Incorporating Cellular Stochasticity in Solid–Fluid Mixture Biofilm Models



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
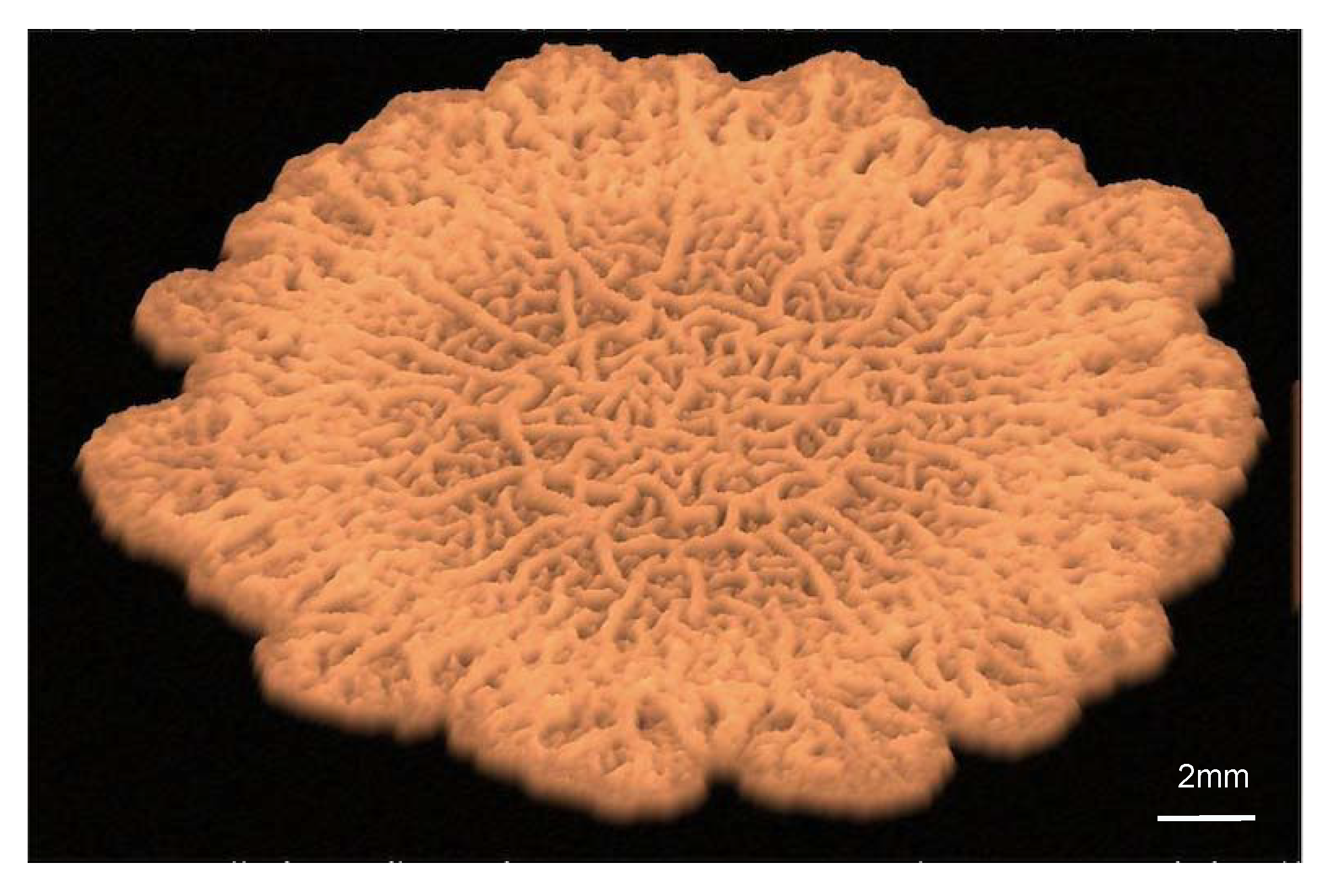
1. Introduction
2. Solid-Fluid Mixture Model of a Biofilm Spreading on an Agar/Air Interface
2.1. Mass Balance
2.2. Driving Forces
2.2.1. Stresses in the Solid and the Fluid
2.2.2. Interaction and Inertial Forces
2.3. Equations of Motion
2.4. Final Equations
2.5. Motion of the Air–Biofilm Interface
2.6. Motion of the Agar/Biofilm Interface
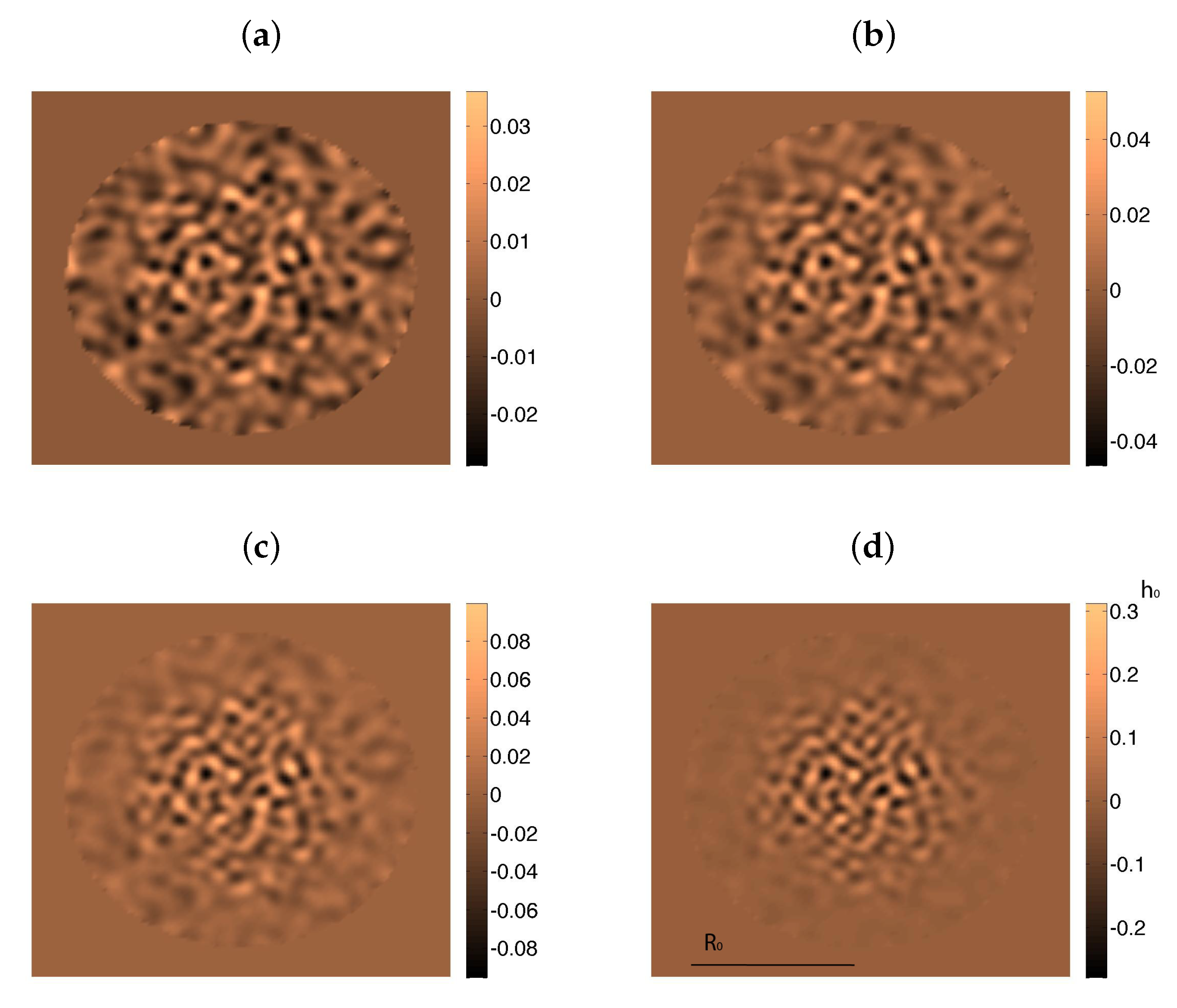
3. Incorporating Cellular Behavior
3.1. Cellular Automata and Dynamic Energy Budget
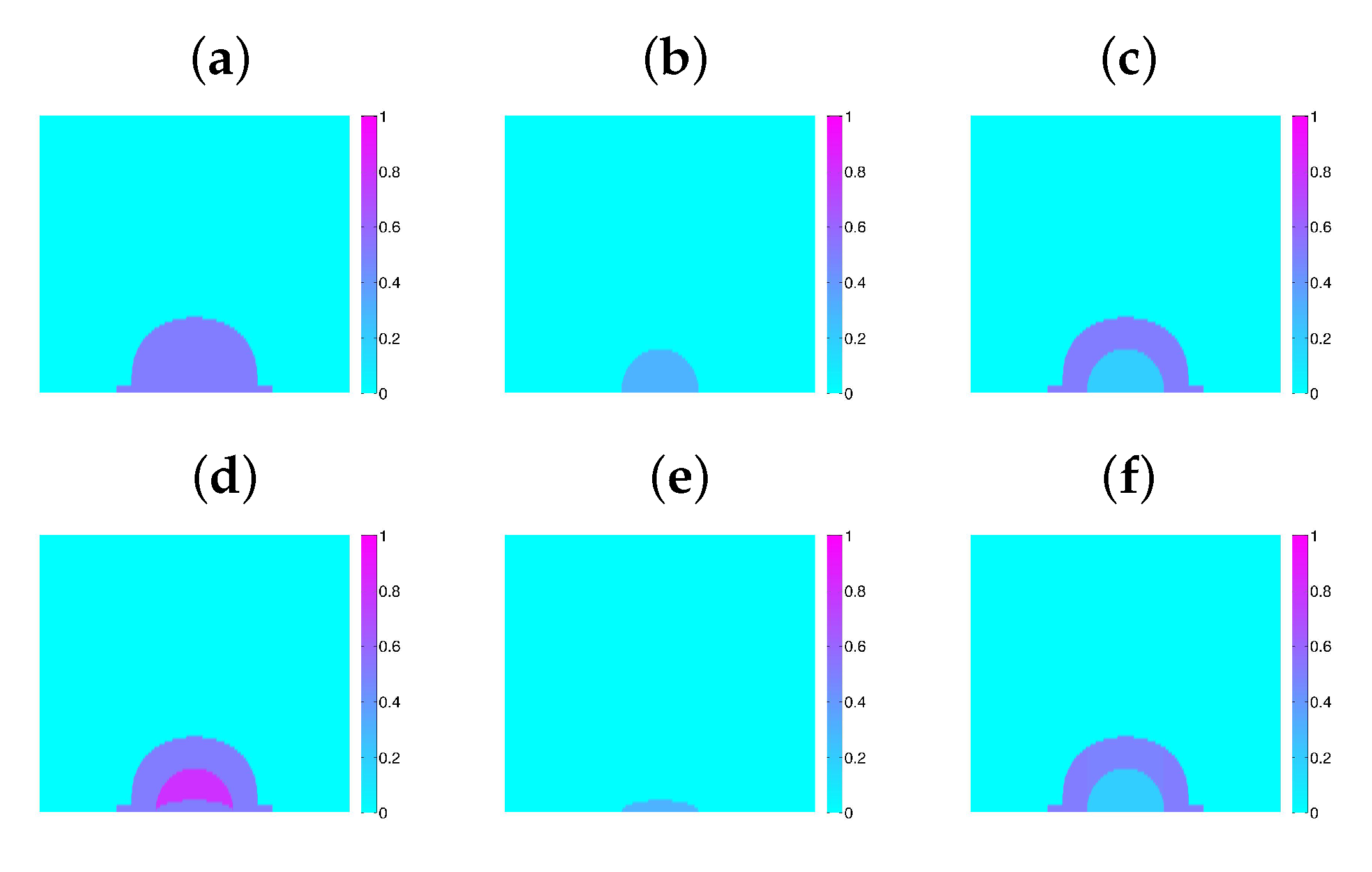
- We set an initial distribution of N bacteria characterized by their energies , volumes , damage , hazard , acclimation , and attached EPS volume , .
- Each bacterium is initially classified as normal, surfactin producer, EPS producer, or inert. Bacteria are distributed in the tiles of the grid. The empty space around them is filled with water and dissolved substances. In this way, we may compute the volume fractions of biomass and fluid in each tile , as well as the osmotic pressure . The pressure is obtained from (13) with and from (29).
- We compute stationary solutions of the Equation (31) for by a relaxation numerical scheme. All except the equation for are solved using the grid defining with no flux boundary conditions. The equation for is solved in the biofilm–agar domain, that is, , imposing continuity of concentrations and fluxes at the agar–biofilm interface and no flux boundary conditions at the air–biofilm interface.
- We update the bacterial status checking whether normal bacteria become surfactin or EPS producers, whether any of them deactivates or dies, and whether they divide, with the probabilities assigned to each situation. In case a bacterium divides, we reallocate the newborn cell.
- In the resulting biofilm configuration , we compute the volume fractions of biomass and fluid in each tile. This also provides the osmotic pressure The fluid pressure is obtained from (13), the displacements from (13), and from (29). The solid velocities are approximated by . Then, the fluid velocity is given by (9).
- We yet need to take into account water absorption from agar. To do so, we solve in the biofilm/agar system. Alternatively, we can solve only in the biofilm, using at the biofilm/agar interface and for boundary conditions, where h and R are reference values for the biofilm height and radius. Then, we revise the biofilm configuration, creating water tiles with probability and shifting the contains of the neighbouring tiles. This provides the final biofilm configuration , that is, the occupied tiles, their contents, the bacterial status and fields, as well as the values of the continuous fields at each tile.
3.2. Balance Equation Approach
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Hoiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Cioufu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S. Mechanisms of antibiotic resistance in bacterial biofilms. Int. J. Med. Microbiol. 2002, 292, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Storck, T.; Picioreanu, C.; Virdis, B.; Batstone, D.J. Variable cell morphology approach for individual-based modeling of microbial communities. Biophys. J. 2014, 106, 2037–2048. [Google Scholar] [CrossRef] [PubMed]
- Grant, M.A.A.; Waclaw, B.; Allen, R.J.; Cicuta, P. The role of mechanical forces in the planar-to-bulk transition in growing Escherichia coli microcolonies. J. R. Soc. Interface 2014, 11, 20140400. [Google Scholar] [CrossRef] [PubMed]
- Laspidou, C.S.; Rittmann, B.E. Modeling the development of biofilm density including active bacteria, inert biomass, and extracellular polymeric substances. Water Res. 2004, 38, 3349–3361. [Google Scholar] [CrossRef]
- Kærn, M.; Elston, T.C.; Blake, W.J.; Collins, J.J. Stochasticity in gene expression: From theories to phenotypes. Nat. Rev. Genet. 2005, 6, 451–464. [Google Scholar] [CrossRef]
- Wilkinson, D.J. Stochastic modelling for quantitative description of heterogeneous biological systems. Nat. Rev. Genet. 2009, 10, 122–133. [Google Scholar] [CrossRef]
- Birnir, B.; Carpio, A.; Cebrián, E.; Vidal, P. Dynamic energy budget approach to evaluate antibiotic effects on biofilms. Commun. Nonlinear Sci. Numer. Simul. 2018, 54, 70–83. [Google Scholar] [CrossRef]
- Chai, L.; Vlamakis, H.; Kolter, R. Extracellular signal regulation of cell differentiation in biofilms. MRS Bull. 2011, 36, 374–379. [Google Scholar] [CrossRef]
- Seminara, A.; Angelini, T.E.; Wilking, J.N.; Vlamakis, H.; Ebrahim, S.; Kolter, R.; Weitz, D.A.; Brenner, M.P. Osmotic spreading of Bacillus subtilis biofilms driven by an extracellular matrix. Proc. Natl. Acad. Sci. USA 2012, 109, 1116–1121. [Google Scholar] [CrossRef] [PubMed]
- Asally, M.; Kittisopikul, M.; Rué, P.; Du, Y.; Hu, Z.; Çağatay, T.; Robinson, A.B.; Lu, H.; Garcia-Ojalvo, J.; Süel, G.M. Localized cell death focuses mechanical forces during 3D patterning in a biofilm. Proc. Natl. Acad. Sci. USA 2012, 109, 18891–18896. [Google Scholar] [CrossRef] [PubMed]
- Espeso, D.R.; Carpio, A.; Einarsson, B. Differential growth of wrinkled biofilms. Phys. Rev. E 2015, 91, 022710. [Google Scholar] [CrossRef] [PubMed]
- Wilking, J.N.; Zaburdaev, V.; De Volder, M.; Losick, R.; Brenner, M.P.; Weitz, D.A. Liquid transport facilitated by channels in Bacillus subtilis biofilms. Proc. Natl. Acad. Sci. USA 2013, 110, 848–852. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Fei, C.; Mao, S.; Moreau, A.; Wingreen, N.S.; Kosmrlj, A.; Stone, H.A.; Bassler, B.L. Mechanical instability and interfacial energy drive biofilm morphogenesis. eLife 2019, 8, e43920. [Google Scholar] [CrossRef]
- Zhang, C.; Li, B.; Huang, X.; Ni, Y.; Feng, X.Q. Morphomechanics of bacterial biofilms undergoing anisotropic differential growth. Appl. Phys. Lett. 2016, 109, 143701. [Google Scholar] [CrossRef]
- Ben Amar, M.; Wu, M. Patterns in biofilms: From contour undulations to fold focussing. Europhys. Lett. 2014, 108, 38003. [Google Scholar] [CrossRef]
- Carpio, A.; Cebrián, E.; Vidal, P. Biofilms as poroelastic materials. Int. J. Non-Linear Mech. 2019, 109, 1–8. [Google Scholar] [CrossRef]
- Drescher, K.; Shen, Y.; Bassler, B.L.; Stone, H.A. Biofilm streamers cause catastrophic disruption of flow with consequences for environmental and medical systems. Proc. Natl. Acad. Sci. USA 2013, 110, 4345–4350. [Google Scholar] [CrossRef]
- Kreft, J.U.; Picioreanu, C.; Wimpenny, J.W.T.; van Loosdrecht, M.C.M. Individual-based modelling of biofilms. Microbiology 2001, 147, 2897–2912. [Google Scholar] [CrossRef]
- Jayathilake, P.G.; Gupta, P.; Li, B.; Madsen, C.; Oyebamiji, O.; González-Cabaleiro, R.; Rushton, S.; Bridgens, B.; Swailes, D.; Allen, B.; et al. A mechanistic Individual-based Model of microbial communities. PLoS ONE 2017, 12, e0181965. [Google Scholar] [CrossRef] [PubMed]
- Lanir, Y. Biorheology and fluid flux in swelling tissues. I. Bicomponent theory for small deformations, including concentration effects. Biorheology 1987, 24, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Shaw, T.; Winston, M.; Rupp, C.J.; Klapper, I.; Stoodley, P. Commonality of elastic relaxation times in biofilms. Phys. Rev. Lett. 2004, 93, 098102. [Google Scholar] [CrossRef] [PubMed]
- Charlton, S.G.V.; White, M.A.; Jana, S.; Eland, L.E.; Jayathilake, P.G.; Burgess, J.G.; Chen, J.; Wipat, A.; Curtis, T.P. Regulating, measuring, and modeling the viscoelasticity of bacterial biofilms. J. Bacteriol. 2019, 201, e00101-19. [Google Scholar] [CrossRef]
- Burridge, R.; Keller, J.B. Poroelasticity equations derived from microstructure. J. Acoust. Soc. Am. 1981, 70, 1140–1146. [Google Scholar] [CrossRef]
- Kapellos, G.E.; Alexiou, T.S.; Payatakes, A.C. Theoretical modeling of fluid flow in cellular biological media: An overview. Math. Biosci. 2010, 225, 83–93. [Google Scholar] [CrossRef]
- Witelski, T.P. Perturbation analysis for wetting fronts in Richard’s equation. Transp. Porous Media 1997, 27, 121–134. [Google Scholar] [CrossRef]
- Wilson, W.; van Donkelaar, C.; Huyghe, J.M. A comparison between mechano-electrochemical and biphasic swelling theories for soft hydrated tissues. J. Biomech. Eng. Trans. ASME 2005, 127, 158–165. [Google Scholar] [CrossRef]
- Ghassemi, A.; Diek, A. Linear chemo-poroelasticity for swelling shales: Theory and application. J. Petrol. Sci. Eng. 2003, 38, 199–212. [Google Scholar] [CrossRef]
- Chen, G.; Gallipoli, D.; Ledesma, A. Chemo-hydro-mechanical coupled consolidation for a poroelastic clay buffer in a radioactive waste repository. Trans. Porous Med. 2007, 69, 189–213. [Google Scholar] [CrossRef]
- Sacco, R.; Causin, P.; Lelli Ch Raimondi, M.T. A poroelastic mixture model of mechanobiological processes in biomass growth: theory and application to tissue engineering. Meccanica 2017, 52, 3273–3297. [Google Scholar] [CrossRef] [PubMed]
- Wood, B.W.; Quintard, M.; Whitaker, S. Calculation of effective diffusivities for biofilms and tissues. Biotech. Bioeng. 2002, 77, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Landau, L.D.; Lifshitz, E.M. Statistical Physics, 3rd ed.; Part 1: Volume 5 (Course of Theoretical Physics, Volume 5); Pergamon Press: Oxford, UK, 1980. [Google Scholar]
- Huang, R.; Im, S.H. Dynamics of wrinkle growth and coarsening in stressed thin films. Phys. Rev. E 2006, 74, 026214. [Google Scholar] [CrossRef] [PubMed]
- Iakunin, S.; Bonilla, L.L. Variational formulation, asymptotic analysis, and finite element simulation of wrinkling phenomena in modified plate equations modeling biofilms growing on agar substrates. Comput. Methods Appl. Mech. Eng. 2018, 333, 257–286. [Google Scholar] [CrossRef]
- Mehta, P.; Mukhopadhyay, R.; Wingreen, N.S. Exponential sensitivity of noise-driven switching in genetic networks. Phys. Biol. 2008, 5, 026005. [Google Scholar] [CrossRef]
- Kooijman, S.A.L.M. Dynamic Energy Budget Theory for Metabolic Organization; Cambridge UP: Cambridge, UK, 2008. [Google Scholar]
- Stotsky, J.A.; Hammond, J.F.; Pavlovsky, L.; Stewart, E.J.; Younger, J.G.; Solomon, M.J.; Bortz, D.M. Variable viscosity and density biofilm simulations using an immersed boundary method, Part II: Experimental validation and the heterogeneous rheology-IBM. J. Comput. Phys. 2016, 317, 204–222. [Google Scholar] [CrossRef]
- Dillon, R.; Owen, M.; Painter, K. A single-cell-based model of multicellular growth using the immersed boundary method. In Moving Interface Problems and Applications in Fluid Dynamics (Contemporary Mathematics); American Mathematical Society: Providence, RI, USA, 2008; pp. 1–16. [Google Scholar]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carpio, A.; Cebrián, E. Incorporating Cellular Stochasticity in Solid–Fluid Mixture Biofilm Models. Entropy 2020, 22, 188. https://doi.org/10.3390/e22020188
Carpio A, Cebrián E. Incorporating Cellular Stochasticity in Solid–Fluid Mixture Biofilm Models. Entropy. 2020; 22(2):188. https://doi.org/10.3390/e22020188
Chicago/Turabian StyleCarpio, Ana, and Elena Cebrián. 2020. "Incorporating Cellular Stochasticity in Solid–Fluid Mixture Biofilm Models" Entropy 22, no. 2: 188. https://doi.org/10.3390/e22020188
APA StyleCarpio, A., & Cebrián, E. (2020). Incorporating Cellular Stochasticity in Solid–Fluid Mixture Biofilm Models. Entropy, 22(2), 188. https://doi.org/10.3390/e22020188