Primitive Membrane Formation, Characteristics and Roles in the Emergent Properties of a Protocell

Abstract
:1. Introduction
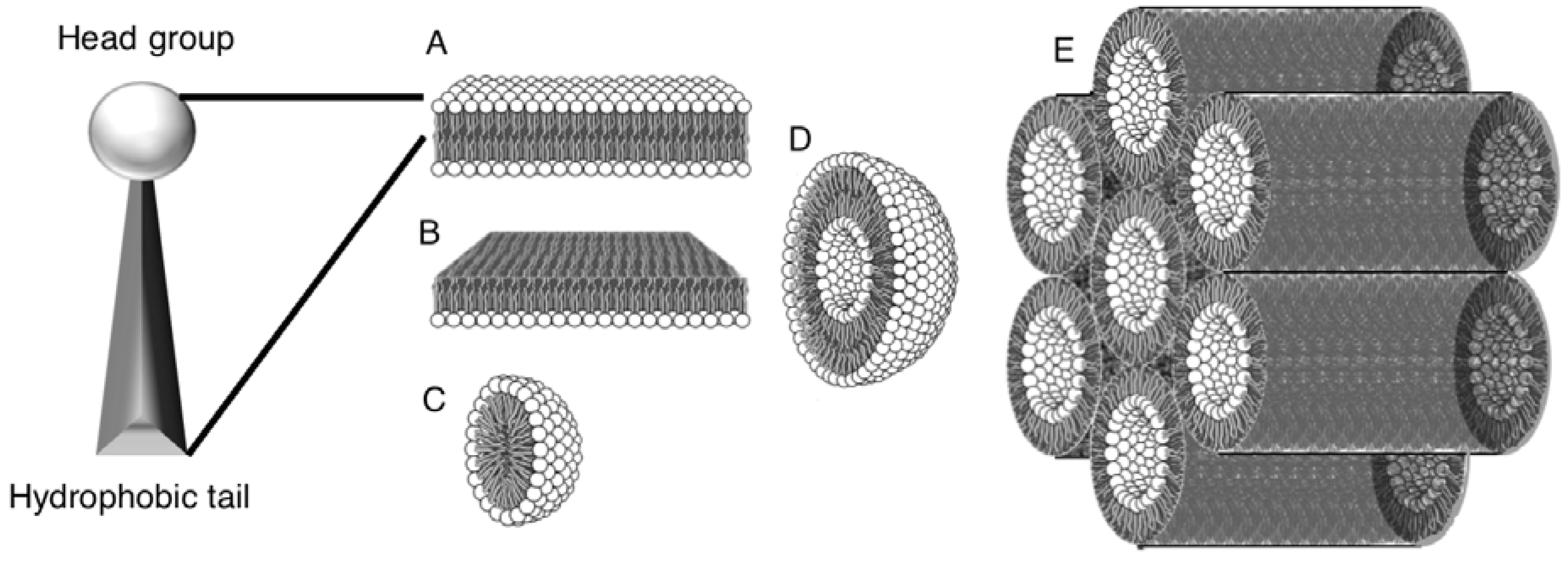
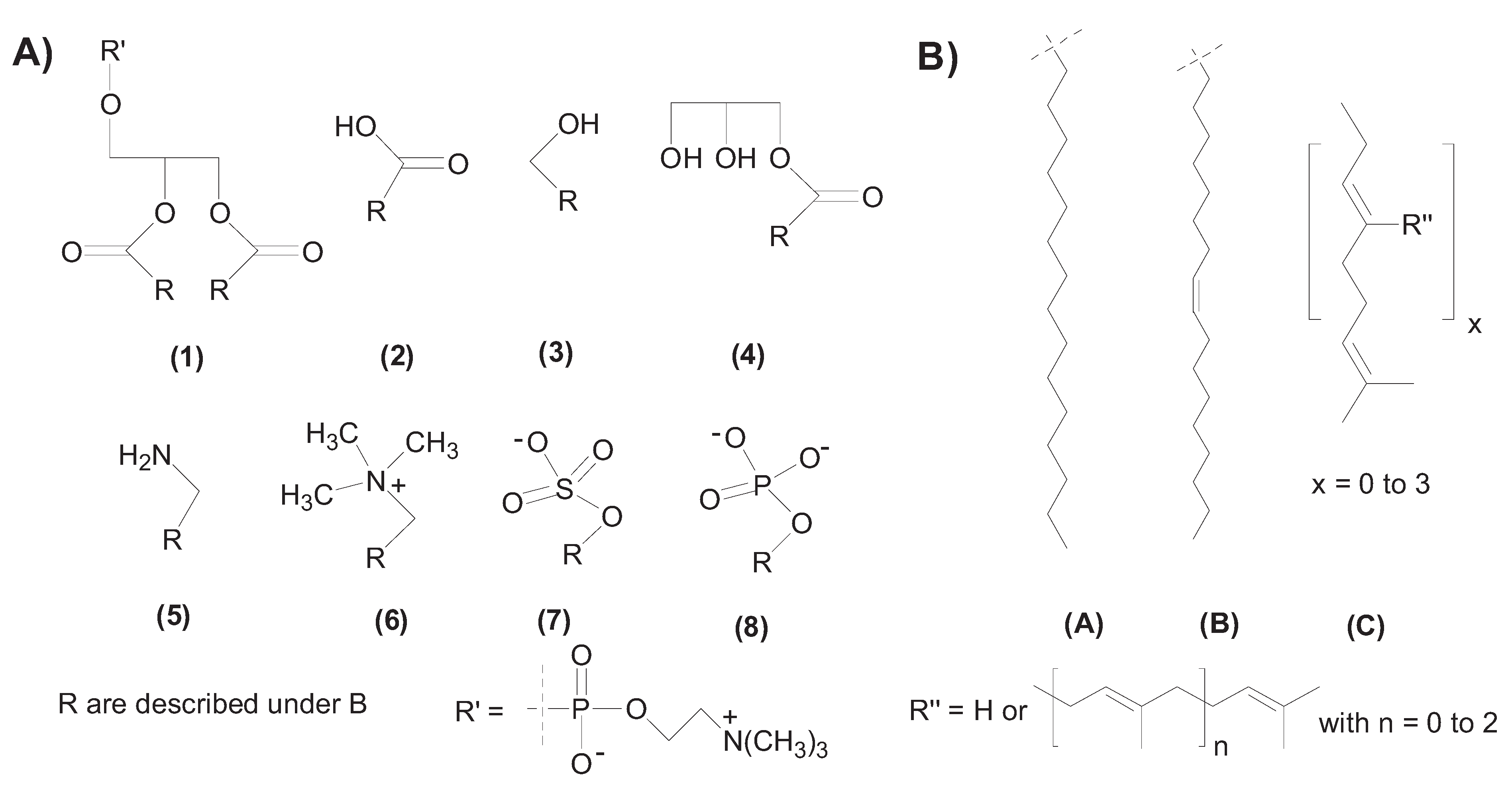
2. Self-Assembly of Amphiphile Compartments
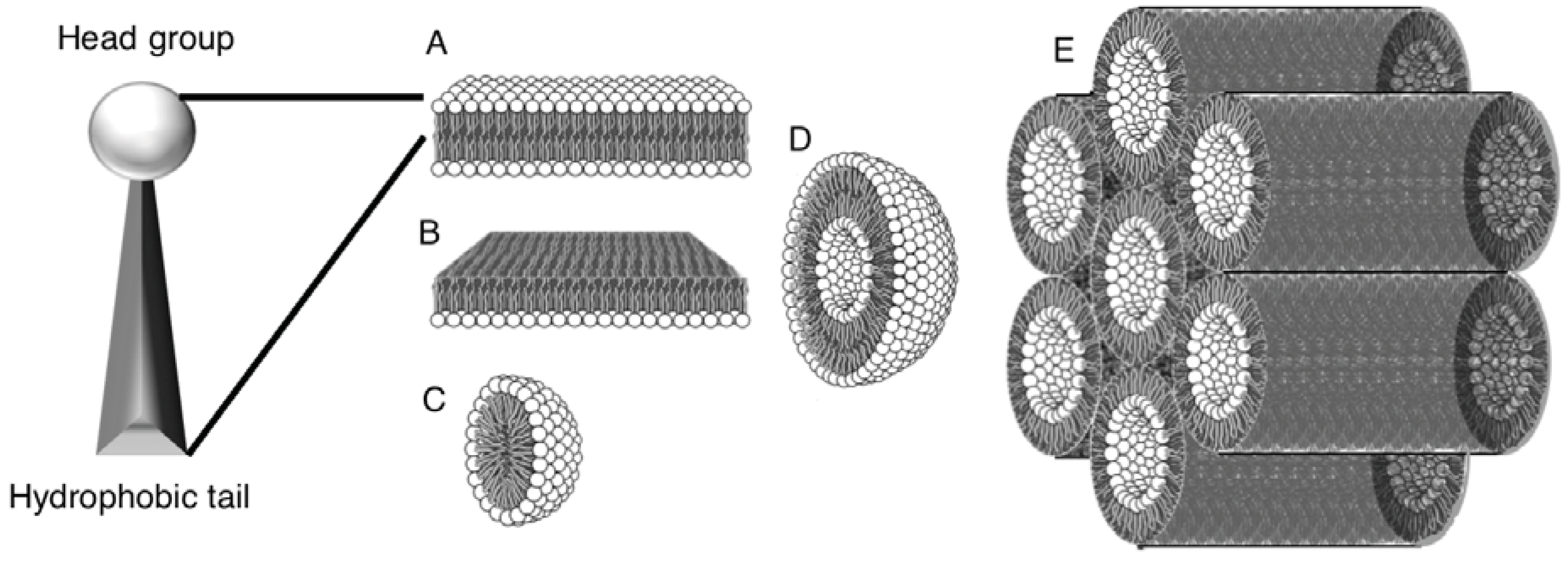
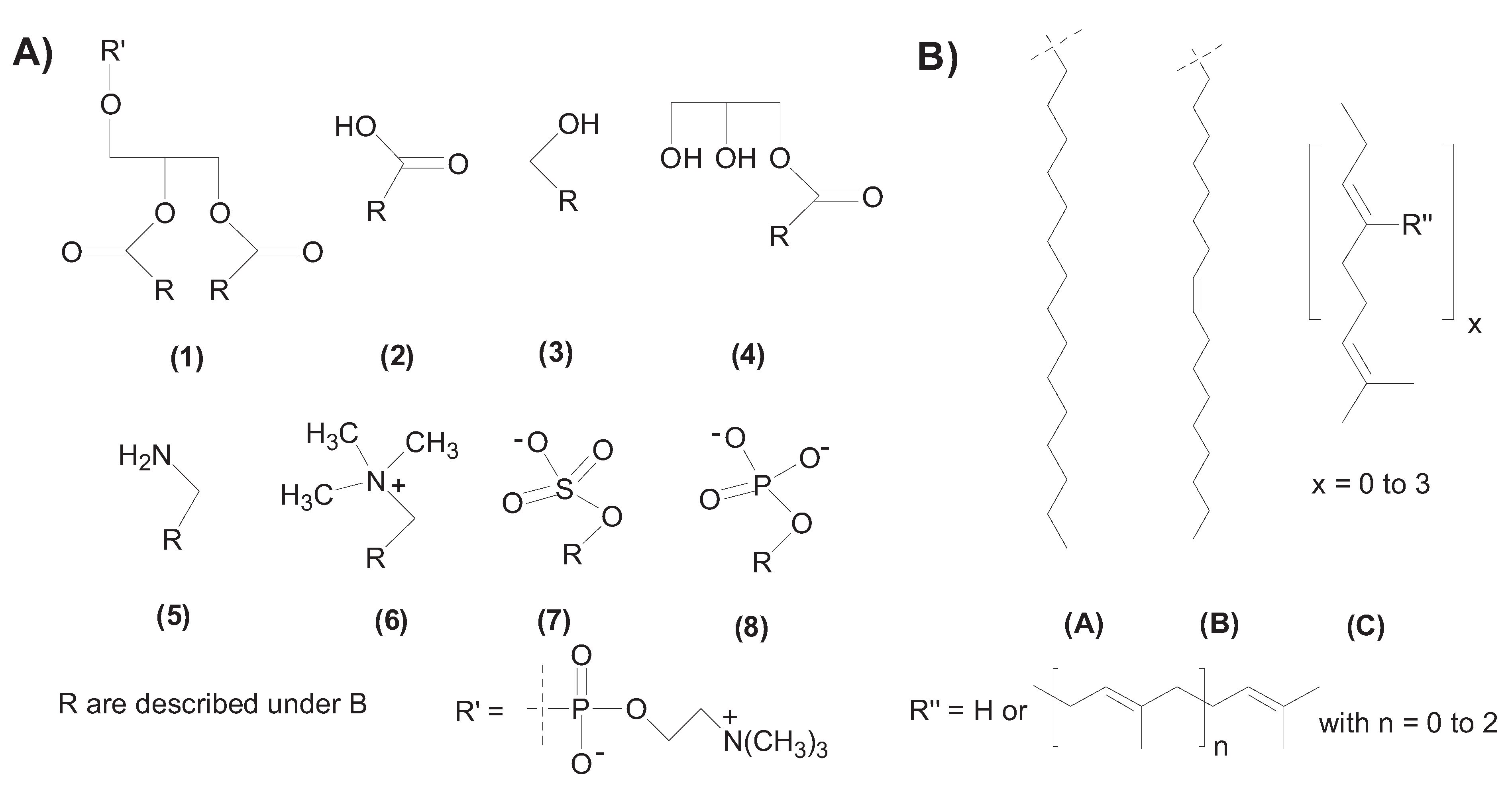
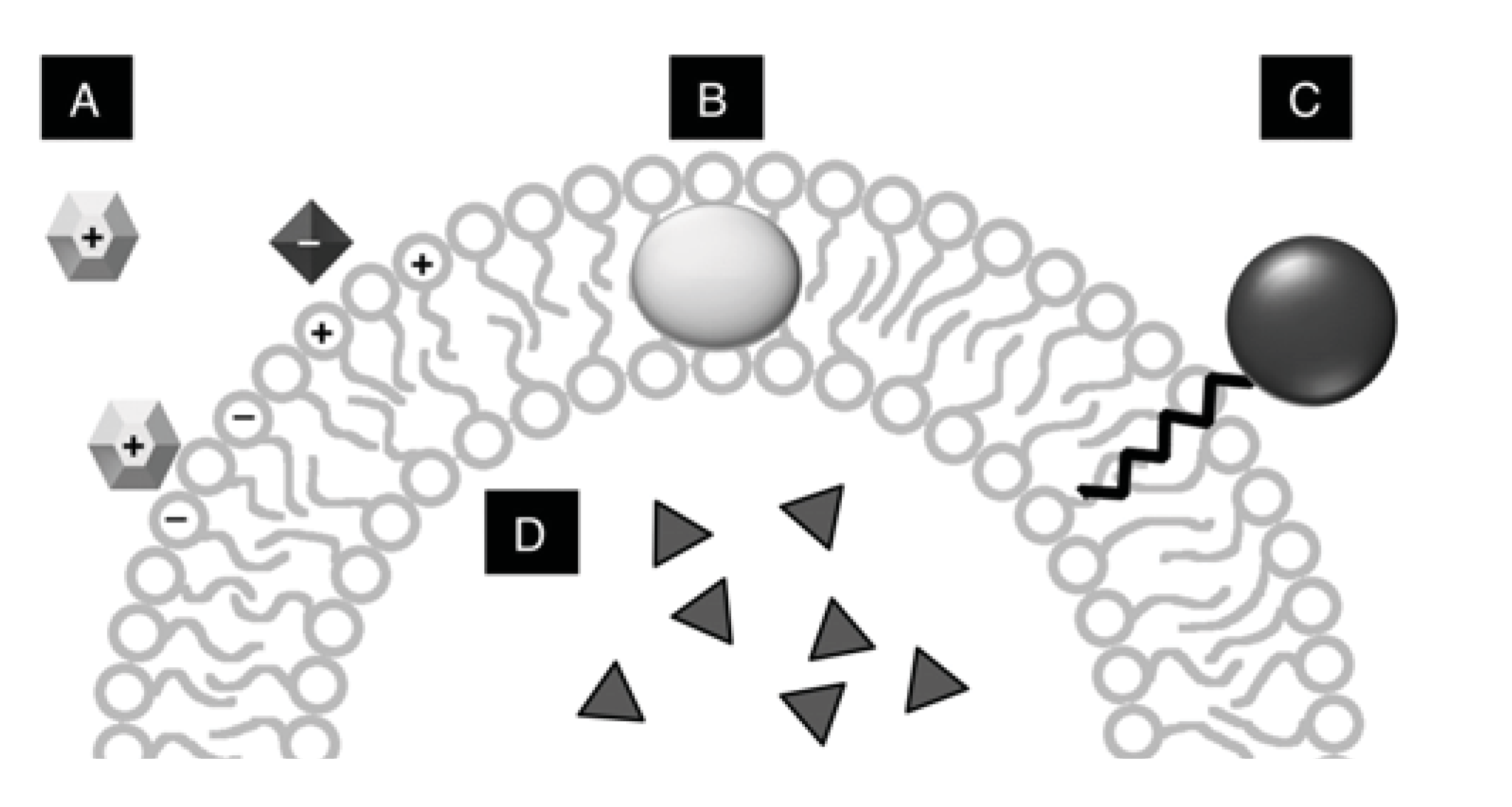
3. Vesicle Interactions with Small Chemicals and Polymers
3.1. Membrane Stability
- Environmental factors such as temperature and pressure of the solutions can change the fluidity of the amphiphiles causing precipitation or dissolution.
- Additional solutes including salts, buffers, biomolecules, and other chemicals like residual organic solvents, or catalytic cofactors can potentially decrease the hydrophobic and electrostatic interactions of bilayers causing membrane disruption. For a full summary containing various instances see for example [27].
- The composition of the membrane considering the head and tail group type and mixtures of co-surfactants will interact with the environment and solutes in species-specific manner (e.g., anionic vesicles are sensitive to cations).
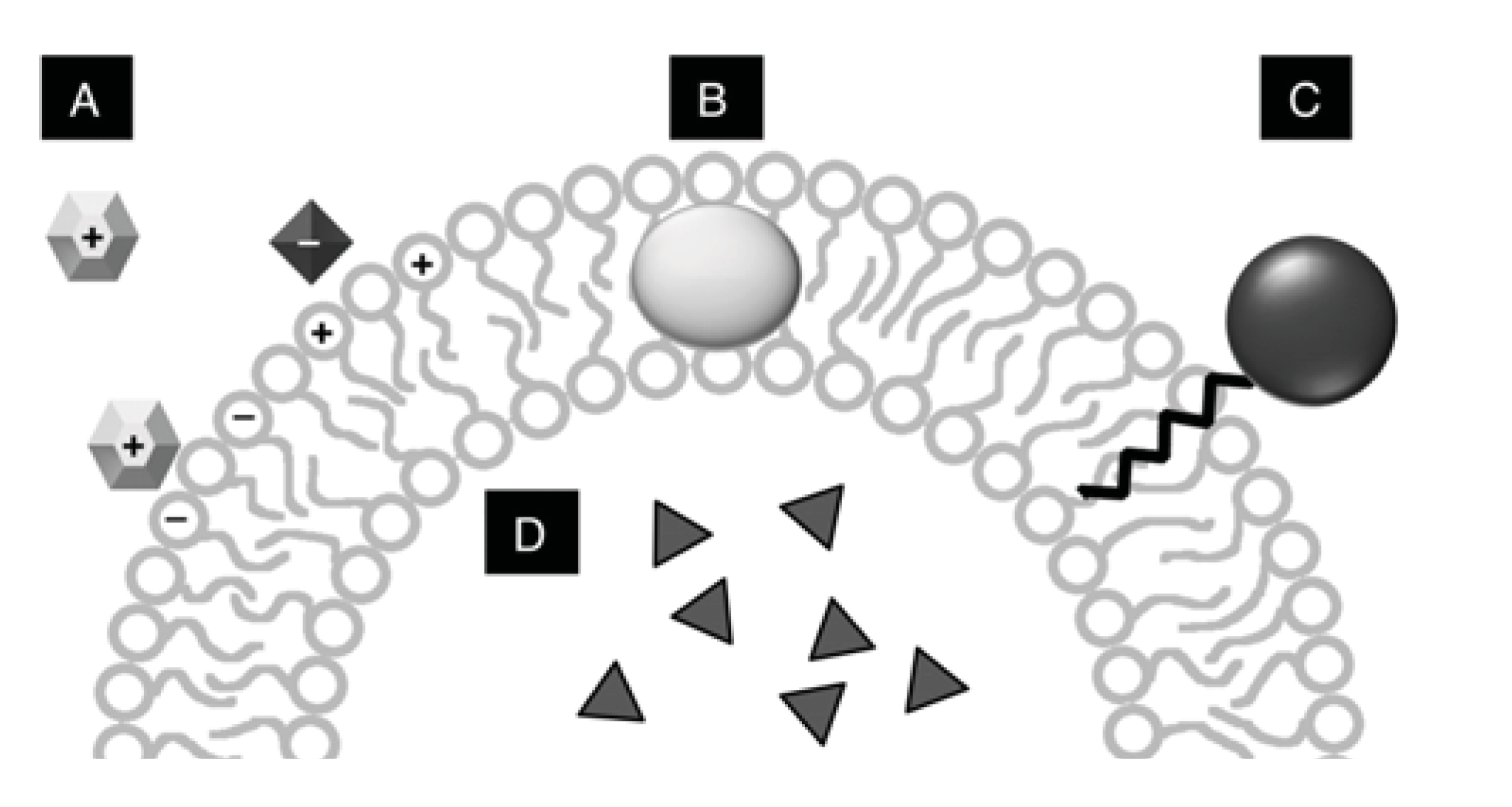

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular species | Internal | Surface association * | Hydrophobic association |
|---|---|---|---|
| DNA | X | X | |
| RNA | X | X | |
| PAH | X | ||
| Protein | X | X | X |
| Fluorescent dyes | X | X | |
| Amino acids | X | X | |
| Molecules derivatized with hydrophobic tails** | X | ||
| Sugars | X |
3.2. Interaction of Vesicles with Biomolecules
3.3. Interaction between Vesicles
4. Membrane Related Reactions
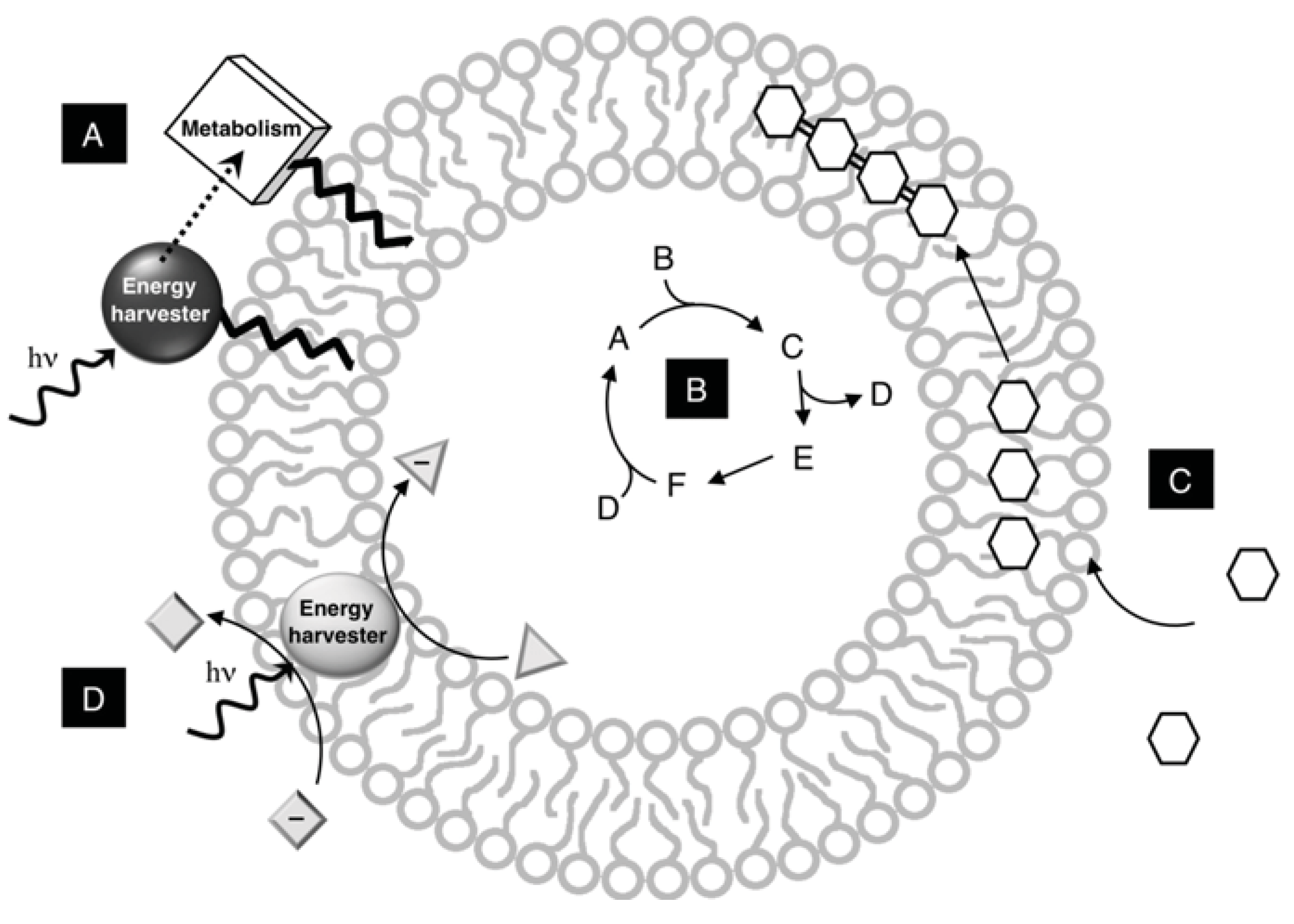
4.1. Membranes as Colocalization Systems
4.2. Membranes as Multiphase Reaction systems and Reaction Scaffolds
5. Conclusions and Outlook
Acknowledgements
References
- Szostak, J.W.; Bartel, D.P.; Luisi, P.L. Synthesizing life. Nature 2001, 409, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Luisi, P.L.; Ferri, F.; Stano, P. Approaches to semi-synthetic minimal cells: A review. Naturwissenschaften 2006, 93, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.; Chen, L.; Nilsson, M.; Abe, S. Bridging nonliving and living matter. Artif. Life 2003, 9, 269–316. [Google Scholar] [CrossRef] [PubMed]
- Agresti, J.J.; Kelly, B.T.; Jaschke, A.; Griffiths, A.D. Selection of ribozymes that catalyse multiple-turnover Diels-Alder cycloadditions by using in vitro compartmentalization. Proc. Natl. Acad. Sci. USA 2005, 102, 16170–16175. [Google Scholar] [CrossRef] [PubMed]
- Nomura, S.M.; Tsumoto, K.; Hamada, T.; Akiyoshi, K.; Nakatani, Y.; Yoshikawa, K. Gene expression within cell-sized lipid vesicles. ChemBioChem 2003, 4, 1172–1175. [Google Scholar] [CrossRef] [PubMed]
- Steinberg-Yfrach, G.; Rigaud, J.-L.; Durantini, E.N.; Moore, A.L.; Gust, D.; Moore, T.A. Light-driven production of ATP catalyzed by F0F1-ATP synthase in an artificial photosynthetic membrane. Nature 1998, 392, 479–482. [Google Scholar] [PubMed]
- Miller, S.L.; Urey, H.C. Organic compound synthesis on the primitive earth. Science 1959, 130, 7. [Google Scholar]
- Cronin, L. Inorganic molecular capsules: From structure to function. Angew. Chem. Int. Ed. 2006, 45, 3576–3578. [Google Scholar] [CrossRef] [PubMed]
- Israelachvili, J. Intermolecular and Surface Forces, 2nd ed.; Academic Press: London, UK, 1991. [Google Scholar]
- Nagarajan, R. Molecular packing parameter and surfactant self-assembly: The neglected role of the surfactant tail. Langmuir 2001, 18, 31–38. [Google Scholar] [CrossRef]
- Milsmann, M.H.W.; Schwendener, R.A.; Weder, H.-G. The preparation of large single bilayer liposomes by a fast and controlled dialysis. Biochim. Biophys. Acta 1978, 512, 147–155. [Google Scholar] [CrossRef]
- Talsma, H.; Van Steenbergen, M.; Borchert, J.C.H.; Crommelin, D.J.A. A novel technique for the One-Step preparation of liposomes and nonionic surfactant vesicles without the use of organic solvent. Liposome formation in acontinuous gas stream: the BUBBLE method. J. Pharm. Sci. 1994, 83, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Mortara, R.A.; Quina, F.H.; Chaimovich, H. Formation of closed vesciles from a simple phosphate diester. Preparation and some properties of vesicles of Dihexadecyl phosphate. Biochem. Biophys. Res. Commun. 1978, 81, 1080–1084. [Google Scholar] [CrossRef]
- Monnard, P.A.; Deamer, D.W. Preparation of vesicles from nonphospholipid amphiphiles. Meth. Enzymology 2003, 372, 133–151. [Google Scholar]
- Domazou, A.S.; Luisi, P.L. Size distribution of spontaneously formed liposomes by the alcohol injection method. J. Liposome Res. 2002, 12, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Castile, J.D.; Taylor, K.M.G. Factors affecting the size distribution of liposomes produced by freeze-thaw extrusion. Int. J. Pharm. 1999, 188, 87–95. [Google Scholar] [CrossRef]
- Maurer, S.E.; Deamer, D.W.; Boncella, J.M.; Monnard, P.A. Chemical evolution of amphiphiles: glycerol monoacyl derivatives stabilize plausible prebiotic membranes. Astrobiology 2009, 9, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, W.R.; Deamer, D.W. Liposomes from ionic, single-chain amphiphiles. Biochem. US 1978, 17, 3759–3768. [Google Scholar] [CrossRef]
- Cistola, D.P.; Atkinson, D.; Hamilton, J.A.; Small, D.M. Phase-behavior and bilayer properties of fatty-acids—hydrated 1-1 acid soaps. Biochem. US 1986, 25, 2804–2812. [Google Scholar] [CrossRef]
- Kanicky, J.R.; Shah, D.O. Effect of premicellar aggregation on the pK(a) of fatty acid soap solutions. Langmuir 2003, 19, 2034–2038. [Google Scholar] [CrossRef]
- Paula, S.; Volkov, A.G.; VanHoek, A.N.; Haines, T.H.; Deamer, D.W. Permeation of protons, potassium ions, and small polar molecules through phospholipid bilayers as a function of membrane thickness. Biophys. J. 1996, 70, 339–348. [Google Scholar] [CrossRef]
- McMullen, T.P.W.; Lewis, R.N.A.H.; McElhaney, R.N. Differential scanning calorimetric study of the effect of cholesterol on the thermotropic phase behavior of a homologous series of linear saturated phosphatidylcholines. Biochem. US 1993, 32, 516–522. [Google Scholar] [CrossRef]
- Komatsu, H.; Okada, S. Ethanol-enhanced permeation of phosphatidylcholine/ phosphatidylethanolamine mixed liposomal membranes due to ethanol-induced lateral phase separation. Biochim. Biophys. Acta 1996, 1283, 73–79. [Google Scholar] [CrossRef]
- Karnovsky, M.J.; Kleinfeld, A.M.; Hoover, R.L.; Klausner, R.D. The concept of lipid domains in membranes. J. Cell. Biol. 1982, 94, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Baumgart, T.; Hess, S.T.; Webb, W.W. Imaging coexisting fluid domains in biomembrane models coupling curvature and line tension. Nature 2003, 425, 821–824. [Google Scholar] [CrossRef] [PubMed]
- Inaoka, Y.; Yamazaki, M. Vesicle fission of giant unilamellar vesicles of liquid-ordered-phase membranes induced by amphiphiles with a single long hydrocarbon chain. Langmuir 2007, 23, 720–728. [Google Scholar] [CrossRef] [PubMed]
- Deamer, D.W.; Dworkin, J.P. Chemistry and physics of primitive membranes. Top. Curr. Chem. 2005, 259, 1–27. [Google Scholar]
- Jakobsen, U.; Simonsen, A.C.; Vogel, S. DNA-controlled assembly of soft nanoparticles. J. Am. Chem. Soc 2008, 130, 10462–10463. [Google Scholar] [CrossRef] [PubMed]
- Deamer, D.W.; Gutknecht, J. Proton permeation through model membranes. Methods Enzymol. 1986, 127, 471–480. [Google Scholar] [PubMed]
- Deamer, D.W. Proton permeation of lipid bilayers. J. Bioenerg. Biomembr. 1987, 19, 457–479. [Google Scholar] [PubMed]
- Paula, S.; Deamer, D.W. Permeation of chloride, bromide, and iodide across phospholipid bilayers. Biophys. J. 1997, 72, Th341–Th341. [Google Scholar]
- Paula, S.; Deamer, D.W. Membrane permeability barriers to ionic and polar solutes. Membr. Permeabil. 1999, 48, 77–95. [Google Scholar]
- Chakrabarti, A.C.; Deamer, D.W. Permeation of membranes by the neutral form of amino-acids and peptides - relevance to the origin of peptide translocation. J. Mol. Evol. 1994, 39, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.C.; Deamer, D.W. Permeability of lipid bilayers to amino-acids and phosphate. Biochim. Biophys. Acta 1992, 1111, 171–177. [Google Scholar] [CrossRef]
- Harang, E.A.; Deamer, D.W. Amino-acid permeability across liposome bilayers. Biophys. J. 1990, 57, A487–A487. [Google Scholar]
- Walde, P.; Goto, A.; Monnard, P.A.; Wessicken, M.; Luisi, P.L. Oparins reactions revisited-enzymatic-synthesis of poly(adenylic acid) in MICELLES and self-reproducing vesicles. J. Am. Chem. Soc. 1994, 116, 7541–7547. [Google Scholar] [CrossRef]
- Monnard, P.A.; Deamer, D.W. Nutrient uptake by protocells: A liposome model system. Orig. Life Evol. Biosphere 2001, 31, 147–155. [Google Scholar] [CrossRef]
- Monnard, P.A.; Oberholzer, T.; Luisi, P. Entrapment of nucleic acids in liposomes. Biochim. Biophys. Acta 1997, 1329, 39–50. [Google Scholar] [CrossRef]
- Mansy, S.S.; Schrum, J.P.; Krishnamurthy, M.; Tobe, S.; Treco, D.A.; Szostak, J.W. Template-directed synthesis of a genetic polymer in a model protocell. Nature 2008, 454, 122–125. [Google Scholar] [CrossRef] [PubMed]
- Sacerdote, M.G.; Szostak, J.W. Semipermeable lipid bilayers exhibit diastereoselectivity favoring ribose. Proc. Natl. Acad. Sci. USA 2005, 102, 6004–6008. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.Y.; Pohorille, A. Permeation of membranes by ribose and its diastereomers. J. Am. Chem. Soc. 2009, 131, 10237–10245. [Google Scholar] [CrossRef] [PubMed]
- Escabi-Perez, J.R.; Romero, A.; Lukac, S.; Fendler, J.H. Aspects of artificial photosynthesis. Photoionization and electron transfer in dihexadecylphosphate vesicles. J. Am. Chem. Soc. 1979, 101, 2231–2233. [Google Scholar] [CrossRef]
- Deamer, D.W. Polycyclic aromatic hydrocarbons: primitive pigment systems in the prebiotic environment. Adv. Space Res. 1992, 12, 183–189. [Google Scholar] [CrossRef]
- Cape, J.L.; Monnard, P.A.; Boncella, J.M. Prebiotically relevant mixed fatty acid vesicles support anionic solute encapsulation and photochemically catalyzed trans-membrane charge transport. Chem. Sci. 2011. [Google Scholar] [CrossRef]
- Namani, T.; Deamer, D.W. Stability of model membranes in extreme environments. Orig. Life Evol. Biosphere 2008, 38, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Sternberg, B.; Sorgi, F.L.; Huang, L. New structures in complex formation between DNA and cationic liposomes visualized by freeze-fracture electron microscopy. FEBS Lett. 1994, 356, 361–366. [Google Scholar] [CrossRef]
- Luisi, P.L.; de Souza, T.P.; Stano, P. Vesicle behavior: In search of explanations. J. Phys. Chem. B 2008, 112, 14655–14664. [Google Scholar] [CrossRef] [PubMed]
- Berti, D.; Luisi, P.L.; Baglioni, P. Molecular recognition in supramolecular structures formed by phosphatidylnucleosides-based amphiphiles. Colloid Surface A 2000, 167, 95–103. [Google Scholar] [CrossRef]
- Hadorn, M.; Eggenberger Hotz, P. DNA-mediated self-assembly of artificial vesicles. PLoS ONE 2010, 5, e9886. [Google Scholar] [CrossRef] [PubMed]
- Caschera, F.; Stano, P.; Luisi, P.L. Reactivity and fusion between cationic vesicles and fatty acid anionic vesicles. J. Coll. Interface Sci. 2010, 345, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Walde, P.; Ichikawa, S. Review. Enzyme inside lipid vesicles: preparation, reactivity and applications. Biomol. Eng. 2001, 18, 143–177. [Google Scholar] [CrossRef]
- Monnard, P.A.; DeClue, M.S.; Ziock, H.J. Organic nano-compartments as biomimetic reactors and protocells. Curr. Nanosci. 2008, 4, 71–87. [Google Scholar] [CrossRef]
- Pereira de Souza, T.; Stano, P.; Luisi, P.L. The minimal size of liposome-based model cells brings about a remarkably enhanced entrapment and protein synthesis. ChemBioChem 2009, 10, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Kuruma, Y.; Stano, P.; Ueda, T.; Luisi, P.L. A synthetic biology approach to the construction of membrane proteins in semi-synthetic minimal cells. Biochim. Biophys. Acta 2009, 1788, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Tawfik, D.S.; Griffiths, A.D. Man-made cell-like compartments for molecular evolution. Nat. Biotechnol. 1998, 16, 652–656. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, K.; Sato, K.; Shima, Y.; Urabe, I.; Yomo, T. Expression of a cascading genetic network within liposomes. FEBS Lett. 2004, 576, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Noireaux, V.; Bar-Ziv, R.; Godefroy, J.; Salman, H.; Libchaber, A. Toward an artificial cell based on gene expression in vesicles. Phys. Biol. 2005, 2, P1–P8. [Google Scholar] [CrossRef] [PubMed]
- Noireaux, V.; Libchaber, A. A vesicle bioreactor as a step toward an artificial cell assembly. Proc. Natl. Acad. Sci. USA 2004, 101, 17669–17674. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Noireaux, V. Study of messenger RNA inactivation and protein degradation in an Escherichia coli cell-free expression system. J. Biol. Eng. 2010, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.; Chen, L.H.; Deamer, D.; Krakauer, D.C.; Packard, N.H.; Stadler, P.F.; Bedau, M.A. Transitions from nonliving to living matter. Science 2004, 303, 963–965. [Google Scholar] [CrossRef] [PubMed]
- Giese, B. Electron transfer in DNA. Curr. Opin. Chem. Biol. 2002, 6, 612–618. [Google Scholar] [CrossRef]
- DeClue, M.S.; Monnard, P.A.; Bailey, J.A.; Maurer, S.E.; Collis, G.E.; Ziock, H.J.; Rasmussen, S.; Boncella, J.M. Nucleobase mediated, photocatalytic vesicle formation from an ester precursor. J. Am. Chem. Soc. 2009, 131, 931–933. [Google Scholar] [CrossRef] [PubMed]
- Maurer, S.E.; DeClue, M.S.; Albertsen, A.N.; Dörr, M.; Kuiper, D.S.; Ziock, H.; Rasmussen, S.; Boncella, J.M.; Monnard, P.-A. Interactions between catalyst and amphiphilic structures and their implications for a protocell model. ChemPhysChem 2011, in press. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, Y.; Hatfield, S.; Wan, R.; Zhu, Q.; Li, X.; McMills, M.; Ma, Y.; Li, J.; Brown, K.L.; He, C.; Liu, F.; Chen, X. Dipeptide seryl-histidine and related oligopeptides cleave DNA, protein, and a carboxyl ester. Bioorg. Med. Chem. 2000, 8, 2675–2680. [Google Scholar] [CrossRef]
- Liu, R.; Orgel, L.E. Polymerization of beta-amino acids in aqueous solution. Orig. Life Evol. Biosphere 1998, 28, 47–60. [Google Scholar] [CrossRef]
- Hill, A.R., Jr.; Orgel, L.E. Oligomerization of negatively-charged amino acids by carbonyldiimidazole. Orig. Life Evol. Biosphere 1996, 26, 539–545. [Google Scholar] [CrossRef]
- Ehler, K.W.; Orgel, L.E. N,N′-carbonyldiimidazole-induced peptide formation in aqueous solution. Biochim. Biophys. Acta 1976, 434, 233–243. [Google Scholar] [CrossRef]
- Brack, A. Selective emergence and survival of early polypeptides in water. Orig. Life Evol. Biosphere 1987, 17, 367–379. [Google Scholar] [CrossRef]
- Blocher, M.; Liu, D.J.; Walde, P.; Luisi, P.L. Liposome-assisted selective polycondensation of alpha-amino acids and peptides. Macromolecules 1999, 32, 7332–7334. [Google Scholar] [CrossRef]
- Blocher, M.; Liu, D.J.; Luisi, P.L. Liposome-assisted selective polycondensation of alpha-amino acids and peptides: The case of charged liposomes. Macromolecules 2000, 33, 5787–5796. [Google Scholar] [CrossRef]
- Gilbert, W. Origin of life - the Rna world. Nature 1986, 319, 618. [Google Scholar] [CrossRef]
- Tohidi, M.; Zielinski, W.S.; Chen, C.H.; Orgel, L.E. Oligomerization of 3′-amino-3′deoxyguanosine-5′phosphorimidazolidate on a d(CpCpCpCpC) template. J. Mol. Evol. 1987, 25, 97–99. [Google Scholar] [CrossRef] [PubMed]
- Kanavarioti, A. Dimerization in highly concentrated solutions of phospho-imidazolide activated mononucleotides. Orig. Life Evol. Biosphere 1997, 27, 357–376. [Google Scholar] [CrossRef]
- Ferris, J.P. Montmorillonite catalysis of 30–50 mer oligonucleotides: Laboratory demonstration of potential steps in the origin of the RNA world. Orig. Life Evol. Biosphere 2002, 32, 311–332. [Google Scholar] [CrossRef]
- Rajamani, S.; Vlassov, A.; Benner, S.; Coombs, A.; Olasagasti, F.; Deamer, D. Lipid-assisted synthesis of RNA-like polymers from mononucleotides. Orig. Life Evol. Biosphere 2008, 38, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Vriezema, D.M.; Aragones, M.C.; Elemans, J.A.A.W.; Cornelissen, J.J.L.M.; Rowan, A.E.; Nolte, R.J.M. Self-assebled nanoreactors. Chem. Rev. 2005, 105, 1445–1489. [Google Scholar] [CrossRef] [PubMed]
- Walde, P.; Wick, R.; Fresta, M.; Mangone, A.; Luisi, P.L. Autopoietic self-reproduction of fatty-acid vesicles. J. Am. Chem. Soc. 1994, 116, 11649–11654. [Google Scholar] [CrossRef]
- Steinberg-Yfrach, G.; Liddell, P.A.; Hung, S.-C.; Moore, A.L.; Gust, D.; Moore, T.A. Conversion of light energy to proton potential in liposomes by artificial photosythetic reaction centres. Nature 1997, 389, 239–241. [Google Scholar] [CrossRef]
- Balazs, A.C.; Epstein, I.R. Chemistry, emergent or just complex? Science 2009, 325, 1632–1634. [Google Scholar] [CrossRef] [PubMed]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Maurer, S.E.; Monnard, P.-A. Primitive Membrane Formation, Characteristics and Roles in the Emergent Properties of a Protocell. Entropy 2011, 13, 466-484. https://doi.org/10.3390/e13020466
Maurer SE, Monnard P-A. Primitive Membrane Formation, Characteristics and Roles in the Emergent Properties of a Protocell. Entropy. 2011; 13(2):466-484. https://doi.org/10.3390/e13020466
Chicago/Turabian StyleMaurer, Sarah Elizabeth, and Pierre-Alain Monnard. 2011. "Primitive Membrane Formation, Characteristics and Roles in the Emergent Properties of a Protocell" Entropy 13, no. 2: 466-484. https://doi.org/10.3390/e13020466
APA StyleMaurer, S. E., & Monnard, P.-A. (2011). Primitive Membrane Formation, Characteristics and Roles in the Emergent Properties of a Protocell. Entropy, 13(2), 466-484. https://doi.org/10.3390/e13020466