
Evaluating the Diversity of Ecotypes of Red Clover (Trifolium pratense L.) from Northwestern Spain by Phenotypic Traits and Microsatellites


 ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Phenotypic Traits
2.2. Microsatellites
3. Results
3.1. Phenotypic Variation
3.2. Diversity by SSRs
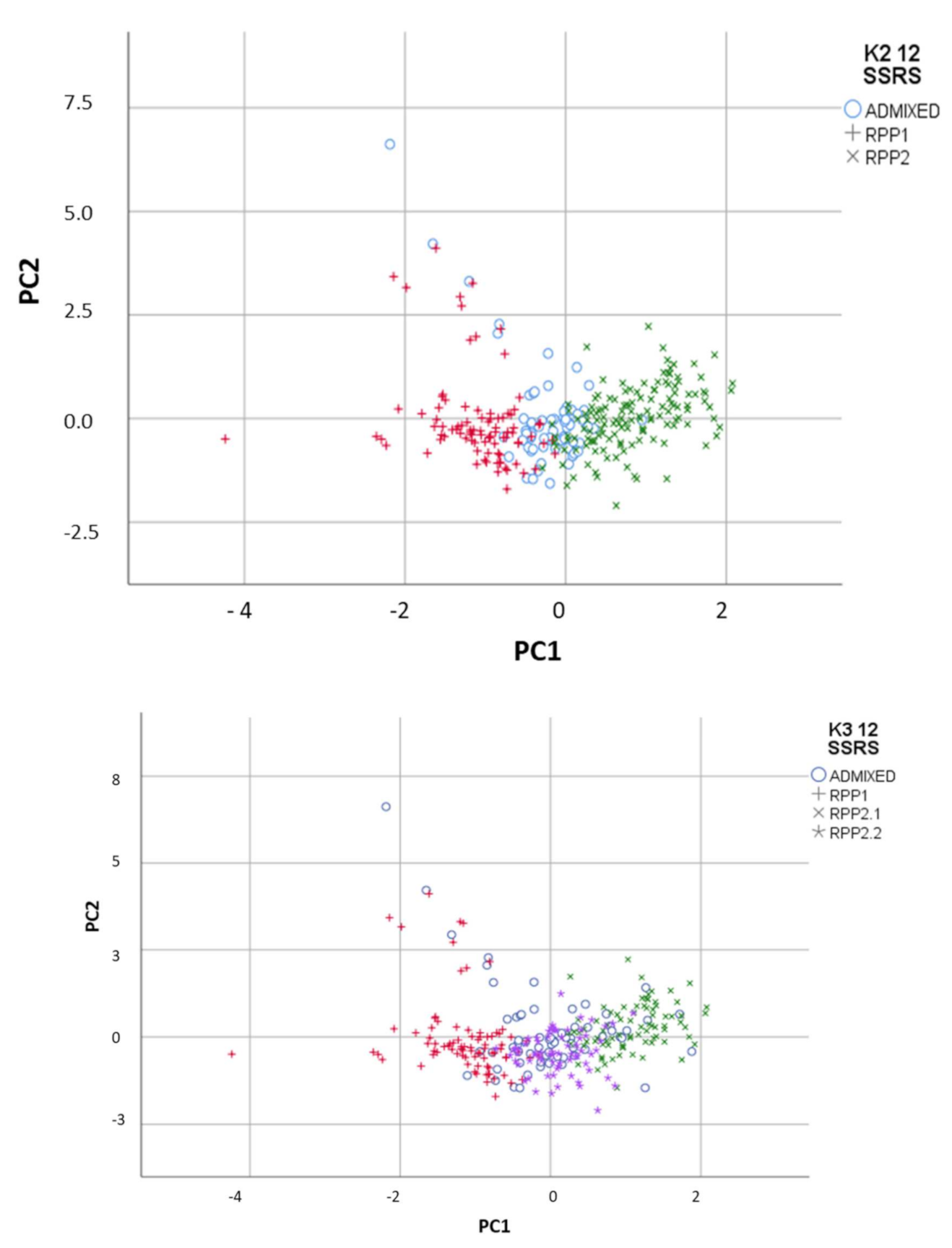
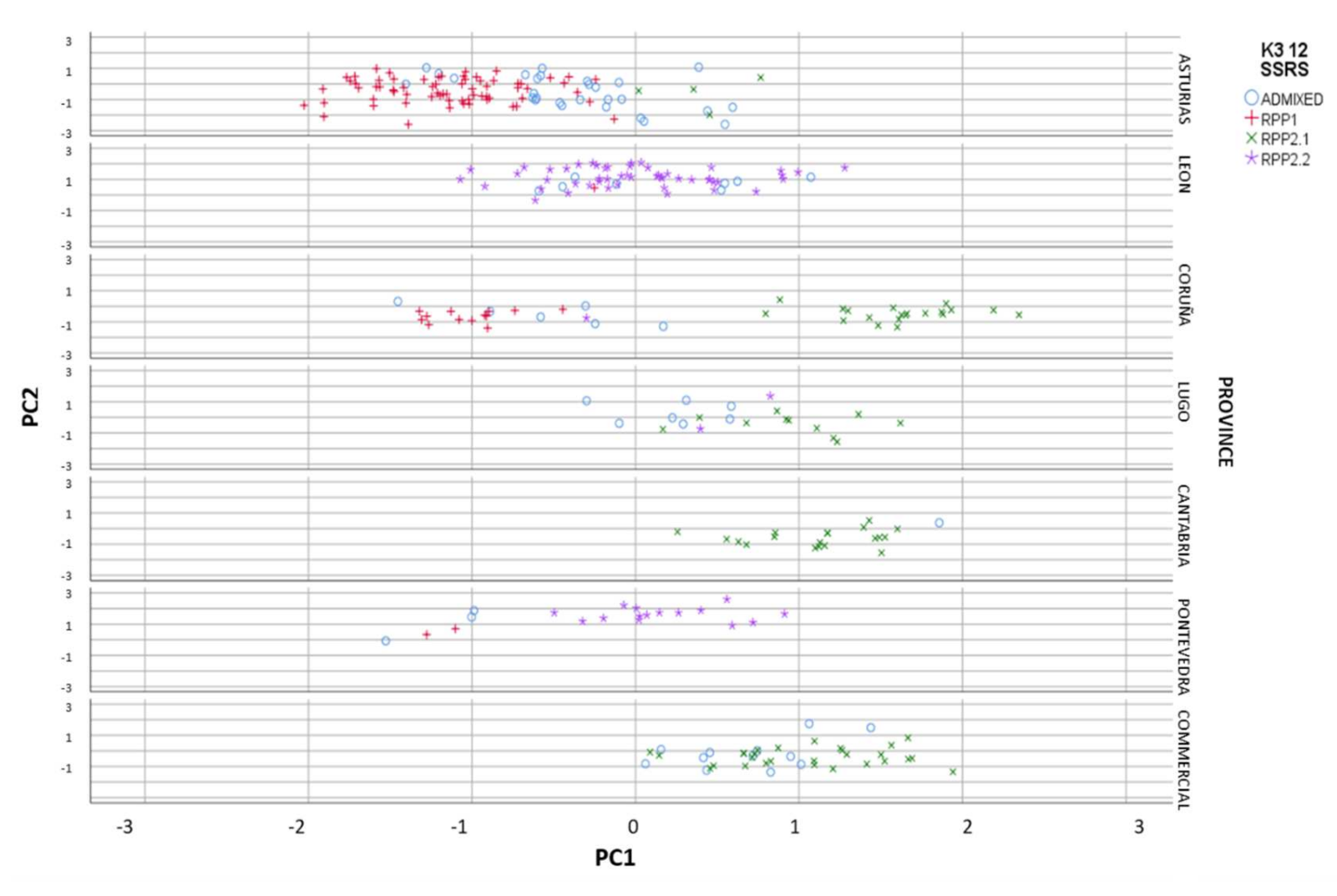
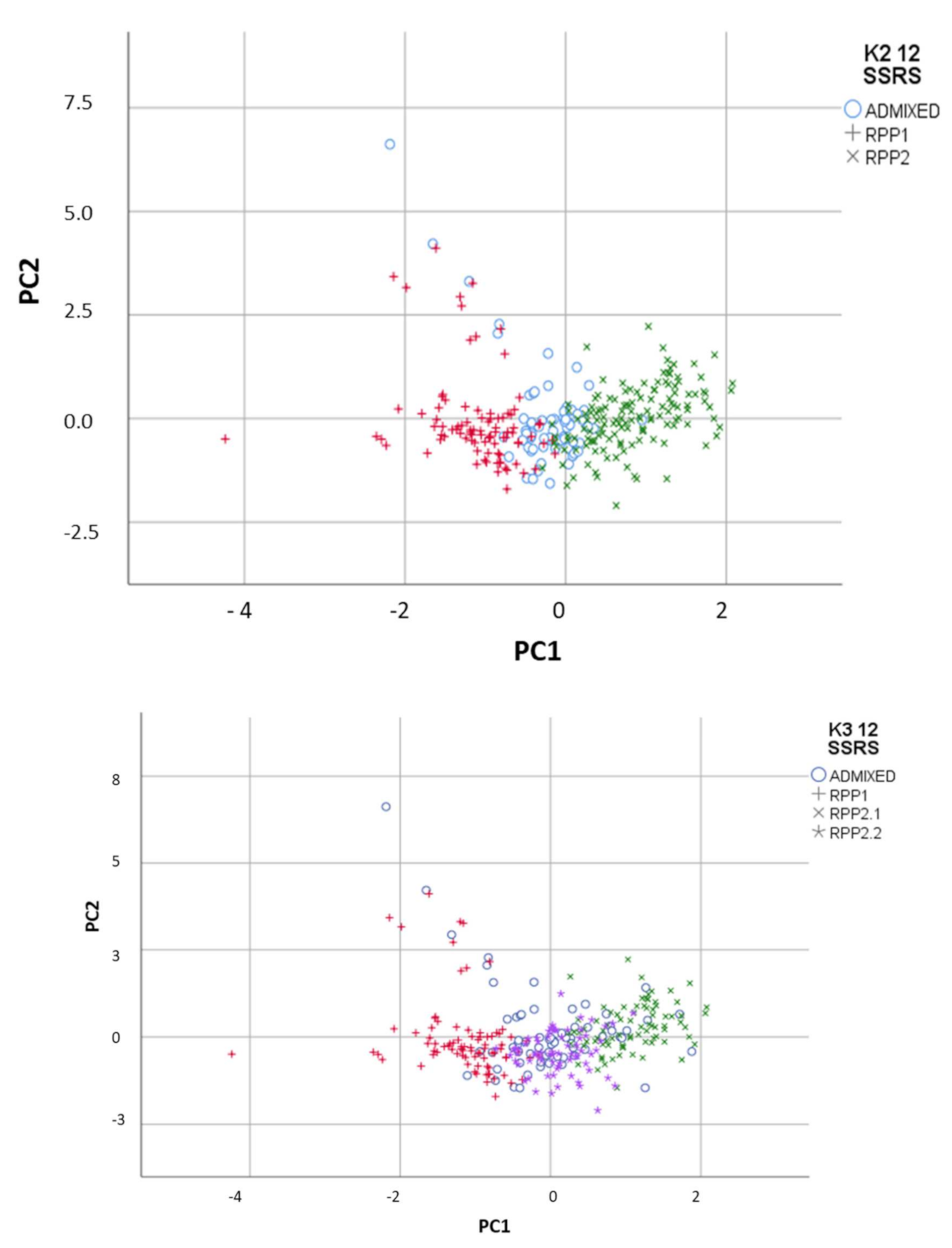
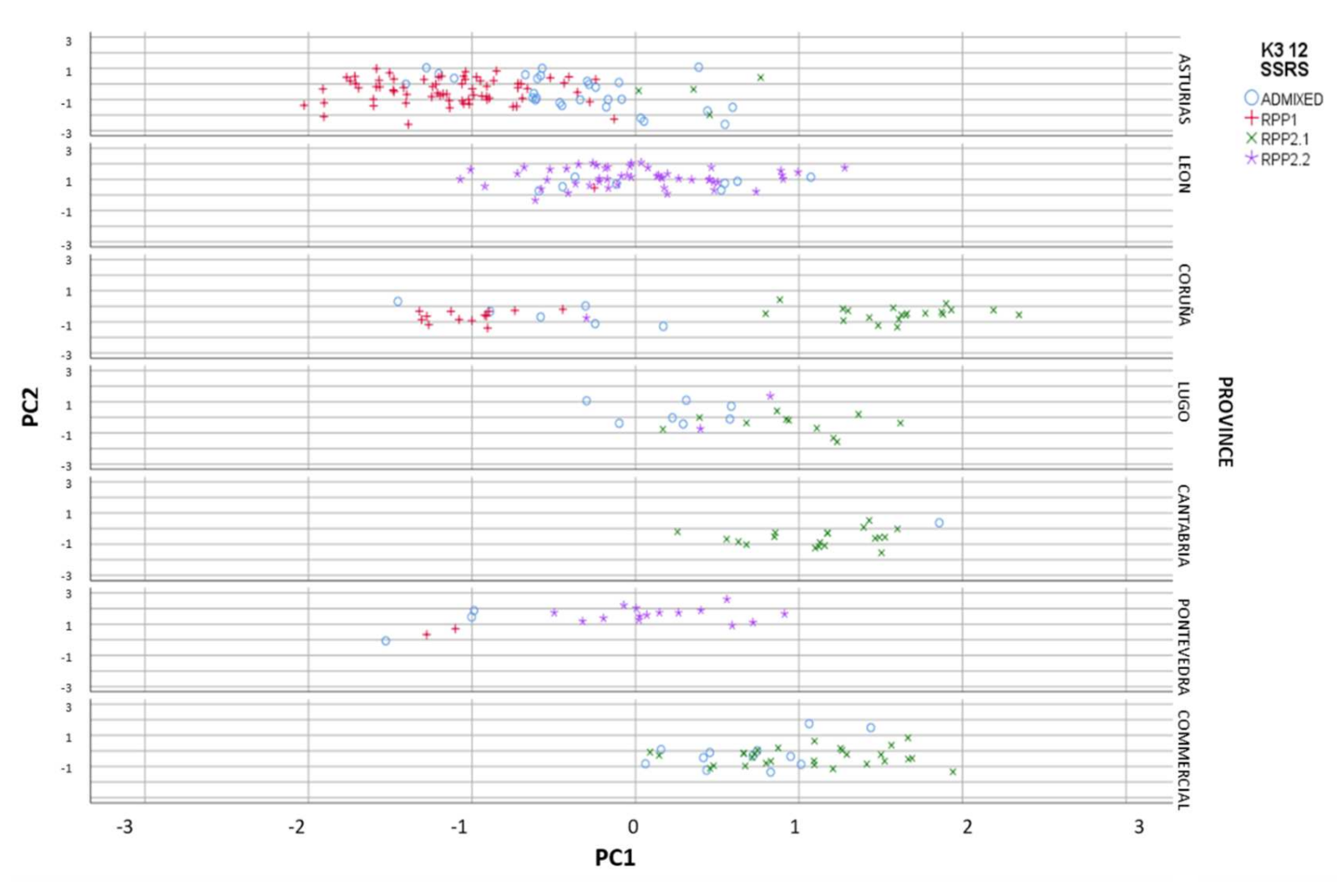
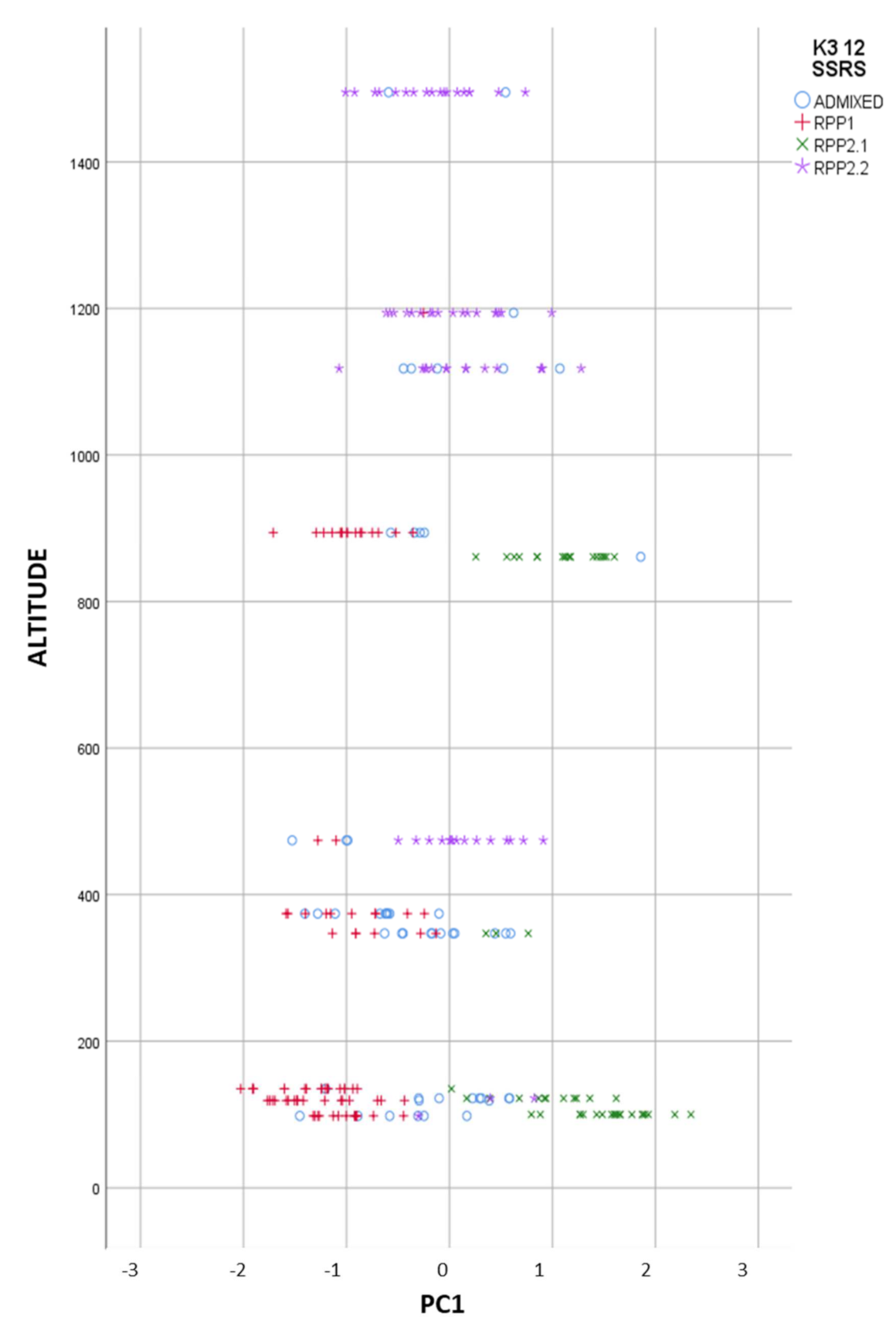
3.2.1. Genetic and Geographic Structure
3.2.2. Intra-Specific Variability
3.2.3. Factorial Component Analysis (FCA)
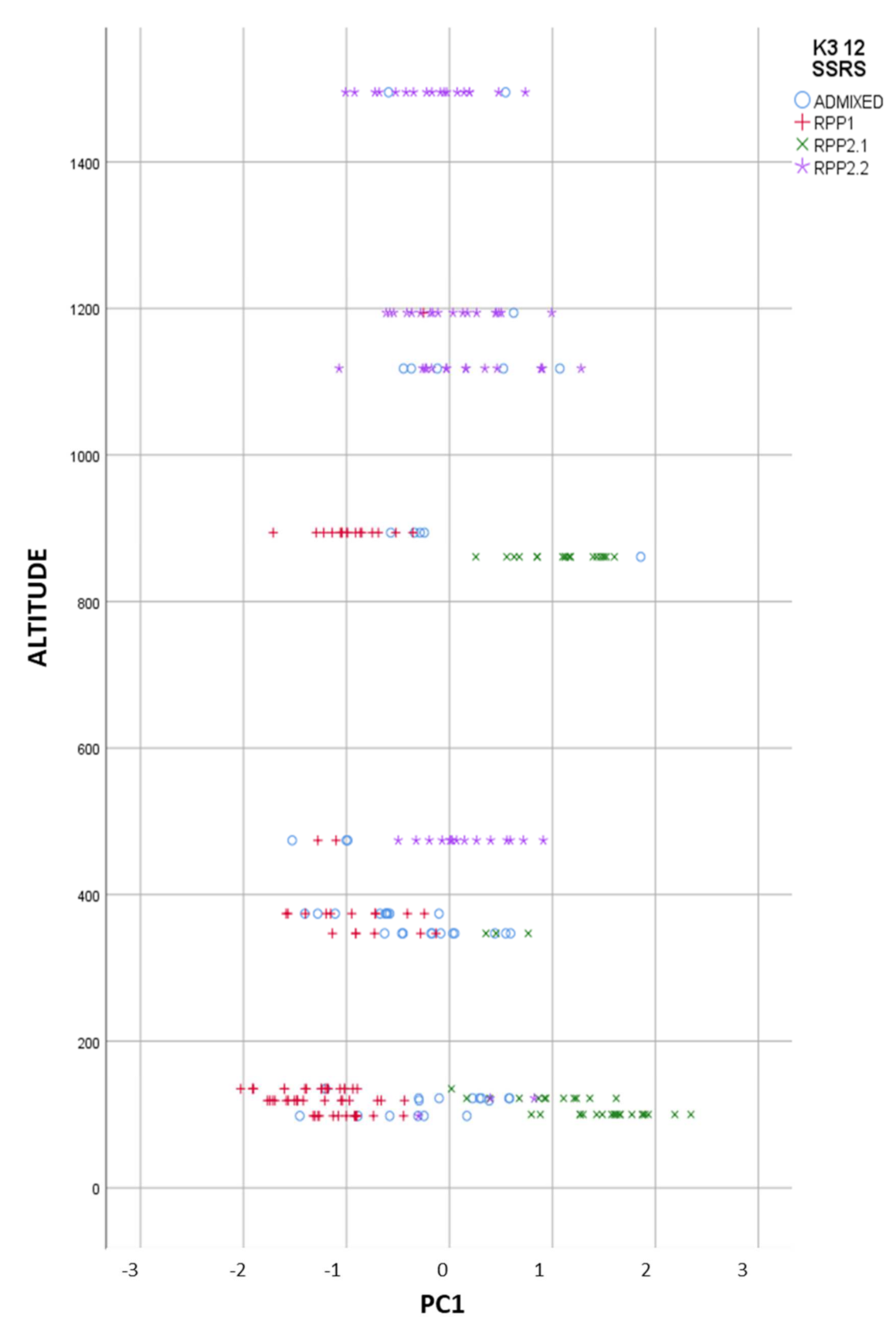
3.2.4. Genetic and Geographical Origin
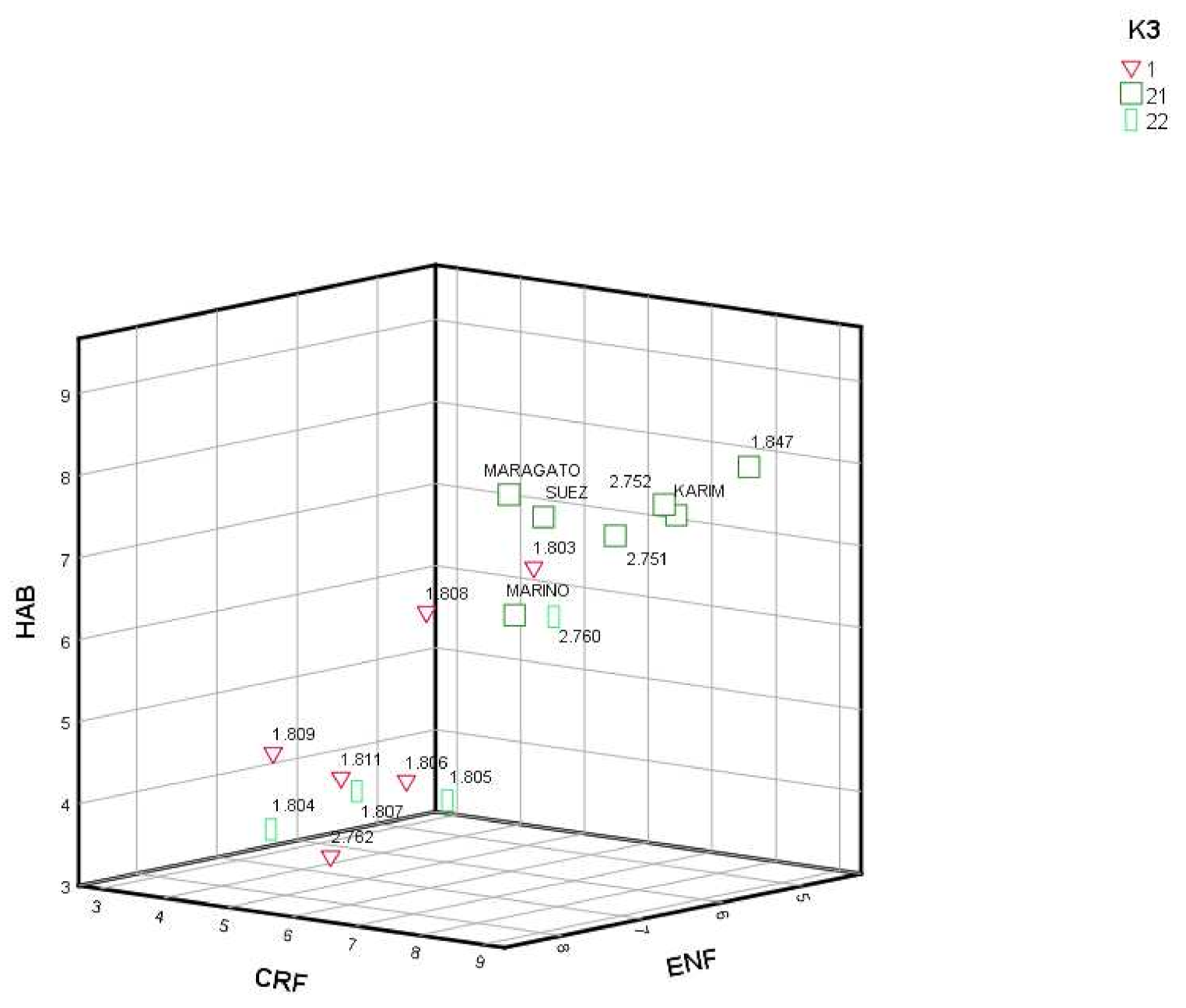
3.3. Genetic and Phenotypic Variation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bowley, S.R.; Taylor, N.L.; Dougherty, C.T. Physiology and morphology of red clover. Adv. Agron. 1984, 37, 317–347. [Google Scholar] [CrossRef]
- López, J.E.; González, E.; Oliveira, J.A.; Vázquez, O.P. Caracterización agronómica y cianogénica de poblaciones naturales de trébol blanco y trébol violeta recolectadas en la Cordillera Cantábrica. In Soberanía Alimentaria e Agricultura Ecolóxica: Propostas de Acción; II Congreso de Agroecoloxía e Agricultura Ecolóxica en Galiza; Universidade de Vigo: Vigo, Spain, 2010; pp. 865–883. [Google Scholar]
- IBPGR. Forage Legume Descriptors; International Board for Plant Genetic Resources: Rome, Italy, 1984. [Google Scholar]
- UPOV. Guidelines for the Conduct of Tests for Distinctness, Homogeneity and Stability; Red Clover; TG/5/4; International Union for the Protection of New varieties of Plants: Geneve, Swisse, 1985. [Google Scholar]
- Isobe, S.; Kölliker, R.; Hisano, H.; Sasamoto, S.; Wada, T.; Klimenko, I.; Okumura, K.; Tabata, S. Construction of a consensus linkage map for red clover (Trifolium pratense L). BMC Plant Biol. 2009, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Radinovic, I.; Vasiljevic, S.; Brankovic, G.; Ahsyee, R.S.; Momirovic, U.; Perovic, D.; Surlan-Momirovic, G. Molecular characterization of red clover genotypes utilizing microsatellite markers. Chil. J. Agric. Res. 2017, 77, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Isobe, S.; Asamizu, E.; Ohmido, N.; Kataoka, R.; Nakamura, Y.; Kaneko, T.; Sakurai, N.; Okumura, K.; Klimenko, I.; et al. Comprehensive structural analysis of the genome of red clover (Trifolium pratense L.). DNA Res. 2005, 12, 301–364. [Google Scholar] [CrossRef] [Green Version]
- Red Clover Marker Database. Available online: http://marker.kazusa.or.jp/Red_clover (accessed on 18 April 2019).
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Rosenberg, N.A.; Donnelly, P. Association mapping in structured populations. Am. J. Hum. Genet. 2000, 67, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Pereira-Lorenzo, S.; Ramos-Cabrer, A.M.; Barreneche, T.; Mattioni, C.; Villani, F.; Díaz-Hernández, B.; Martín, L.M.; Robles-Loma, A.; Cáceres, Y.; Martín, A. Instant domestication process of European chestnut cultivars. Ann. Appl. Biol 2019, 174, 74–85. [Google Scholar] [CrossRef] [Green Version]
- Porras-Hurtado, L.; Ruiz, Y.; Santos, C.; Phillips, C.; Carracedo, Á.; Lareu, M. An overview of STRUCTURE: Applications, parameter settings, and supporting software. Front. Genet. 2013, 4, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [CrossRef] [PubMed]
- Meirmans, P.G.; Van Tienderen, P.H. genotype and genodive: Two programs for the analysis of genetic diversity of asexual organisms. Mol. Ecol. Notes 2004, 4, 792–794. [Google Scholar] [CrossRef]
- Goodman, L.A. The analysis of multidimensional contingency tables when some variables are posterior to others: A modified path analysis approach. Biometrika 1973, 60, 179–192. [Google Scholar] [CrossRef]
- Llauradó, M.; Moreno-Gonzalez, J. Classification of Northern Spanish populations of maize by methods of numerical taxonomy. I. Morphological traits. Maydica 1993, 38, 15–21. [Google Scholar]
- Pereira-Lorenzo, S.; Fernández-Loópez, J.; Moreno-Gonzaález, J. Variability and grouping of northwestern Spanish chestnut cultivars. I. morphological traits. J. Am. Soc. Hort. Sci. 1996, 121, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Dugar, Y.N.; Popov, V.N. Genetic structure and diversity of Ukrainian red clover cultivars revealed by microsatellite markers. Open J. Genet. 2013, 3, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Dias, P.M.B.; Julier, B.; Sampoux, J.-P.; Barre, P.; Dall’Agnol, M. Genetic diversity in red clover (Trifolium pratense L.) revealed by morphological and microsatellite (SSR) markers. Euphytica 2008, 160, 189–205. [Google Scholar] [CrossRef]
- Soengas, P.; Cartea, M.E.; Francisco, M.; Lema, M.; Velasco, P. Genetic structure and diversity of a collection of Brassica rapa subsp. rapa L. revealed by simple sequence repeat markers. J. Agric. Sci. 2011, 149, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Joukhadar, R.; Daetwyler, H.D.; Bansal, U.K.; Gendall, A.R.; Hayden, M.J. Genetic Diversity, population structure and ancestral origin of Australian wheat. Front. Plant Sci. 2017, 8, 2115. [Google Scholar] [CrossRef] [Green Version]
- Atnaf, M.; Yao, N.; Martina, K.; Dagne, K.; Wegary, D.; Tesfaye, K. Molecular genetic diversity and population structure of Ethiopian white lupin landraces: Implications for breeding and conservation. PLoS ONE 2017, 12, e0188696. [Google Scholar] [CrossRef] [Green Version]
- Pereira-Lorenzo, S.; Dos Santos, A.; Ramos-Cabrer, A.M.; Sau, F.; Díaz-Hernández, M.B. Morphological variation in local pears from north-western Spain. Sci. Hortic. 2012, 138, 176–182. [Google Scholar] [CrossRef]
- Xiong, H.; Qin, J.; Shi, A.; Mou, B.; Wu, D.; Sun, J.; Shu, X.; Wang, Z.; Lu, W.; Ma, J.; et al. Genetic differentiation and diversity upon genotype and phenotype in cowpea (Vigna unguiculata L. Walp.). Euphytica 2018, 214, 4. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
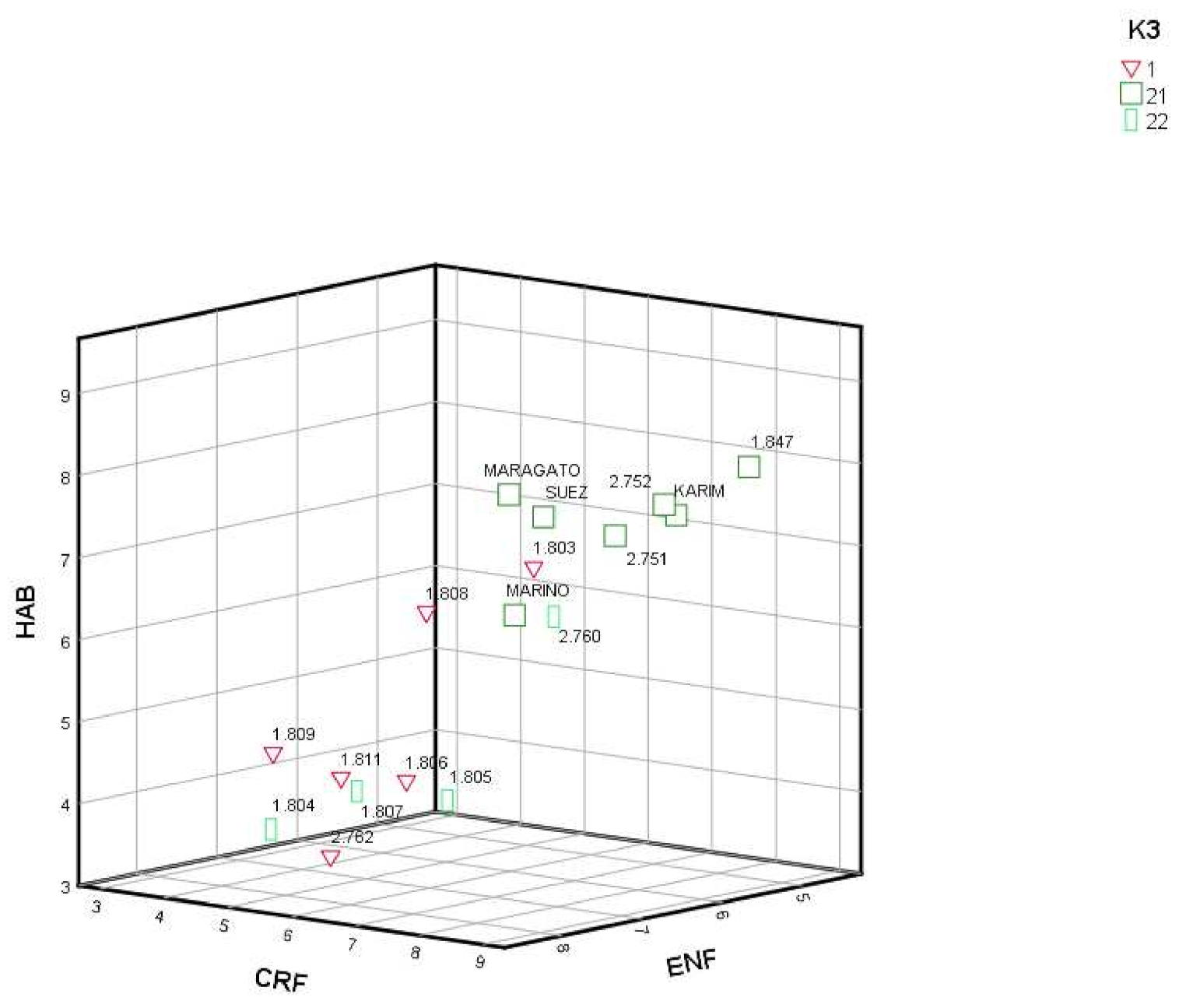
| Pop | K2 | K3 | FLO | CRE | CRF | HAB | ENF | PROVINCE (Origin) | ALTITUDE |
|---|---|---|---|---|---|---|---|---|---|
| 1803 | 1 | 1 | 120.60 | 4.92 | 5.89 | 6.62 | 5.62 | Asturias | 347 |
| 1804 | 2 | 22 | 140.08 | 4.02 | 3.58 | 3.45 | 7.05 | Leon | 1495 |
| 1805 | 2 | 22 | 142.14 | 3.75 | 4.31 | 3.56 | 5.44 | Leon | 1118 |
| 1806 | 1 | 1 | 147.75 | 4.26 | 4.21 | 3.88 | 5.87 | Asturias | 894 |
| 1807 | 2 | 22 | 138.15 | 3.74 | 3.76 | 3.75 | 6.13 | Leon | 1194 |
| 1808 | 1 | 1 | 98.83 | 4.80 | 6.53 | 6.52 | 7.46 | Asturias | 374 |
| 1809 | 1 | 1 | 101.50 | 3.97 | 4.93 | 4.75 | 8.10 | Asturias | 135 |
| 1811 | 1 | 1 | 93.97 | 4.18 | 5.00 | 4.29 | 7.31 | Asturias | 119 |
| 1847 | 2 | 21 | 142.53 | 4.43 | 8.14 | 7.92 | 4.73 | A Coruña | 100 |
| 2751 | 2 | 21 | 111.66 | 5.58 | 7.17 | 7.15 | 5.62 | Lugo | 122 |
| 2752 | 2 | 21 | 146.05 | 4.60 | 7.61 | 7.52 | 5.36 | Cantabria | 861 |
| 2760 | 2 | 22 | 146.97 | 6.23 | 5.59 | 5.88 | 5.13 | Pontevedra | 474 |
| 2762 | 1 | 1 | 136.92 | 4.36 | 5.14 | 3.40 | 7.55 | A Coruña | 98 |
| KARIM | 2 | 21 | 147.43 | 4.45 | 7.66 | 7.38 | 5.25 | France | |
| MARAGATO | 2 | 21 | 145.09 | 4.33 | 8.18 | 8.19 | 7.74 | Spain | |
| MARINO | 2 | 21 | 143.79 | 4.12 | 6.22 | 6.17 | 6.12 | Germany | |
| SUEZ | 2 | 21 | 148.18 | 3.11 | 6.93 | 7.49 | 6.32 | France |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Otero, C.I.; Ramos-Cabrer, A.M.; López-Díaz, J.E.; Pereira-Lorenzo, S. Evaluating the Diversity of Ecotypes of Red Clover (Trifolium pratense L.) from Northwestern Spain by Phenotypic Traits and Microsatellites. Agronomy 2021, 11, 2275. https://doi.org/10.3390/agronomy11112275
Fernández-Otero CI, Ramos-Cabrer AM, López-Díaz JE, Pereira-Lorenzo S. Evaluating the Diversity of Ecotypes of Red Clover (Trifolium pratense L.) from Northwestern Spain by Phenotypic Traits and Microsatellites. Agronomy. 2021; 11(11):2275. https://doi.org/10.3390/agronomy11112275
Chicago/Turabian StyleFernández-Otero, Cristina Isabel, Ana María Ramos-Cabrer, Julio Enrique López-Díaz, and Santiago Pereira-Lorenzo. 2021. "Evaluating the Diversity of Ecotypes of Red Clover (Trifolium pratense L.) from Northwestern Spain by Phenotypic Traits and Microsatellites" Agronomy 11, no. 11: 2275. https://doi.org/10.3390/agronomy11112275
APA StyleFernández-Otero, C. I., Ramos-Cabrer, A. M., López-Díaz, J. E., & Pereira-Lorenzo, S. (2021). Evaluating the Diversity of Ecotypes of Red Clover (Trifolium pratense L.) from Northwestern Spain by Phenotypic Traits and Microsatellites. Agronomy, 11(11), 2275. https://doi.org/10.3390/agronomy11112275