Abstract
We extend techniques and learnings about the stochastic properties of nonlinear responses from finance to medicine, particularly oncology, where it can inform dosing and intervention. We define antifragility. We propose uses of risk analysis for medical problems, through the properties of nonlinear responses (convex or concave). We (1) link the convexity/concavity of the dose-response function to the statistical properties of the results; (2) define “antifragility” as a mathematical property for local beneficial convex responses and the generalization of “fragility” as its opposite, locally concave in the tails of the statistical distribution; (3) propose mathematically tractable relations between dosage, severity of conditions, and iatrogenics. In short, we propose a framework to integrate the necessary consequences of nonlinearities in evidence-based oncology and more general clinical risk management.
1. Introduction: Where the Idea of Antifragility Came From
The notions of fragility and antifragility were inspired by the payoffs and the intricacies of financial derivatives. The concept was introduced in Taleb (2012) [1] and more formalized in Taleb and Douady (2013) [2]. While, in the real world, many phenomena are intuitively known to benefit from an increase in “volatility” (that is, the standard deviation of a random variable, or the variability of a nonrandom one), only quantitative finance had names for such attributes, such as “long gamma” (where the financial “derivative” contract has a positive local second mathematical derivative with respect to the underlying security), “long vega” (where the financial derivative has a positive first derivative with respect to the standard deviation of the underlying security), and similar measures, always associated with some range of variation as these sensitivities are local and have, themselves, higher derivatives. By “local” we refer to the fact that most payoff functions in finance are convex over a certain range, then linear or concave, with the second derivative changing in sign, the so-called “higher Greeks” in [3]) Furthermore, finance links some nonlinear attributes of portfolios to the risk of “blowups”, that is, a loss large enough to be irreversible, such as irrecoverable financial ruin. Such quantitative and qualitative models of ruin can give us a tractable generalizable definition of fragility. However, centrally, derivative risk management, at its core, lies in distinguishing between the properties of a random variable X and a payoff function , almost always nonlinear.
While Jensen’s inequality (on which more in Appendix B) is concerned with the first moment of the distribution, monotone convex (or concave) functions, and a static expectation operator, financial payoffs are more complicated; the first static moment, while relevant, is not the sole focus as:
- The expectation must be conditioned on absence of “blowup”, that is, the left tail of the distribution must be constrained (see Geman et al., 2015) [4], which involves all higher moments of .
- The payoff functions are almost never monotone.
- Taking into account higher moments of the distributions is analogous to going beyond second-order effects: third, fourth, etc.
Fragility, as defined in Taleb (2012) and Taleb and Douady (2013) [1], Ref. [2], is related to how a system suffers from the variability of its environment beyond a certain preset threshold (when the threshold is K, it is called K-fragility), see Figure 1, while antifragility refers to when it benefits from this variability—in a similar way to as what we saw quantitative finance calls “vega” for an option or a nonlinear payoff, that is, its sensitivity to volatility or some similar measure of scale of a distribution. (Tail fragility maps to a risk of financial ruin, while local fragility does not necessarily mean ruin). In [2]:
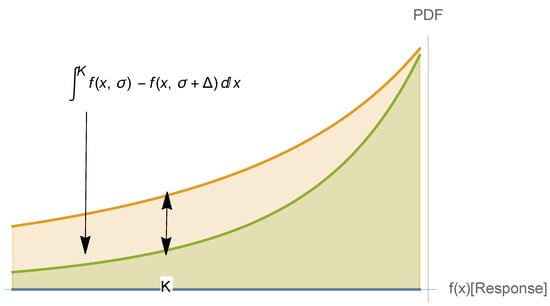
Figure 1.
Fragility below level K as indicative of survival. It is not quite symmetric because global antifragility is conditioned on tail robustness (“to do well, one must first survive”). The Taleb and Douady (2013) [2] paper shows that the gap between and , where is the scale of the distribution, is proportional to the concavity of . Hence, without knowing the distribution (PDF above), one can gauge such an effect by looking at the nonlinearity of below the threshold K.
Simply, a coffee cup on a table suffers more from large deviations than from the cumulative effect of some shocks—conditional on being unbroken, it has to suffer more from “tail” events than regular ones around the center of the distribution, the ‘at-the-money’ category. This is the case of elements of nature that have survived: conditional on being in existence, then the class of events around the mean should matter considerably less than tail events, particularly when the probabilities decline faster than the inverse of the harm, which is the case of all used monomodal probability distributions. Further, what has exposure to tail events suffers from uncertainty; typically, when systems—a building, a bridge, a nuclear plant, an airplane, or a bank balance sheet—are made robust to a certain level of variability and stress but may fail or collapse if this level is exceeded, then they are particularly fragile to uncertainty about the distribution of the stressor, hence to model error, as this uncertainty increases the probability of dipping below the robustness level, bringing a higher probability of collapse. In the opposite case, the natural selection of an evolutionary process is particularly antifragile, indeed a more volatile environment increases the relative survival rate of robust species and eliminates those whose superiority over other species is highly dependent on environmental parameters.
The paper above produced theorems linking the second derivative of in some ranges of variation to sensitivity to the scale of the distribution of X. Thus, the same sensitivity to the scale of the distribution can also express sensitivity to a stressor (dose increase) in medicine or other fields in its effect on either tail. Thus, one single measure will allow us to express with comfortable precision the exposure to the disorder cluster: (i) uncertainty, (ii) variability, (iii) imperfect, incomplete knowledge, (iv) chance, (v) chaos, (vi) volatility, (vii) disorder, (viii) entropy, (ix) time, (x) the unknown, (xi) randomness, (xii) turmoil, (xiii) stressor, (xiv) error, and (xv) dispersion of outcomes. A positive (negative) sensitivity to one means positive (negative) to all others in the group. Finally—and critically—the paper showed that one does not need to have an exact probability distribution to obtain a useful idea of the exposure, since these metrics are based on acceleration, which washes out up to one order of magnitude the precision errors. Note that for multivariate situations, an additive approach is used without any loss of effectiveness.
Asymmetry of Fragile/Antifragile: The opposite of globally fragile (with respect to a random variable X) is not naively “antifragile”, but is both convex with respect to that variable and has a left tail constraint. In probabilistic representation, must have a positively skewed distribution. Furthermore, as with the fragile, antifragility is limited to a specific range of variations, and with respect to a single random variable.
The rest of this article will present medicine and convexity, then apply the notions of fragility–antifragility at two levels: efficient dosing in oncology and an examination of iatrogenics as linked to convexity. Finally, in the appendix, we present an overview of convex responses in medicine.
Note: we use, by convention, the term “convexity” or “convexity effect” in the presence of consequential nonlinearity, which can be concave: if the harm function is defined as positive, it shows as convex; if negative, it shows as concave. Finance uses the expression “convexity bias” for both concave and convex responses (and with the possible additional designation “positive” or “negative” convexity).
2. Medicine and Convexity
Medicine has much simpler payoffs than quantitative finance. Most are generalizations around simple sigmoids, see Figure 2, which were described in the mapping in Taleb and Douady (2013) [2] as belonging to the benign class: the distribution of is necessarily thinner-tailed than that of X, owing to the boundedness of the function, dubbed more “binary” than “vanilla”, see [5].
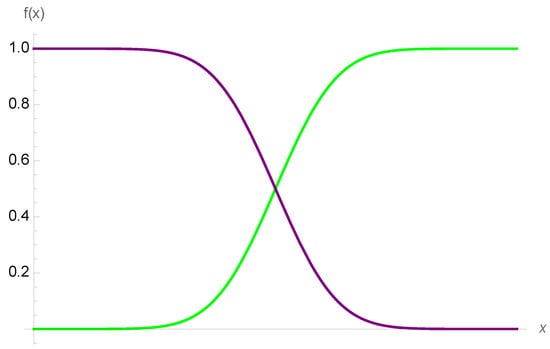
Figure 2.
Simple (first-order) nonincreasing or nondecreasing sigmoids, defined as floored and capped increasing functions. They map to the payoff in finance of a binary option with time left to expiration. As the sigmoid loses smoothness (with the decreased time to expiration), it becomes, at the limit, a Heaviside function, see Figure 3.
However, in spite of such simplicity, little work in medicine has been conducted about probabilistic effects on convexity—almost always limited to first-order effects and comparative statics. The probabilistic dimension of variability has been made explicitly in some medical domains, for instance, there are a few studies connecting Jensen’s inequality to patient responses with pulmonary ventilators: papers such as Brewster et al. (2005) [6], Amato et al. [7], Funk (2004) [8], Arold et al. (2003) [9], Graham et al. (2005) [10], and Mutch et al. (2007). To summarize the literature, continuous high pressures have been shown to be harmful (leading to increased mortality), but episodic spikes of ventilation pressures can be helpful with the recruitment of collapsed alveoli (natural breathing exhibits some variability, with some breaths deeper than others). However, these papers stop at Jensen’s inequality, and, further, explicit probabilistic formulations are still missing in other domains where the applications of these techniques are most needed, such as intermittent fasting, episodic energy deficit, uneven distribution of sub-groups (say, proteins), vitamin absorption, moderate- and low-intensity training, fractional dosage, the comparative effects of low-intensity and distributed interventions vs. intense and concentrated ones, the chronic vs. the acute, and similar effects. As to the psychology literature, the notion of overcompensation is present Den Hartigh and Hill (2022) [11]. The identification of convexity is still confined to local responses and did not generalize to decision-making under uncertainty and inferences concerning silent risks from the nonlinearity in dose response. For instance, the results did not reach the obvious relation between tumor size and the trade-offs of the intervention, or the extrapolation between the numbers needed to treat (NNT) and the potential severity of the side effects.
The connections we are investigating are necessary and mathematical: they work in both directions. We can illustrate as follows:
- A convex response to energy balance over a fixed time window necessarily implies gains from intermittent fasting in some situations and under some strict conditions (that is, higher variance in the distribution of nutrients) over some range within the limits of that time window;
- The presence of metabolic problems in populations that have a steady supply of food intake, as well as evidence of human fitness to an environment that provides moderate variations in the availability of food, both necessarily imply a concave response to food within a range and time frame.
Finally, a short summary of the above is as follows. Convexity analysis in medicine is in two dimensions; first, working with the nonlinearity of dosing, second, for risk analysis for patients and groups.
Missing second-order effects: One frequent lacuna in the literature is ignoring the second-order effects when making statements derived from from empirical data. One example is dietary recommendations for food group composition rather than frequency. Epidemiological interpretations of the Cretan diet relied solely on composition. A simplified intuition of the second-order effect in nutrition is as follows. Eating once a day vs., say, three times (in an isocaloric way) presents a difference if the response function is nonlinear: an average of response functions is not a response function of an average. However, frequency matters: the Eastern Orthodox Church has, with minor local variations, around two hundred days of vegan fasts per year. This is an episodic protein deprivation; fatty meats are consumed in lumps (on Sundays and holidays), which compensates for such a deprivation (recall the threshold in Figure 4. As shown in the literature review in Appendix A, there is a need for a mathematical bridge between studies of variability, say Martin et al. (2006) [12] and Fontana et al. (2008) [13], on one hand, and the focus on composition—the Longo and Fontana studies, furthermore, narrow the effect of the frequency to a given food type, namely proteins (Lee and Longo (2011) [14] “In the prokaryote E. coli, lack of glucose or nitrogen (comparable to protein restriction in mammals) increase resistance to high levels of (15 mm) (Jenkins et al., 1988) [15]”). Further, the computation of the “recommended daily” units may vary markedly if one assumes necessary stochasticity.
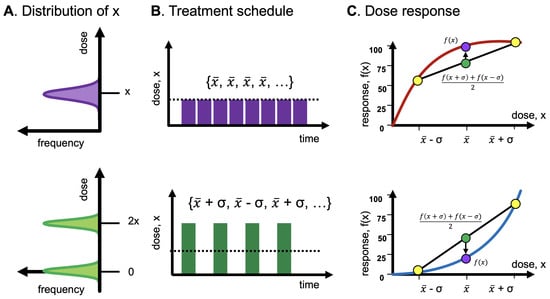
Figure 4.
Example treatment-scheduling protocols. (A) Input distribution of dosing is typically unimodal (“even”) or bimodal (“uneven”). (B) Protocols are typically fixed, with doses administered at regular intervals. It may be feasible to temporarily increase the dose (green), with periodic treatment holidays. (C) Even treatment is optimal to maximize response for concavity; uneven for convexity.
Extracting past statistical attributes and frequencies: A central question is if we need a certain dose of stressors, whether in intermittence of nutrition or necessary exercise, might these represent the attributes of an “ideal” environment. Whether evolutionary or not, this is the one to whose stochastic properties we are most adapted. We can, therefore, reverse engineer the stochastic nature of such an “ideal” environment by finding the various conditions that result from a reduction in stressors. We noted that papers such as Kaiser (2003) [16] and Calabrese and Baldwin (2003), [17] do not bridge the results to the point that hormesis may correspond to a “fitness dose”, beyond and below which one departs from such an ideal dispersion of the dose x per time period.
Such a reverse engineering uses the visible dose–response curve to make inferences about the ideal parametrization of the probability distribution of nutritional balance and vice-versa. For example, assessing the benefits of episodic fasting and the length of windows for neoplasms, insulin resistance, and other conditions can lead to understanding some kind of “fitness” to an environment endowed with a certain structure of randomness, either with the above or some more sophisticated attributes of the probability distributions (such as higher moments, hence different shapes). For example, if insulin resistance can be reduced thanks to occasional deprivation (a certain variance), say one 24-h fast every week, 3 days of fasting per trimester, and a complete week every five years, then we can extract and parametrize a probability distribution of ancestral deprivations. A comprehension of the exact mechanism by which such intermittences work can be helpful but is not needed given the robustness of the mathematical connection between the functional and probabilistic.
Antifragility in Treatment Scheduling
In Figure 5, the input distribution of dose x is subject to the convexity (or concavity or linearity) of the dose-response function, which influences the tail of the outcome distribution. Importantly, an oncologist has “first-mover” advantage [18] and has the benefit of prescribing an “even” treatment protocol with no variance (Figure 4, top row) or an “uneven” treatment protocol with positive variance (Figure 4, bottom row).
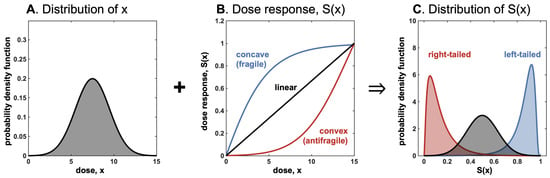
Figure 5.
These three graphs (related to the convex (concave) transformations of random variables) summarize and simplify our main idea; they show how we can go from the reaction or dose response , combined with the probability distribution of x, to the probability distribution of and its properties: mean, expected benefits or harm, variance of . Thus, we can play with the various parameters that can affect and those that can affect the distribution of x, and extract results from the output. , as we show, can take different forms (we chose a monotone convex or concave , but a second-order mixed sigmoid can also be used).
In medical practice, treatment protocols are typically fixed with doses administered at regular intervals (e.g., Figure 4). The distribution of dosing is unimodal (purple; continuous dosing), or, at most, bimodal (green; intermittent dosing). Manipulation of dose volatility when designing treatment protocols is under-utilized as a strategy in cancer treatment. In place of a dose x, one can give, say, 120% of x, then 80% of x, with a more favorable outcome if one is in a zone that benefits from unevenness. If antifragile, more unevenness is more beneficial: 140% followed by 60% produces better effects [19].
3. Antifragility in Oncology
Across all treatment modalities, cancer treatment is intended to induce perturbations to environmental conditions within a tumor leading to cell death, altering vasculature, or impacting immune response. However, the most common treatment paradigm is the “maximum tolerable dose” (MTD) dosing protocol, whereby the dose is maximized, and only limited by tolerability, toxicity, and side effects. To re-phrase, oncology research is implicitly focused on maximizing “first-order” treatment effects by increasing the cumulative dose [20] or shortening the time between doses [21]. The “log-kill” law proposes an MTD protocol for cytotoxic chemotherapy agents that decreases the amount of time over which a cumulative dose is delivered as toxicity allows [22]. More recently, metronomic therapy proposes frequent, low doses known to provide an anti-angiogenic effect during chemotherapy, still implicitly optimizing a first-order effect of cumulative dose [23].
Oncology must consider convexity in strategizing treatment protocols. Although convexity was not a consideration of initial clinical design, recent approaches have had success in managing second-order effects through the practice of high/low dosing. Intermittent high dosing of tyrosine kinase inhibitors (TKI) in HER2-driven breast cancers was administered with concentrations of the drugs that would otherwise far exceed toxicity thresholds if dosed continuously [24]. Continuous letrozole in combination with high-dose intermittent ribociclib is currently in clinical trial (NCT02712723; ER-positive breast cancer) [25]. Intermittent high-dose erlotinib delays resistance in an EGFR-mutant non–small cell lung cancer in vivo model [26,27]. Intermittent weekly EGFR-inhibitors reduced tumor load in vivo, compared with daily regimens with identical cumulative doses [28]. Intermittent “pulsatile” high-dose erlotinib once weekly maintains efficacy even after failure of low-dose continuous treatment [29]. Ideal treatment protocols will maximize both first-order effects (cumulative dose) and second-order effects (variance of dose delivered). Studies mentioned previously provide evidence of the tolerability of temporary dose escalation by also employing off-treatment periods to alleviate therapy toxicity.
3.1. Defining (Local) Fragility in Oncology
Local fragility, F, is a measurable quantity, similar to the Jensen gap, defined as the difference in the result of unevenness over evenness (with corresponding unevenness range parameter ):
Which property of cancer cells, , is important in oncology? Here, we are interested in the advantage of “uneven” high/low schedule over the “even” schedule. More precisely, fragility is the difference between (1) a schedule of two constant doses (termed an “even” dosing strategy), and (2) a schedule of a high dose followed by a low dose (termed an “uneven” dosing strategy), . We consider the response to two doses over an interval of time T, where the first dose is given at and the second dose is given at . Given a tumor with an initial population size of , the final size of an exponentially growing population is given by:
where is the decay rate of the population associated with a dose of x. Fragility can be defined as:
which simplifies to:
We are interested in the antifragile–fragile boundary, the point at which the population is no longer fragile but antifragile. If , the cell population is antifragile by definition (a benefit conferred to uneven dosing for minimizing tumor growth rate), and if , then it is fragile. It follows that the previous equation will be negative if:
Thus, the important domain for convexity is the dose-dependent growth rate, . It is an unfortunate common practice to normalize dose–response curves to obtain fractional survival at the final time point (e.g., ), which obscures convexity. Drug-induced growth rate inhibition (GR curves) has been introduced as a method to remove the artifactual dependency of IC50 and Emax on the cellular division rate [30]. GR curves preserve convexity, unlike fractional survival.
3.2. Fragility and Taylor Series Approximations
Antifragility is a “second-order” effect. This is shown by first taking a Taylor expansion about x:
where represents all third-order or higher terms. Using this expansion, fragility can be written:
The zeroth-order terms and the first-order terms cancel out:
For small values of , fragility is proportional to the second derivative, and, thus, known as a second-order effect. Next, we connect the concept to finite difference methods.
3.3. Fragility and Finite Differences
We can approximate the derivative of using finite differences. Here, we use the well-known central difference approximation to the second derivative, , where h is the width of the interval over which the finite difference is estimated ():
The term on the right-hand side in brackets is related to fragility (Equation (1)), giving us the relationship between and :
This limit illustrates that approaches F only when h is small. When the value of h is large, the approximation is poor, and, therefore, we employ Equation (1). Figure 6 provides an example of approximation error for the Hill function (see next section).
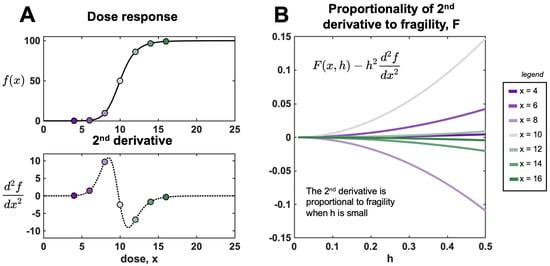
Figure 6.
The second derivative is an approximation for fragility for low values of h. (A) Hill function, (Equation (11)) shown for , , , and . Analytically derived second derivative (Equation (12)) is shown in the bottom panel. (B) Difference between fragility and second derivative at various dose values (red to blue) corresponding to panel A. As , the error approaches zero: .
3.4. Applications of Hill Function
The Hill function is commonly used to describe drug pharmacodynamics [31], where is the cell viability in response to a dose x.
where n is the Hill shape parameter, and are the minimal and maximal response (respectively), and C is the half-maximal response (the EC50 value). The second derivative can be written:
Figure 6A shows a sample Hill function and corresponding second derivative. Figure 6B illustrates decreasing error between the numerical (F) and analytical () as . The error scales like , as predicted in Equation (10). The inflection point, found where , defines the boundary between convex and concave regions.
It can be shown that as n increases, . Importantly, determines the boundary between the antifragile and fragile regions of . Benefit can, thus, be derived from uneven dosing if . The inverse implies that uneven treatment schedules provide no additional benefit. For a discussion on relaxing the assumption of fixed treatment schedules (e.g., Figure 4) using a probability density function describing dose distribution, see Appendix C, Equations (A4) and (A5).
As shown in Figure 5, the input distribution passing through convex dose response (e.g., on the Hill function below its inflection point) results in a left-tailed outcome distribution, and a concave response function results in a right-tailed distribution.
Thus, we have defined as locally antifragile a situation in which, over a specific interval , either the expectation rises with the scale parameter of the probability distribution as in Equation (A2), or the dose response is convex (on average) over the same range. The designation in Taleb (2012) [1] meant to accurately describe such situations: anything that gains from an increase in stochasticity or variability (since the scale parameter represents both). Terms such as “resilience”, since they were not mapped mathematically, are vague and even confusing as they meant either resistance or gains from stressors, depending on context. Figure 7 and Figure 8 illustrates the threshold effect of the asymmetric response, and gives the intuition of how they can be described as as antifragile.
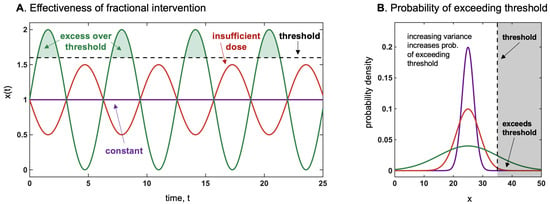
Figure 7.
(A) How a fractional intervention is more effective to surpass a threshold than a constant dosage of the same average. This is akin to stochastic resonance (in physics), by which the presence of noise causes the signal to rise above the detection threshold. For instance, genetically modified BT crops produce a constant level of pesticide, which appears to be much less effective than occasional manual interventions to add doses to conventional plants. The same may apply to antibiotics, chemotherapy, and radiation therapy [32]. (B) How more variance impacts the exceedance over the threshold. If threshold ≥ mean, we have convexity, and the variance increases the payoff more than variations in the mean. Such an effect is proportional to the remoteness of such threshold. Note that the harm function is defined as positive.
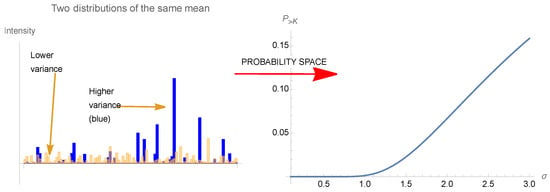
Figure 8.
(Left) A time series illustration of how a higher variance (hence scale), given the same mean, allow more spikes, hence an antifragile effect. We have random paths of two gamma distributions of the same mean, different variances, and , showing higher spikes and maxima for . The effect depends on the norm , more sensitive to tail events, even more than just the scale which is related to the norm . (Right) Representation of antifragility of (Left) in distribution space: we show the probability of exceeding a certain threshold for a variable, as a function of , the scale of the distribution, while keeping the mean constant.
3.5. The First-Order Sigmoid Curve
Next, we outline the variety of sigmoids as catalogued by [33]. Define the sigmoid or sigmoidal function as having membership in a class of function , , with additional membership in the class (twice differentiable), monotonic nonincreasing or nondecreasing, that is let be the first derivative with respect to x: for all x or . Thus, we have:
which can of course be normalized with and if S is increasing, or vice versa, or alternatively and if S is increasing. We can define the simple (or first-order) sigmoid curve as having equal convexity in one portion and concavity in another: if .
Now, all functions starting at zero will have three possible properties at inception, as in Figure 9 and Figure 10: concave, linear, and convex. The point of our discussion is the latter becoming sigmoid. Although few medical examples appear, under scrutiny, to belong to the first two cases, one cannot exclude them from analysis. We note that given that the inception of these curves is zero, no linear combination can be initially convex unless the curve is convex, which would not be the case if the start of the reaction is at a level different from zero.
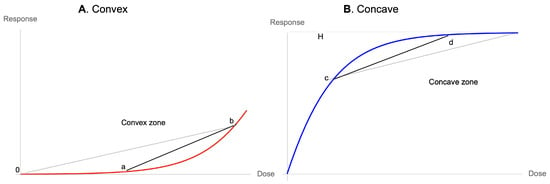
Figure 9.
(A) Every (relatively) smooth dose response with a floor has to be initially convex, hence prefers variations. (B) Every (relatively) smooth dose response with a ceiling has to be concave while approaching the ceiling, hence prefers stability.
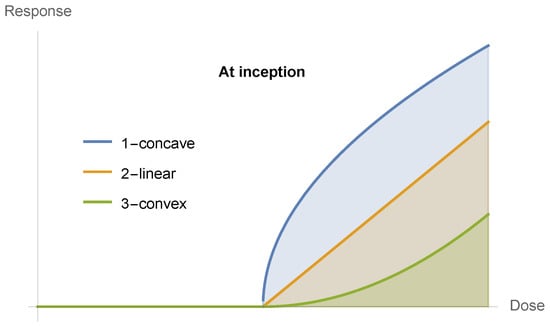
Figure 10.
The three possibilities at inception.
There are many sub-classes of functions producing a sigmoidal effect. Examples include:
- Pure sigmoids with smoothness characteristics expressed in trigonometric or exponential form, :
- Gompertz functions (a vague classification that includes above curves but can also mean special functions).
- Special functions with support in such as the error function
- Special functions with support in , such aswhere is the beta regularized function.
- Special functions with support inwhere is the gamma regularized function.
- Piecewise sigmoids, such as the CDF of the Student distribution
We note that the “smoothing” of the step function, or Heaviside theta produces a sigmoid (in a situation of a distribution or convoluted with a test function with compact support), such as , with , see Figure 3.
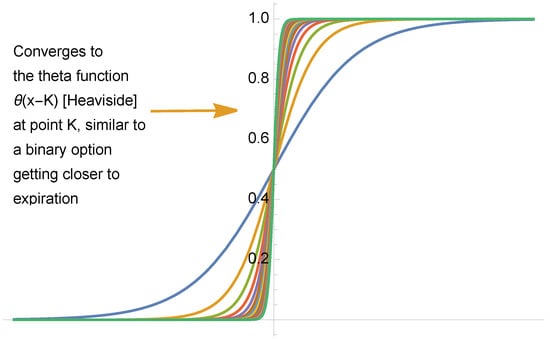
Figure 3.
The smoothing of the Heaviside function as distribution or Schwartz function.
3.6. Some Necessary Relations Leading to a Sigmoid Curve
Let , , of class be the first-order dose-response function, satisfying , , and , monotonic nondecreasing, that is, , with a continuous second derivative, and analytic in the vicinity of 0. Then, we conjecture that:
A-There is a zone in which is convex, that is, , with the implication that a policy of variation of dosage produces beneficial effects:
(The acute outperforms the chronic).
B-There is a zone in which is concave, that is, , with the implication that a policy of stability of dosage produces beneficial effects:
(The chronic outperforms the acute).
4. The Generalized Dose–Response Curve
Let : → [, ], be a continuous function possessing derivatives of all orders, expressed as an N-summed and scaled standard sigmoid function:
where are scaling constants ∈, satisfying:
- (−∞) = , and
- () = , and (equivalently for the first and last of the following conditions)
- ≥ 0 for x∈ (−∞, ), < 0 for x∈ (, ), and ≥ 0 for x∈ (, ∞), with .
By increasing N, we can approximate a continuous function’s density in a metric space, see Cybenko (1989) [34].
The shapes at different calibrations are shown in Figure 11, in which we combined different values of N = 2 and the standard sigmoid , with = 1, = 1, and = 0. As we can see, unlike the common sigmoid, the asymptotic response can be lower than the maximum, as our curves are not monotonically increasing. The sigmoid shows benefits increasing rapidly (the convex phase), then increasing at a slower and slower rate until saturation. Our more general case starts by increasing, but the reponse can actually be negative beyond the saturation phase, though in a convex manner. Harm slows down and becomes “flat” when something is totally broken.
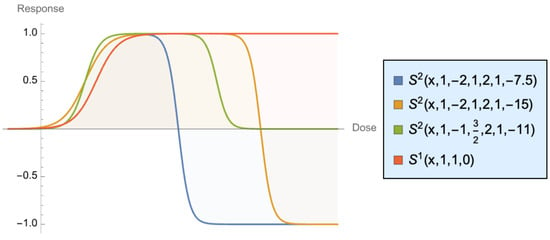
Figure 11.
Generalizing the Dose–Response Curve, The convex part in the increasing section is what we call “antifragile”.
Antifragility and Heterogeneity
Tumors are composed of a heterogeneous collection of subpopulations with varied treatment sensitivity. Given N non-interacting populations, the fragility of the total population is given by the sum of each subpopulation i’s fragility, , weighted by its frequency within the total population, , such that .
where fragility for a single population is given by Equation (1).
The simplest case of a heterogeneous mixture of two populations, sensitive (with associated dose response ) and resistant (), is shown in Figure 12A. In the case that each dose response, , is non-increasing, then the mixed dose response will also be non-increasing, but changes in convexity may occur. As seen in Figure 12A, the mixed dose response has an internal plateau (shown for ). Local convexity (fragility) may switch signs multiple times.
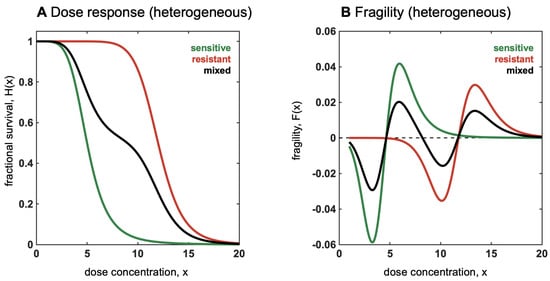
Figure 12.
Relationship between convexity and mixed, heterogeneous populations. (A) Dose response shown for sensitive (green) and resistant (red) cell lines. When mixed, dose response is a weighted average of each (Equation 15; black). (B) Fragility shown for sensitive (green) and resistant (red) cell lines. When mixed, fragility (black) switches from locally convex to locally concave multiple times.
5. Nonlinearities and Medical Iatrogenics
Next, we connect nonlinearity to iatrogenics (that is, harm done by the healer) for medicine in general, broadly defined as all manner of net deficit of benefits minus harm from a given intervention.
The Taleb and Douady (2013) [2] theorems state:
- Convexity for a dose-response function increases fragility (from the expansion of the left tail in response to the increase in the scale of the distribution).
- Detection of a nonlinearity allows the prediction of fragility and helps formulate probabilistic decisions without much knowledge of the probability distribution beyond minimum standard attributes.
- The presence of concavity in the tails of the distribution implies a silent risk.
This approach was used in stress testing by the International Monetary Fund (IMF), where the degree of concavity in the tail was used as an indicator of the severity of tail exposure, see Taleb, Canetti et al. [35]. Such a method can transfer to medicine as the convexity of the dose response can be estimated via titration applied to Equation (1).
5.1. Effect Reversal
Radiation might be beneficial in small doses, with reversal later on. In Neumaier et al. (2012) [36] titled “Evidence for formation of DNA repair centers and dose-response nonlinearity in human cells”:
The standard model currently in use applies a linear scale, extrapolating cancer risk from high doses to low doses of ionizing radiation. However, our discovery of DSB clustering over such large distances casts considerable doubts on the general assumption that risk to ionizing radiation is proportional to dose, and instead provides a mechanism that could more accurately address risk dose dependency of ionizing radiation.
Therefore, low-level radiation may cause hormetic overreaction, producing protective effects. Also see Tubiana et al. (2005) [37]. Bharadwaj and Stafford (2010) present similar general sigmoidal effects in hormonal disruptions by chemicals [38].
5.2. Nonlinearity of NNT and the Consequences
Below are applications of convexity analysis in decision-making in dosage, shown in Figure 13 and Figure 14. In short, it is fallacious to translate a policy derived from acute conditions and apply it to milder ones. Mild conditions are different in treatment from an acute ones. Likewise, high risk is qualitatively different from mild risk.
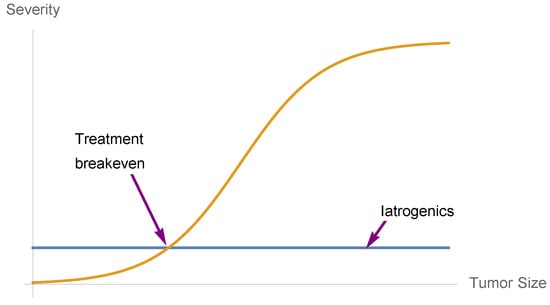
Figure 13.
Drug benefits when convex to numbers needed to treat (NNT) in the left part, with gross iatrogenics invariant to condition (the constant line). We are assuming a standard sigmoidal benefit function.
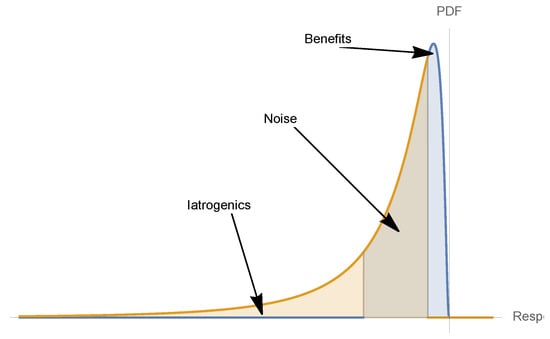
Figure 14.
Unseen risks and mild gains: translation of Figure 13 into a probabilistic representation, showing to the skewness of a decision involving iatrogenics when the condition is mild. This also gives the intuition of the Taleb and Douady [2] translation theorems from concavity for into probabilistic attributes.
There is active literature on “overdiagnosis”, see Kalager et al. (2012) [39] and Morell et al. (2012) [40]. The point is that treating a tumor that does not kill reduces life expectancy; hence the need to balance iatrogenics and risk of cancer. An application of nonlinearity can shed some light on the approach and clarify the public debate [1].
In a similar spirit of avoiding over-treatment, adaptive therapy in metastatic castrate-resistant prostate cancer (clinical trial NCT02415621) has illustrated the feasibility of irregular treatment protocols based on algorithms that react to tumor response. Adaptive treatment protocols maintain a stable population of sensitive cells in order to suppress the emergence of resistance [41,42]. Resistance, in some cases, is similar to the irreconcilable ruin of a financial “blowup” as patients may develop multi-drug resistance to structurally or functionally different drugs. The irreversibility of such clinical outcomes is similar to that of financial ruin, a tail fragility situation from which the agent cannot exit. Adaptive algorithms decrease the cumulative dose administered to a patient, lessening the selection for resistance [43]. While it is not an explicitly stated goal of adaptive therapy, these schedules increase both intra- and inter-patient dosing variance [44].
Last year (2022) saw the publication of calls from within the FDA to revamp the dose-finding protocols to be suitable for targeted therapies [45]. Traditional dose selection protocols invented for use with cytotoxic chemotherapies may not apply to targeted therapies that have exposure–response curves which plateau at low toxicities to the patient. Differences in convexity between chemotherapies and newly developed targeted therapies lead to differing outcomes in diminished returns of dose escalation and differing curvature of dose–response curves.
Author Contributions
N.N.T. and J.W. contributed equally to this work. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not Applicable.
Data Availability Statement
Not Applicable.
Acknowledgments
Yaneer Bar Yam and participants in the International Conference On Complex Systems, Boston, 2018, where early ideas leading to this paper were presented [33] (synthesized in the appendices), as well as Luke Pierik, Sandy Anderson, Maximilian Strobl, and Andriy Marusyk.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A. Antifragility Indirectly Detected in the Various Literature
Table A1 reviews the medical literature on embedded antifragility, defined as indirectly producing evidence of benefits from the “disorder cluster”.

Table A1.
Review of medical research on embedded antifragility.
Table A1.
Review of medical research on embedded antifragility.
| Field | Papers |
|---|---|
| Mithridatization and hormesis | Kaiser (2003) [16], Rattan (2008) [46], Calabrese and Baldwin (2002, 2003a, 2003b) [17,47,48], Aruguman et al. (2006) [49]. |
| Caloric restriction and hormesis | Martin, Mattson et al. (2006) [12] |
| Cancer treatment and fasting | Longo et al. (2010) [50], Safdie et al. (2009) [51], Raffaghelo et al. (2010), [52], Lee et al. (2012) [53] |
| Aging and intermittence | Fontana et al. [54] |
| For brain effects | Anson, Guo, et al. (2003) [55], Halagappa, Guo, et al. (2007) [56], Stranahan and Mattson (2012) [57]. The long-held belief that the brain needed glucose, not ketones, and that the brain does not go through autophagy, has been progressively replaced. |
| Yeast and longevity under restriction | Fabrizio et al. (2001) [58]; SIRT1, Longo et al. (2006) [59], Michan et al. (2010) [60] |
| Diabetes, remission or reversal | Taylor (2008) [61], Lim et al. (2011) [62], Boucher et al. (2004) [63]; diabetes management by diet alone, early insights in Wilson et al. (1980) [64]. Couzin (2008) [65] gives insight that blood sugar stabilization does not have the effect anticipated (there need to be stressors). The ACCORD study (Action to Control Cardiovascular Risk in Diabetes) found no benefits from lowering blood glucose levels. Synthesis, Skyler et al. (2009) [66], old methods, Westman and Vernon (2008) [67]. Bariatric (or other) surgery as alternative to intermittent fasting: Pories (1995) [68], Guidone et al. (2006) [69], Rubino et al. 2006 [70] |
| Ramadan and effect of fasting | Trabelsi et al. (2012) [71], Akanji et al. (2012). Note that the Ramadan time window is short (12 to 17 h) and possibly fraught with overeating so conclusions need to take into account energy balance and that the considered effect is at the low-frequency part of the timescale. |
| Caloric restriction | Harrison (1984), Wiendruch (1996), Pischon (2008) |
| Autophagy for cancer | Kondo et al. (2005) [72] |
| Autophagy (general) | Danchin et al. (2011) [73], He et al. (2012) [74] |
| Fractional dosage | Wu et al. (2016) [75] |
| Jensen’s inequality in exercise | Many such as Schnohr and Marott (2011) [76], intermittent extremes vs. moderate physical activity. |
| Cluster of ailments | Yaffe and Blackwell (2004) [77], Alzheimer and hyperinsulenemia, Razay and Wilcock (1994) [78]; Luchsinger, Tang, et al. (2002) [79], Luchsinger Tang et al. (2004) [80] Janson, Laedtke, et al. (2004) [81]. |
| Benefits ofsome type ofstress (and convexity of the effect) | For the different results from the two types of stressors, short and chronic, Dhabar (2009) “A hassle a day may keep the pathogens away: the fight-or-flight stress response and the augmentation of immune function” [82]. For the benefits of stress on boosting immunity and cancer resistance (squamous cell carcinoma), Dhabhar et al. (2010) [83], Dhabhar et al. (2012) [84], Ansbacher et al. (2013) [85] |
| Iatrogenics of hygiene and systematic elimination of germs | Rook (2011) [86], Rook (2012) [87] (auto-immune diseases from absence of stressors), Mégraud and Lamouliatte (1992) [88] for Helyobacter Pilori and incidence of cancer. |
Appendix B. Simple Convexity and Its Effects
To eliminate ambiguity, let us define convexity. Let be the response function, and be a twice-differentiable function. If, over a range x , over a set time period , 0, or more practically (by relaxing the assumptions of differentiability), , with and , then there are benefits or harm from the unevenness of distribution, depending on whether f is defined as positive or favorable or modeled as a harm function (in which case one needs to reverse the sign for the interpretation).
We can generalize to comparing linear combinations: , , ; thus, we have situations where, for and , . This last property describes a “stressor” as having higher intensity than zero: there may be no harm from , yet there will be some at higher levels of x.
Now, if X is a random variable with support in and f is convex over the interval as per above, then
which is commonly known as Jensen’s inequality, see Jensen(1906) [89], Figure 4. Further (without loss of generality), if its continuous distribution with density and support in belongs to the location scale family distribution, with and , then, with , the indexing representing the expectation under a probability distribution indexed by the scale , we have:
The last property implies that the convexity effect increases the expectation operator. We can verify that since is an increasing function of . A more simple approach (inspired by mathematical finance heuristics) is to consider , where and are the mean expected deviations or, alternatively, the results of a simplified two-state system, each with probability :
This is, of course, a simplification here, since dose response is rarely monotone in its nonlinearity, as we will see in later sections. However, we can at least make claims in a certain interval and it can produce useful heuristics.
What are we measuring? Clearly, the dose (represented on the x line) is hardly ambiguous: any measurable quantity can suffice, such as systolic blood pressure, ejection fraction, caloric deficit, pounds per square inch, temperature, etc. The response, harm or benefits, , on the other hand, need to be equally precise, with nothing vague, such as hazard ratios, some quantifiable index of health, median life expectancy, and similar quantities. If one cannot express the response quantitatively, then such an analysis cannot apply.
Appendix C. Relaxing the Assumption of Fixed Treatment Schedules
We can relax the assumption of fixed treatment schedules (e.g., Figure 4). Given some input probability density function describing the distribution of dose, , the probability density can be determined for the Hill function analytically. Let X (dose concentration) and be random variables. The probability density function transformation, , is given by:
References
- Taleb, N.N. Antifragile: Things That Gain from Disorder; Random House and Penguin: New York, NY, USA, 2012. [Google Scholar]
- Taleb, N.N.; Douady, R. Mathematical definition, mapping, and detection of (anti) fragility. Quant. Financ. 2013, 13, 1677–1689. [Google Scholar] [CrossRef]
- Taleb, N.N. Dynamic Hedging: Managing Vanilla and Exotic Options; John Wiley and Sons (Wiley Series in Financial Engineering): Hoboken, NJ, USA, 1997. [Google Scholar]
- Geman, D.; Geman, H.; Taleb, N. Tail Risk Constraints and Maximum Entropy. Entropy 2015, 17, 3724. [Google Scholar] [CrossRef]
- Taleb, N.N. The Statistical Consequences of Fat Tails; STEM Academic Press: New York, NY, USA, 2020. [Google Scholar]
- Brewster, J.F.; Graham, M.R.; Mutch, W.A.C. Convexity, jensen’s inequality and benefits of noisy mechanical ventilation. J. R. Soc. Interface 2005, 2, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Amato, M.B.P.; Barbas, C.S.V.; Medeiros, D.M.; Magaldi, R.B.; Schettino, G.P.; Lorenzi-Filho, G.; Kairalla, R.A.; Deheinzelin, D.; Munoz, C.; Oliveira, R.; et al. Effect of a Protective-Ventilation Strategy on Mortality Acute Respiratory Distress Syndrome. N. Engl. J. Med. 1998, 338, 347–354. [Google Scholar] [CrossRef]
- Funk, D.J.; Graham, M.R.; Girling, L.G.; Thliveris, J.A.; McManus, B.M.; Walker, E.K.; Rector, E.S.; Hillier, C.; Scott, J.E.; Mutch, W.A.C. A comparison of biologically variable ventilation to recruitment manoeuvres in a porcine model of acute lung injury. Respir. Res. 2004, 5, 22. [Google Scholar] [CrossRef] [PubMed]
- Arold, S.P.; Suki, B.; Alencar, A.M.; Lutchen, K.R.; Ingenito, E.P. Variable ventilation induces endogenous surfactant release in normal guinea pigs. Am. J. Physiol.-Lung Cell. Mol. 2003, 285, L370–L375. [Google Scholar] [CrossRef]
- Graham, M.R.; Haberman, C.J.; Brewster, J.F.; Girling, L.G.; McManus, B.M.; Mutch, W.A.C. Mathematical modelling to centre low tidal volumes following acute lung injury: A study with biologically variable ventilation. Respir. Res. 2005, 6, 64. [Google Scholar] [CrossRef][Green Version]
- Den Hartigh, R.; Hill, Y. Conceptualizing and measuring psychological resilience: What can we learn from physics? New Ideas Psychol. 2022, 66, 100934. [Google Scholar] [CrossRef]
- Martin, B.; Mattson, M.P.; Maudsley, S. Caloric restriction and intermittent fasting: Two potential diets for successful brain aging. Ageing Res. Rev. 2006, 5, 332–353. [Google Scholar] [CrossRef]
- Fontana, L.; Weiss, E.; Villareal, D.; Klein, S.; Holloszy, J. Long-term effects of calorie or protein restriction on serum IGF-1 and IGFBP-3 concentration in humans. Aging Cell. 2008, 7, 681–687. [Google Scholar] [CrossRef]
- Lee, C.; Longo, V. Fasting vs dietary restriction in cellular protection and cancer treatment: From model organisms to patients. Oncogene 2011, 30, 3305–3316. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.; Schultz, J.; Matin, A. Starvation-induced cross protection against heat or H2O2 challenge in Escherichia coli. J. Ofbacteriol. 1988, 170, 3910–3914. [Google Scholar] [CrossRef]
- Kaiser, J. Sipping from a poisoned chalice. Science 2003, 302, 376. [Google Scholar] [CrossRef]
- Calabrese, E.; Baldwin, L. Hormesis: The dose-response revolution. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 175–197. [Google Scholar] [CrossRef]
- Staňková, K.; Brown, J.; Dalton, W.; Gatenby, R. Optimizing cancer treatment using game theory: A review. JAMA Oncol. 2019, 5, 96–103. [Google Scholar] [CrossRef] [PubMed]
- West, J.; Desai, B.; Strobl, M.; Pierik, L.; Ver Velde, R.; Armagost, C.; Miles, R.; Robertson-Tessi, M.; Marusyk, A.; Anderson, A. Antifragile therapy. BioRxiv 2021. [Google Scholar] [CrossRef]
- Piccart, M.; Biganzoli, L.; Leo, A.D. The impact of chemotherapy dose density and dose intensity on breast cancer outcome: What have we learned? Eur. J. Cancer 2000, 36, 4–10. [Google Scholar] [CrossRef]
- Citron, M. Dose-dense chemotherapy: Principles, clinical results and future perspectives. Breast Care 2008, 3, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Skipper, H.; Schabel, F.; Wilcox, W. Experimental evaluation of potential anticancer agents XIII, on the criteria and kinetics associated with“ curability” of experimental leukemia. Cancer Chemother. Rep. 1964, 35, 3–111. [Google Scholar]
- Kerbel, R.; Kamen, B. The anti-angiogenic basis of metronomic chemotherapy. Nat. Rev. Cancer 2004, 4, 423–436. [Google Scholar] [CrossRef]
- Amin, D.; Sergina, N.; Ahuja, D.; McMahon, M.; Blair, J.; Wang, D.; Hann, B.; Koch, K.; Shokat, K.; Moasser, M. Resiliency and vulnerability in the HER2-HER3 tumorigenic driver. Sci. Transl. Med. 2010, 2, 16ra7. [Google Scholar] [CrossRef]
- Griffiths, J.; Chen, J.; Cosgrove, P.; O’Dea, A.; Sharma, P.; Ma, C.; Trivedi, M.; Kalinsky, K.; Wisinski, K.; O’Regan, R.; et al. Serial single-cell genomics reveals convergent subclonal evolution of resistance as patients with early-stage breast cancer progress on endocrine plus CDK4/6 therapy. Nat. Cancer 2021, 2, 658–671. [Google Scholar] [CrossRef] [PubMed]
- Chmielecki, J.; Foo, J.; Oxnard, G.; Hutchinson, K.; Ohashi, K.; Somwar, R.; Wang, L.; Amato, K.; Arcila, M.; Sos, M.; et al. Optimization of dosing for EGFR-mutant non–small cell lung cancer with evolutionary cancer modeling. Sci. Transl. Med. 2011, 3, 90ra59. [Google Scholar] [CrossRef] [PubMed]
- Schöttle, J.; Chatterjee, S.; Volz, C.; Siobal, M.; Florin, A.; Rokitta, D.; Hinze, Y.; Dietlein, F.; Plenker, D.; König, K.; et al. Intermittent high-dose treatment with erlotinib enhances therapeutic efficacy in EGFR-mutant lung cancer. Oncotarget 2015, 6, 38458. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, Q.; Li, R.; Chen, X.; Lee, S.; Pan, J.; Xiong, D.; Hu, J.; Miller, M.; Szabo, E.; Lubet, R.; et al. Effect of weekly or daily dosing regimen of Gefitinib in mouse models of lung cancer. Oncotarget 2017, 8, 72447. [Google Scholar] [CrossRef]
- Grommes, C.; Oxnard, G.; Kris, M.; Miller, V.; Pao, W.; Holodny, A.; Clarke, J.; Lassman, A. Pulsatile high-dose weekly erlotinib for CNS metastases from EGFR mutant non-small cell lung cancer. Neuro-oncology 2011, 13, 1364–1369. [Google Scholar] [CrossRef]
- Hafner, M. Growth Rate Inhib. Metrics Correct Confounders Meas. Sensit. Cancer Drugs. Nat. Methods 2016, 13, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.; Wooten, D.; Paudel, B.; Bauer, J.; Hardeman, K.; Westover, D.; Lovly, C.; Harris, L.; Tyson, D.; Quaranta, V. Quantifying drug combination synergy along potency and efficacy axes. Cell Syst. 2019, 8, 97–108. [Google Scholar] [CrossRef]
- Cunningham, J. A call for integrated metastatic management. Nat. Ecol. Evol. 2019, 3, 996–998. [Google Scholar] [CrossRef]
- Taleb, N.N. (Anti)Fragility and Convex Responses in Medicine. In Proceedings of the International Conference On Complex Systems, Proceedings, Cambridge, MA, USA, 22–27 July 2018; pp. 299–325. [Google Scholar]
- Cybenko, G. Approximation by superpositions of a sigmoidal function. Math. Control. Signals Syst. 1989, 2, 303–314. [Google Scholar] [CrossRef]
- Taleb, N.N.; Canetti, E.; Kinda, T.; Loukoianova, E.; Schmieder, C. A new heuristic measure of fragility and tail risks: Application to stress testing. International Monetary Fund. 2018. Available online: https://www.imf.org/external/pubs/ft/wp/2012/wp12216.pdf (accessed on 8 February 2023).
- Neumaier, T.; Swenson, J.; Pham, C.; Polyzos, A.; Lo, A.T.; Yang, P.; Dyball, J.; Asaithamby, A.; Chen, D.J.; Bissell, M.J.; et al. Evidence for formation of dna repair centers and dose-response nonlinearity in human cells. Proc. Natl. Acad. Sci. USA 2012, 109, 443–448. [Google Scholar] [CrossRef]
- Tubiana, M.; Aurengo, A.; Averbeck, D.; Masse, R. Recent reports on the effect of low doses of ionizing radiation and its dose–effect relationship. Radiat. Environ. Biophys. 2006, 44, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, A.; Stafford, K.C., III. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. J. Med. Entomol 2010, 47, 862–867. [Google Scholar] [CrossRef] [PubMed]
- Kalager, M.; Adami, H.-O.; Bretthauer, M.; Tamimi, R.M. Overdiagnosis of invasive breast cancer due to mammography screening: Results from the norwegian screening program. Ann. Intern. Med. 2012, 156, 491–499. [Google Scholar] [CrossRef]
- Morrell, S.; Barratt, A.; Irwig, L.; Howard, K.; Biesheuvel, C.; Armstrong, B. Estimates of overdiagnosis of invasive breast cancer associated with screening mammography. Cancer Causes Control 2010, 21, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Gatenby, R.; Silva, A.; Gillies, R.; Frieden, B. Adaptive therapy. Cancer Res. 2009, 69, 4894–4903. [Google Scholar] [CrossRef]
- West, J.; You, L.; Zhang, J.; Gatenby, R.; Brown, J.; Newton, P.; Anderson, A. Towards Multidrug Adaptive Therapy. Cancer Res. 2020, 80, 1578–1589. [Google Scholar] [CrossRef]
- Cunningham, J.; Brown, J.; Gatenby, R.; Staňková, K. Optimal control to develop therapeutic strategies for metastatic castrate resistant prostate cancer. J. Theor. Biol. 2018, 459, 67–78. [Google Scholar] [CrossRef]
- Zhang, J.; Cunningham, J.; Brown, J.; Gatenby, R. Integrating evolutionary dynamics into treatment of metastatic castrate-resistant prostate cancer. Nat. Commun. 2017, 8, 1816. [Google Scholar] [CrossRef]
- Tracey, A. FDA’s Brian Booth: “We need to reconsider our approach to dose selection”. Cancer Lett. 2022, 48, 12–16. [Google Scholar]
- Rattan, S.I. Hormesis in aging. Ageing Res. Rev. 2008, 7, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Baldwin, L.A. Defining hormesis. Hum. Exp. Toxicol. 2002, 21, 91–97. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Baldwin, L.A. The hormetic dose-response model is more common than the threshold model in toxicology. Toxicol. Sci. 2003, 71, 246–250. [Google Scholar] [CrossRef]
- Arumugam, T.V.; Gleichmann, M.; Tang, S.-C.; Mattson, M.P. Hormesis/preconditioning mechanisms, the nervous system and aging. Ageing Res. Rev. 2006, 5, 165–178. [Google Scholar] [CrossRef]
- Longo, V.D.; Fontana, L. Calorie restriction and cancer prevention: Metabolic and molecular mechanisms. Trends Pharmacol. 2010, 31, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Safdie, F.M.; Dorff, T.; Quinn, D.; Fontana, L.; Wei, M.; Lee, C.; Cohen, P.; Longo, V.D. Fasting and cancer treatment in humans: A case series report. Aging (Albany NY) 2009, 1, 988–1007. [Google Scholar] [CrossRef]
- Raffaghello, L.; Safdie, F.; Bianchi, G.; Dorff, T.; Fontana, L.; Longo, V.D. Fasting and differential chemotherapy protection in patients. Cell Cycle 2010, 9, 4474–4476. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Raffaghello, L.; Brandhorst, S.; Safdie, F.M.; Bianchi, G.; Martin-Montalvo, A.; Pistoia, V.; Wei, M.; Hwang, S.; Merlino, A.; et al. Fasting Cycles Retard Growth Tumors Sensitize A Range Cancer Celltypes Chemotherapy. Sci. Transl. Med. 2012, 4, 124ra27. [Google Scholar] [CrossRef]
- Fontana, L.; Kennedy, B.K.; Longo, V.D.; Seals, D.; Melov, S. Medical research: Treat ageing. Nature 2014, 511, 405–407. [Google Scholar] [CrossRef]
- Anson, R.M.; Guo, Z.; de Cabo, R.; Iyun, T.; Rios, M.; Hagepanos, A.; Ingram, D.K.; Lane, M.A.; Mattson, M.P. Intermittent fasting dissociates beneficial effects of dietary restriction on glucose metabolism and neuronal resistance to injury from calorie intake. Proc. Natl. Acad. Sci. USA 2003, 100, 6216–6220. [Google Scholar] [CrossRef]
- Halagappa, V.K.M.; Guo, Z.; Pearson, M.; Matsuoka, Y.; Cutler, R.G.; LaFerla, F.M.; Mattson, M.P. Intermittent fasting and caloric restriction ameliorate age-related behavioral deficits in the triple-transgenic mouse model of alzheimer’s disease. Neurobiol. Dis. 2007, 26, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Stranahan, A.M.; Mattson, M.P. Recruiting adaptive cellular stress responses for successful brain ageing. Nat. Rev. Neurosci. 2012, 13, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Fabrizio, P.; Pozza, F.; Pletcher, S.D.; Gendron, C.M.; Longo, V.D. Regulation of longevity and stress resistance by sch9 in yeast. Science 2001, 292, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.D.; Kennedy, B.K. Sirtuins in aging and age-related disease. Cell 2006, 126, 257–268. [Google Scholar] [CrossRef]
- Michán, S.; Li, Y.; Chou, M.M.-H.; Parrella, E.; Ge, H.; Long, J.M.; Allard, J.S.; Lewis, K.; Miller, M.; Xu, W.; et al. Sirt1 is essential for normal cognitive function and synaptic plasticity. J. Neurosci. 2010, 30, 9695–9707. [Google Scholar] [CrossRef]
- Taylor, R. Pathogenesis of type 2 diabetes: Tracing the reverse route from cure to cause. Diabetologia 2008, 51, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.L.; Hollingsworth, K.; Aribisala, B.S.; Chen, M.; Mathers, J.; Taylor, R. Reversal of type 2 diabetes: Normalisation of beta cell function in association with decreased pancreas and liver triacylglycerol. Diabetologia 2011, 54, 2506–2514. [Google Scholar] [CrossRef]
- Boucher, A.; Lu, D.; Burgess, S.C.; Telemaque-Potts, S.; Jensen, M.V.; Mulder, H.; Wang, M.-Y.; Unger, R.H.; Sherry, A.D.; Newgard, C.B. Biochemical mechanism of lipid-induced impairment of glucose-stimulated insulin secretion and reversal with a malate analogue. J. Biol. 2004, 279, 27263–27271. [Google Scholar] [CrossRef]
- Wilson, E.A.; Hadden, D.; Merrett, J.; Montgomery, D.; Weaver, J. Dietary management of maturity-onset diabetes. Br. Med. J. 1980, 280, 1367–1369. [Google Scholar] [CrossRef]
- Couzin, J. Deaths in diabetes trial challenge a long-held theory. Science 2008, 319, 884–885. [Google Scholar] [CrossRef]
- Skyler, J.S.; Bergenstal, R.; Bonow, R.O.; Buse, J.; Deedwania, P.; Gale, E.A.; Howard, B.V.; Kirkman, M.S.; Kosiborod, M.; Reaven, P.; et al. Intensive glycemic control and the prevention of cardiovascular events: Implications of the accord, advance, and va diabetes trials: A position statement of the american diabetes association and a scientific statement of the american college of cardiology foundation and the american heart association. J. Am. Coll. Cardiol. 2009, 53, 298–304. [Google Scholar] [PubMed]
- Westman, E.C.; Vernon, M.C. Has carbohydrate-restriction been forgotten as a treatment for diabetes mellitus? A perspective on the accord study design. Nutr. Metab. 2008, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Pories, W.J.; Swanson, M.S.; MacDonald, K.G.; Long, S.B.; Morris, P.G.; Brown, B.M.; Barakat, H.A. Who would have thought it? An operation proves to be the most effective therapy for adult-onset diabetes mellituss. Ann. Surg. 1995, 222, 339. [Google Scholar] [CrossRef]
- Guidone, C.; Manco, M.; Valera-Mora, E.; Iaconelli, A.; Gniuli, D.; Mari, A.; Nanni, G.; Castagneto, M.; Calvani, M.; Mingrone, G. Mechanisms of recovery from type 2 diabetes after malabsorptive bariatric surgery. Diabetes 2006, 55, 2025–2031. [Google Scholar] [CrossRef]
- Rubino, F.; Forgione, A.; Cummings, D.E.; Vix, M.; Gnuli, D.; Mingrone, G.; Castagneto, M.; Marescaux, J. The mechanism of diabetes control after gastrointestinal bypass surgery reveals a role of the proximal small intestine in the pathophysiology of type 2 diabetes. Ann. Surg. 2006, 244, 741–749. [Google Scholar] [CrossRef]
- Trabelsi, K.; Stannard, S.R.; Maughan, R.J.; Jamoussi, K.; Zeghal, K.M.; Hakim, A. Effect of resistance training during ramadan on body composition, and markers of renal function, metabolism, inflammation and immunity in tunisian recreational bodybuilders. Intern. J. Sport Nutr. Exer. Metabo. 2012, 22, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y.; Kanzawa, T.; Sawaya, R.; Kondo, S. The role of autophagy in cancer development and response to therapy. Nat. Rev. Cancer 2005, 5, 726–734. [Google Scholar] [CrossRef]
- Danchin, A.; Binder, P.M.; Noria, S. Antifragility and tinkering in biology (and in business) flexibility provides an efficient epigenetic way to manage risk. Genes 2011, 2, 998–1016. [Google Scholar] [CrossRef]
- He, C.; Bassik, M.C.; Moresi, V.; Sun, K.; Wei, Y.; Zou, Z.; An, Z.; Loh, J.; Fisher, J.; Sun, Q.; et al. Exercise-induced bcl2-regulated autophagy is required for muscle glucose homeostasis. Nature 2012, 481, 511–515. [Google Scholar] [CrossRef]
- Wu, J.T.; Peak, C.M.; Leung, G.M.; Lipsitch, M. Fractional dosing of yellow fever vaccine to extend supply: A modeling study. Lancet 2016, 388, 2904–2911. [Google Scholar] [CrossRef]
- Schnohr, P.; Marott, J.L.; Jensen, J.S.; Jensen, G.B. Intensity versus duration of cycling, impact on all-cause and coronary heart disease mortality: The copenhagen city heart study. Eur. J. Cardiovascular Prev. Rehabil. 2011, 19, 73–80. [Google Scholar] [CrossRef]
- Yaffe, K.; Blackwell, T.; Kanaya, A.; Davidowitz, N.; Barrett-Connor, E.; Krueger, K. Diabetes, impaired fasting glucose, and development of cognitive impairment in older women. Neurology 2004, 63, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Razay, G.; Wilcock, G.K. Hyperinsulinaemia and alzheimer’s disease. Age Ageing 1994, 23, 396–399. [Google Scholar] [CrossRef]
- Luchsinger, J.A.; Tang, M.-X.; Shea, S.; Mayeux, R. Caloric intake and the risk of alzheimer disease. Arch. Neurol. 2002, 59, 1258–1263. [Google Scholar] [CrossRef] [PubMed]
- Luchsinger, J.A.; Tang, M.-X.; Shea, S.; Mayeux, R. Hyperinsulinemia and risk of alzheimer disease. Neurology 2004, 63, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Janson, J.; Laedtke, T.; Parisi, J.E.; O’Brien, P.; Petersen, R.C.; Butler, P.C. Increased risk of type 2 diabetes in alzheimer disease. Diabetes 2004, 53, 474–481. [Google Scholar] [CrossRef]
- Dhabhar, F.S. A hassle a day may keep the pathogens away: The fight-or-flight stress response and the augmentation of immune function. Integr. Comp. Biol. 2009, 49, 215–236. [Google Scholar] [CrossRef] [PubMed]
- Dhabhar, F.S.; Saul, A.N.; Daugherty, C.; Holmes, T.H.; Bouley, D.M.; Oberyszyn, T.M. Short-term stress enhances cellular immunity and increases early resistance to squamous cell carcinoma. Brain, Behav. Andimmunity 2010, 24, 127–137. [Google Scholar] [CrossRef]
- Dhabhar, F.S.; Saul, A.N.; Holmes, T.H.; Daugherty, C.; Neri, E.; Tillie, J.M.; Kusewitt, D.; Oberyszyn, T.M. High-anxious individuals show increased chronic stress burden, decreased protective immunity, and increased cancer progression in a mouse model of squamous cell carcinoma. PLoS ONE 2012, 7, e33069. [Google Scholar] [CrossRef]
- Aschbacher, K.; O’Donovan, A.; Wolkowitz, O.M.; Dhabhar, F.S.; Su, Y.; Epel, E. Good stress, bad stress and oxidative stress: Insights from anticipatory cortisol reactivity. Psychoneuroendocrinology 2013, 38, 1698–1708. [Google Scholar] [CrossRef]
- Rook, G.A. Hygiene and other early childhood influences on the subsequent function of the immune system. Dig. Dis. 2011, 29, 144–153. [Google Scholar] [CrossRef]
- Rook, G.A. Hygiene hypothesis and autoimmune diseases. Clin. Rev. Allergy Immunol. 2012, 42, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Mégraud, F.; Lamouliatte, H. Helicobacter pylori and duodenal ulcer. Dig. Dis. Sci. 1992, 37, 769–772. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.L.W.V. Sur les fonctions convexes et les inégalités entre les valeurs moyennes. Acta Math. 1906, 30, 175–193. [Google Scholar] [CrossRef]














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).