
The Stiffness of Cardiac Fibroblast Substrates Exerts a Regulatory Influence on Collagen Metabolism via α2β1 Integrin, FAK and Src Kinases


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Polyacrylamide Gel Preparation
2.2. Cell Culture
2.3. Reagent Preparation
2.4. ITGA2 Gene Silencing
2.5. qPCR
2.6. Collagen Content
2.7. Enzyme-Linked Immunosorbent Assay (ELISA)
2.8. Statistical Analysis
3. Results
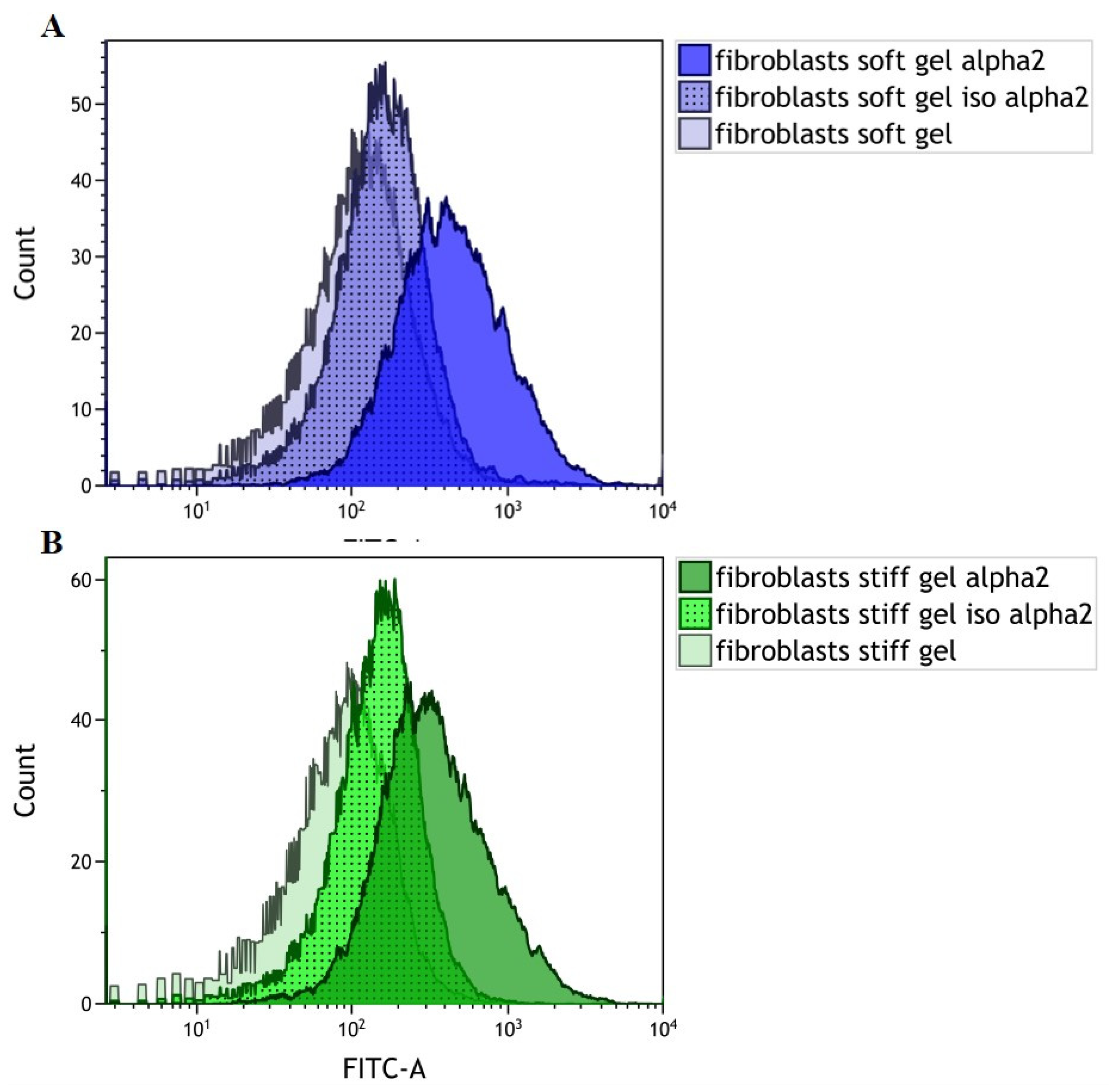
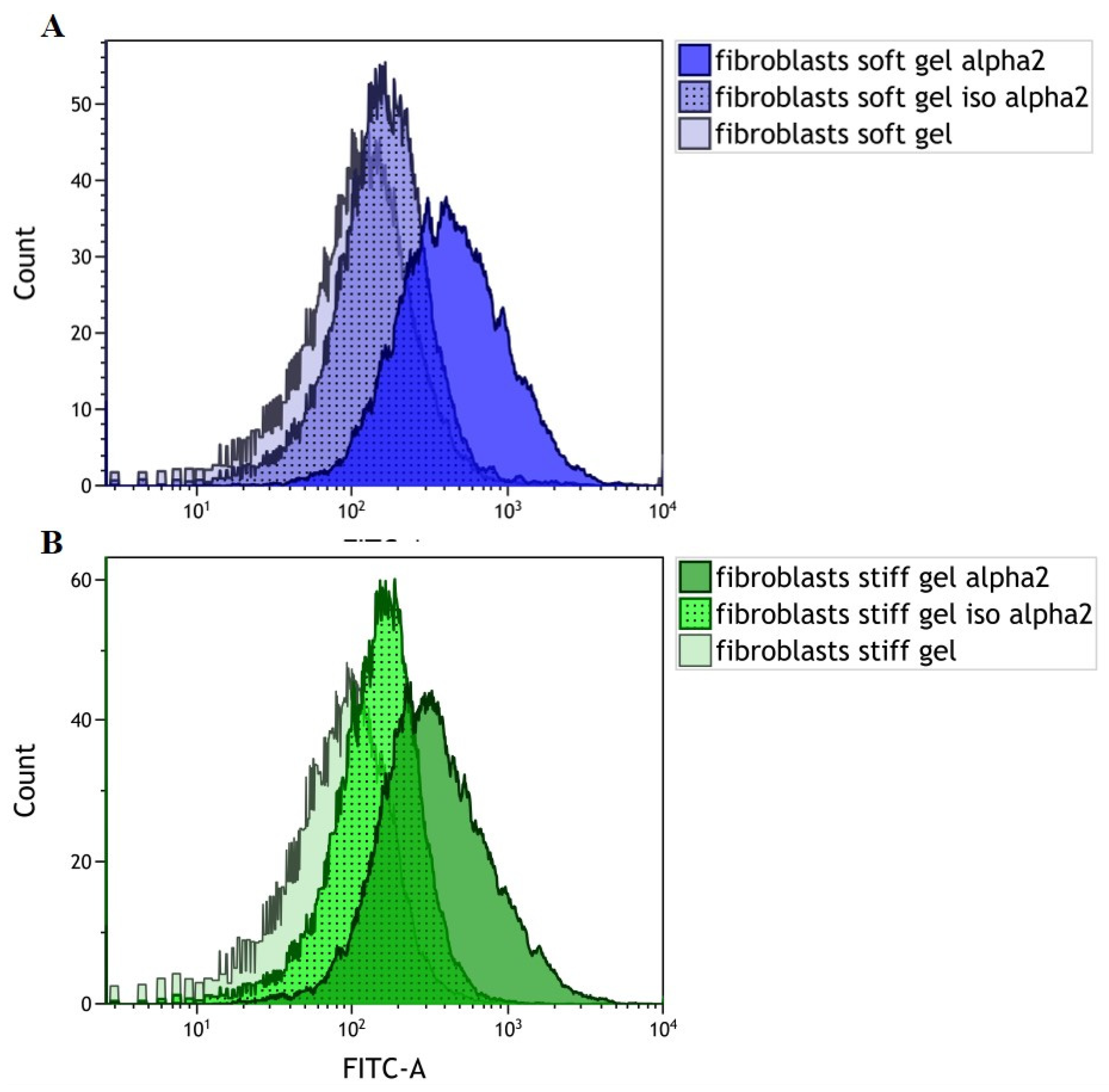
3.1. Evaluation of α2 Integrin Subunit Density in Cardiac Fibroblasts Cultured on Gels with Different Rigidities
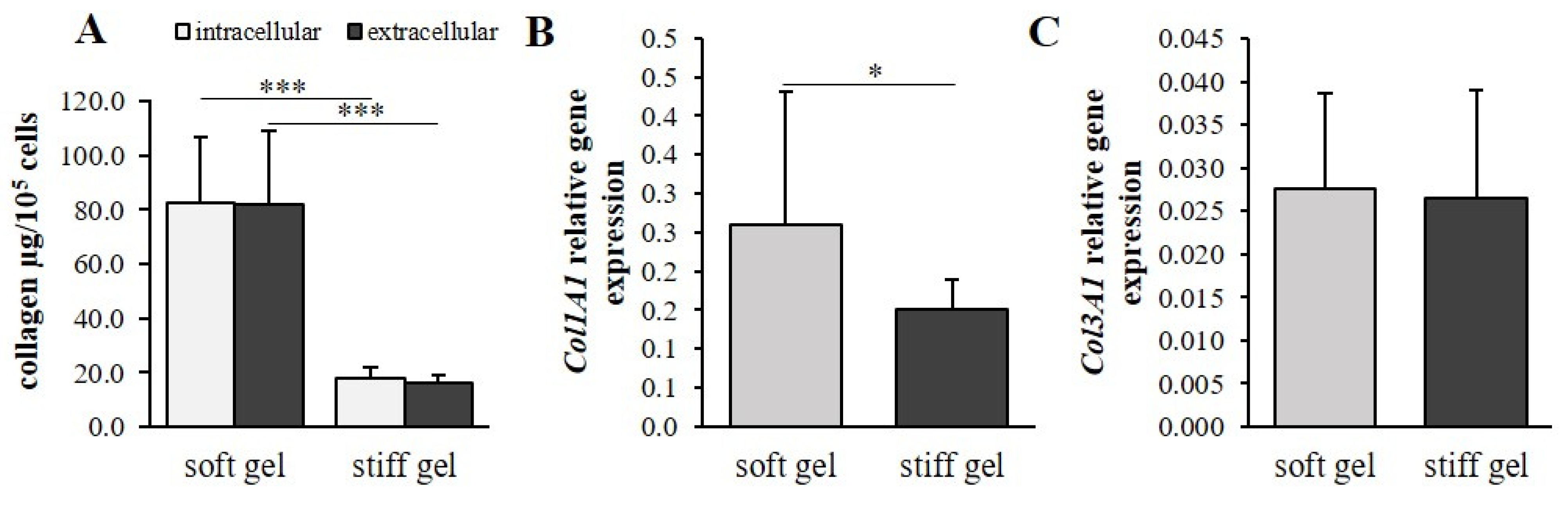
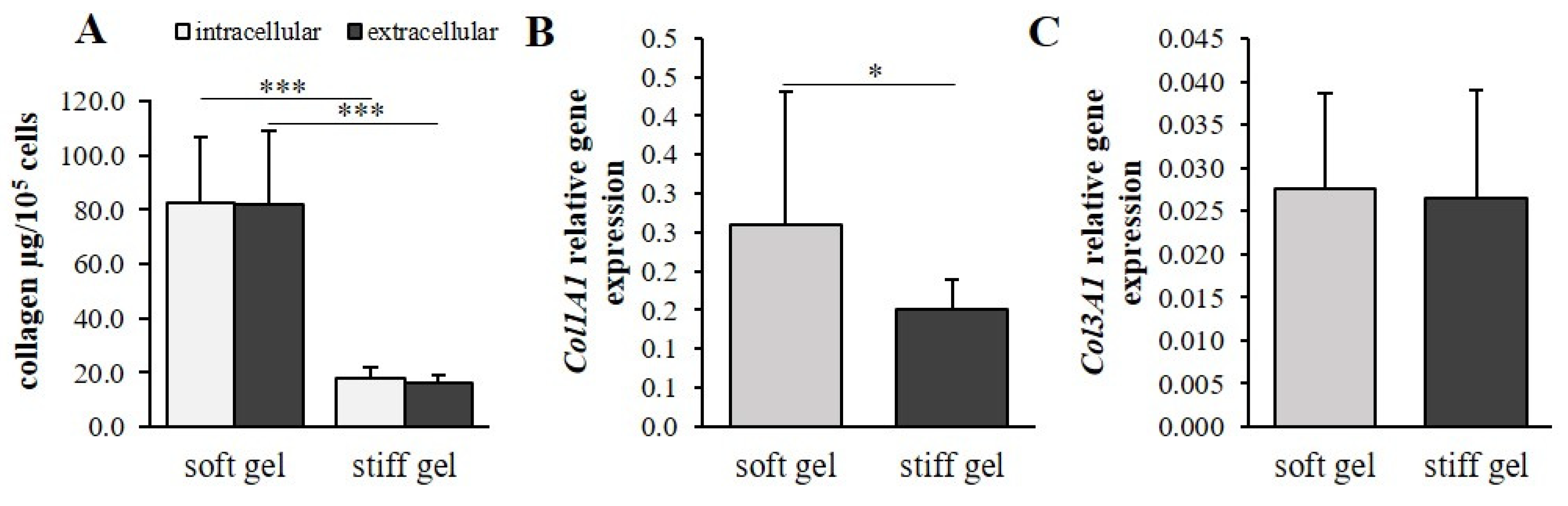
3.2. The Mechanical Properties of the Cell Environment Modulate Collagen Content in Cardiac Fibroblasts
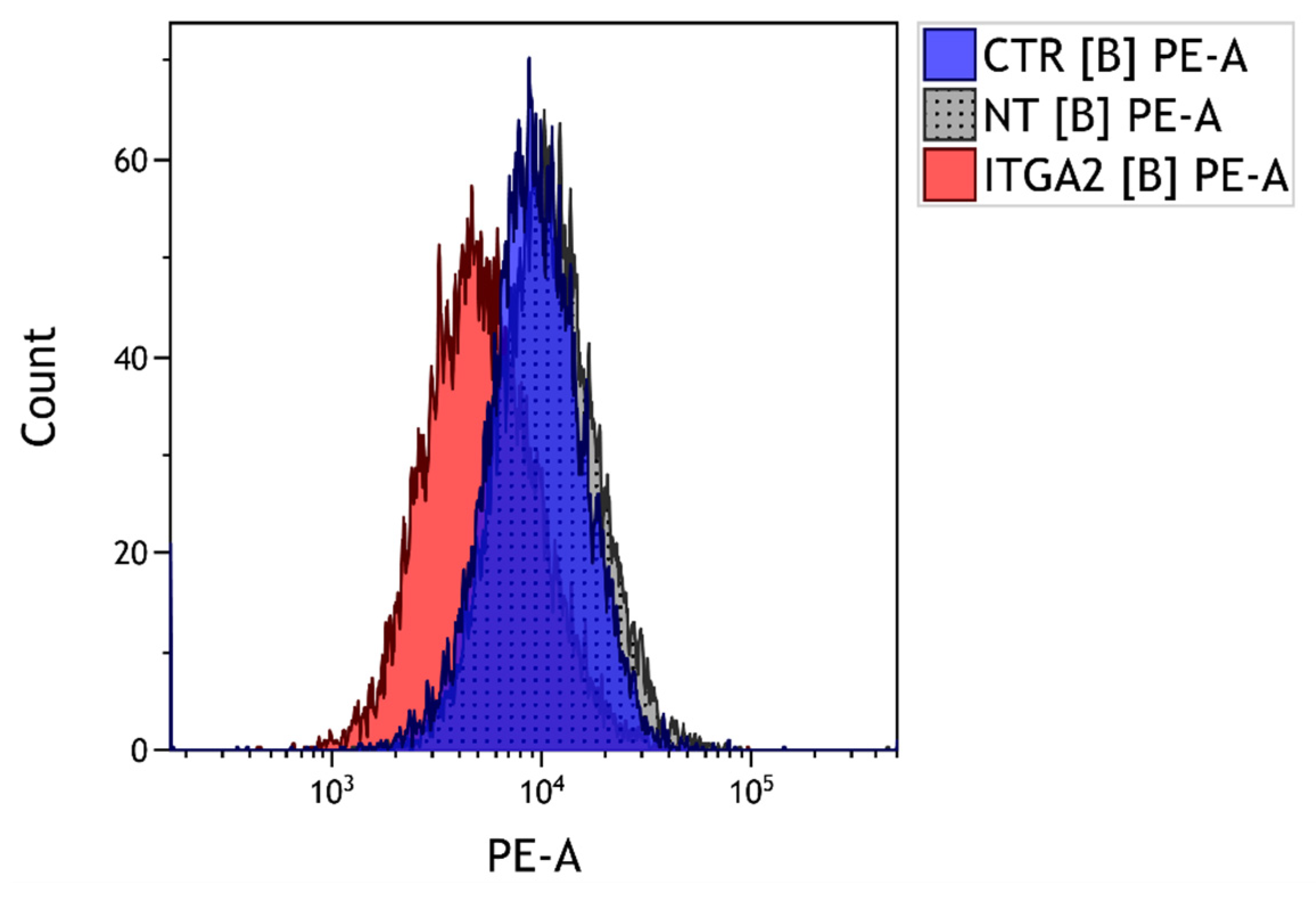
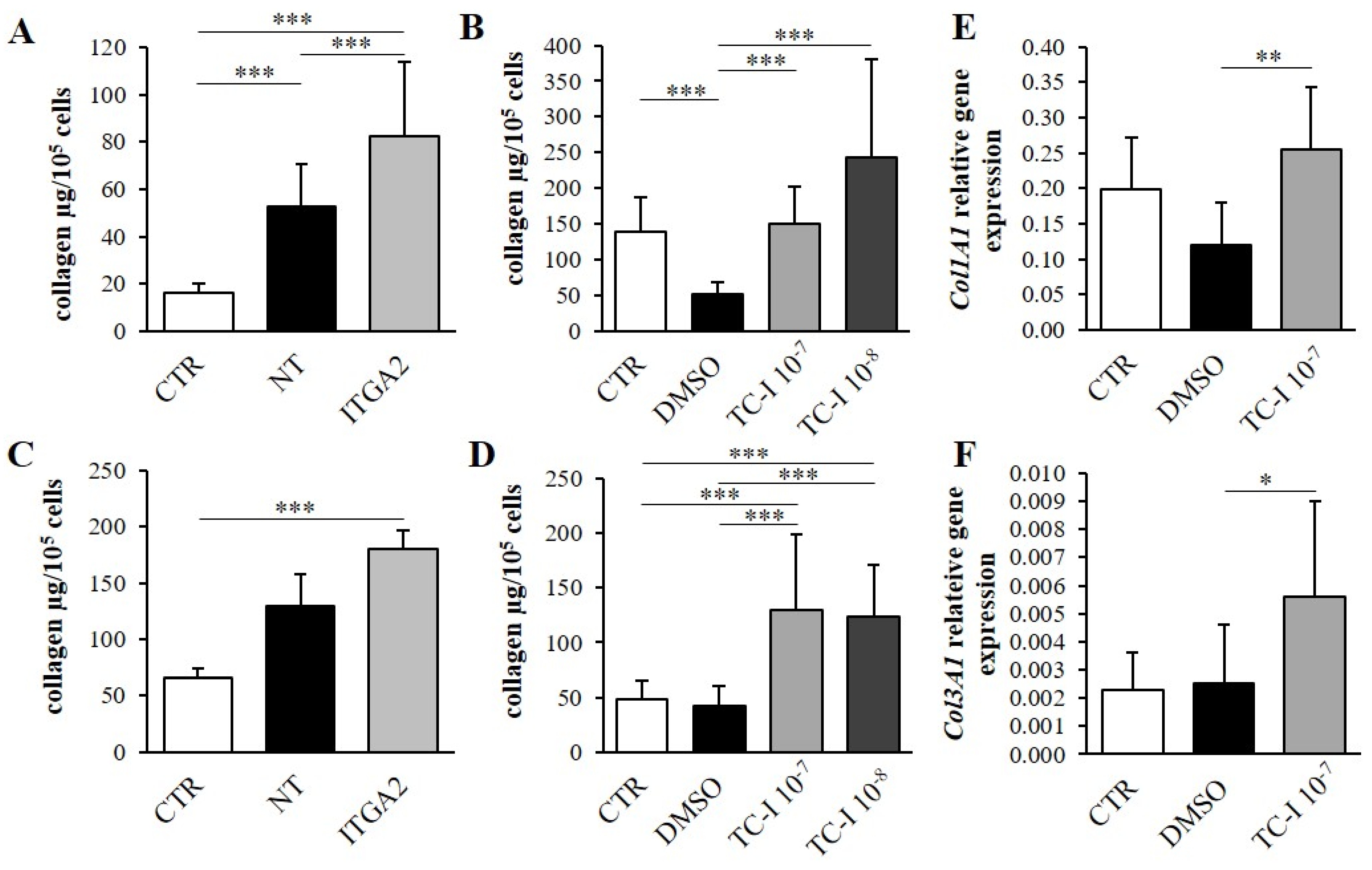
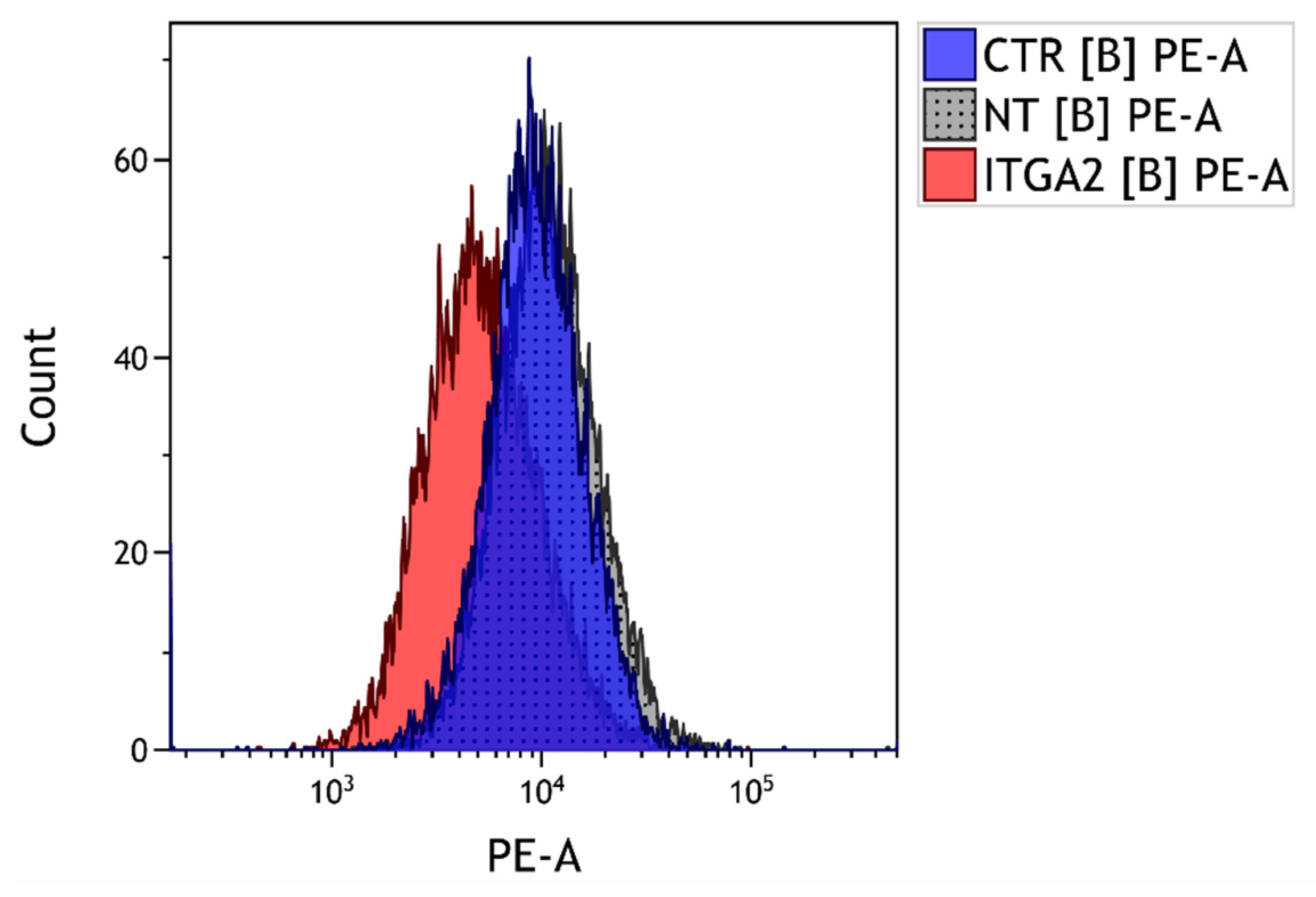
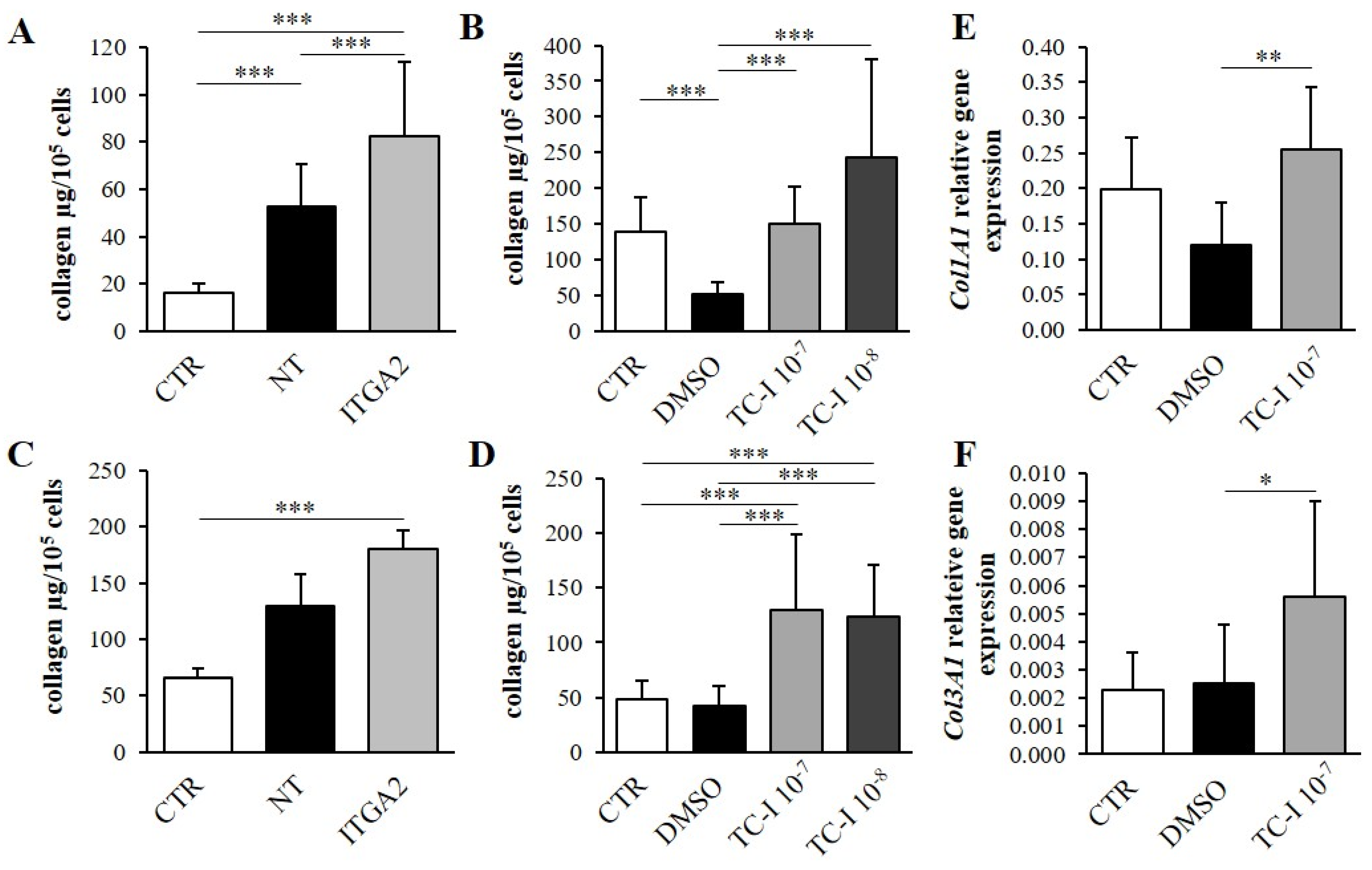
3.3. The Effect of α2 Integrin Subunit Inhibition on Collagen Content in Vitro
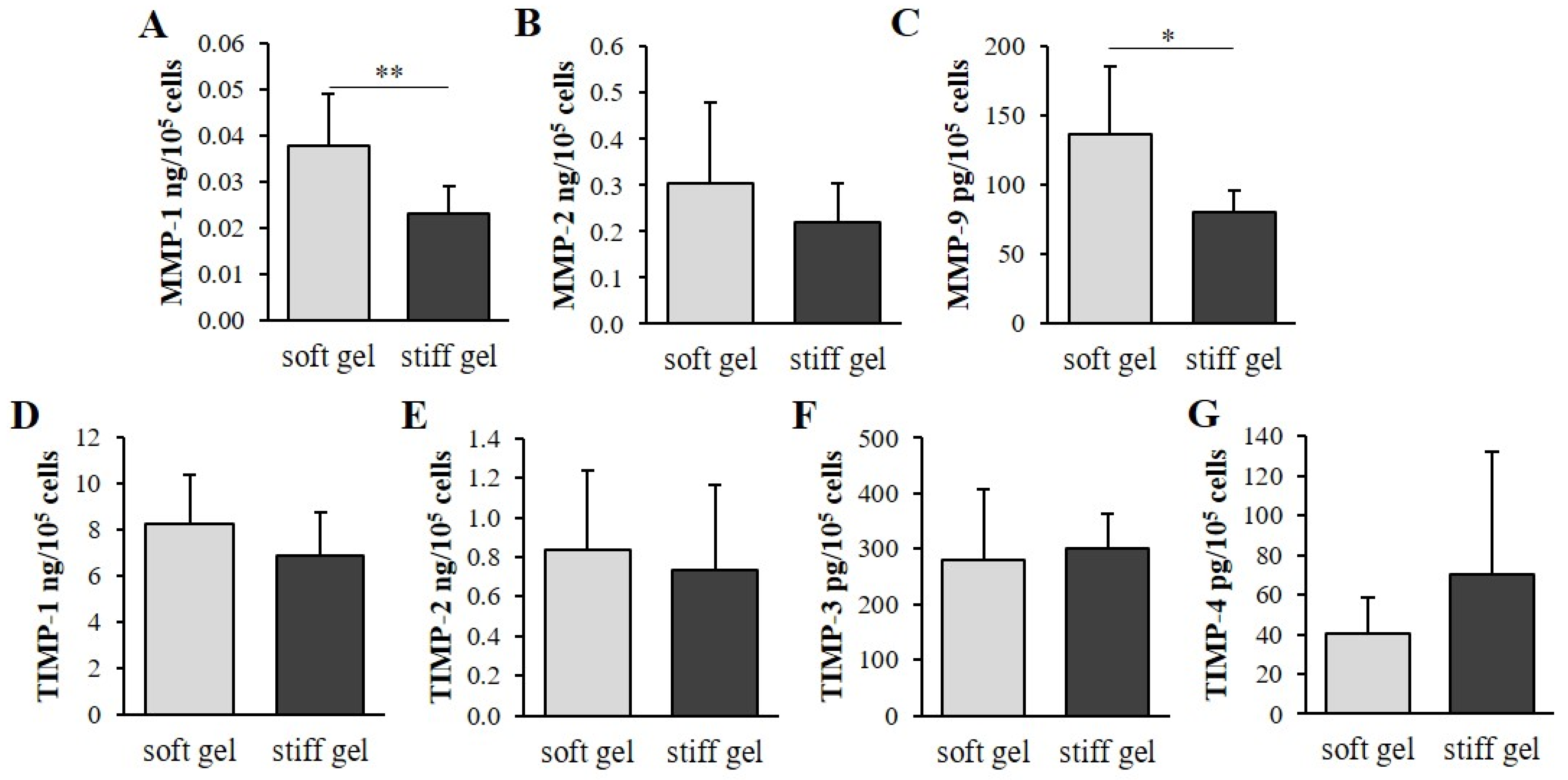
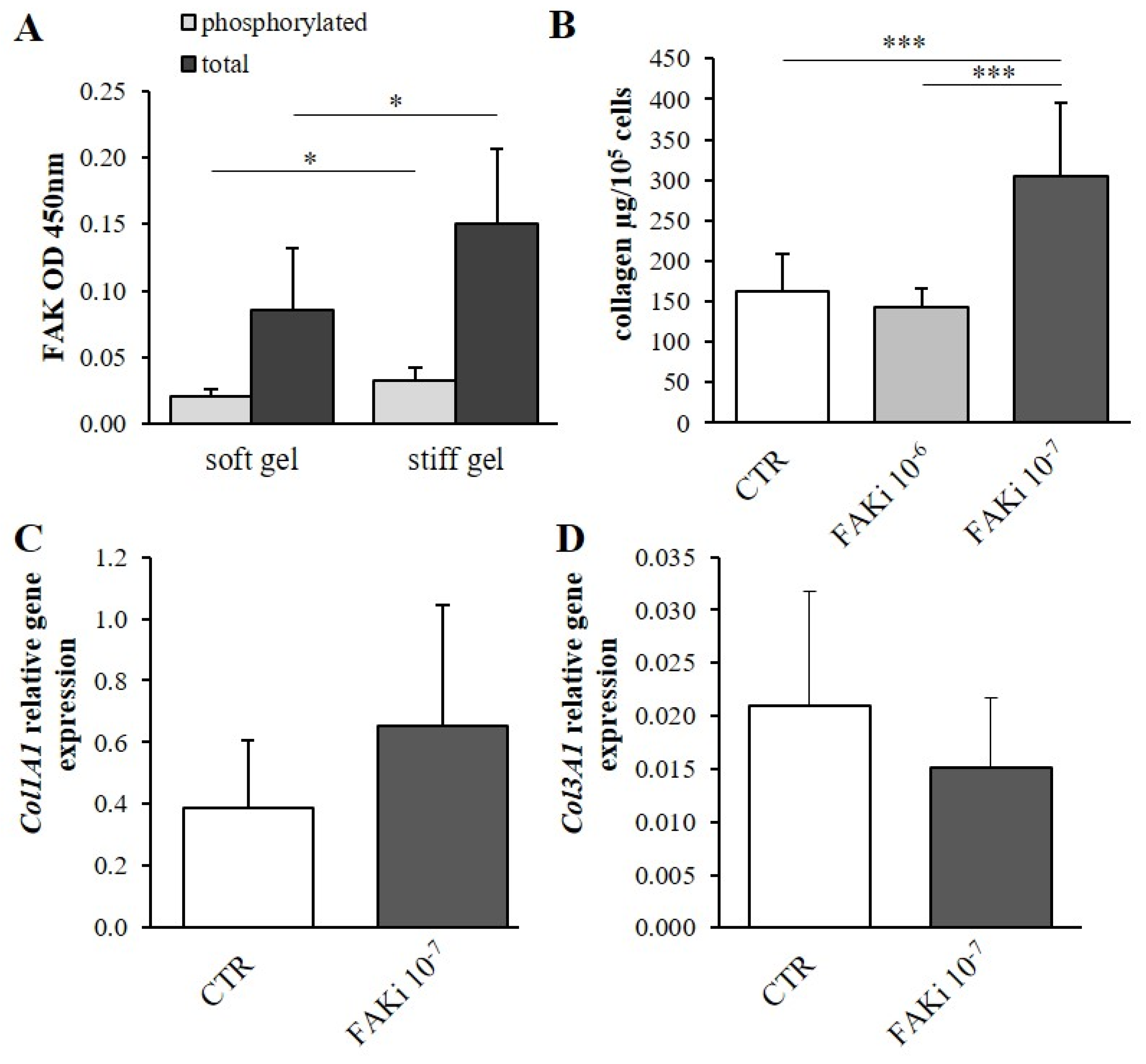
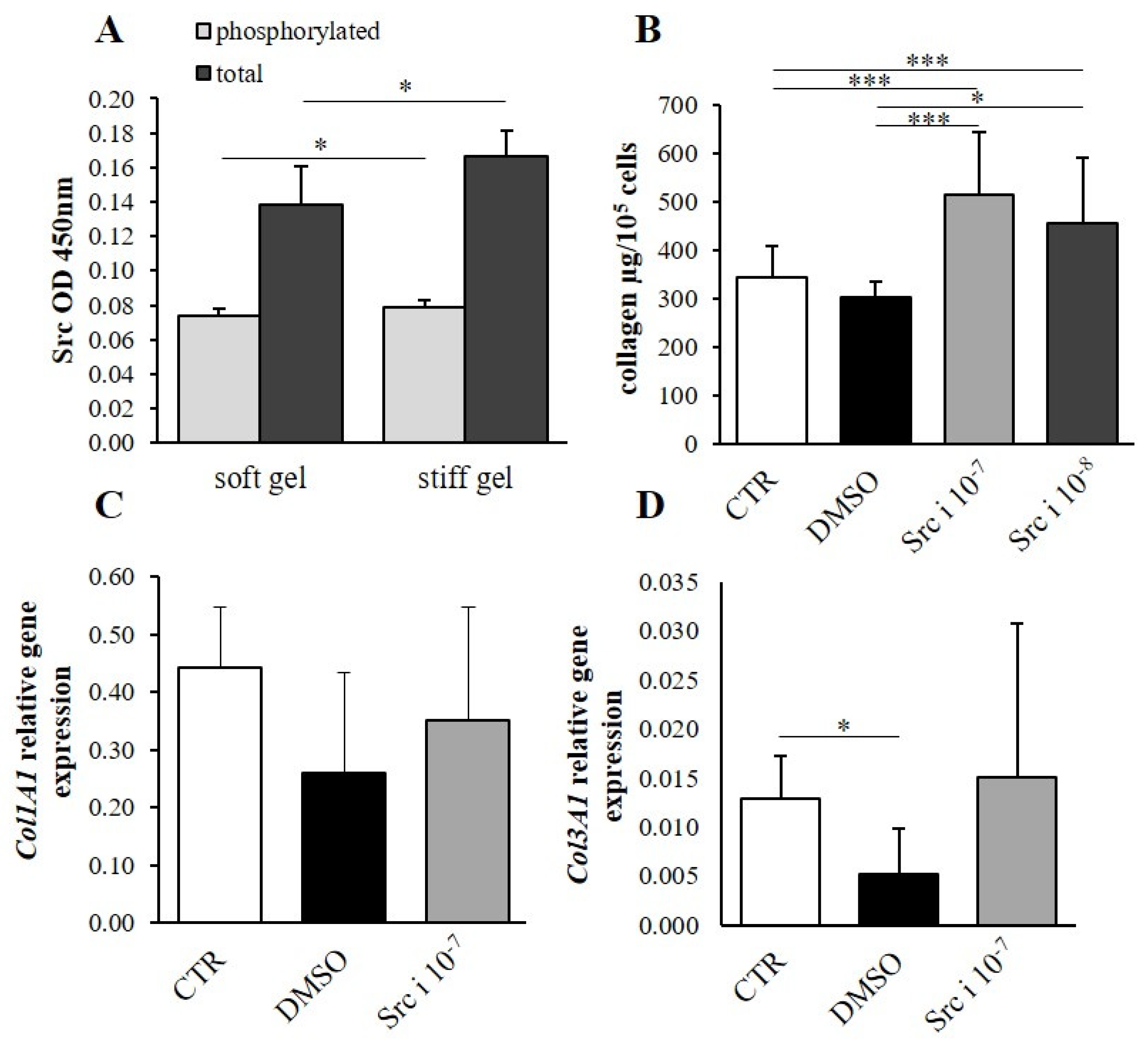
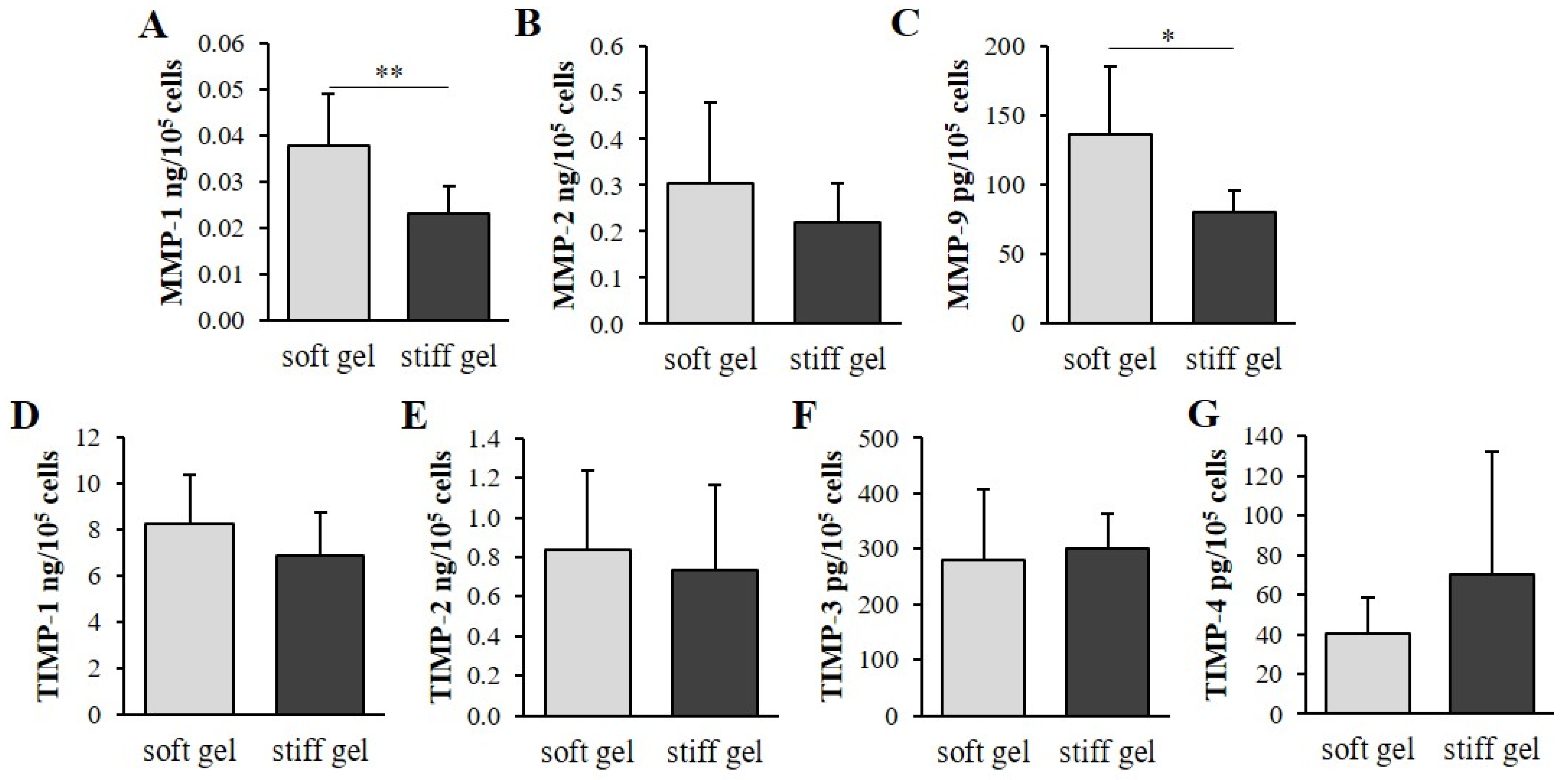
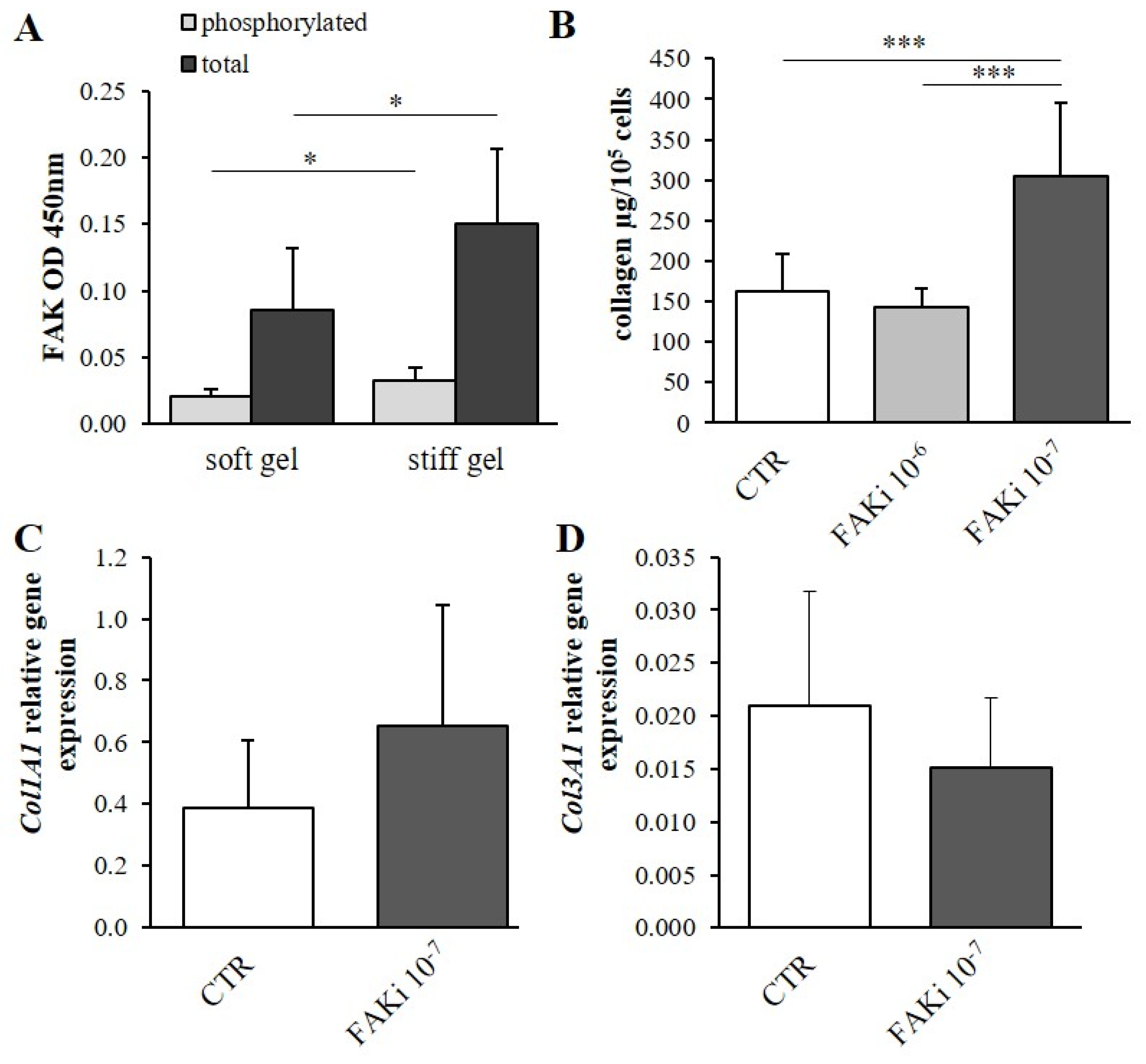
3.4. Determination of the Effect of Mechanical Properties on α2β1 Integrin Second Messengers
3.5. The Effect of Second Messenger Inhibition on Collagen Content
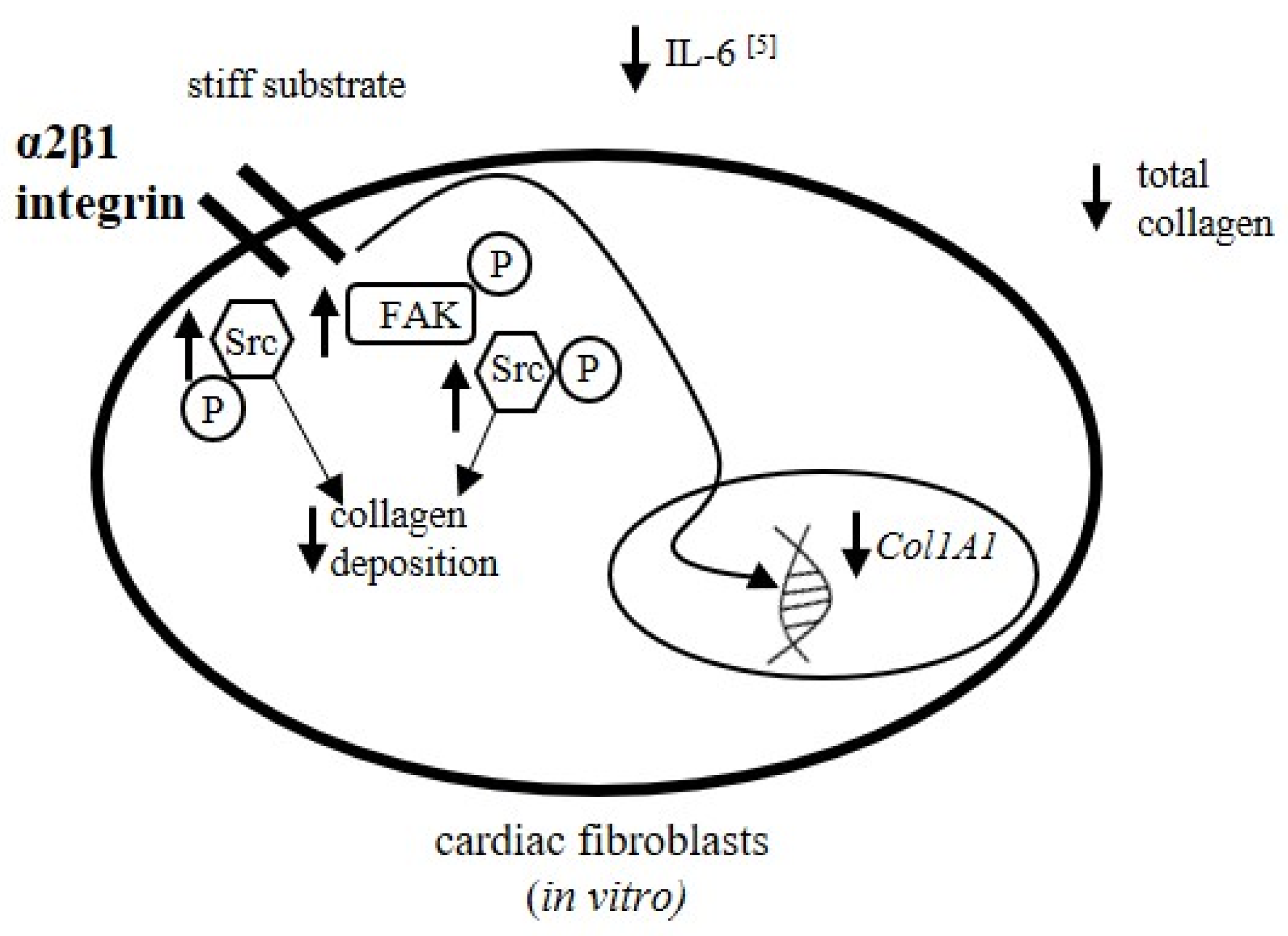
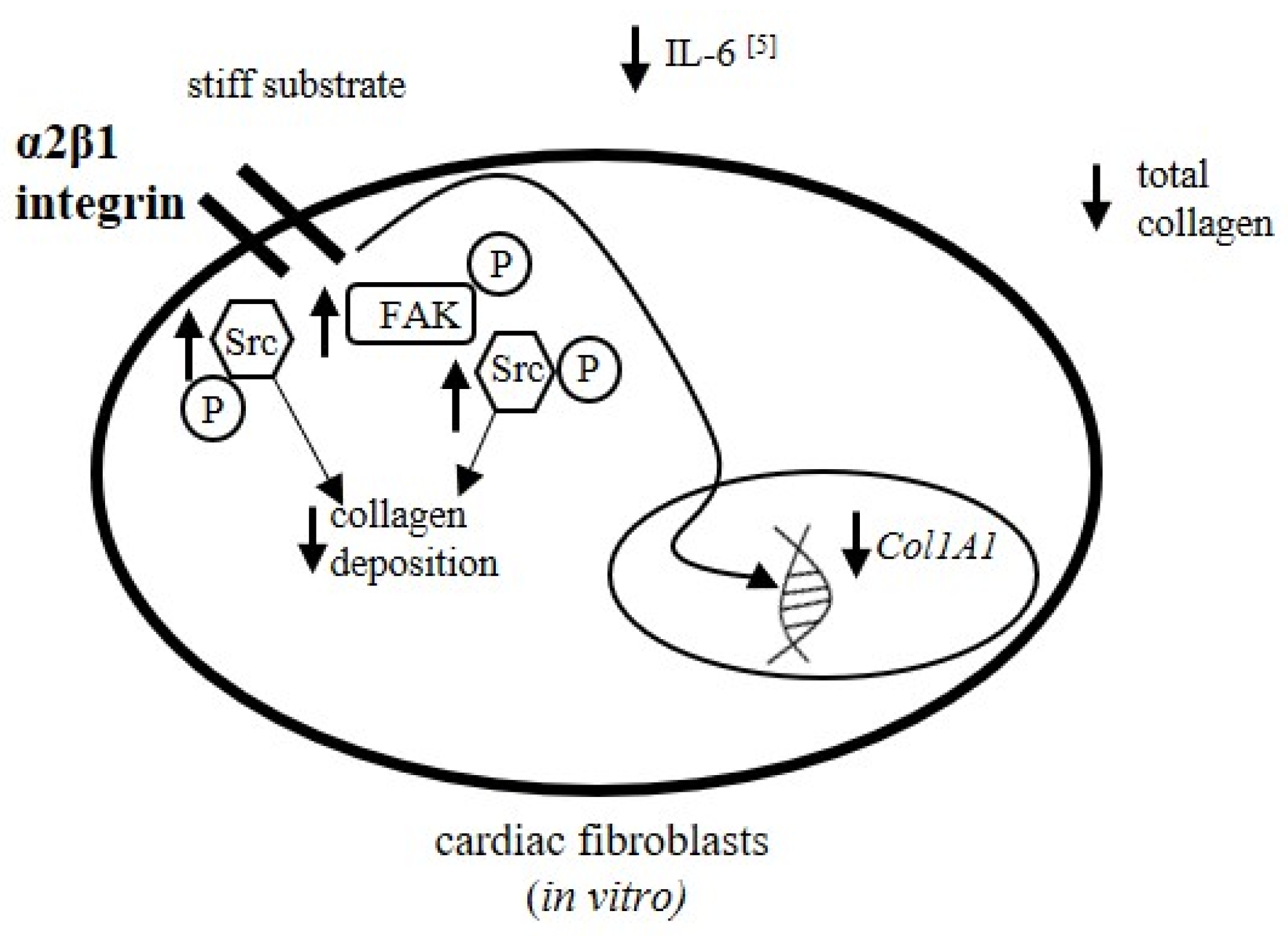
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Frangogiannis, N.G. Cardiac fibrosis: Cell biological mechanisms, molecular pathways and therapeutic opportunities. Mol. Asp. Med. 2019, 65, 70–99. [Google Scholar] [CrossRef] [PubMed]
- Kong, P.; Christia, P.; Frangogiannis, N.G. The pathogenesis of cardiac fibrosis. Cell. Mol. Life Sci. 2014, 71, H549–H574. [Google Scholar] [CrossRef] [Green Version]
- Krenning, G.; Zeisberg, E.M.; Kalluri, R. The origin of fibroblasts and mechanism of cardiac fibrosis. J. Cell. Physiol. 2010, 225, 631–637. [Google Scholar] [CrossRef] [Green Version]
- Meléndez, G.C.; McLarty, J.L.; Levick, S.P.; Du, Y.; Janicki, J.S.; Brower, G.L. Interleukin 6 mediates myocardial fibrosis, concentric hypertrophy, and diastolic dysfunction in rats. Hypertension 2010, 56, 225–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gałdyszyńska, M.; Bobrowska, J.; Lekka, M.; Radwańska, P.; Piera, L.; Szymański, J.; Drobnik, J. The stiffness-controlled release of interleukin-6 by cardiac fibroblasts is dependent on integrin α2β1. J. Cell. Mol. Med. 2020, 24, 13853–13862. [Google Scholar] [CrossRef]
- Katsumi, A.; Orr, A.W.; Tzima, E.; Schwartz, M.A. Integrins in mechanotransduction. J. Biol. Chem. 2004, 279, 12001–12004. [Google Scholar] [CrossRef] [Green Version]
- Drobnik, J.; Owczarek, K.; Piera, L.; Tosik, D.; Olczak, S.; Ciosek, J.; Hrabec, E. Melatonin-induced augmentation of collagen deposition in cultures of fibroblasts and myofibroblasts is blocked by luzindole—A melatonin membrane receptors inhibitor. Pharm. Rep. 2013, 65, 642–649. [Google Scholar] [CrossRef]
- Egemnazarov, B.; Crnkovic, S.; Nagy, B.M.; Olschewski, H.; Kwapiszewska, G. Right ventricular fibrosis and dysfunction: Actual concepts and common misconceptions. Matrix Biol. 2018, 68–69, 507–521. [Google Scholar] [CrossRef]
- Eckes, B.; Zweers, M.C.; Zhang, Z.G.; Hallinger, R.; Mauch, C.; Aumailley, M.; Krieg, T. Mechanical tension and integrin alpha 2 beta 1 regulate fibroblast functions. J. Investig. Dermatol. Symp. Proc. 2006, 11, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Mitra, S.K.; Schlaepfer, D.D. Integrin-regulated FAK-Src signaling in normal and cancer cells. Curr. Opin. Cell Biol. 2006, 18, 516–523. [Google Scholar] [CrossRef]
- Rustad, K.C.; Wong, V.W.; Gurtner, G.C. The role of focal adhesion complexes in fibroblast mechanotransduction during scar formation. Differentiation 2013, 86, 87–91. [Google Scholar] [CrossRef]
- Chen, C.; Li, R.; Ross, R.S.; Manso, A.M. Integrins and integrin-related proteins in cardiac fibrosis. J. Mol. Cell. Cardiol. 2016, 93, 162–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hynes, R.O. Integrins: Bidirectional, allosteric signaling machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef] [Green Version]
- Civitarese, R.A.; Talior-Volodarsky, I.; Desjardins, J.F.; Kabir, G.; Switzer, J.; Mitchell, M.; Kapus, A.; McCulloch, C.A.; Gullberg, D.; Connelly, K.A. The alpha11 integrin mediates fibroblast-extracellular matrix-cardiomyocyte interactions in health and disease. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H96–H106. [Google Scholar] [CrossRef] [PubMed]
- González, G.E.; Rhaleb, N.E.; D’Ambrosio, M.A.; Nakagawa, P.; Liu, Y.; Leung, P.; Dai, X.; Yang, X.P.; Peterson, E.L.; Carretero, O.A. Deletion of interleukin-6 prevents cardiac inflammation, fibrosis and dysfunction without affecting blood pressure in angiotensin II-high salt-induced hypertension. J. Hypertens. 2015, 33, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, J.H.; Zhang, Y.Y.; Wang, Y.Z.; Wang, J.; Zhao, Y.; Jin, X.X.; Xue, G.L.; Li, P.H.; Sun, Y.L.; et al. Deletion of interleukin-6 alleviated interstitial fibrosis in streptozotocin-induced diabetic cardiomyopathy of mice through affecting TGFβ1 and miR-29 pathways. Sci. Rep. 2016, 6, 23010. [Google Scholar] [CrossRef]
- Kobayashi, T.; Tanaka, K.; Fujita, T.; Umezawa, H.; Amano, H.; Yoshioka, K.; Naito, Y.; Hatano, M.; Kimura, S.; Tatsumi, K.; et al. Bidirectional role of IL-6 signal in pathogenesis of lung fibrosis. Respir. Res. 2015, 16, 99. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, I.; Fuseler, J.W.; Intwala, A.R.; Baudino, T.A. IL-6 loss causes ventricular dysfunction, fibrosis, reduced capillary density, and dramatically alters the cell populations of the developing and adult heart. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H1694–H1704. [Google Scholar] [CrossRef]
- Bartunek, J.; Wijns, W.; Heyndrickx, G.R.; Vanderheyden, M. Timing of intracoronary bone-marrow-derived stem cell transplantation after ST-elevation myocardial infarction. Nat. Clin. Pract. Cardiovasc. Med. 2006, 3 (Suppl. 1), S52–S56. [Google Scholar] [CrossRef]
- Jones, D.L.; Meridew, J.A.; Link, P.A.; Ducharme, M.T.; Lydon, K.L.; Choi, K.M.; Caporarello, N.; Tan, Q.; Diaz Espinosa, A.M.; Xiong, Y.; et al. ZNF416 is a pivotal transcriptional regulator of fibroblast mechanoactivation. J. Cell Biol. 2021, 220, e202007152. [Google Scholar] [CrossRef]
- Matsumura, T.; Fujimoto, T.; Futakuchi, A.; Takihara, Y.; Watanabe-Kitamura, F.; Takahashi, E.; Inoue-Mochita, M.; Tanihara, H.; Inoue, T. TGF-beta-induced activation of conjunctival fibroblasts is modulated by FGF-2 and substratum stiffness. PLoS ONE 2020, 15, e0242626. [Google Scholar] [CrossRef]
- Allijn, I.; Ribeiro, M.; Poot, A.; Passier, R.; Stamatialis, D. Membranes for Modelling Cardiac Tissue Stiffness In Vitro Based on Poly(trimethylene carbonate) and Poly(ethylene glycol) Polymers. Membranes 2020, 10, 274. [Google Scholar] [CrossRef]
- Glaubitz, M.; Block, S.; Witte, J.; Empen, K.; Gross, S.; Schlicht, R.; Weitmann, K.; Klingel, K.; Kandolf, R.; Hoffmann, W.; et al. Stiffness of left ventricular cardiac fibroblasts is associated with ventricular dilation in patients with recent-onset nonischemic and nonvalvular cardiomyopathy. Circ. J. 2014, 78, 1693–1700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belinchón-Romero, I.; Bellot, P.; Romero-Pérez, D.; Herraiz-Romero, I.; Marco, F.; Frances, R.; Ramos-Rincón, J.M. Non-alcoholic fatty liver disease is associated with bacterial translocation and a higher inflammation response in psoriatic patients. Sci. Rep. 2021, 11, 8593. [Google Scholar] [CrossRef]
- Childers, R.C.; Lucchesi, P.A.; Gooch, K.J. Decreased Substrate Stiffness Promotes a Hypofibrotic Phenotype in Cardiac Fibroblasts. Int. J. Mol. Sci. 2021, 22, 6231. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Qu, J.; He, L.; Zhu, Y.; Yang, S.Z.; Zhang, F.; Guo, T.; Peng, H.; Chen, P.; Zhou, Y. Stiff matrix instigates type I collagen biogenesis by mammalian cleavage factor I complex-mediated alternative polyadenylation. JCI Insight 2020, 5, e133972. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Cao, S.; Stowe, J.C.; Valdez-Jasso, D. Substrate Stiffness and Stretch Regulate Profibrotic Mechanosignaling in Pulmonary Arterial Adventitial Fibroblasts. Cells 2021, 10, 1000. [Google Scholar] [CrossRef]
- Chen, L.C.; Shibu, M.A.; Liu, C.J.; Han, C.K.; Ju, D.T.; Chen, P.Y.; Viswanadha, V.P.; Lai, C.H.; Kuo, W.W.; Huang, C.Y. ERK1/2 mediates the lipopolysaccharide-induced upregulation of FGF-2, uPA, MMP-2, MMP-9 and cellular migration in cardiac fibroblasts. Chem. Biol. Interact. 2019, 306, 62–69. [Google Scholar] [CrossRef]
- Rodriguez, W.E.; Tyagi, N.; Deng, A.Y.; Adeagbo, A.; Joshua, I.G.; Tyagi, S.C. Congenic expression of tissue inhibitor of metalloproteinase in Dahl-salt sensitive hypertensive rats is associated with reduced LV hypertrophy. Arch Physiol. Biochem. 2008, 114, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Purcell, B.P.; Barlow, S.C.; Perreault, E.P.; Freeburg, L.; Doviak, H.; Jacobs, J.; Hoenes, A.; Zellars, K.N.; Khakoo, A.Y.; Lee, T.W.; et al. Delivery of a matrix metalloproteinase-responsive hydrogel releasing TIMP-3 after myocardial infarction: Effects on left ventricular remodeling. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H814–H825. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xiao, Y.; Liu, J.; Feng, L.; Kang, Y.J. Copper-induced reduction in myocardial fibrosis is associated with increased matrix metalloproteins in a rat model of cardiac hypertrophy. Metallomics 2018, 10, 201–208. [Google Scholar] [CrossRef]
- Riikonen, T.; Westermarck, J.; Koivisto, L.; Broberg, A.; Kähäri, V.M.; Heino, J. Integrin alpha 2 beta 1 is a positive regulator of collagenase (MMP-1) and collagen alpha 1(I) gene expression. J. Biol. Chem. 1995, 270, 13548–13552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajshankar, D.; Wang, B.; Worndl, E.; Menezes, S.; Wang, Y.; McCulloch, C.A. Focal adhesion kinase regulates tractional collagen remodeling, matrix metalloproteinase expression, and collagen structure, which in turn affects matrix-induced signaling. J. Cell. Physiol. 2020, 235, 3096–3111. [Google Scholar] [CrossRef]
- Thannickal, V.J.; Lee, D.Y.; White, E.S.; Cui, Z.; Larios, J.M.; Chacon, R.; Horowitz, J.C.; Day, R.M.; Thomas, P.E. Myofibroblast differentiation by transforming growth factor-beta1 is dependent on cell adhesion and integrin signaling via focal adhesion kinase. J. Biol. Chem. 2003, 278, 12384–12389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, L.; Li, X.; Wu, X.; Hui, B.; Han, S.; Gao, J.; Li, Y.; Shi, J.; Zhu, H.; Zhao, B.; et al. Simultaneous deactivation of FAK and Src improves the pathology of hypertrophic scar. Sci. Rep. 2016, 6, 26023. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.M.; Ferrari, G.D.; Alberici, L.C.; Malaspina, O.; Moraes, K.C.M. Cellular and molecular effects of silymarin on the transdifferentiation processes of LX-2 cells and its connection with lipid metabolism. Mol. Cell. Biochem. 2020, 468, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.B.; Tang, X.L.; Zhao, J.; Fischer, A.G.; Wu, W.J.; Uchida, S.; Gumpert, A.M.; Stowers, H.; Wysoczynski, M.; Bolli, R. Epigenetically modified cardiac mesenchymal stromal cells limit myocardial fibrosis and promote functional recovery in a model of chronic ischemic cardiomyopathy. Basic Res. Cardiol. 2018, 114, 3. [Google Scholar] [CrossRef]
- Chen, W.; Yuan, H.; Cao, W.; Wang, T.; Chen, W.; Yu, H.; Fu, Y.; Jiang, B.; Zhou, H.; Guo, H.; et al. Blocking interleukin-6 trans-signaling protects against renal fibrosis by suppressing STAT3 activation. Theranostics 2019, 9, 3980–3991. [Google Scholar] [CrossRef]
- Kumar, S.; Wang, G.; Zheng, N.; Cheng, W.; Ouyang, K.; Lin, H.; Liao, Y.; Liu, J. HIMF (Hypoxia-Induced Mitogenic Factor)-IL (Interleukin)-6 Signaling Mediates Cardiomyocyte-Fibroblast Crosstalk to Promote Cardiac Hypertrophy and Fibrosis. Hypertension 2019, 73, 1058–1070. [Google Scholar] [CrossRef]
- Fujimura, T.; Moriwaki, S.; Imokawa, G.; Takema, Y. Crucial role of fibroblast integrins alpha2 and beta1 in maintaining the structural and mechanical properties of the skin. J. Dermatol. Sci. 2007, 45, 45–53. [Google Scholar] [CrossRef]
- Wang, J.H.; Zhao, L.; Pan, X.; Chen, N.N.; Chen, J.; Gong, Q.L.; Su, F.; Yan, J.; Zhang, Y.; Zhang, S.H. Hypoxia-stimulated cardiac fibroblast production of IL-6 promotes myocardial fibrosis via the TGF-β1 signaling pathway. Lab. Investig. 2016, 96, 839–852. [Google Scholar] [CrossRef] [PubMed]
- Ceccato, T.L.; Starbuck, R.B.; Hall, J.K.; Walker, C.J.; Brown, T.E.; Killgore, J.P.; Anseth, K.S.; Leinwand, L.A. Defining the Cardiac Fibroblast Secretome in a Fibrotic Microenvironment. J. Am. Heart Assoc. 2020, 9, e017025. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gałdyszyńska, M.; Radwańska, P.; Szymański, J.; Drobnik, J. The Stiffness of Cardiac Fibroblast Substrates Exerts a Regulatory Influence on Collagen Metabolism via α2β1 Integrin, FAK and Src Kinases. Cells 2021, 10, 3506. https://doi.org/10.3390/cells10123506
Gałdyszyńska M, Radwańska P, Szymański J, Drobnik J. The Stiffness of Cardiac Fibroblast Substrates Exerts a Regulatory Influence on Collagen Metabolism via α2β1 Integrin, FAK and Src Kinases. Cells. 2021; 10(12):3506. https://doi.org/10.3390/cells10123506
Chicago/Turabian StyleGałdyszyńska, Małgorzata, Paulina Radwańska, Jacek Szymański, and Jacek Drobnik. 2021. "The Stiffness of Cardiac Fibroblast Substrates Exerts a Regulatory Influence on Collagen Metabolism via α2β1 Integrin, FAK and Src Kinases" Cells 10, no. 12: 3506. https://doi.org/10.3390/cells10123506
APA StyleGałdyszyńska, M., Radwańska, P., Szymański, J., & Drobnik, J. (2021). The Stiffness of Cardiac Fibroblast Substrates Exerts a Regulatory Influence on Collagen Metabolism via α2β1 Integrin, FAK and Src Kinases. Cells, 10(12), 3506. https://doi.org/10.3390/cells10123506