
Biophotons and Emergence of Quantum Coherence—A Diffusion Entropy Analysis

 , ,
, ,  , , , ,
, , , , 
Abstract
1. Introduction
1.1. Introduction to Biophotons
1.2. Introduction to Complexity
2. Material and Methods
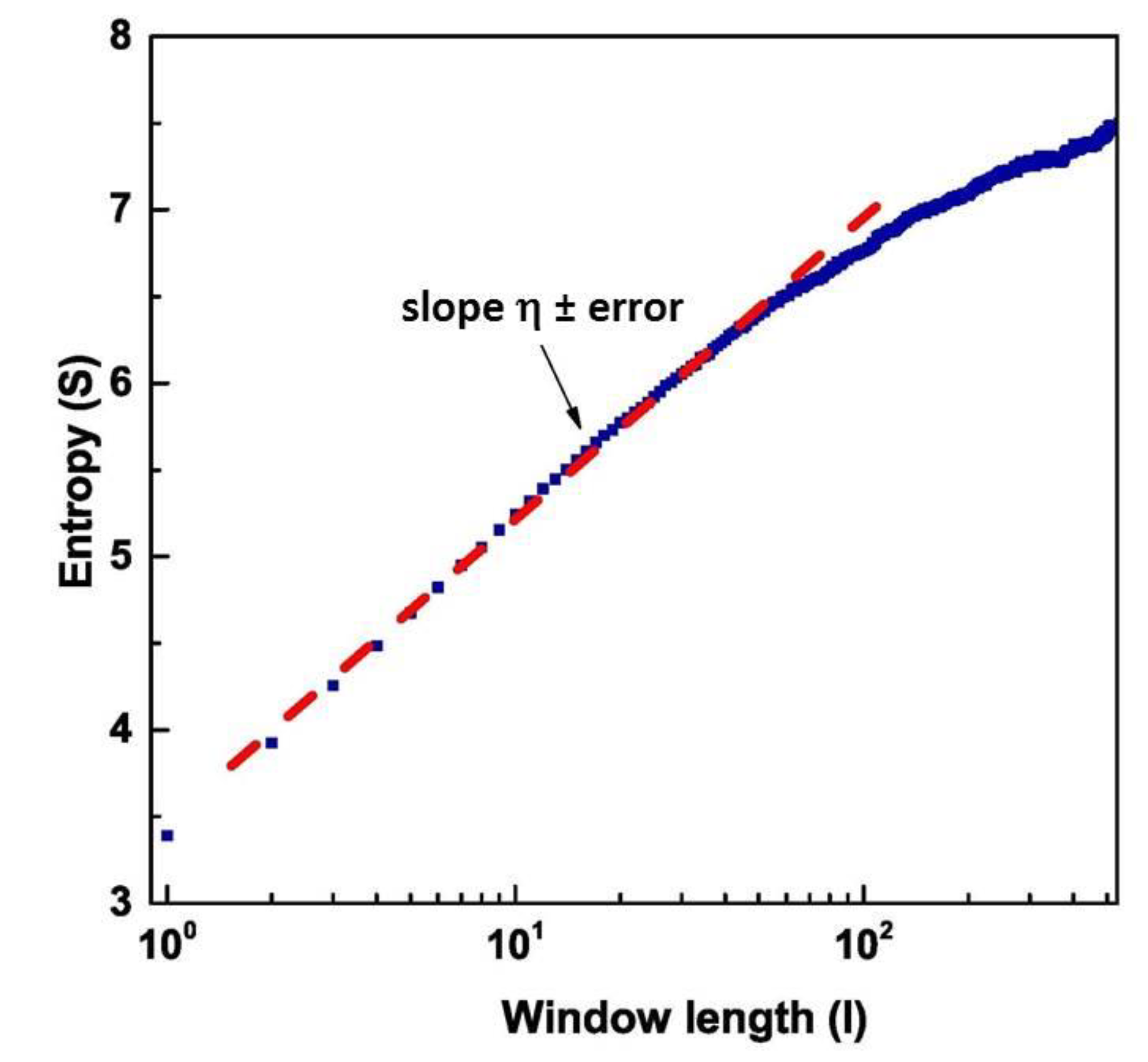
3. Theoretical Foundation
Definition of μ
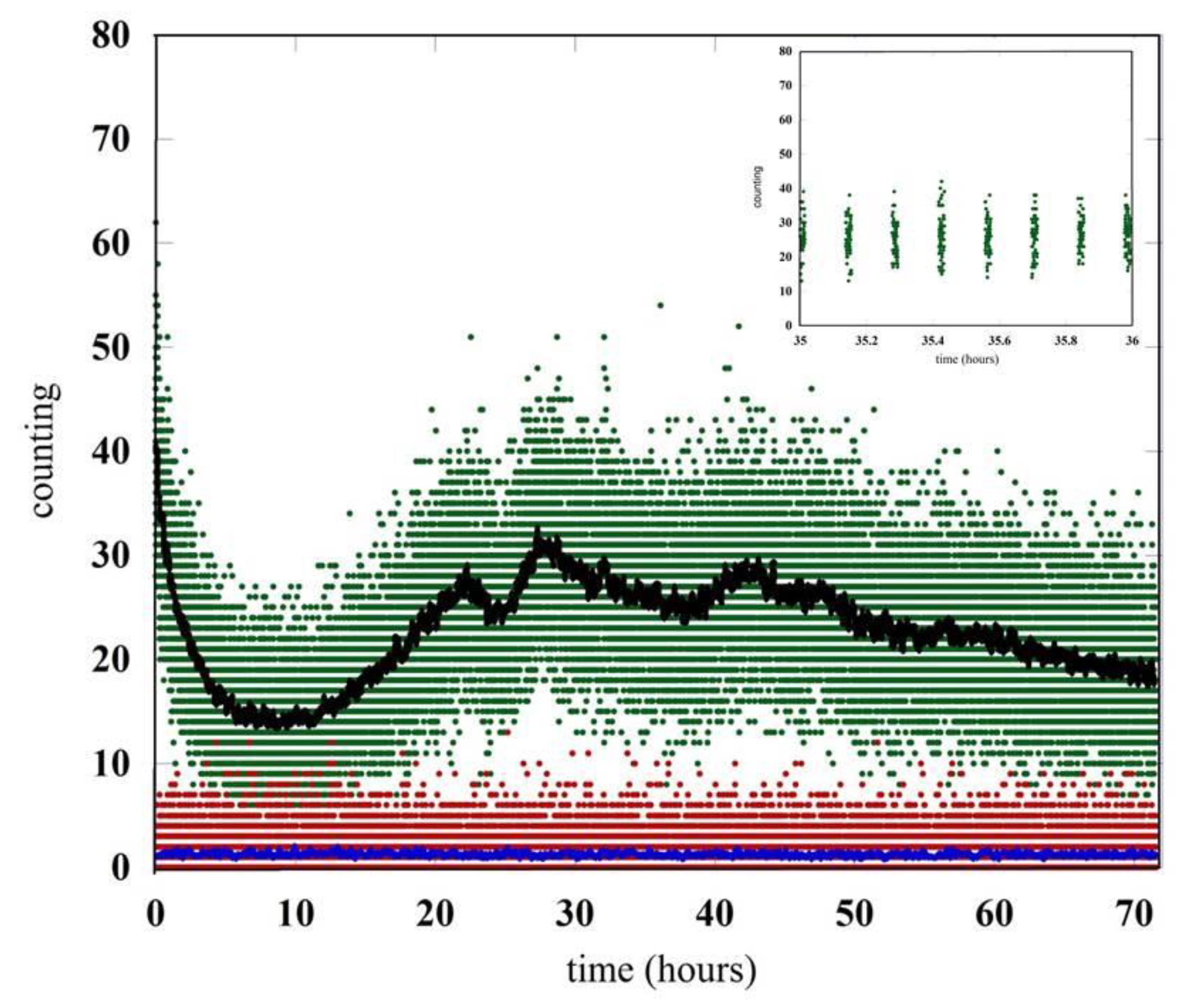
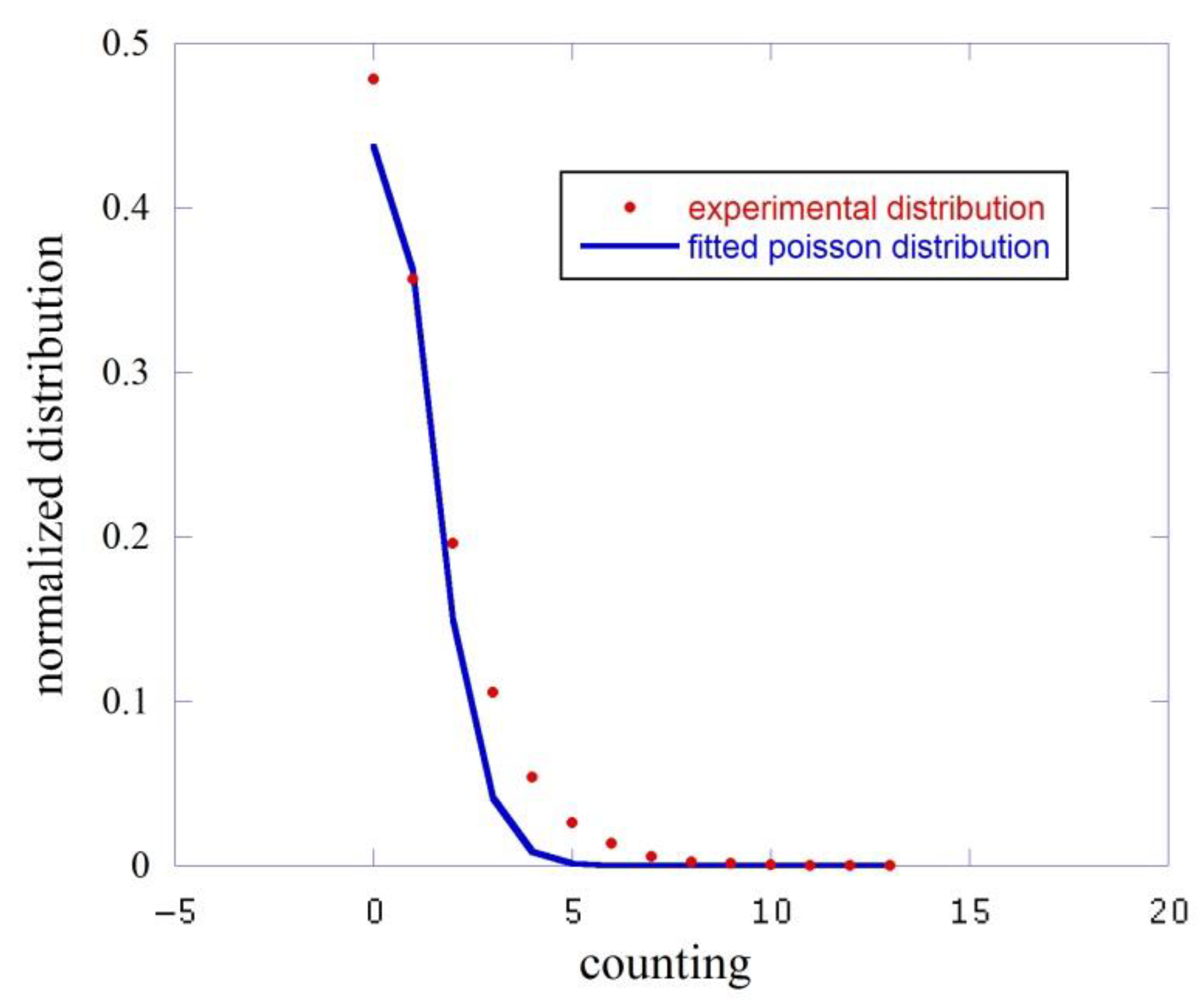
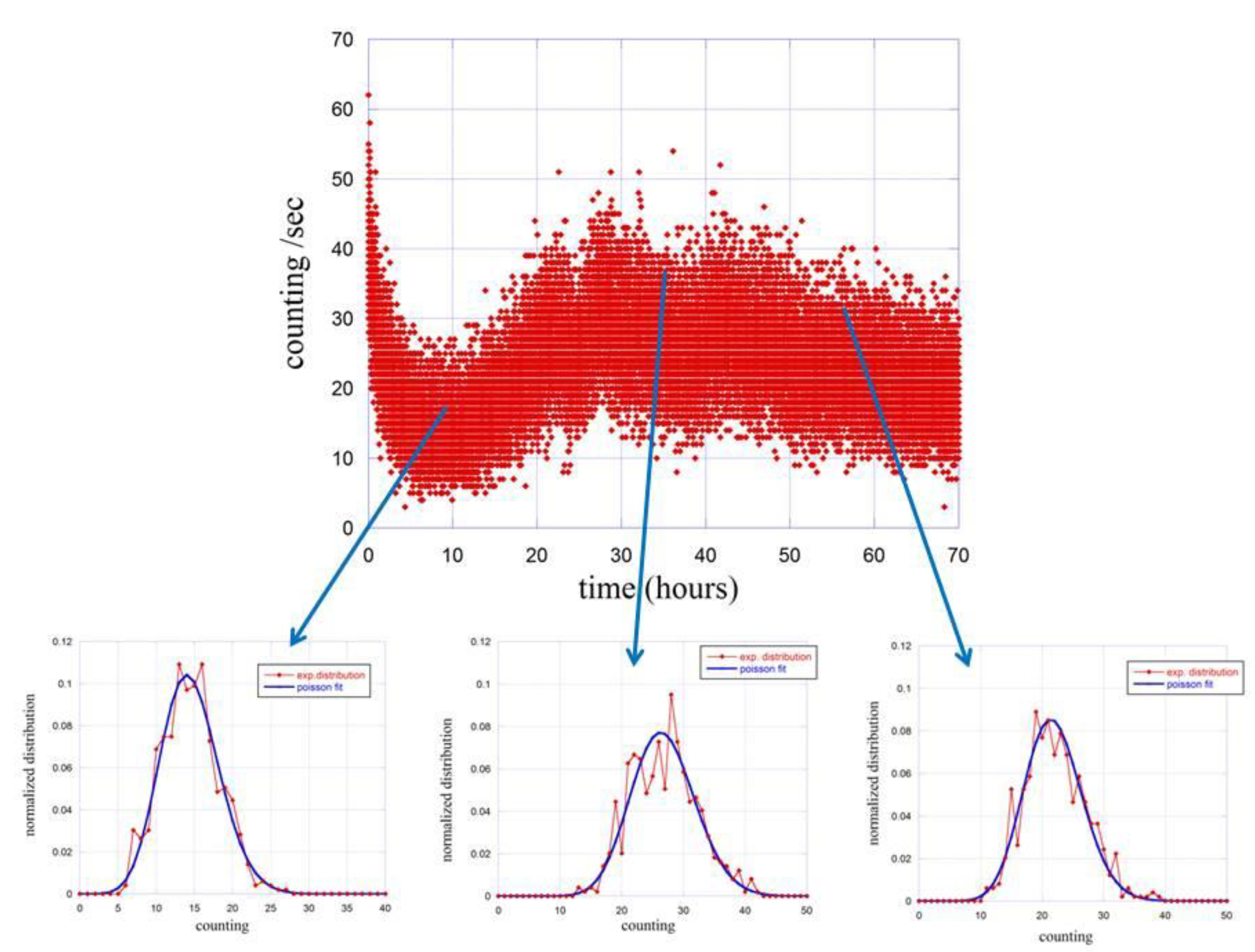
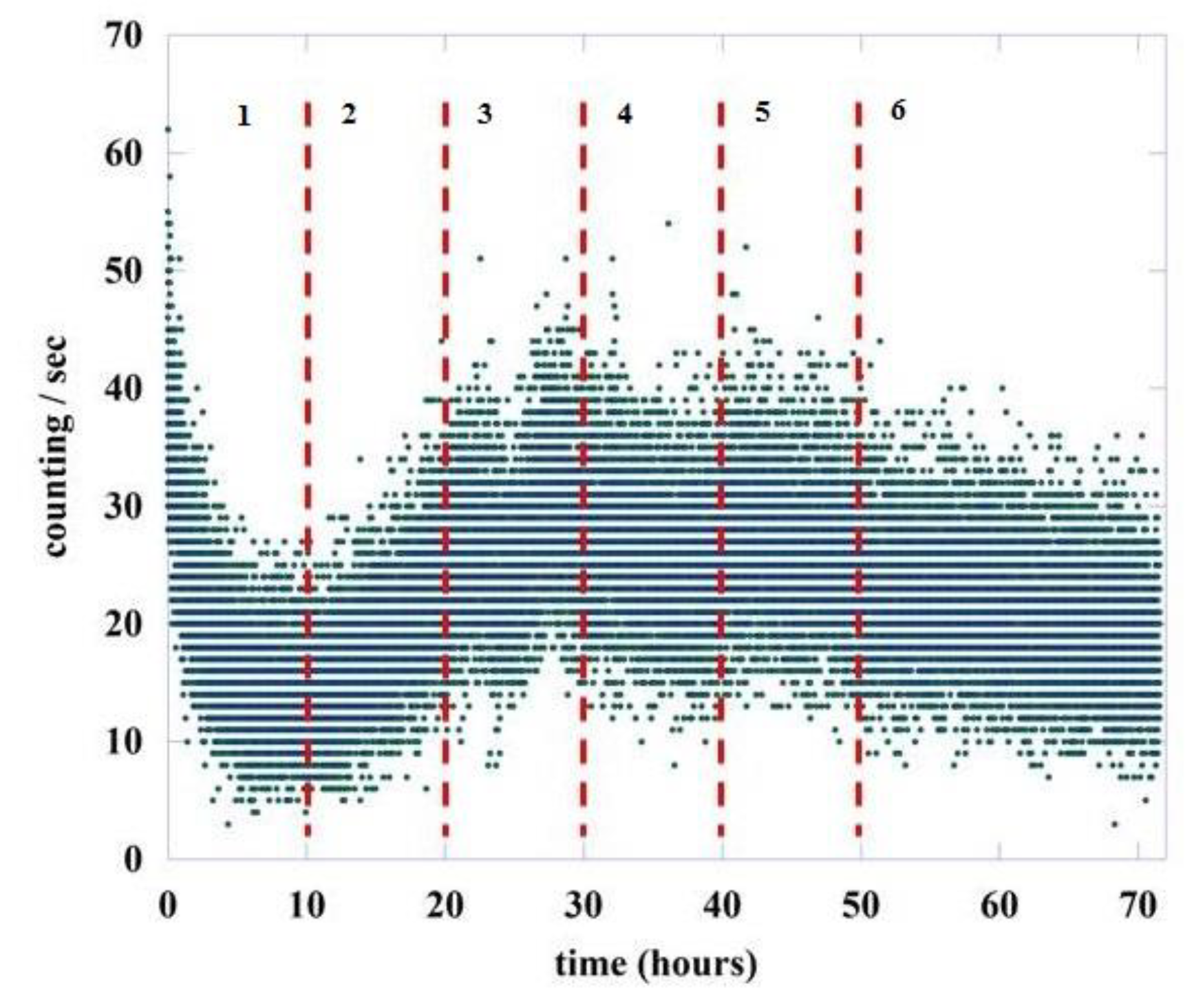
4. Results
5. Discussion and Conclusions
5.1. Results of This Paper
5.2. Suggestions for Future Research Work
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gurwitsch, A.G. Die Natur des spezifischen Erregers der Zellteilung. Arch. Entw. Mech. Org. 1923, 100, 11–40. [Google Scholar] [CrossRef]
- Reiter, T.; Gabor, D. Ultraviolette Strahlung und Zellteilung. Wiss. Verffentlichungen Aus Dem Siemens-Konzern 1928, 4, 184. [Google Scholar]
- Colli, L.; Facchini, U. Light Emission by Germinating Plants. Il Nuovo Cim. 1954, 12, 150–153. [Google Scholar] [CrossRef]
- Colli, L.; Facchini, U.; Guidotti, G.; Dugnani Lonati, R.; Orsenigo, M.; Sommariva, O. Further Measurements on the Bioluminescence of the Seedlings. Experientia 1955, 11, 479–481. [Google Scholar] [CrossRef]
- Popp, F.A.; Gu, Q.; Li, K.H. Biophoton Emission: Experimental Background and Theoretical Approaches. Mod. Phys. Lett. B 1994, 8, 1269–1296. [Google Scholar] [CrossRef]
- Van Wijk, R. Light in Shaping Life: Biophotons in Biology and Medicine; Boekenservice: Almere, The Netherlands, 2014. [Google Scholar]
- Gallep, C.M.; Dos Santos, S.R. Photon-count during germination of wheat (Triticum aestivum) in waste water sediment solution correlated with seedling growth. Seed Sci. Technol. 2007, 35, 607–614. [Google Scholar] [CrossRef]
- Musumeci, F.; Scordino, A.; Triglia, A.; Blandino, G.; Milazzo, I. Intercellular communication during yeast cell growth. Europhys. Lett. 1999, 47, 736. [Google Scholar] [CrossRef]
- Grasso, F.; Grillo, C.; Musumeci, F.; Triglia, A.; Rodolico, G.; Cammisuli, F.; Rinzivillo, C.; Fragati, G.; Santuccio, A.; Rodolico, M. Photon emission from normal and tumor human tissue. Experientia 1992, 48, 10–13. [Google Scholar] [CrossRef]
- Mauburov, S.N. Photonic Communications in Biological Systems. J. Samara State Tech. Univ. Ser. Phys. Math. Sci. 2011, 15, 260–265. [Google Scholar]
- Kucera, O.; Cifra, M. Cell-to-cell signaling through light: Just a ghost of chance? Cell Comm. Signal. 2013, 11, 87. [Google Scholar] [CrossRef]
- Fels, D. Cellular Communication through light. PLoS ONE 2009, 4, e5086. [Google Scholar] [CrossRef]
- Beloussov, L.V.; Burlakov, A.B.; Louchinskaia, N.N. Biophotonic Pattern of optical interaction between fish eggs and embryos. Indian J. Exp. Biol. 2003, 41, 424–430. [Google Scholar]
- Tang, R.; Dai, J. Biophoton signal transmission and processing in the brain. J. Photochem. Photobiol. B Biol. 2014, 139, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Boone, K.; Tuszynski, J.; Barclay, P.; Simon, C. Possible existence of optical communication channels in the brain. Sci. Rep. 2016, 6, 36508. [Google Scholar] [CrossRef]
- Zangari, A.; Micheli, D.; Galeazzi, R.; Tozzi, A. Node of Ranvier as an Array of Bio-Nanoantennas for Infrared Communication in Nerve Tissue. Sci. Rep. 2018, 8, 539. [Google Scholar] [CrossRef]
- Cifra, M.; Brouder, C.; Nerudova, M.; Kucera, O. Biophotons, coherence and photocount statistics: A critical review. J. Lumin. 2015, 164, 38–51. [Google Scholar] [CrossRef]
- Duan, S.; Wang, F.; Zhang, Y. Research on the biophoton emission of wheat kernels based on permutation entropy. Opt. Int. J. Light Electron Opt. 2019, 178, 723–730. [Google Scholar] [CrossRef]
- Bandt, C.; Pompe, B. Permutation Entropy: A Natural Complexity Measure for Time Series. Phys. Rev. Lett. 2002, 88, 174102. [Google Scholar] [CrossRef] [PubMed]
- Falconi, M.; Loreto, V.; Vulpiani, A. Kolmogorov Legacy about Entropy, Chaos and Complexity; Springer: Berlin/Heidelberg, Germany, 2003; pp. 85–108. [Google Scholar]
- Latora, V.; Baranger, M. Kolomogorov-Sinai Entropy Rate versus Physical Entropy. Phys. Rev. Lett. 1999, 82, 520. [Google Scholar] [CrossRef]
- Allegrini, P.; Benci, V.; Grigolini, P.; Hamilton, P.; Ignaccolo, M.; Menconi, G.; Palatella, L.; Raffaelli, G.; Scafetta, N.; Virgilio, M.; et al. Compression and diffusion: A joint approach to detect complexity. Chaos Solitons Fractals 2003, 15, 517–535. [Google Scholar] [CrossRef]
- Scafetta, N.; Hamilton, P.; Grigolini, P. The thermodynamics of social processes: The teen birth phenomenon. Fractals 2001, 9, 193–208. [Google Scholar] [CrossRef]
- Scafetta, N.; Grigolini, P. Scaling detection in time series: Diffusion Entropy analysis. Phys. Rev. E 2002, 66, 036130. [Google Scholar] [CrossRef] [PubMed]
- Culbreth, G.; West, B.J.; Grigolini, P. Entropic Approach to the Detection of Crucial Events. Entropy 2019, 21, 178. [Google Scholar] [CrossRef]
- Grigolini, P. Emergence of biological complexity: Criticality, renewal and memory. Chaos Solitons Fractals 2015, 81, 575–588. [Google Scholar] [CrossRef]
- Bohara, G.; West, B.J.; Grigolini, P. Bridging Waves and Crucial Events in the Dynamic of the Brain. Front. Physiol. 2018, 9, 1174. [Google Scholar] [CrossRef]
- Attanasi, A.; Cavagna, A.; Del Castello, L.; Giardina, I.; Melillo, S.; Parisi, L.; Pohl, O.; Rossaro, B.; Shen, E.; Silvestri, E.; et al. Finite-Size Scaling as a Way to Probe Near-Criticality in Natural Swarms. Phys. Rev. Lett. 2014, 113, 238102. [Google Scholar] [CrossRef]
- Vanni, F.; Luković, M.; Grigolini, P. Criticality and Transmission of Information in a Swarm of Cooperative Units. Phys. Rev. Lett. 2011, 107, 078103. [Google Scholar] [CrossRef] [PubMed]
- Contoyiannis, Y.F.; Diakonos, F.K.; Malakis, A. Intermittent Dynamics of Critical Fluctuations. Phys. Rev. Lett. 2002, 89, 035701. [Google Scholar] [CrossRef] [PubMed]
- Schuster, H.G. Deterministic Chaos; VCH: New York, NY, USA, 1988. [Google Scholar]
- Bohara, G.; Lambert, D.; West, B.J.; Grigolini, P. Crucial events, randomness and multifractality in heartbeat. Phys. Rev. E 2017, 96, 062216. [Google Scholar] [CrossRef] [PubMed]
- Mahmoodi, K.; West, B.J.; Grigolini, P. Self-Organizing Complex Networks: Individual versus global rules. Front. Physiol. 2017, 8, 478. [Google Scholar] [CrossRef] [PubMed]
- Mahmoodi, K.; West, B.J.; Grigolini, P. Self-Organized Temporal Criticality: Bottom-up resilience versus top-down vulnerability. Complexity 2018, 2018, 8139058. [Google Scholar] [CrossRef]
- Mahmoodi, K.; Grigolini, P.; West, B.J. On social sensitivity to either zealot or independent minorities. Chaos Solitons Fractals 2018, 110, 185–190. [Google Scholar] [CrossRef]
- Photon Counting Head H12386-210 Datasheet. Available online: https://www.hamamatsu.com/eu/en/product/type/H12386-210/index.html (accessed on 20 February 2021).
- Test Sheet Hamamatsu for the Phototube H12386-210, Serial Number 30050260. Available online: https://www.hamamatsu.com/resources/pdf/etd/H12386_TPMO1073E.pdf (accessed on 20 February 2021).
- Fox, M. Quantum Optics—An Introduction; OUP Oxford: Oxford, UK, 2006. [Google Scholar]
- Stanley, H.E.; Buldyrev, S.V.; Goldberger, A.L.; Goldberger, Z.D.; Havlin, S.; Mantegna, R.N.; Ossadnik, S.M.; Peng, C.K.; Simons, M. Statistical mechanics in biology: How ubiquitous are long-range correlations? Physica A 1994, 205, 214–253. [Google Scholar] [CrossRef]
- Hurst, H.E. Long-term storage capacity of reservoirs. Trans. Am. Soc. Civ. Eng. 1951, 116, 770–799. [Google Scholar] [CrossRef]
- Mandelbrot, B.B.; Walls, J.R. Noah, Joseph and operational hydrology. Water Resourc. Res. 1968, 4, 909. [Google Scholar] [CrossRef]
- Cakir, R.A.Ş.İ.T.; Grigolini, P.; Krokhin, A.A. Dynamical origin of memory and renewal. Phys. Rev. E 2006, 74, 021108. [Google Scholar] [CrossRef] [PubMed]
- Weiss, U. Quantum Dissipative Systems, 2nd ed.; World Scientific: Singapore, 1992. [Google Scholar]
- Grigolini, P.; Palatella, L.; Raffaelli, G. Anomalous Diffusion: An Efficient Way to Detect Memory in Time Series. Fractals 2001, 9, 439–449. [Google Scholar] [CrossRef]
- Paradisi, P.; Allegrini, P.; Gemignani, A.; Laurino, M.; Menicucci, D.; Piarulli, A. Scaling and intermittency of brain events as a manifestation of consciousness. In AIP Conference Proceedings; American Institute of Physics: College Park, MD, USA, 2013; Volume 1510, p. 151. [Google Scholar] [CrossRef]
- Allegrini, P.; Menicucci, D.; Bedini, R.; Fronzoni, L.; Gemignani, A.; Grigolini, P.; West, B.J.; Paradisi, P. Spontaneous brain activity as a source of ideal 1/f noise. Phys. Rev. E 2009, 80, 061914. [Google Scholar] [CrossRef] [PubMed]
- Barenblatt, G.I. Scaling, Self-Similarity and Intermediate Asymptotics; Cambridge Press: Cambridge, UK, 1996. [Google Scholar]
- Culbreth, G.; West, B.J.; Grigolini, P. Caputo Fractional Derivative versus Quantum Coherence. Entropy 2021, 23, 211. [Google Scholar] [CrossRef]
- Mori, H. Transport, Collective Motion and Brownian Motion. Prog. Theor. Phyiscs 1965, 33, 423. [Google Scholar] [CrossRef]
- Scafetta, N.; Latora, V.; Grigolini, P. Levy scaling: The diffusion entropy analysis applied to DNA sequences. Phys. Rev. E 2002, 66, 031906. [Google Scholar] [CrossRef]
- Vanni, F.; Grigolini, P. Music as a mirror of mind. In Esthétique de la Complexité; Hermann: Paris, France, 2017. [Google Scholar]
- Pease, A.; Mahmoodi, K.; West, B.J. Complexity measures of music. Chaos Solitons Fractals 2018, 108, 82–86. [Google Scholar] [CrossRef]
- Jelinek, H.F.; Tuladhar, R.; Culbreth, G.; Bohara, G.; Cornforth, D.; West, B.J.; Grigolini, P. Diffusion Entropy versus Multiscale and Renyi Entropy to detect progression of Autonomic Neuropathy. Front. Physiol. 2020, 11, 607324. [Google Scholar] [CrossRef] [PubMed]
- Lukovic, M.; Grigolini, P. Power spectra for both interrupted and perennial aging process. J. Chem. Phys. 2008, 129, 184102. [Google Scholar] [CrossRef]
- Dlask, M.; Kukal, J.; Poplová, M.; Sovka, P.; Cifra, M. Short–time fractal analysis of biological autoluminescence. PLoS ONE 2019, 14, e0214427. [Google Scholar] [CrossRef]
- Miller, W.B., Jr. Biological information systems: Evolution as cognition-based information management. Prog. Biophys. Mol. Biol. 2018, 134, 1–26. [Google Scholar] [CrossRef]
- Eigen, M. From Strange Simplicity to Complex Familiarity: A Treatise on Matter, Information, Life and Thought; OUP: Oxford, UK, 2013. [Google Scholar]
- Ford, B.J. On Intelligence in Cells: The Case for Whole Cell Biology. Interdiscip. Sci. Rev. 2009, 34, 350–365. [Google Scholar] [CrossRef]
- Dodig-Crnkovic, G. Modeling Life as Cognitive Info-Computation. In Language, Life, Limits; Beckmann, A., Csuhaj-Varj, E., Meer, K., Eds.; Springer: Cham, Switzerland, 2014; Volume 8493. [Google Scholar]
- Maturana, H.; Varela, F. Autopoiesis and Cognition: The Realization of the Living; D. Reidel Pub. Co.: Dordrecht, The Netherlands, 1980. [Google Scholar]
- Mahmoodi, K.; West, B.J.; Grigolini, P. Complexity Matching and Requisite Variety. arXiv 2019, arXiv:1806.08808. [Google Scholar]
- Popp, F.A. Consciousness as Evolutionary Process Based on Coherent States. NeuroQuantology 2008, 6, 431. [Google Scholar] [CrossRef]
- Piao, D. On the stress-induced photon emission from organism: II, how will the stress-transfer kinetics affect the photo-genesis? SN Appl. Sci. 2020, 6, 1556. [Google Scholar] [CrossRef]
- Piao, D. On the stress-induced photon emission from organism: I, will the scattering-limited delay affect the temporal course? SN Appl. Sci. 2020, 2, 1566. [Google Scholar] [CrossRef]
- Mancuso, S. The Revolutionary Genius of Plants: A New Understanding of Plant Intelligence and Behaviour; Simon and Shuster: New York, NY, USA, 2018. [Google Scholar]
- Mancuso, S.; Viola, A. Brilliant Green: The Surprising History and Science of Plant Intelligence; Island Press: Washington, DC, USA, 2015. [Google Scholar]
- Van Wijk, R.; Van Wijk, E.; Pang, J.; Yang, M.; Yan, Y.; Han, J. Integrating Ultra-Weak Photon Emission Analysis in Mitochondrial Research. Front. Physiol. 2020, 11, 717. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Gouripeddi, P.; Devireddy, H.R.N.; Ovsii, A.; Rachakonda, D.P.; Wijk, R.V.; Pospíšil, P. Spectral Distribution of Ultra-Weak Photon Emission as a Response to Wounding in Plants: An in Vivo Study. Biology 2020, 9, 139. [Google Scholar] [CrossRef] [PubMed]
- Dal Lin, C.; Falanga, M.; De Lauro, E.; De Martino, S.; Vitiello, G. Biochemical and biophysical mechanisms underlying the heart and the brain dialog. Biophysics 2020, 8, 1–33. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exp. <n> | Exp. σ2 | Poisson <n> | |
|---|---|---|---|
| dark | 1.56 | 2.24 | 0.827 ± 0.03 |
| 1 | 14.40 | 14.60 | 14.51 ± 0.10 |
| 2 | 26.83 | 29.12 | 26.80 ± 0.21 |
| 3 | 22.21 | 24.84 | 21.90 ± 0.13 |
| Without Stripes | With Stripes | |||
|---|---|---|---|---|
| dark counts | 0.575 | 2.739 | 0.508 | 2.968 |
| region #1 | 0.773 | 2.293 | 0.596 | 2.677 |
| region #2 | 0.796 | 2.254 | 0.558 | 2.792 |
| region #3 | 0.736 | 2.358 | 0.526 | 2.901 |
| region #4 | 0.737 | 2.355 | 0.496 | 3.016 |
| region #5 | 0.694 | 2.440 | 0.509 | 2.964 |
| region #6 | 0.725 | 2.377 | 0.504 | 2.984 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benfatto, M.; Pace, E.; Curceanu, C.; Scordo, A.; Clozza, A.; Davoli, I.; Lucci, M.; Francini, R.; De Matteis, F.; Grandi, M.; et al. Biophotons and Emergence of Quantum Coherence—A Diffusion Entropy Analysis. Entropy 2021, 23, 554. https://doi.org/10.3390/e23050554
Benfatto M, Pace E, Curceanu C, Scordo A, Clozza A, Davoli I, Lucci M, Francini R, De Matteis F, Grandi M, et al. Biophotons and Emergence of Quantum Coherence—A Diffusion Entropy Analysis. Entropy. 2021; 23(5):554. https://doi.org/10.3390/e23050554
Chicago/Turabian StyleBenfatto, Maurizio, Elisabetta Pace, Catalina Curceanu, Alessandro Scordo, Alberto Clozza, Ivan Davoli, Massimiliano Lucci, Roberto Francini, Fabio De Matteis, Maurizio Grandi, and et al. 2021. "Biophotons and Emergence of Quantum Coherence—A Diffusion Entropy Analysis" Entropy 23, no. 5: 554. https://doi.org/10.3390/e23050554
APA StyleBenfatto, M., Pace, E., Curceanu, C., Scordo, A., Clozza, A., Davoli, I., Lucci, M., Francini, R., De Matteis, F., Grandi, M., Tuladhar, R., & Grigolini, P. (2021). Biophotons and Emergence of Quantum Coherence—A Diffusion Entropy Analysis. Entropy, 23(5), 554. https://doi.org/10.3390/e23050554