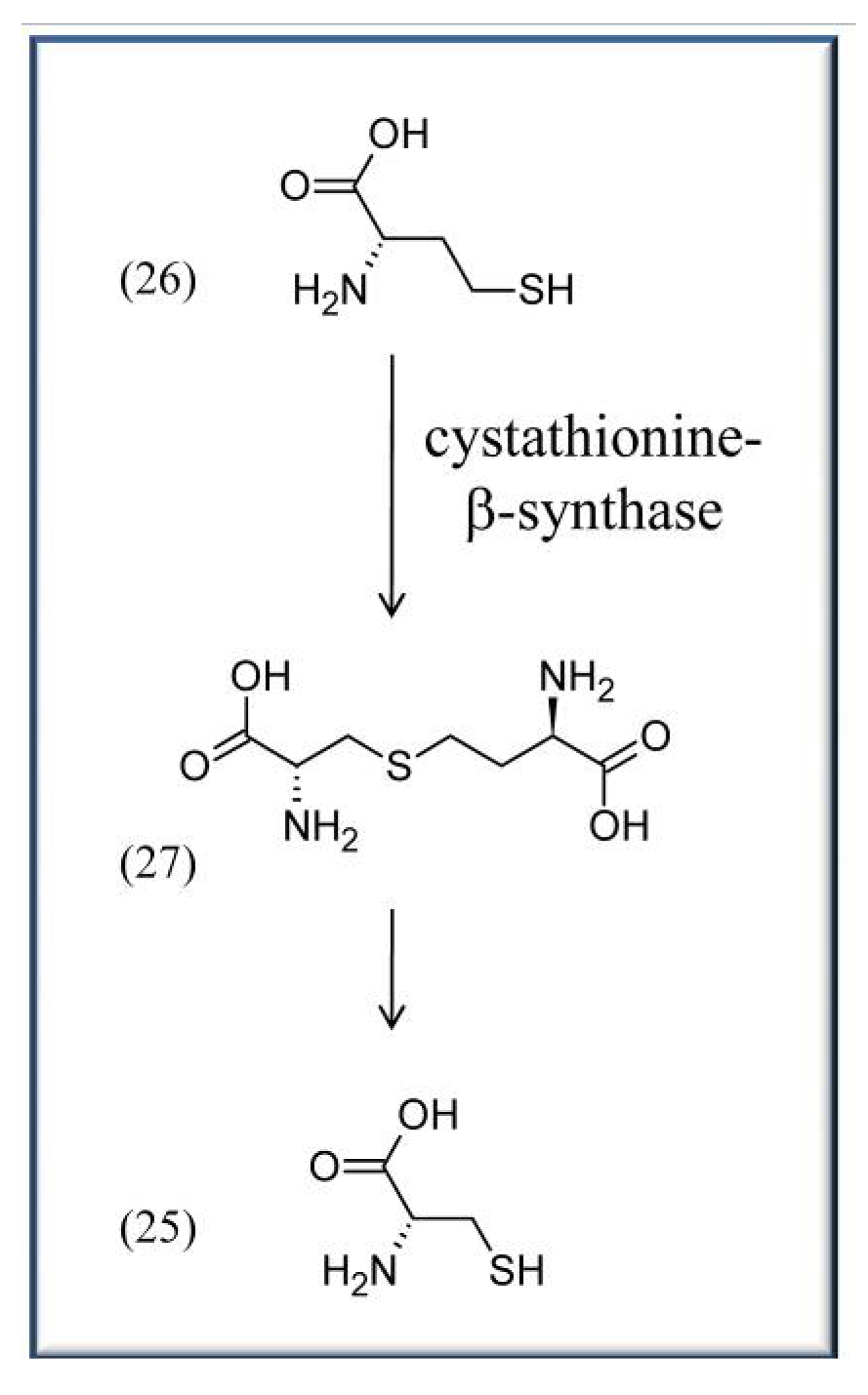
4.2. VitB6 in context with cardiovascular disease and blood pressure
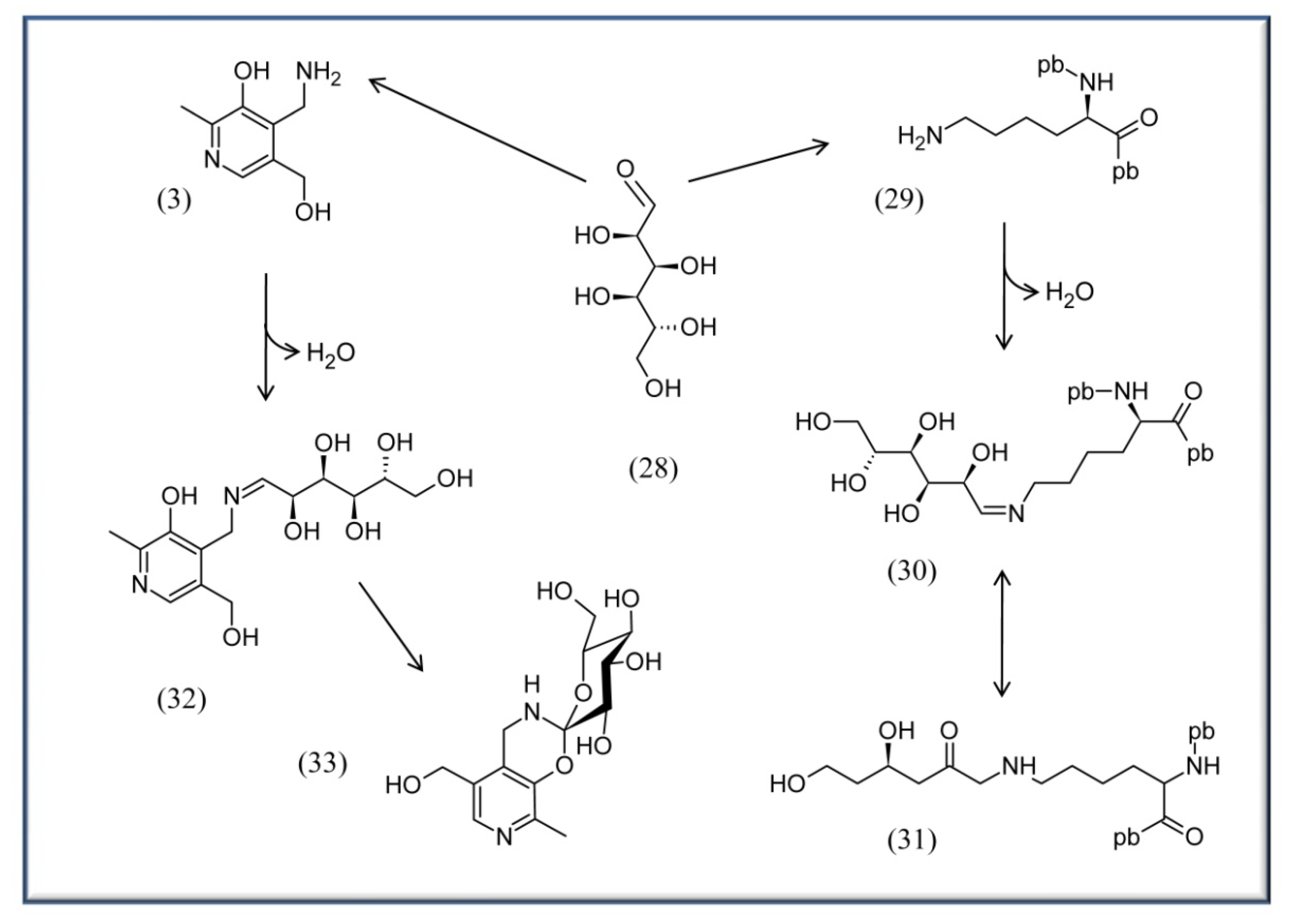
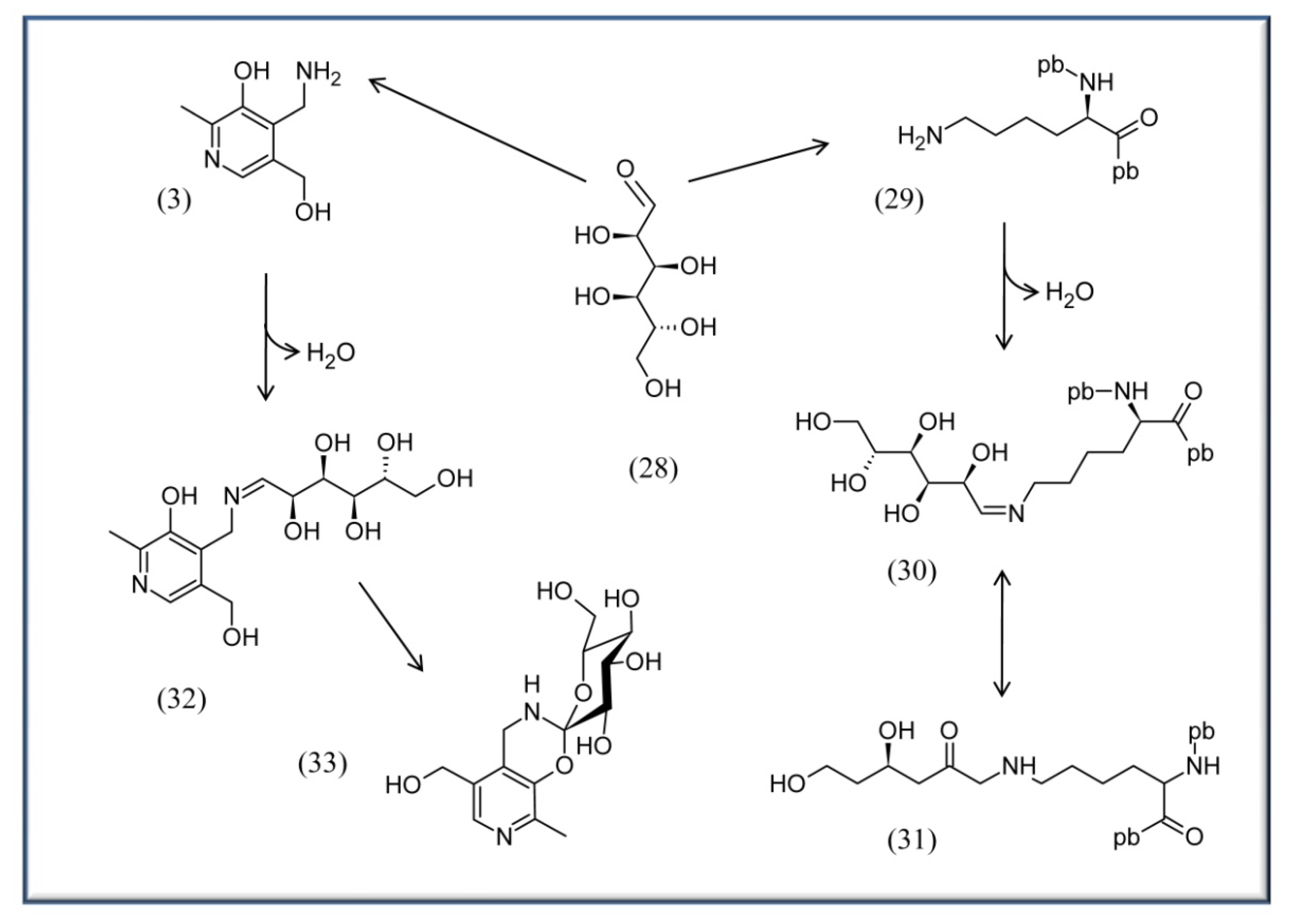
4.3. VitB6 in context with diabetes, AGE and ALE
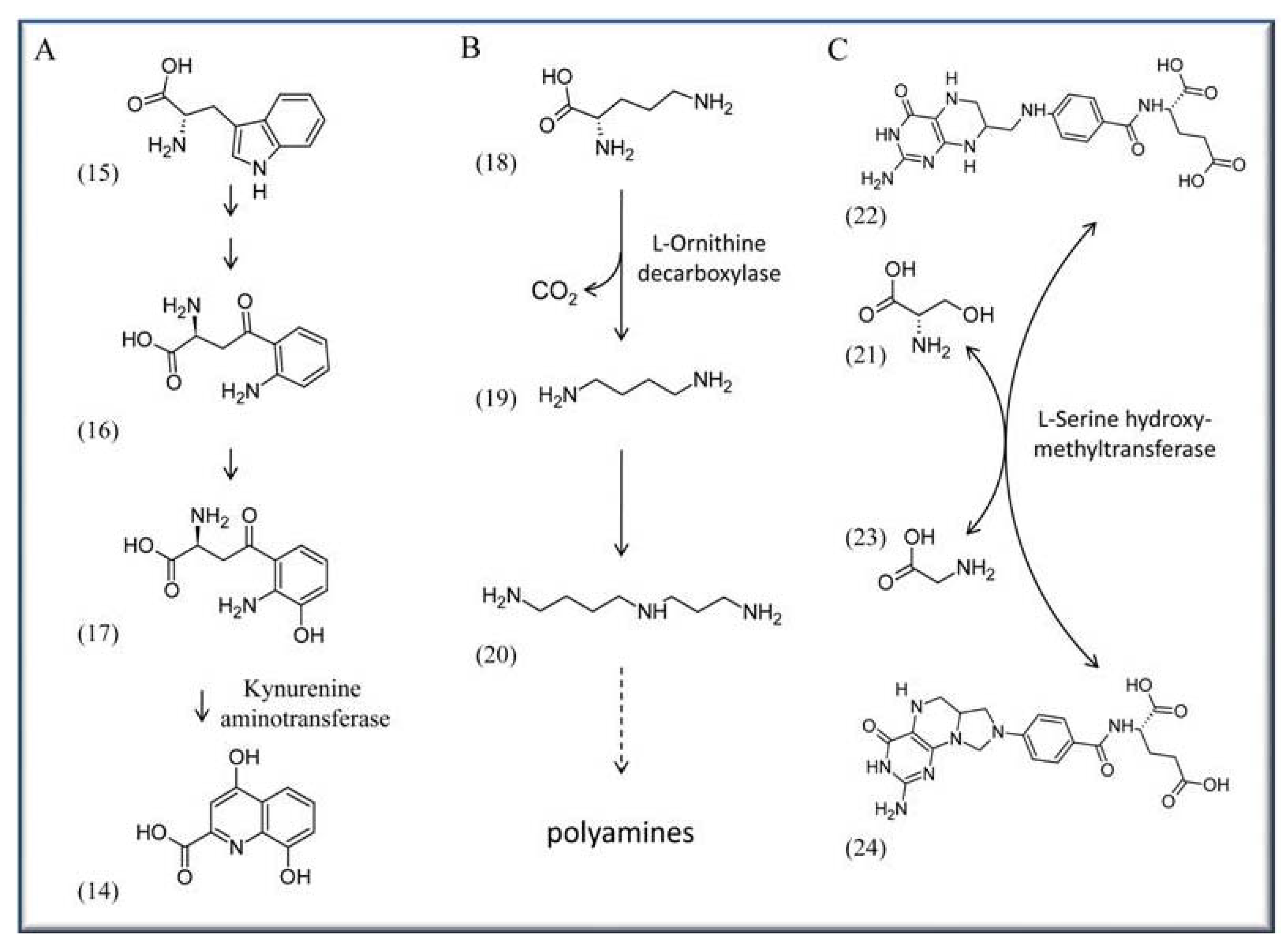
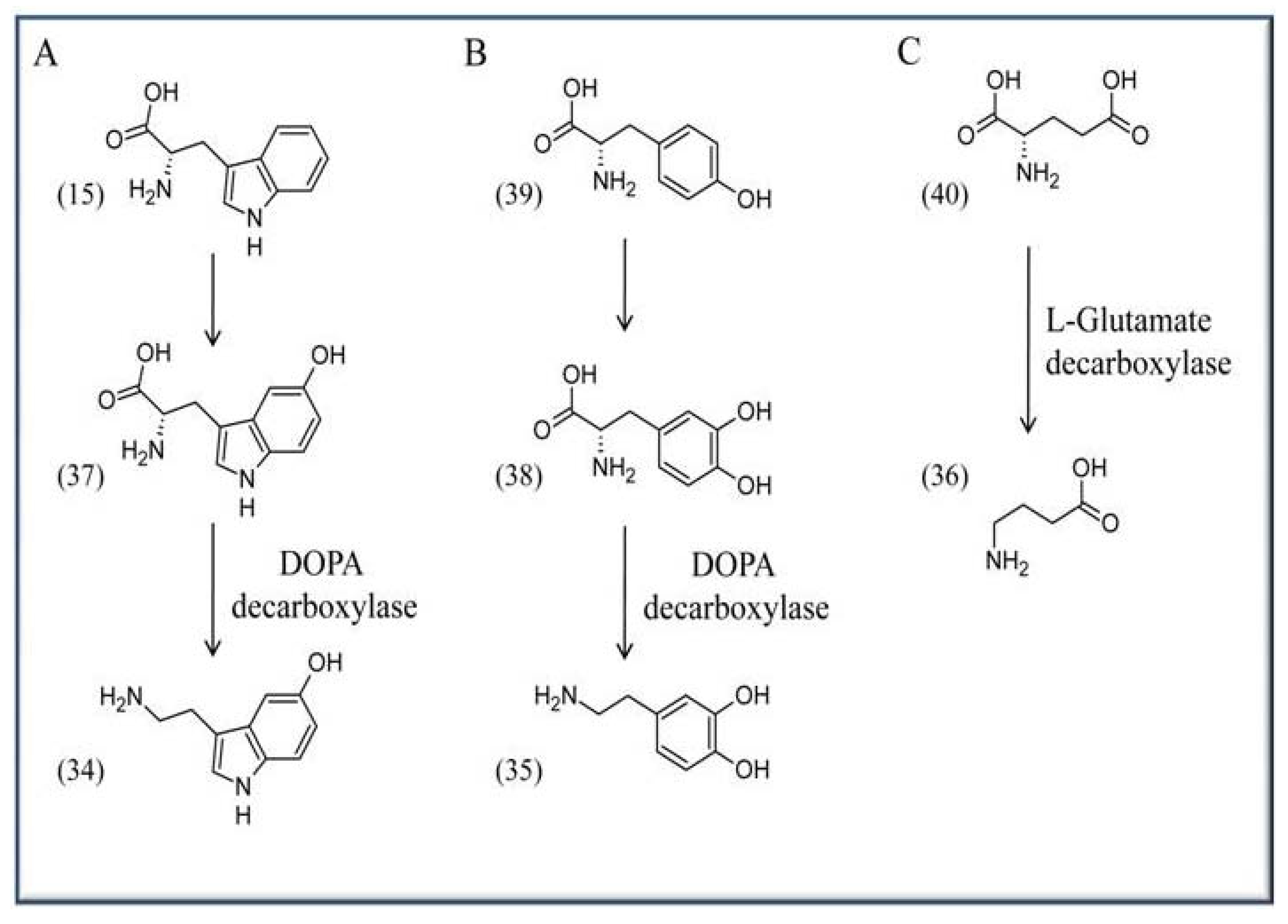
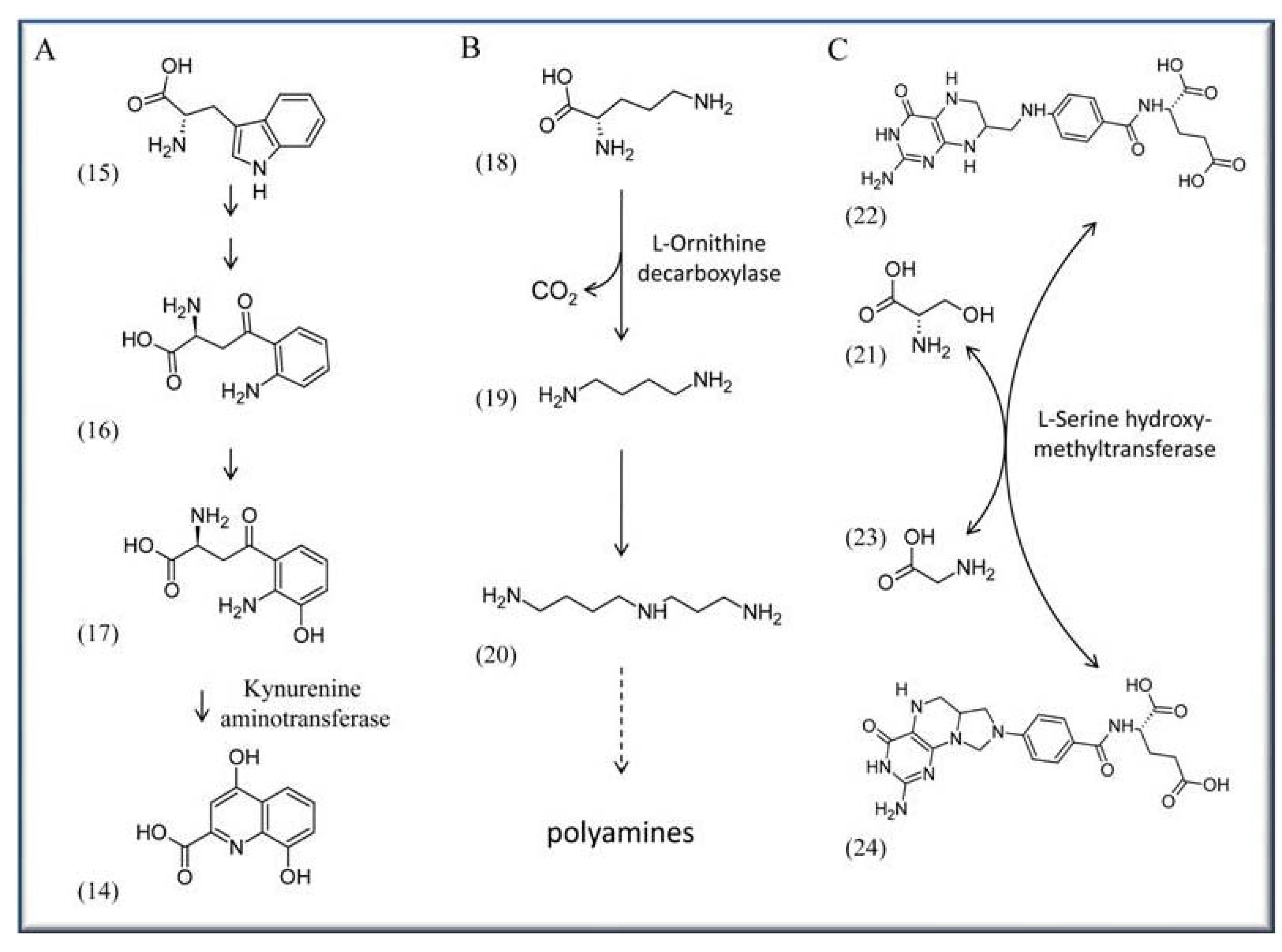
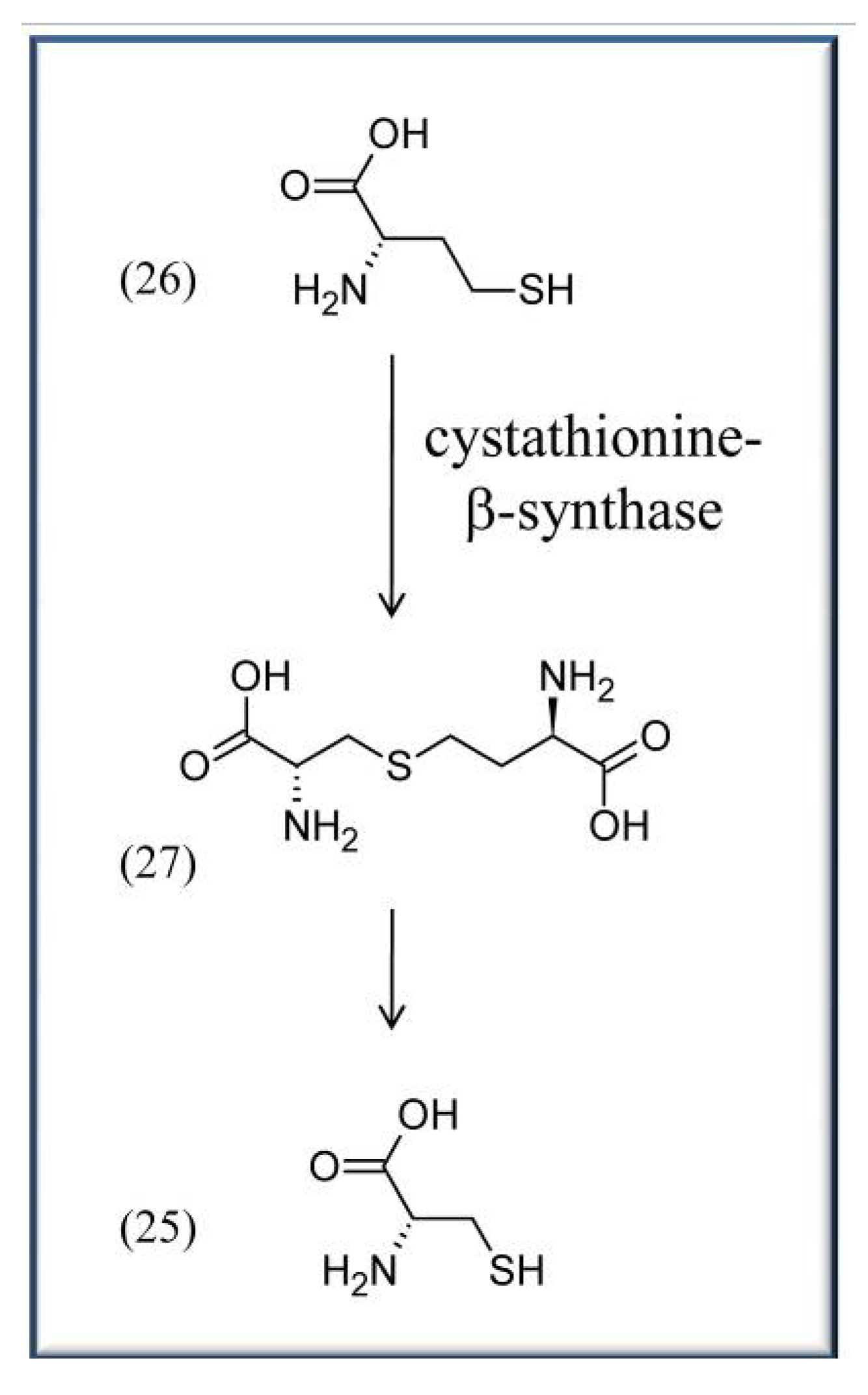
4.4. VitB6 in context with neurological activity
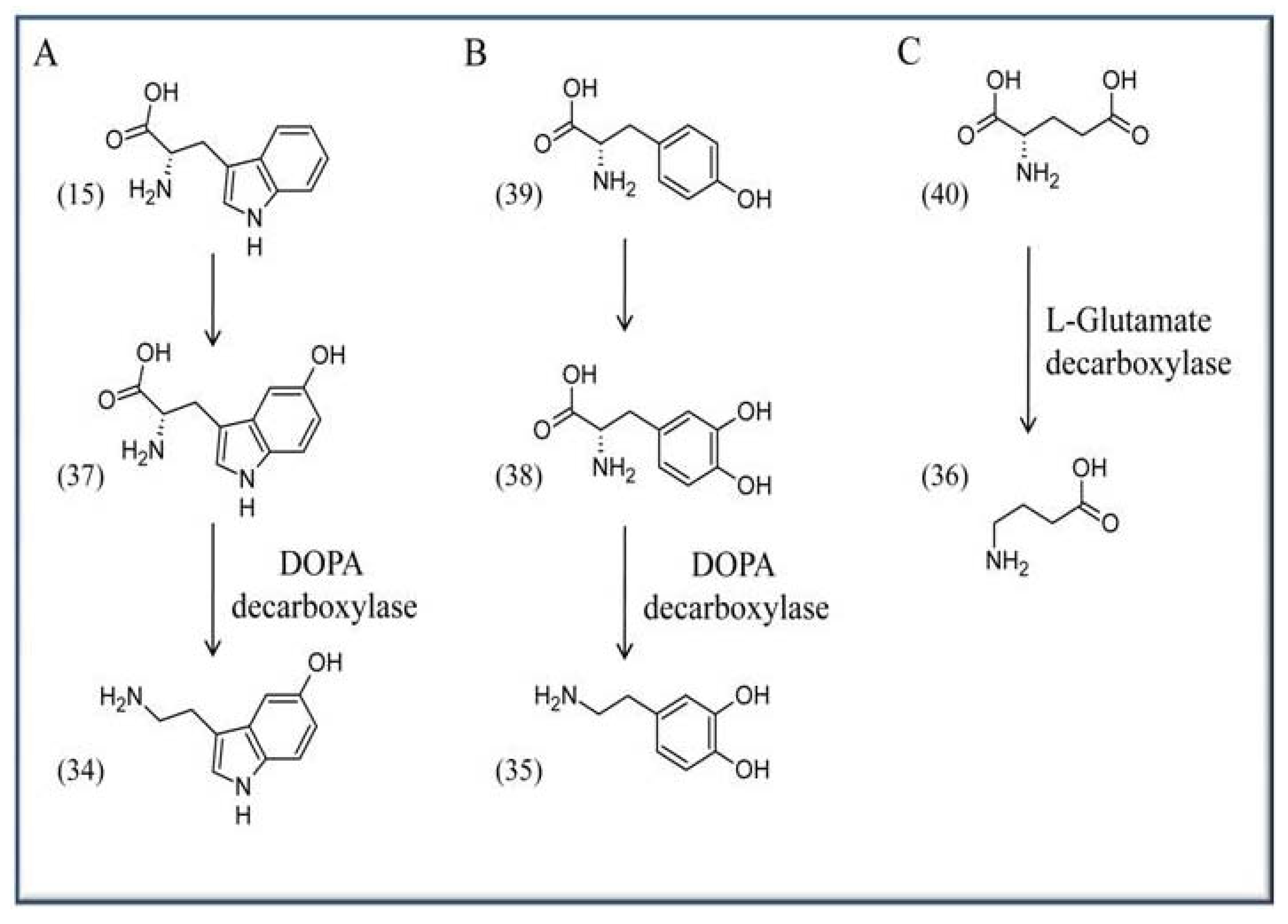
4.5. The antioxidative capacity of VitB6
5. Conclusions
Acknowledgments
- Samples Availability: Not applicable.
References and Notes
- Roje, S. Vitamin B biosynthesis in plants. Phytochemistry 2007, 68, 1904–1921. [Google Scholar] [CrossRef]
- Birch, T.W.; Gyorgy, P.; Harris, L.J. The vitamin B(2) complex. Differentiation of the antiblacktongue and the "P.-P." factors from lactoflavin and vitamin B(6) (so-called "rat pellagra" factor). Parts I-VI. Biochem. J. 1935, 29, 2830–2850. [Google Scholar]
- Percudani, R.; Peracchi, A. The B6 database: a tool for the description and classification of vitamin B6-dependent enzymatic activities and of the corresponding protein families. BMC Bioinformatics 2009, 10, 273. [Google Scholar] [CrossRef]
- Brouwer, D.A.; Welten, H.T.; van Doormaal, J.J.; Reijngoud, D.J.; Muskiet, F.A. Recommended dietary allowance of folic acid is insufficient for optimal homocysteine levels. Ned. Tijdschr. Geneeskd. 1998, 142, 782–786. [Google Scholar]
- Amadasi, A.; Bertoldi, M.; Contestabile, R.; Bettati, S.; Cellini, B.; di Salvo, M.L.; Borri-Voltattorni, C.; Bossa, F.; Mozzarelli, A. Pyridoxal 5'-phosphate enzymes as targets for therapeutic agents. Curr. Med. Chem. 2007, 14, 1291–1324. [Google Scholar] [CrossRef]
- Di Giovine, P. PLPMDB: pyridoxal-5'-phosphate dependent enzymes mutants database. Bioinformatics 2004, 20, 3652–3653. [Google Scholar] [CrossRef]
- Denesyuk, A.I.; Denessiouk, K.A.; Korpela, T.; Johnson, M.S. Functional attributes of the phosphate group binding cup of pyridoxal phosphate-dependent enzymes. J. Mol. Biol. 2002, 316, 155–172. [Google Scholar] [CrossRef]
- Mooney, S.; Leuendorf, J.E.; Hendrickson, C.; Hellmann, H. Vitamin B6: a long known compound of surprising complexity. Molecules 2009, 14, 329–351. [Google Scholar] [CrossRef]
- Fitzpatrick, T.B.; Amrhein, N.; Kappes, B.; Macheroux, P.; Tews, I.; Raschle, T. Two independent routes of de novo vitamin B6 biosynthesis: not that different after all. Biochem. J. 2007, 407, 1–13. [Google Scholar] [CrossRef]
- Drewke, C.; Leistner, E. Biosynthesis of vitamin B6 and structurally related derivatives. Vitam. Horm. 2001, 61, 121–155. [Google Scholar] [CrossRef]
- Drewke, C.; Klein, M.; Clade, D.; Arenz, A.; Muller, R.; Leistner, E. 4-O-phosphoryl-L-threonine, a substrate of the pdxC(serC) gene product involved in vitamin B6 biosynthesis. FEBS Lett. 1996, 390, 179–182. [Google Scholar] [CrossRef]
- Notheis, C.; Drewke, C.; Leistner, E. Purification and characterization of the pyridoxol-5'-phosphate:oxygen oxidoreductase (deaminating) from Escherichia coli. Biochim. Biophys. Acta 1247, 265–271. [Google Scholar]
- Mittenhuber, G. Phylogenetic analyses and comparative genomics of vitamin B6 (pyridoxine) and pyridoxal phosphate biosynthesis pathways. J. Mol. Microbiol. Biotechnol. 2001, 3, 1–20. [Google Scholar]
- Tambasco-Studart, M.; Titiz, O.; Raschle, T.; Forster, G.; Amrhein, N.; Fitzpatrick, T.B. Vitamin B6 biosynthesis in higher plants. Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 13687–13692. [Google Scholar]
- Burns, K.E.; Xiang, Y.; Kinsland, C.L.; McLafferty, F.W.; Begley, T.P. Reconstitution and biochemical characterization of a new pyridoxal-5'-phosphate biosynthetic pathway. J. Am. Chem. Soc. 2005, 127, 3682–3683. [Google Scholar]
- Tanaka, T.; Tateno, Y.; Gojobori, T. Evolution of vitamin B6 (pyridoxine) metabolism by gain and loss of genes. Mol. Biol. Evol. 2005, 22, 243–2450. [Google Scholar]
- Wagner, S.; Bernhardt, A.; Leuendorf, J. E.; Drewke, C.; Lytovchenko, A.; Mujahed, N.; Gurgui, C.; Frommer, W.B.; Leistner, E.; Fernie, A.R.; Hellmann, H. Analysis of the Arabidopsis rsr4-1/pdx1-3 mutant reveals the critical function of the PDX1 protein family in metabolism, development, and vitamin B6 biosynthesis. Plant Cell 2006, 18, 1722–1735. [Google Scholar] [CrossRef]
- Neuwirth, M.; Strohmeier, M.; Windeisen, V.; Wallner, S.; Deller, S.; Rippe, K.; Sinning, I.; Macheroux, P.; Tews, I. X-ray crystal structure of Saccharomyces cerevisiae Pdx1 provides insights into the oligomeric nature of PLP synthases. FEBS Lett. 2009, 583, 2179–2186. [Google Scholar] [CrossRef]
- Raschle, T.; Speziga, D.; Kress, W.; Moccand, C.; Gehrig, P.; Amrhein, N.; Weber-Ban, E.; Fitzpatrick, T.B. Intersubunit crosstalk in pyridoxal 5'-phosphate synthase, co-ordinated by the C-terminus of the synthase subunit. J. Biol. Chem. 2008, 284, 7706–7718. [Google Scholar]
- Gdynia, H.J.; Muller, T.; Sperfeld, A.D.; Kuhnlein, P.; Otto, M.; Kassubek, J.; Ludolph, A.C. Severe sensorimotor neuropathy after intake of highest dosages of vitamin B6. Neuromuscul. Disord. 2008, 18, 156–158. [Google Scholar] [CrossRef]
- Neuwirth, M.; Flicker, K.; Strohmeier, M.; Tews, I.; Macheroux, P. Thermodynamic characterization of the protein-protein interaction in the heteromeric Bacillus subtilis pyridoxalphosphate synthase. Biochemistry 2007, 46, 5131–5139. [Google Scholar] [CrossRef]
- Strohmeier, M.; Raschle, T.; Mazurkiewicz, J.; Rippe, K.; Sinning, I.; Fitzpatrick, T.B.; Tews, I. Structure of a bacterial pyridoxal 5'-phosphate synthase complex. Proc. Natl. Acad. Sci. USA 2006, 103, 19284–19289. [Google Scholar]
- Gengenbacher, M.; Fitzpatrick, T.B.; Raschle, T.; Flicker, K.; Sinning, I.; Muller, S.; Macheroux, P.; Tews, I.; Kappes, B. Vitamin B6 biosynthesis by the malaria parasite Plasmodium falciparum: biochemical and structural insights. J. Biol. Chem. 2006, 281, 3633–3641. [Google Scholar]
- Zein, F.; Zhang, Y.; Kang, Y.N.; Burns, K.; Begley, T.P.; Ealick, S.E. Structural insights into the mechanism of the PLP synthase holoenzyme from Thermotoga maritima. Biochemistry 2006, 45, 14609–14620. [Google Scholar]
- Gonzalez, E.; Danehower, D.; Daub, M.E. Vitamer levels, stress response, enzyme activity, and gene regulation of Arabidopsis lines mutant in the pyridoxine/pyridoxamine 5'-phosphate oxidase (PDX3) and the pyridoxal kinase (SOS4) genes involved in the vitamin B6 salvage pathway. Plant Physiol. 2007, 145, 985–996. [Google Scholar] [CrossRef]
- Ohdake, S. Isolation of "Oryzanin" (Antineuritic Vitamin) from Rice-polishings. Bull Agri. Chem. Soc. Jpn. 1932, 8, 11–46. [Google Scholar] [CrossRef]
- Oka, T. Vitamin B6. Nippon Rinsho 1999, 57, 2199–2204. [Google Scholar]
- Nogueira, A.; Duarte, A.F.; Magina, S.; Azevedo, F. Pellagra associated with esophageal carcinoma and alcoholism. Dermatol. Online J. 2009, 15, 8. [Google Scholar]
- Hendricks, W.M. Pellagra and pellagralike dermatoses: etiology, differential diagnosis, dermatopathology, and treatment. Semin. Dermatol. 1991, 10, 282–292. [Google Scholar]
- Morris, M.S.; Picciano, M.F.; Jacques, P.F.; Selhub, J. Plasma pyridoxal 5'-phosphate in the US population: the National Health and Nutrition Examination Survey, 2003–2004. Am. J. Clin. Nutr. 2008, 87, 1446–1454. [Google Scholar]
- Perry, T.; Holloway, H.W.; Weerasuriya, A.; Mouton, P.R.; Duffy, K.; Mattison, J.A.; Greig, N.H. Evidence of GLP-1-mediated neuroprotection in an animal model of pyridoxine-induced peripheral sensory neuropathy. Exp. Neurol. 2007, 203, 293–301. [Google Scholar] [CrossRef]
- Albin, R.L.; Albers, J.W.; Greenberg, H.S.; Townsend, J.B.; Lynn, R.B.; Burke, J.M. Jr.; Alessi, A.G. Acute sensory neuropathy-neuronopathy from pyridoxine overdose. Neurology 1987, 37, 1729–1732. [Google Scholar] [CrossRef]
- Lu, T.; Xu, Y.; Monttinen, E.S.; Kato, N. Supplementing vitamin B6 to a low vitamin B6 diet exaggerates UVB-induced skin tumorigenesis in DMBA-treated hairless mice. J. Nutr. Sci. Vitaminol. (Tokyo) 2008, 54, 262–265. [Google Scholar]
- Wondrak, G.T.; Roberts, M.J.; Jacobson, M.K.; Jacobson, E.L. 3-hydroxypyridine chromophores are endogenous sensitizers of photooxidative stress in human skin cells. J. Biol. Chem. 2004, 279, 30009–30020. [Google Scholar]
- Maeda, T.; Taguchi, H.; Minami, H.; Sato, K.; Shiga, T.; Kosaka, H.; Yoshikawa, K. Vitamin B6 phototoxicity induced by UVA radiation. Arch. Dermatol. Res. 2000, 292, 562–567. [Google Scholar] [CrossRef]
- Muhia, D.K.; Swales, C.A.; Deng, W.; Kelly, J.M.; Baker, D.A. The gametocyte-activating factor xanthurenic acid stimulates an increase in membrane-associated guanylyl cyclase activity in the human malaria parasite Plasmodium falciparum. Mol. Microbiol. 2001, 42, 553–560. [Google Scholar] [CrossRef]
- Rossi, F.; Garavaglia, S.; Giovenzana, G.B.; Arca, B.; Li, J.; Rizzi, M. Crystal structure of the Anopheles gambiae 3-hydroxykynurenine transaminase. Proc. Natl. Acad. Sci. USA 2006, 103, 5711–576. [Google Scholar]
- Rossi, F.; Lombardo, F.; Paglino, A.; Cassani, C.; Miglio, G.; Arca, B.; Rizzi, M. Identification and biochemical characterization of the Anopheles gambiae 3-hydroxykynurenine transaminase. Febs.J. 2005, 272, 5653–5662. [Google Scholar] [CrossRef]
- Marti-Carvajal, A.J.; Sola, I.; Lathyris, D.; Salanti, G. Homocysteine lowering interventions for preventing cardiovascular events. Cochrane Database Syst. Rev. 2009, CD006612. [Google Scholar]
- Muller, I.B.; Wu, F.; Bergmann, B.; Knockel, J.; Walter, R.D.; Gehring, H.; Wrenger, C. Poisoning pyridoxal 5-phosphate-dependent enzymes: a new strategy to target the malaria parasite Plasmodium falciparum. PLoS One 2009, 4, 4406. [Google Scholar]
- Wrenger, C.; Eschbach, M.L.; Muller, I.B.; Warnecke, D.; Walter, R.D. Analysis of the vitamin B6 biosynthesis pathway in the human malaria parasite Plasmodium falciparum. J. Biol. Chem. 2005, 280, 5242–5248. [Google Scholar]
- Pegg, A.E.; McGovern, K.A.; Wiest, L. Decarboxylation of alpha-difluoromethylornithine by ornithine decarboxylase. Biochem. J. 1987, 241, 305–307. [Google Scholar]
- Poulin, R.; Lu, L.; Ackermann, B.; Bey, P.; Pegg, A.E. Mechanism of the irreversible inactivation of mouse ornithine decarboxylase by alpha-difluoromethylornithine. Characterization of sequences at the inhibitor and coenzyme binding sites. J. Biol. Chem. 1992, 267, 15015–15018. [Google Scholar]
- Kern, A.D.; Oliveira, M.A.; Coffino, P.; Hackert, M.L. Structure of mammalian ornithine decarboxylase at 1.6 A resolution: stereochemical implications of PLP-dependent amino acid decarboxylases. Structure 1999, 7, 567–581. [Google Scholar]
- Kernebeck, T.; Lohse, A.W.; Grotzinger, J. A bioinformatical approach suggests the function of the autoimmune hepatitis target antigen soluble liver antigen/liver pancreas. Hepatology 2001, 34, 230–233. [Google Scholar]
- Krauth-Siegel, L.R.; Comini, M.A.; Schlecker, T. The trypanothione system. Subcell. Biochem. 2007, 44, 231–251. [Google Scholar] [CrossRef]
- Fairlamb, A.H.; Blackburn, P.; Ulrich, P.; Chait, B.T.; Cerami, A. Trypanothione: a novel bis(glutathionyl)spermidine cofactor for glutathione reductase in trypanosomatids. Science 1985, 227, 1485–1487. [Google Scholar]
- Snell, K.; Natsumeda, Y.; Eble, J.N.; Glover, J.L.; Weber, G. Enzymic imbalance in serine metabolism in human colon carcinoma and rat sarcoma. Br. J. Cancer 1988, 57, 87–90. [Google Scholar] [CrossRef]
- Snell, K.; Natsumeda, Y.; Weber, G. The modulation of serine metabolism in hepatoma 3924A during different phases of cellular proliferation in culture. Biochem. J. 1987, 245, 609–612. [Google Scholar]
- Chang, S.J. Vitamin B6 antagonists alter the function and ultrastructure of mice endothelial cells. J. Nutr. Sci. Vitaminol. (Tokyo) 2000, 46, 149–153. [Google Scholar]
- Renwick, S.B.; Skelly, J.V.; Chave, K.J.; Sanders, P.G.; Snell, K.; Baumann, U. Purification, crystallization and preliminary X-ray analysis of human recombinant cytosolic serine hydroxymethyltransferase. Acta Crystallogr. D. Biol. Crystallogr. 1998, 54, 1030–1031. [Google Scholar] [CrossRef]
- Pires, A.R.; Araujo, A.N.; Montenegro, M.C.; Chocholous, P.; Solich, P. New ionophores for vitamin B1 and vitamin B6 potentiometric sensors for multivitaminic control. J. Pharm. Biomed. Anal. 2008, 46, 683–691. [Google Scholar] [CrossRef]
- Ishihara, J.; Iso, H.; Inoue, M.; Iwasaki, M.; Okada, K.; Kita, Y.; Kokubo, Y.; Okayama, A.; Tsugane, S. Intake of folate, vitamin B6 and vitamin B12 and the risk of CHD: the Japan Public Health Center-Based Prospective Study Cohort I. J. Am. Coll. Nutr. 2008, 27, 127–136. [Google Scholar]
- Czeizel, A.E.; Puho, E.; Banhidy, F.; Acs, N. Oral pyridoxine during pregnancy : potential protective effect for cardiovascular malformations. Drugs R. D. 2004, 5, 259–269. [Google Scholar] [CrossRef]
- Merrill, R.M.; Taylor, P.; Aldana, S.G. Coronary Health Improvement Project (CHIP) is associated with improved nutrient intake and decreased depression. Nutrition 2008, 24, 314–321. [Google Scholar] [CrossRef]
- Booth, G.L.; Wang, E.E. Preventive health care, 2000 update: screening and management of hyperhomocysteinemia for the prevention of coronary artery disease events. The Canadian Task Force on Preventive Health Care. CMAJ 2000, 163, 21–29. [Google Scholar]
- Nozaki, T.; Shigeta, Y.; Saito-Nakano, Y.; Imada, M.; Kruger, W.D. Characterization of transsulfuration and cysteine biosynthetic pathways in the protozoan hemoflagellate, Trypanosoma cruzi. Isolation and molecular characterization of cystathionine beta-synthase and serine acetyltransferase from Trypanosoma. J. Biol. Chem. 2001, 276, 6516–6523. [Google Scholar]
- Graham, I.M.; Daly, L.E.; Refsum, H.M.; Robinson, K.; Brattstrom, L.E.; Ueland, P.M.; Palma-Reis, R.J.; Boers, G.H.; Sheahan, R.G.; Israelsson, B.; Uiterwaal, C.S.; Meleady, R.; McMaster, D.; Verhoef, P.; Witteman, J.; Rubba, P.; Bellet, H.; Wautrecht, J.C.; de Valk, H.W.; Sales Luis, A.C.; Parrot-Rouland, F.M.; Tan, K.S.; Higgins, I.; Garcon, D.; Andria, G. Plasma homocysteine as a risk factor for vascular disease. The European Concerted Action Project. JAMA 1997, 277, 1775–1781. [Google Scholar] [Green Version]
- Lal, K.J.; Dakshinamurti, K. The relationship between low-calcium-induced increase in systolic blood pressure and vitamin B6. J. Hypertens. 1995, 13, 327–332. [Google Scholar]
- Lal, K.J.; Dakshinamurti, K.; Thliveris, J. The effect of vitamin B6 on the systolic blood pressure of rats in various animal models of hypertension. J. Hypertens. 1996, 14, 355–363. [Google Scholar] [CrossRef]
- Dakshinamurti, K.; Lal, K.J.; Ganguly, P.K. Hypertension, calcium channel and pyridoxine (vitamin B6). Mol. Cell Biochem. 1998, 188, 137–148. [Google Scholar] [CrossRef]
- Aybak, M.; Sermet, A.; Ayyildiz, M.O.; Karakilcik, A.Z. Effect of oral pyridoxine hydrochloride supplementation on arterial blood pressure in patients with essential hypertension. Arzneimittelforschung 1995, 45, 1271–1273. [Google Scholar]
- Dakshinamurti, K.; Dakshinamurti, S. Blood pressure regulation and micronutrients. Nutr. Res. Rev. 2001, 14, 3–44. [Google Scholar] [CrossRef]
- Okuda, T.; Sumiya, T.; Iwai, N.; Miyata, T. Pyridoxine 5'-phosphate oxidase is a candidate gene responsible for hypertension in Dahl-S rats. Biochem. Biophys. Res. Commun. 2004, 313, 647–53. [Google Scholar] [CrossRef]
- Lieber, C.S. Mechanism of ethanol induced hepatic injury. Pharmacol. Ther. 1990, 46, 1–41. [Google Scholar] [CrossRef]
- Vasdev, S.; Whalen, M.; Ford, C. A.; Longerich, L.; Prabhakaran, V.; Parai, S. Ethanol- and threonine-induced hypertension in rats: a common mechanism. Can. J. Cardiol. 1995, 11, 807–815. [Google Scholar]
- Vasdev, S.; Wadhawan, S.; Ford, C.A.; Parai, S.; Longerich, L.; Gadag, V. Dietary vitamin B6 supplementation prevents ethanol-induced hypertension in rats. Nutr. Metab. Cardiovasc. Dis. 1999, 9, 55–63. [Google Scholar]
- Gloria, L.; Cravo, M.; Camilo, M.E.; Resende, M.; Cardoso, J.N.; Oliveira, A.G.; Leitao, C.N.; Mira, F.C. Nutritional deficiencies in chronic alcoholics: relation to dietary intake and alcohol consumption. Am. J. Gastroenterol. 1997, 92, 485–489. [Google Scholar]
- Huber, K.H.; Rexroth, W.; Werle, E.; Koeth, T.; Weicker, H.; Hild, R. Sympathetic neuronal activity in diabetic and non-diabetic subjects with peripheral arterial occlusive disease. Klin. Wochenschr. 1991, 69, 233–238. [Google Scholar] [CrossRef]
- Cicila, G.T. Strategy for uncovering complex determinants of hypertension using animal models. Curr. Hypertens. Rep. 2000, 2, 217–226. [Google Scholar] [CrossRef]
- MacKenzie, K.E.; Wiltshire, E.J.; Gent, R.; Hirte, C.; Piotto, L.; Couper, J.J. Folate and vitamin B6 rapidly normalize endothelial dysfunction in children with type 1 diabetes mellitus. Pediatrics 2006, 118, 242–253. [Google Scholar] [CrossRef]
- Nakamura, S.; Li, H.; Adijiang, A.; Pischetsrieder, M.; Niwa, T. Pyridoxal phosphate prevents progression of diabetic nephropathy. Nephrol. Dial. Transplant 2007, 22, 2165–2174. [Google Scholar] [CrossRef]
- Adrover, M.; Vilanova, B.; Munoz, F.; Donoso, J. Inhibition of glycosylation processes: the reaction between pyridoxamine and glucose. Chem. Biodivers. 2005, 2, 964–975. [Google Scholar] [CrossRef]
- Nagore, E.; Insa, A.; Sanmartin, O. Antineoplastic therapy-induced palmar plantar erythrodysesthesia ('hand-foot') syndrome. Incidence, recognition and management. Am. J. Clin. Dermatol. 2000, 1, 225–234. [Google Scholar]
- Taylor, B.V.; Oudit, G.Y.; Evans, M. Homocysteine, vitamins, and coronary artery disease. Comprehensive review of the literature. Can. Fam. Physician 2000, 46, 2236–2245. [Google Scholar]
- Matsubara, K.; Matsumoto, H.; Mizushina, Y.; Lee, J.S.; Kato, N. Inhibitory effect of pyridoxal 5'-phosphate on endothelial cell proliferation, replicative DNA polymerase and DNA topoisomerase. Int. J. Mol. Med. 2003, 12, 51–55. [Google Scholar]
- Bostom, A.G.; Carpenter, M.A.; Kusek, J.W.; Hunsicker, L.G.; Pfeffer, M.A.; Levey, A.S.; Jacques, P.F.; McKenney, J. Rationale and design of the Folic Acid for Vascular Outcome Reduction In Transplantation (FAVORIT) trial. Am. Heart J. 2006, 152, 448. [Google Scholar]
- Menon, V.; Wang, X.; Greene, T.; Beck, G.J.; Kusek, J.W.; Selhub, J.; Levey, A.S.; Sarnak, M.J. Homocysteine in chronic kidney disease: Effect of low protein diet and repletion with B vitamins. Kidney Int. 2005, 67, 1539–1546. [Google Scholar] [CrossRef]
- Adrover, M.; Vilanova, B.; Frau, J.; Munoz, F.; Donoso, J. A comparative study of the chemical reactivity of pyridoxamine, Ac-Phe-Lys and Ac-Cys with various glycating carbonyl compounds. Amino Acids 2009, 36, 437–448. [Google Scholar] [CrossRef]
- Metz, T.O.; Alderson, N.L.; Thorpe, S.R.; Baynes, J.W. Pyridoxamine, an inhibitor of advanced glycation and lipoxidation reactions: a novel therapy for treatment of diabetic complications. Arch. Biochem. Biophys. 2003, 419, 41–49. [Google Scholar] [CrossRef]
- Voziyan, P.A.; Metz, T.O.; Baynes, J.W.; Hudson, B.G. A post-Amadori inhibitor pyridoxamine also inhibits chemical modification of proteins by scavenging carbonyl intermediates of carbohydrate and lipid degradation. J. Biol. Chem. 2002, 277, 3397–3403. [Google Scholar]
- Berger, M.; Gray, J.A.; Roth, B.L. The expanded biology of serotonin. Annu. Rev. Med. 2009, 60, 355–66. [Google Scholar] [CrossRef]
- Matxain, J.M.; Padro, D.; Ristila, M.; Strid, A.; Eriksson, L.A. Evidence of high *OH radical quenching efficiency by vitamin B6. J. Phys. Chem. B. 2009, 113, 9629–9632. [Google Scholar] [CrossRef]
- Belelli, D.; Harrison, N.L.; Maguire, J.; Macdonald, R.L.; Walker, M.C.; Cope, D.W. Extrasynaptic GABAA receptors: form, pharmacology, and function. J. Neurosci. 2009, 29, 12757–12763. [Google Scholar]
- Hvas, A.M.; Juul, S.; Bech, P.; Nexo, E. Vitamin B6 level is associated with symptoms of depression. Psychother. Psychosom. 2004, 73, 340–343. [Google Scholar] [CrossRef]
- Hoffmann, G.F.; Schmitt, B.; Windfuhr, M.; Wagner, N.; Strehl, H.; Bagci, S.; Franz, A. R.; Mills, P.B.; Clayton, P.T.; Baumgartner, M.R.; Steinmann, B.; Bast, T.; Wolf, N.I.; Zschocke, J. Pyridoxal 5'-phosphate may be curative in early-onset epileptic encephalopathy. J. Inherit. Metab. Dis. 2007, 30, 96–99. [Google Scholar] [CrossRef]
- Lott, I.T.; Coulombe, T.; Di Paolo, R.V.; Richardson, E.P. Jr.; Levy, H.L. Vitamin B6-dependent seizures: pathology and chemical findings in brain. Neurology 1978, 28, 47–54. [Google Scholar] [CrossRef]
- McCarty, M.F. High-dose pyridoxine as an 'anti-stress' strategy. Med. Hypotheses 2000, 54, 803–807. [Google Scholar] [CrossRef]
- Leuendorf, J.E.; Genau, A.; Szewczyk, A.; Mooney, S.; Drewke, C.; Leistner, E.; Hellmann, H. The Pdx1 family is structurally and functionally conserved between Arabidopsis thaliana and Ginkgo biloba. FEBS J. 2008, 275, 960–969. [Google Scholar]
- Kästner, U.; Hallmen, C.; Wiese, M.; Leistner, E.; Drewke, C. The human pyridoxal kinase, a plausible target for ginkgotoxin from Ginkgo biloba. FEBS J. 2007, 274, 1036–1045. [Google Scholar] [CrossRef]
- Denslow, S.A.; Walls, A.A.; Daub, M.E. Regulation of biosynthetic genes and antioxidant properties of vitamin B6 vitamers during plant defense responses. Physiol. Mol. Plant Path. 2005, 66, 244–255. [Google Scholar] [CrossRef]
- Bilski, P.; Li, M.Y.; Ehrenshaft, M.; Daub, M.E.; Chignell, C.F. Vitamin B6 (pyridoxine) and its derivatives are efficient singlet oxygen quenchers and potential fungal antioxidants. Photochem. Photobiol. 2000, 71, 129–134. [Google Scholar] [CrossRef]
- Ehrenshaft, M.; Jenns, A.E.; Chung, K.R.; Daub, M.E. SOR1, a gene required for photosensitizer and singlet oxygen resistance in Cercospora fungi, is highly conserved in divergent organisms. Mol. Cell 1998, 1, 603–609. [Google Scholar] [CrossRef]
- Chen, H.; Xiong, L. Pyridoxine is required for post-embryonic root development and tolerance to osmotic and oxidative stresses. Plant J. 2005, 44, 396–408. [Google Scholar] [CrossRef]
- Chumnantana, R.; Yokochi, N.; Yagi, T. Vitamin B6 compounds prevent the death of yeast cells due to menadione, a reactive oxygen generator. Biochim. Biophys. Acta 2005, 1722, 84–91. [Google Scholar]
- DiSorbo, D.M.; Wagner, R. Jr.; Nathanson, L. In vivo and in vitro inhibition of B16 melanoma growth by vitamin B6. Nutr. Cancer 1985, 7, 43–52. [Google Scholar] [CrossRef]
- DiSorbo, D.M.; Nathanson, L. High-dose pyridoxal supplemented culture medium inhibits the growth of a human malignant melanoma cell line. Nutr. Cancer 1983, 5, 10–15. [Google Scholar] [CrossRef]
- Komatsu, S.; Yanaka, N.; Matsubara, K.; Kato, N. Antitumor effect of vitamin B6 and its mechanisms. Biochim. Biophys. Acta 2003, 1647, 127–130. [Google Scholar]
- Stolzenberg-Solomon, R.Z.; Albanes, D.; Nieto, F.J.; Hartman, T.J.; Tangrea, J.A.; Rautalahti, M.; Sehlub, J.; Virtamo, J.; Taylor, P.R. Pancreatic cancer risk and nutrition-related methyl-group availability indicators in male smokers. J. Natl. Cancer Inst. 1999, 91, 535–541. [Google Scholar] [CrossRef]
- Harbige, L.S. Nutrition and immunity with emphasis on infection and autoimmune disease. Nutr. Health 1996, 10, 285–312. [Google Scholar] [CrossRef]
- Delport, R.; Ubbink, J.B.; Bosman, H.; Bissbort, S.; Vermaak, W.J. Altered vitamin B6 homeostasis during aminophylline infusion in the beagle dog. Int. J. Vitam. Nutr. Res. 1990, 60, 35–40. [Google Scholar]
- Delport, R.; Ubbink, J.B.; Serfontein, W.J.; Becker, P.J.; Walters, L. Vitamin B6 nutritional status in asthma: the effect of theophylline therapy on plasma pyridoxal-5'-phosphate and pyridoxal levels. Int. J. Vitam. Nutr. Res. 1988, 58, 67–72. [Google Scholar]
- van den Berg, H.; Louwerse, E.S.; Bruinse, H.W.; Thissen, J.T.; Schrijver, J. Vitamin B6 status of women suffering from premenstrual syndrome. Hum. Nutr. Clin. Nutr. 1986, 40, 441–450. [Google Scholar]
- Bernstein, A.L. Vitamin B6 in clinical neurology. Ann. N. Y. Acad. Sci. 1990, 585, 250–260. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hellmann, H.; Mooney, S. Vitamin B6: A Molecule for Human Health? Molecules 2010, 15, 442-459. https://doi.org/10.3390/molecules15010442
Hellmann H, Mooney S. Vitamin B6: A Molecule for Human Health? Molecules. 2010; 15(1):442-459. https://doi.org/10.3390/molecules15010442
Chicago/Turabian StyleHellmann, Hanjo, and Sutton Mooney. 2010. "Vitamin B6: A Molecule for Human Health?" Molecules 15, no. 1: 442-459. https://doi.org/10.3390/molecules15010442
APA StyleHellmann, H., & Mooney, S. (2010). Vitamin B6: A Molecule for Human Health? Molecules, 15(1), 442-459. https://doi.org/10.3390/molecules15010442