
Production of Plant Beneficial and Antioxidants Metabolites by Klebsiellavariicola under Salinity Stress

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
:
1. Introduction
2. Results
2.1. Production of Plant Growth-Promoting Traits
2.1.1. Screening for Phytase Production
2.1.2. Screening for Nitrogen Fixation, Production of Ammonia and Siderophore
2.2. Production of Salinity Ameliorating Traits
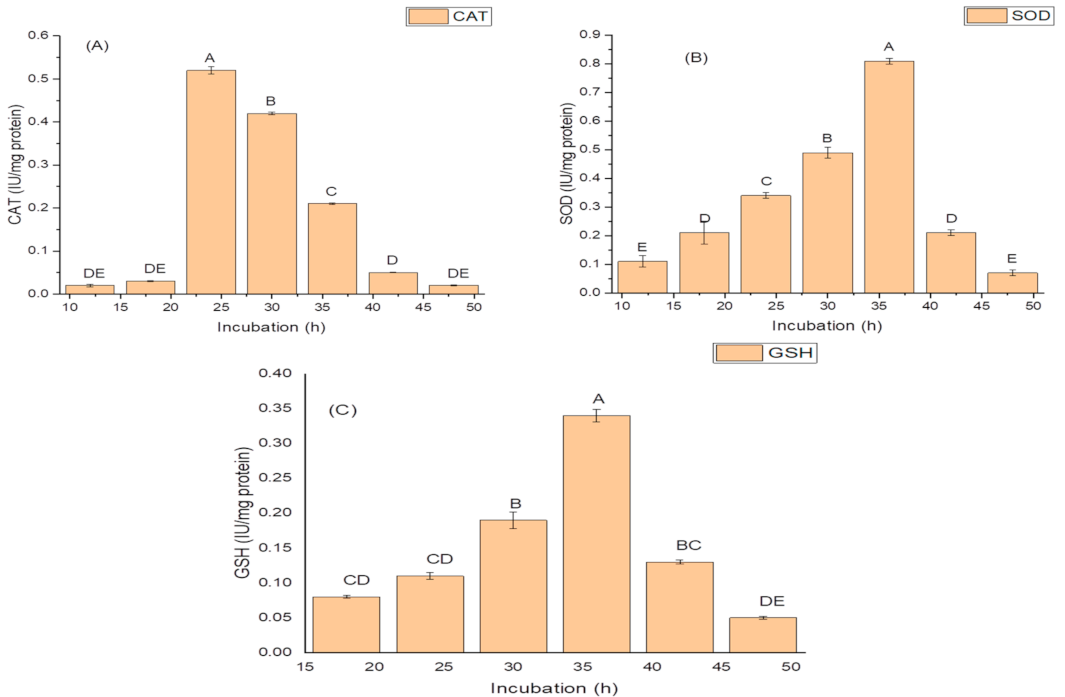
2.3. Screening for Antioxidant Enzymes
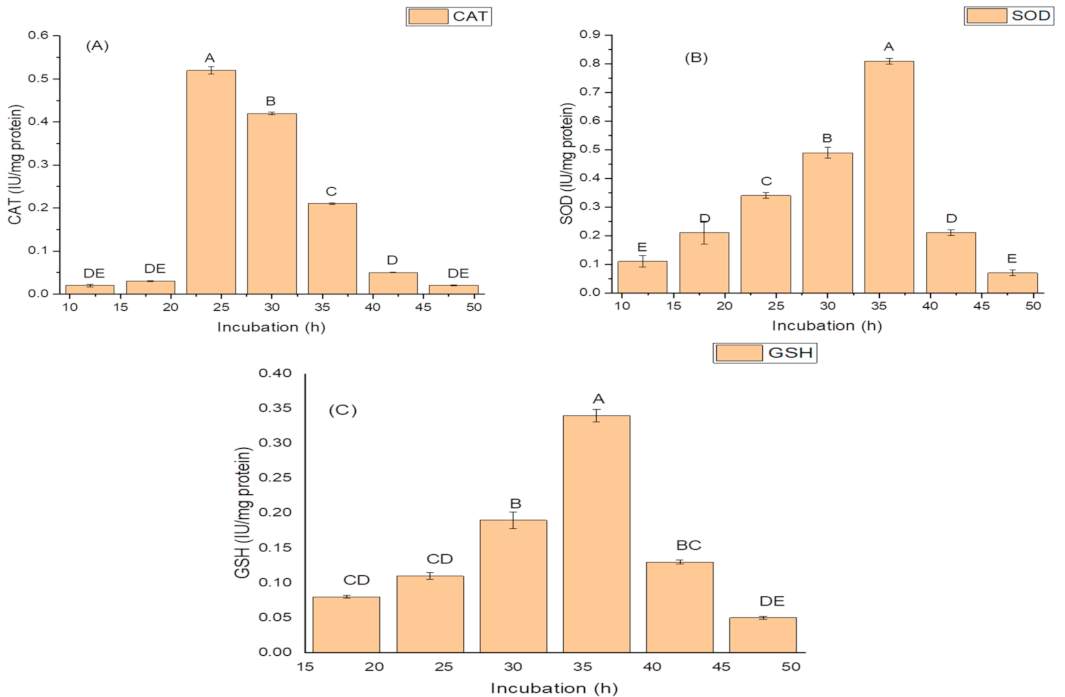
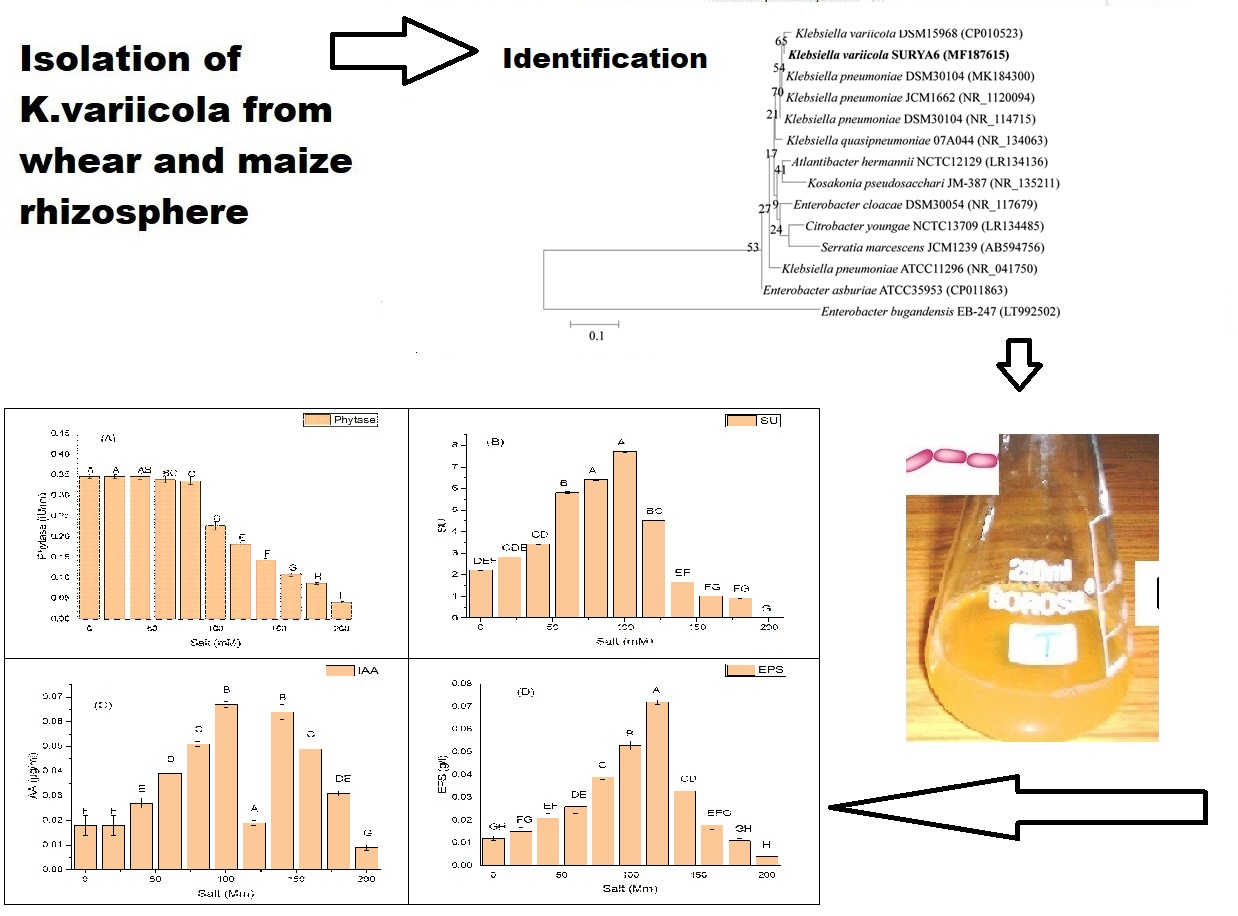
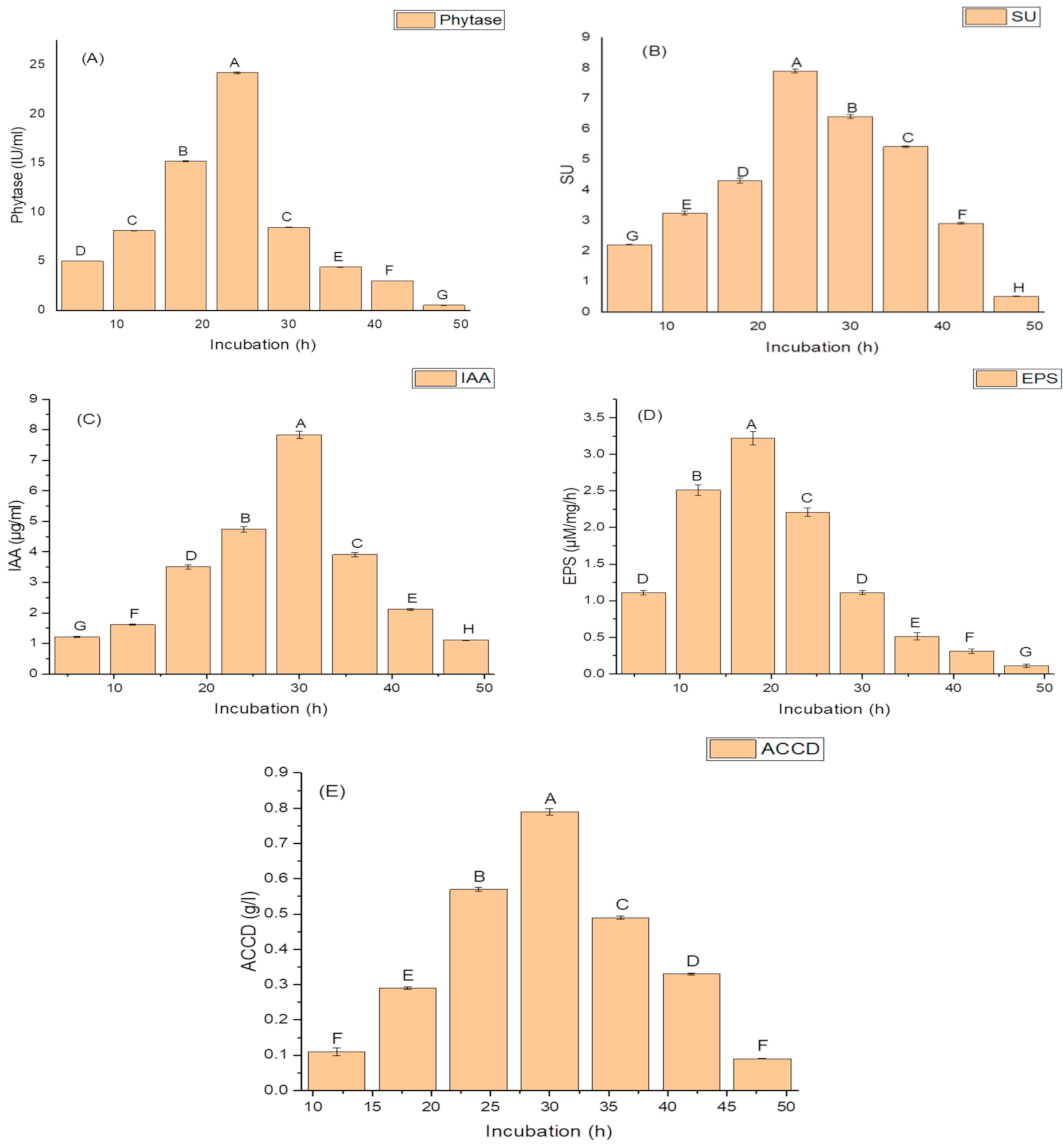
2.4. Effect of Incubation Period on Growth, and Production of PGP and Antioxidant Traits
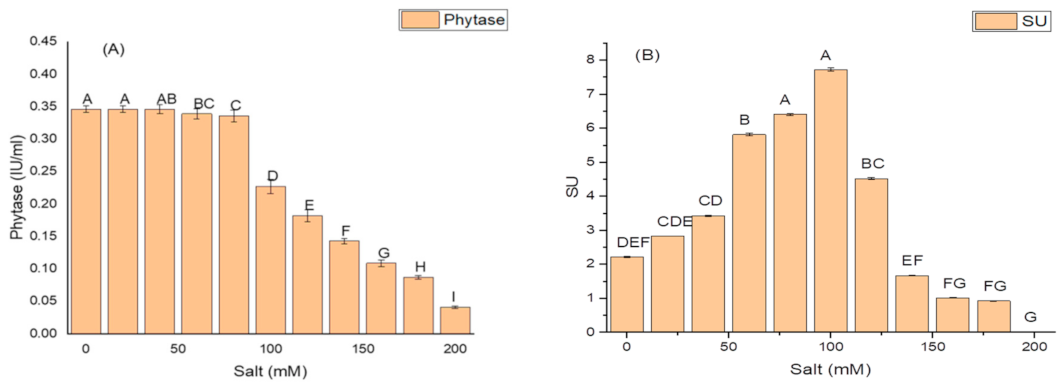
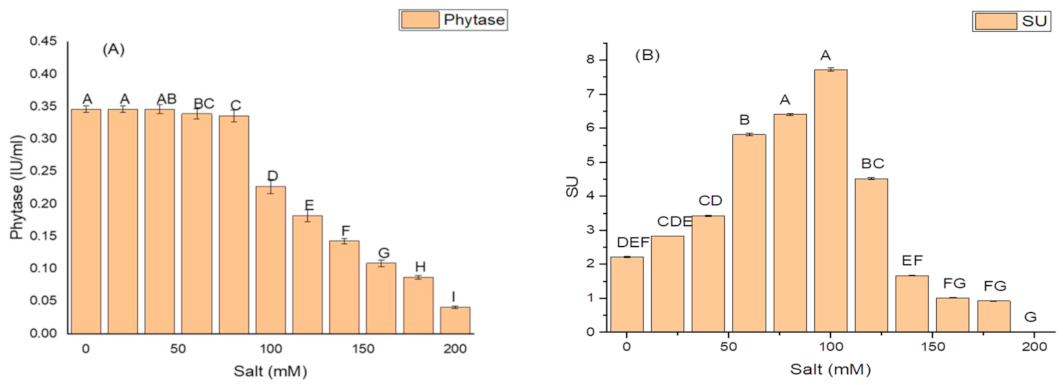
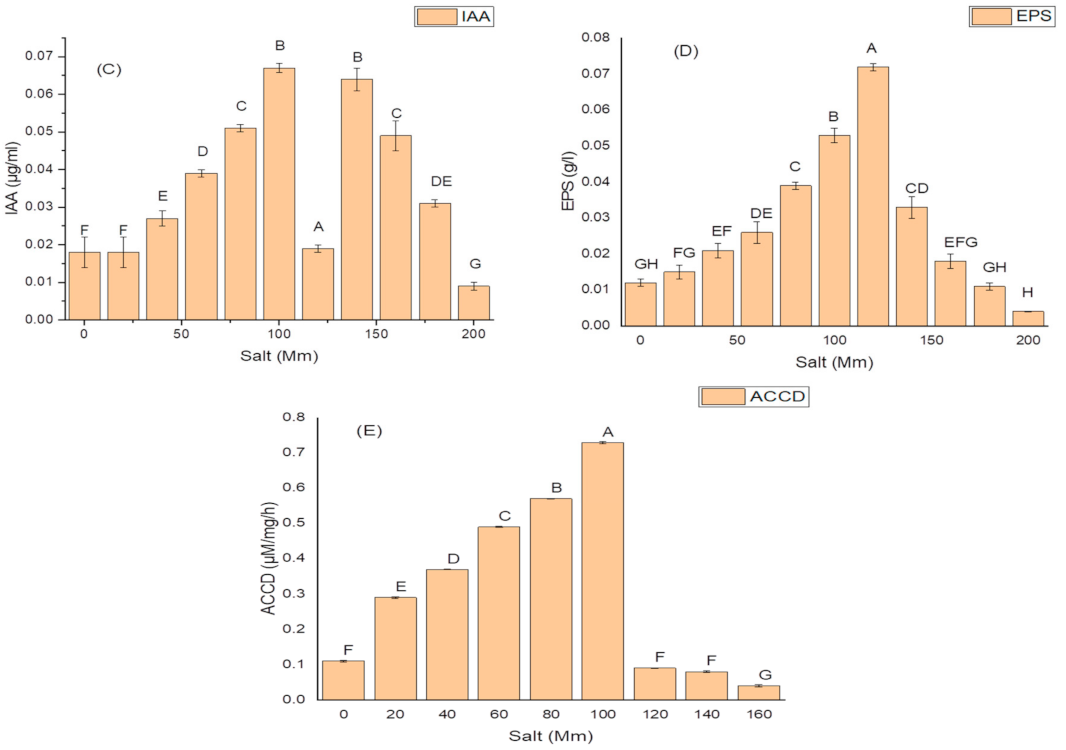
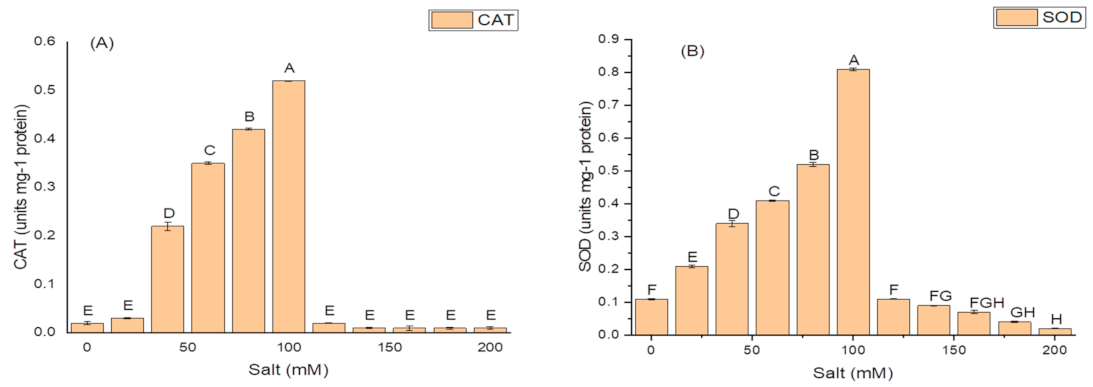
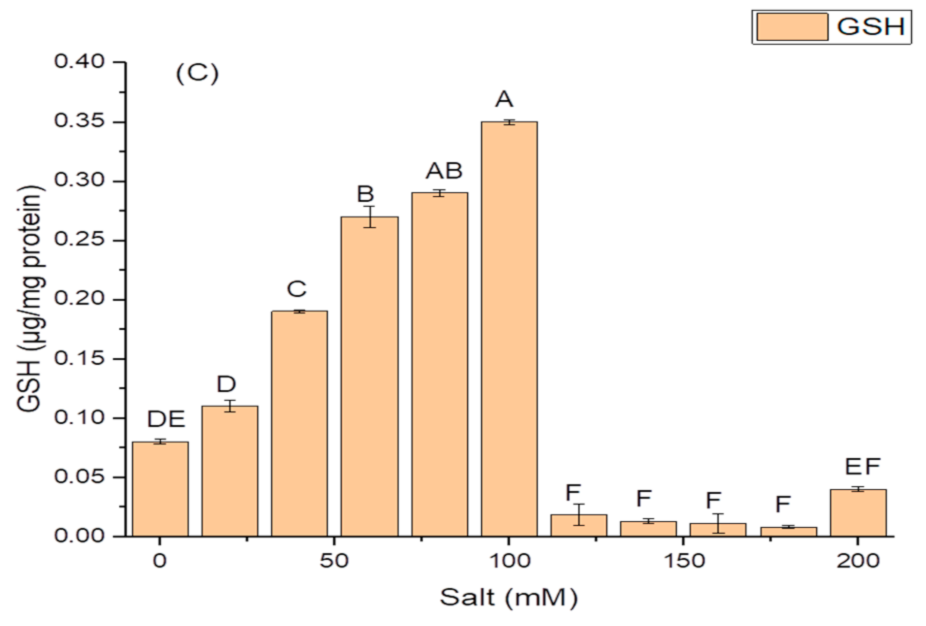
2.5. Effect of Salt Concentration on Growth and Production of PGP Taits and Antioxidant Enzymes in K. variicola SURYA6
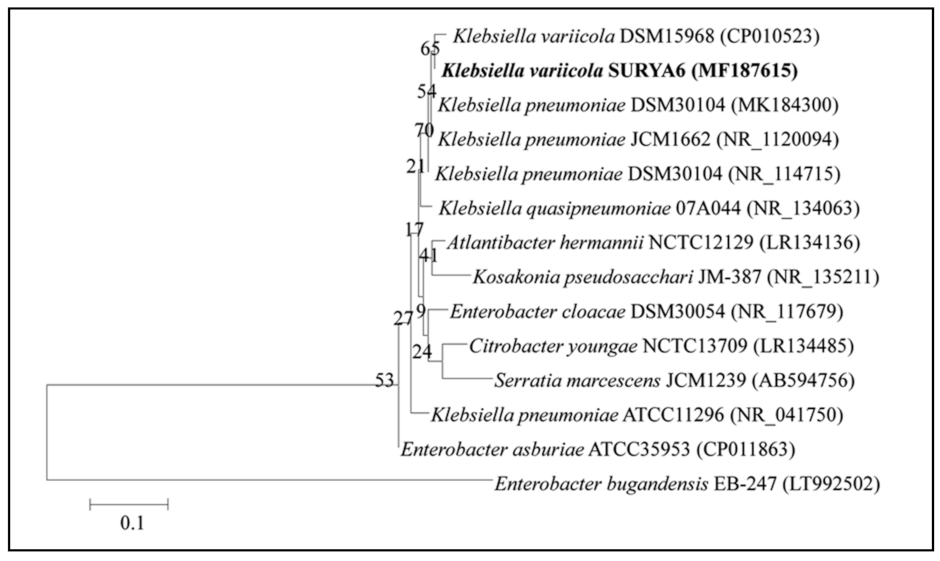
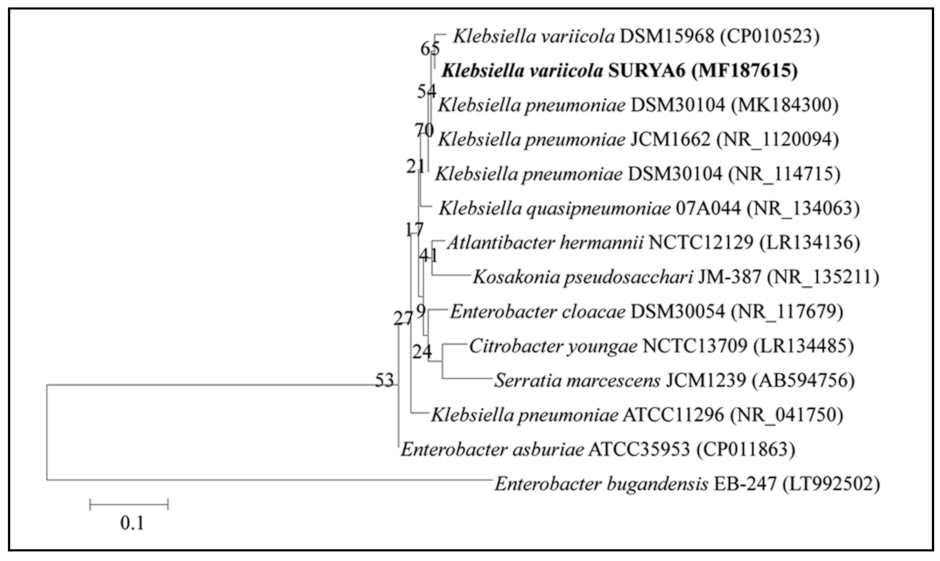
2.6. Identification of the Potent Isolate-Ribotyping
3. Discussion
4. Material and Methods
4.1. Soil Samples
4.2. Screening for Multifarious PGPR Isolate
4.2.1. Screening for Phytase Production
4.2.2. Phosphate Solubilization and Organic Acid Production
4.2.3. Screening for Nitrogen Fixation and Production of Ammonia, Siderophore and Indole Acetic Acid (IAA)
4.3. Screening for Salinity Ameliorating Traits
4.3.1. Production of ACCD and EPS
4.3.2. Screening for Production of Antioxidant Enzymes
4.4. Selection of Potent PGPR Isolate
4.5. Growth Kinetics and Production of PGP and Salinity Ameliorating Traits in Potent PGPR
4.6. The Effect of Salt Stress on the Production of PGP and Salinity Ameliorating Traits
4.7. Identification of the Potent Isolate-Ribotyping
4.8. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Shaikh, S.; Sayyed, R. Role of plant growth-promoting rhizobacteria and their formulation in biocontrol of plant diseases. In Plant Microbes Symbiosis: Applied Facets; Springer: Singapore, 2015; pp. 337–351. [Google Scholar]
- Fazeli-Nasab, B.; Sayyed, R. Plant Growth-Promoting Rhizobacteria and Salinity Stress: A Journey into the Soil. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management; Springer: Singapore, 2019; pp. 21–34. [Google Scholar]
- Lubna; Asaf, S.; Hamayun, M.; Gul, H.; Lee, I.J.; Hussain, A. Aspergillus niger CSR3 regulates plant endogenous hormones and secondary metabolites by producing gibberellins and indoleacetic acid. J. Plant Interact. 2018, 13, 100–111. [Google Scholar]
- Shaikh, S.S.; Sayyed, R.Z.; Reddy, M.S. Plant growth-promoting rhizobacteria: An eco-friendly approach for sustainable agroecosystem. In Plant, Soil and Microbes; Springer: Singapore, 2016; pp. 181–201. [Google Scholar]
- Khan, A.; Sayyed, R.; Seifi, S. Rhizobacteria: Legendary Soil Guards in Abiotic Stress Management. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management; Reddy, M.S., Sayyed, R.Z., Arora, N.K., Eds.; Springer: Singapore, 2019; pp. 327–343. [Google Scholar]
- Jabborova, D.; Wirth, S.; Kannepalli, A.; Narimanov, A.; Desouky, S.; Davranov, K.; Sayyed, R.; El Enshasy, H.; Malek, R.A.; Syed, A. Co-Inoculation of rhizobacteria and biochar application improves growth and nutrients in soybean and enriches soil nutrients and enzymes. Agronomy 2020, 10, 1142. [Google Scholar] [CrossRef]
- Shaikh, S.; Patel, P.; Patel, S.; Nikam, S.; Rane, T.; Sayyed, R. Production of biocontrol traits by banana field fluorescent Pseudomonads and comparison with chemical fungicide. Indian J. Exp. Biol. 2014, 52, 917–920. [Google Scholar] [PubMed]
- Sayyed, R.; Seifi, S.; Patel, P.; Shaikh, S.; Jadhav, H.; El Enshasy, H. Siderophore production in groundnut rhizosphere isolate, Achromobacter sp. RZS2 influenced by physicochemical factors and metal ions. Env. Sustain. 2019, 2, 117–124. [Google Scholar] [CrossRef]
- Pandya, N.; Desai, P.; Jadhav, H.; Sayyed, R. Plant growth-promoting potential of Aspergillus sp. NPF7, isolated from wheat rhizosphere in South Gujarat, India. Environ. Sustain. 2018, 1, 245–252. [Google Scholar] [CrossRef]
- Jabborova, D.; Annapurna, K.; Fayzullaeva, M.; Sulaymonov, K.; Kadirova, D.; Jabbarov, Z.; Sayyed, R. Isolation and characterization of endophytic bacteria from ginger (Zingiber officinale Rosc.). Ann. Phytomed. 2020, 9, 116–121. [Google Scholar] [CrossRef]
- Shaikh, S.; Wani, S.; Sayyed, R.; Thakur, R.; Gulati, A. Production, purification and kinetics of chitinase of Stenotrophomonas maltophilia isolated from rhizospheric soil. Indian J. Exp. Biol. 2018, 56, 274–278. [Google Scholar]
- Khan, I.; Awan, S.A.; Ikram, R.; Rizwan, M.; Akhtar, N.; Yasmin, H.; Sayyed, R.Z.; Ali, S.; Ilyas, N. Effect of 24-Epibrassinolide on plant growth, antioxidants defense system and endogenous hormones in two wheat varieties under drought stress. Physiol. Plant. 2020, 1–11. [Google Scholar] [CrossRef]
- Sayyed, R.; Jamadar, D.; Patel, P. Production of exo-polysaccharide by Rhizobium sp. Ind. J. Microbiol. 2011, 51, 294–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayyed, R.; Patel, P.; Shaikh, S. Plant growth promotion and root colonization by EPS producing Enterobacter sp. RZS5 under heavy metal contaminated soil. Indian J. Exp. Biol. 2015, 53, 116–123. [Google Scholar] [PubMed]
- Sagar, A.; Shukla, P.; Sayyed, R.; Ramteke, P. Stimulation of Seed Germination and Growth Parameters of Rice var. Sahbhagi by Enterobacter cloacae in the presence of ammonium sulfate as a substitute of ACC. In Plant Growth Promoting Rhizobacteria (PGPR): Prospects for Sustainable Agriculture; Springer: Singapore, 2019; pp. 117–124. [Google Scholar]
- Sagar, A.; Riyazuddin, R.; Shukla, P.; Ramteke, P.; Sayyed, R. Heavy metal stress tolerance in Enterobacter sp. PR14 is mediated by plasmid. Indian J. Exp. Biol. 2020, 58, 115–121. [Google Scholar]
- Sagar, A.; Sayyed, R.; Ramteke, P.; Sharma, S.; Marraiki, N.; Elgorban, A.M.; Syed, A. ACC deaminase and antioxidant enzymes producing halophilic Enterobacter sp. PR14 promotes the growth of rice and millets under salinity stress. Physiol. Mol. Biol. Plants 2020, 26, 1847–1854. [Google Scholar] [CrossRef]
- Ilyas, N.; Mumtaz, K.; Akhtar, N.; Yasmin, H.; Sayyed, R.; Khan, W.; Enshasy, H.A.E.; Dailin, D.J.; Elsayed, E.A.; Ali, Z. Exopolysaccharides Producing Bacteria for the Amelioration of Drought Stress in Wheat. Sustainability 2020, 12, 8876. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2013, 2, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.B.; Sayyed, R.Z.; Sonawane, M.; Trivedi, M.H.; Thivakaran, G.A. Neurospora sp. SR8, a novel phosphate solubilizer from rhizosphere soil of Sorghum in Kachchh, Gujarat, India. Indian J. Exp. Biol. 2016, 54, 644–649. [Google Scholar] [PubMed]
- Suriani, N.L.; Suprapta, D.N.; Nazir, N.; Parwanayoni, N.M.S.; Darmadi, A.A.K.; Dewi, D.A.; Sudatri, N.W.; Fudholi, A.; Sayyed, R.; Syed, A. A Mixture of Piper Leaves Extracts and Rhizobacteria for Sustainable Plant Growth Promotion and Bio-Control of Blast Pathogen of Organic Bali Rice. Sustainability 2020, 12, 8490. [Google Scholar] [CrossRef]
- Dailin, D.J.; Hanapi, S.Z.; Elsayed, E.A.; Sukmawati, D.; Azelee, N.I.W.; Eyahmalay, J.; Siwapiragam, V.; El Enshasy, H. Fungal Phytases: Biotechnological applications in food and feed industries. In Recent Advancements in White Biotechnology through Fungi; Springer: Cham, Switzerland, 2019; pp. 65–99. [Google Scholar]
- El Enshasy, H.; Dailin, D.J.; Abd Manas, N.H.; Azlee, N.I.W.; Eyahmalay, J.; Yahaya, S.A.; Abd Malek, R.; Siwapiragam, V.; Siwapiragam, D. Current and future applications of phytases in poultry industry: A critical review. J. Adv. Vet. Bio Sci. Techniq. 2018, 3, 65–74. [Google Scholar]
- Barrios-Camacho, H.; Aguilar-Vera, A.; Beltran-Rojel, M.; Baguilar-Vera, E.; Duran-Bedolla, J.; Rodriguez-Medina, N.; Lozano-Aguirre, L.; Perez-Carrascal, O.M.; Rojas, J.; Garza-Ramos, U. Molecular epidemiology of Klebsiella variicola obtained from different sources. Sci. Rep. 2019, 9, 1–10. [Google Scholar]
- Balaban, N.P.; Suleimanova, A.D.; Valeeva, L.R.; Chastukhina, I.B.; Rudakova, N.L.; Sharipova, M.R.; Shakirov, E.V. Microbial phytases and phytate: Exploring opportunities for sustainable phosphorus management in agriculture. Amer. J. Mol. Biol. 2016, 7, 11–29. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Singh, P.; Jorquera, M.A.; Sangwan, P.; Kumar, P.; Verma, A.K.; Agrawal, S. Isolation of phytase-producing bacteria from Himalayan soils and their effect on growth and phosphorus uptake of Indian mustard (Brassica juncea). World. J. Microbiol. Biotechnol. 2013, 29, 1361–1369. [Google Scholar] [CrossRef]
- Singh, N.K.; Joshi, D.K.; Gupta, R.K. Isolation of phytase producing bacteria and optimization of phytase production parameters. Jund. J. Microbiol. 2013, 6. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Li, A.; Chen, J.; Su, Y.; Li, Y.; Ma, S. Isolation of a phytase-producing bacterial strain from agricultural soil and its characterization and application as an effective eco-friendly phosphate solubilizing bioinoculant. Comm. Soil Sci. Plant Anal. 2018, 49, 984–994. [Google Scholar] [CrossRef]
- Rosenblueth, M.; Martínez, L.; Silva, J.; Martínez-Romero, E. Klebsiella variicola, a novel species with clinical and plant-associated isolates. System. Appl. Microbiol. 2004, 27, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, G.; Shah, R.; Joshi, B.; Patel, P. Klebsiella pneumoniae VRE36 as a PGPR isolated from Saccharum officinarum cultivar Co99004. J. Appl. Biol. Biotech. 2017, 5, 047–052. [Google Scholar] [CrossRef] [Green Version]
- Pavlova, A.; Leontieva, M.; Smirnova, T.; Kolomeitseva, G.; Netrusov, A.; Tsavkelova, E. Colonization strategy of the endophytic plant growth-promoting strains of Pseudomonas fluorescens and Klebsiella oxytoca on the seeds, seedlings, and roots of the epiphytic orchid, Dendrobium nobile Lindl. J. Appl. Microbiol. 2017, 123, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Mazumdar, D.; Saha, S.P.; Ghosh, S. Klebsiella pneumoniae rs26 as a potent PGPR isolated from chickpea (Cicer arietinum) rhizosphere. Pharma. Innov. J. 2018, 7, 56–62. [Google Scholar]
- Sharma, S.; Trivedi, M.; Sayyed, R.; Thivakaran, G. Status of soil phosphorus in context with phosphate solubilizing microorganisms in different agricultural amendments in Kachchh, Gujarat, Western India. Ann. Res. Rev. Biol. 2014, 2901–2909. [Google Scholar] [CrossRef]
- Panda, P.; Choudhury, A.; Chakraborty, S.; Ray, D.P.; Deb, S.; Patra, P.S.; Mahato, B.; Paramanik, B.; Singh, A.K.; Chauhan, R.K. Phosphorus Solubilizing Bacteria from Tea Soils and their Phosphate Solubilizing Abilities. Int. J. Biores. Sci. 2017, 4, 113–125. [Google Scholar] [CrossRef]
- Ahemad, M.; Khan, M.S. Effects of insecticides on plant-growth-promoting activities of phosphate solubilizing rhizobacterium Klebsiella sp. strain PS19. Pesti. Biochem. Physiol. 2011, 100, 51–56. [Google Scholar] [CrossRef]
- Kuan, K.B.; Othman, R.; Abdul Rahim, K.; Shamsuddin, Z.H. Plant growth-promoting rhizobacteria inoculation to enhance vegetative growth, nitrogen fixation, and nitrogen remobilization of maize under greenhouse conditions. PLoS ONE 2016, 11, e0152478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachdev, D.P.; Chaudhari, H.G.; Kasture, V.M.; Dhavale, D.D.; Chopade, B.A. Isolation and characterization of indole acetic acid (IAA) producing Klebsiella pneumoniae strains from the rhizosphere of wheat (Triticum aestivum) and their effect on plant growth. Ind. J. Exp. Biol. 2009, 47, 993–1000. [Google Scholar]
- Govindarajan, M.; Kwon, S.-W.; Weon, H.-Y. Isolation, molecular characterization, and growth-promoting activities of endophytic sugarcane diazotroph Klebsiella sp. GR9. World J. Microbiol. Biotechnol. 2007, 23, 997–1006. [Google Scholar] [CrossRef]
- Haahtela, K.; Ronkko, R.; Laakso, T.; Williams, P.H.; Korhonen, T.K. Root-associated Enterobacter and Klebsiella in Poa pratensis: Characterization of an iron-scavenging system and a substance stimulating root hair production. Mol. Plant-Microbe. Interact. 1990, 3, 358–365. [Google Scholar] [CrossRef]
- Moghannem, S.A.; Farag, M.M.; Shehab, A.; Azab, M.S. Media optimization for exopolysaccharide producing Klebsiella oxytoca KY498625 under varying cultural conditions. Int. J. Adv. Res. Biol. Sci. 2017, 4, 16–30. [Google Scholar] [CrossRef]
- AMAO, J.A.; Omojasola, P.F.; Barooah, M. Isolation, and characterization of some exopolysaccharide producing bacteria from cassava peel heaps. Sci. Afr. 2019, 4, e00093. [Google Scholar] [CrossRef]
- Sivakumar, T.; Shankar, T.; Thangapandian, V.; Mahendran, S. Media optimization for exopolysaccharide producing Klebsiella pneumoniae KU215681 under varying cultural conditions. Int. J. Biochem. Biophys. 2016, 4, 16–23. [Google Scholar] [CrossRef]
- Ramírez-Castillo, M.; Uribelarrea, J. Improved process for exopolysaccharide production by Klebsiella pneumoniae sp. pneumoniae by a fed-batch strategy. Biotechnol. Lett. 2004, 26, 1301–1306. [Google Scholar] [PubMed]
- Zhao, M.; Cui, N.; Qu, F.; Huang, X.; Yang, H.; Nie, S.; Zha, X.; Cui, S.W.; Nishinari, K.; Phillips, G.O. Novel nano-particulated exopolysaccharide produced by Klebsiella sp. PHRC1. 001. Carbon Poly. 2017, 171, 252–258. [Google Scholar] [CrossRef]
- Hariyono, D. The effect of planting hole size and manure on vegetative growth of golden teak (Tectona grandis L.). J. Degrad. Min. Lands Manag. 2018, 5, 1293–1297. [Google Scholar] [CrossRef] [Green Version]
- Patel, P.; Shaikh, S.; Sayyed, R. Dynamism of PGPR in bioremediation and plant growth promotion in heavy metal contaminated soil. Ind. J. Exp. Biol. 2016, 54, 286–290. [Google Scholar]
- Dlamini, A.M.; Peiris, P.S.; Bavor, J.H.; Kailasapathy, K. Rheological characteristics of an exopolysaccharide produced by a strain of Klebsiella oxytoca. J. Biosci. Bioeng. 2009, 107, 272–274. [Google Scholar] [CrossRef]
- Ng, W.V.; Kennedy, S.P.; Mahairas, G.G.; Berquist, B.; Pan, M.; Shukla, H.D.; Lasky, S.R.; Baliga, N.S.; Thorsson, V.; Sbrogna, J. Genome sequence of Halobacterium species NRC-1. Proc. Natl. Acad. Sci. USA 2000, 97, 12176–12181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.P.; Jha, P.; Jha, P.N. The plant-growth-promoting bacterium Klebsiella sp. SBP-8 confers induced systemic tolerance in wheat (Triticum aestivum) under salt stress. J. Plant Physiol. 2015, 184, 57–67. [Google Scholar] [CrossRef]
- Arora, M.; Kaushik, A.; Rani, N.; Kaushik, C. Effect of cyanobacterial exopolysaccharides on salt stress alleviation and seed germination. J. Environ. Biol. 2010, 31, 701–704. [Google Scholar] [PubMed]
- Othman, N.Z.; Elsayed, E.A.; Malek, R.A.; Ramli, S.; Masri, H.J.; Sarmidi, M.R.; Aziz, R.; Wadaan, M.A.; El Enshasy, H.A. Aeration rate effect on the growth kinetics, phytase production and plasmid stability of recombinant Escherichia coli BL21(DE3). J. Pure Appl. Microbiol. 2014, 8, 2721–2728. [Google Scholar]
- Reshma, P.; Naik, M.; Aiyaz, M.; Niranjana, S.; Chennappa, G.; Shaikh, S.; Sayyed, R. Induced systemic resistance by 2, 4-diacetylphloroglucinol positive fluorescent Pseudomonas strains against rice sheath blight. Indian J. Exp. Biol. 2018, 56, 207–212. [Google Scholar]
- Ahemad, M.; Khan, M.S. Plant growth-promoting activities of phosphate solubilizing Enterobacter asburiae as influenced by fungicides. Eurasian J. Biosci. 2010, 4, 88–95. [Google Scholar] [CrossRef]
- Shrivastava, U.P.; Kumar, A. Characterization and optimization of 1-aminocyclopropane-1-carboxylate deaminase (ACCD) activity in different rhizospheric PGPR along with Microbacterium sp. strain ECI-12A. Int. J. Appl. Sci. Biotechnol. 2013, 1, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Acuña, J.J.; Campos, M.; de la Luz Mora, M.; Jaisi, D.P.; Jorquera, M.A. ACCD-producing rhizobacteria from an Andean altiplano native plant (Parastrephia quadrangularis) and their potential to alleviate salt stress in wheat seedlings. Appl. Soil Ecol. 2019, 136, 184–190. [Google Scholar] [CrossRef]
- Sapre, S.; Gontia-Mishra, I.; Tiwari, S. Klebsiella sp. confers enhanced tolerance to salinity and plant growth promotion in oat seedlings (Avena sativa). Microbiol. Res. 2018, 206, 25–32. [Google Scholar] [CrossRef]
- Rodríguez-Medina, N.; Barrios-Camacho, H.; Duran-Bedolla, J.; Garza-Ramos, U. Klebsiella variicola: An emerging pathogen in humans. Emerg. Microbes Infect. 2019, 8, 973–988. [Google Scholar] [CrossRef] [Green Version]
- Kalam, S.; Basu, A.; Ahmad, I.; Sayyed, R.; El Enshasy, H.; Dailin, D.J.; Suriani, N. Recent understanding of soil acidobacteria and their ecological significance: A critical review. Front. Microbiol. 2020, 11, 580024. [Google Scholar] [CrossRef] [PubMed]
- Villamizar, G.A.C.; Funkner, K.; Nacke, H.; Foerster, K.; Daniel, R. Functional metagenomics unwraps a new catalytic domain associated to phytase activity: The Metallo-β-lactamase superfamily domain, discovery and functional characterization of novel soil-metagenome-derived phosphatases. mBio 2019, 10, e01918–e01966. [Google Scholar]
- Pikovskaya, R. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya 1948, 17, 362–370. [Google Scholar]
- Fiske, C.H.; Subbarow, Y. The colorimetric determination of phosphorus. J. Biol. Chem. 1925, 66, 375–400. [Google Scholar] [CrossRef]
- Katznelson, H.; Bose, B. Metabolic activity and phosphate-dissolving capability of bacterial isolates from wheat roots, rhizosphere, and non-rhizosphere soil. Can. J. Microbiol. 1959, 5, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.; Khalil, S.; Ayub, N.; Alam, S.; Latif, F. Organic acids production and phosphate solubilization by phosphate solubilizing microorganisms (PSM) under in vitro conditions. Pak. J. Biol. Sci. 2004, 7, 187–196. [Google Scholar] [CrossRef]
- Bremner, J. Determination of nitrogen in soil by the Kjeldahl method. J. Agric. Sci. 1960, 55, 11–33. [Google Scholar] [CrossRef]
- Dutta, J.; Thakur, D. Evaluation of multifarious plant growth-promoting traits, antagonistic potential, and phylogenetic affiliation of rhizobacteria associated with commercial tea plants grown in Darjeeling, India. PLoS ONE 2017, 12, e0182302. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.; Shaikh, S.; Sayyed, R. Modified chrome azurol S method for detection and estimation of siderophores having an affinity for metal ions other than iron. Environ. Sustain. 2018, 1, 81–87. [Google Scholar] [CrossRef]
- Meyer, J.M.; Abdallah, M. The fluorescent pigment of Pseudomonas fluorescens: Biosynthesis, purification, and physicochemical properties. Microbiology 1978, 107, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Payne, S.M. Detection, isolation, and characterization of siderophores. Methods Enzymol. 1994, 35, 329–344. [Google Scholar]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Safronova, V.I.; Stepanok, V.V.; Engqvist, G.L.; Alekseyev, Y.V.; Belimov, A.A. root-associated bacteria containing 1-aminocyclopropane-1-carboxylate deaminase improve growth and nutrient uptake by pea genotypes cultivated in cadmium supplemented soil. Biol. Fert. Soils. 2006, 42, 267–272. [Google Scholar] [CrossRef]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Planta. 2003, 118, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.A.; Anandapandian, K.T.K.; Parthiban, K. Production and characterization of exopolysaccharides (EPS) from biofilm-forming marine bacterium. Braz. Arch. Biol. Technol. 2011, 54, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Marklund, S. Gudrun Marklund. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Euro. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Beers, R.F.; Sizer, I.W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [CrossRef]
- Nürnberg, D.; Danon, A. Determination of reduced and total glutathione content in extremophilic microalgae Galdieria phlegrea. Plant. Cell. Physiol. 2016, 7, e2372. [Google Scholar] [CrossRef] [Green Version]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Bric, J.M.; Bostock, R.M.; Silverstone, S.E. Rapid in situ assay for indoleacetic acid production by bacteria immobilized on a nitrocellulose membrane. Appl. Env. Microbiol. 1991, 57, 535–538. [Google Scholar] [CrossRef] [Green Version]
- Damodaran, T.; Sah, V.; Rai, R.; Sharma, D.; Mishra, V.; Jha, S.; Kannan, R. Isolation of salt-tolerant endophytic and rhizospheric bacteria by natural selection and screening for promising plant growth-promoting rhizobacteria (PGPR) and growth vigor in tomato under sodic environment. Afr. J. Microbiol. Res. 2013, 7, 5082–5089. [Google Scholar]
- Sambrook, J.; Russell, D.W. The Condensed Protocols from Molecular Cloning: A Laboratory MANUAL; Cold Spring Harbor Laboratory Press Cold Spring Harbor: New York, NY, USA, 2001; p. 800. [Google Scholar]
- Pediyar, V.; Adam, K.A.; Badri, N.N.; Patole, M.; Shouche, Y.S. Aeromonas culicicola sp. nov., from the midgut of Culexquinque fasciatus. Int. J. Syst. Evol. Microbiol. 2002, 52, 1723–1728. [Google Scholar]
- Thompson, J.D.; Toby, J.G.; Frederic, P.; François, J.; Desmond, G.H. The CLUSTAL × windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.N.M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, R.E. Continuous distribution: Tests of significance, In Introductory Statistics for Biology, 2nd ed.; Arnold, B.S., Ed.; Cambridge University Press: London, UK, 1979; pp. 18–42. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Isolates | |||
|---|---|---|---|---|
| N6 | H7 | B1 | V8 | |
| Plant Growth-Promoting (PGP) Traits | ||||
| Phytase activity (IU/mL) | 346.67 ± 2.77 | 329.73 ± 1.33 | 261.67 ± 3.41 | 312.71 ± 2.11 |
| P solubilization (µg/mL) | 3229.02 ± 8.52 | 876.04 ± 3.21 | 507.01 ± 4.56 | 513.05 ± 6.78 |
| Organic acid production | +++ | + | + | + |
| Nitrogen fixation | +++ | + | + | + |
| Ammonia production | +++ | + | + | + |
| Siderophore production | 79.0 ± 0.01 | 43.10 ± 0.01 | 52.11 ± 0.01 | 63.12 ± 0.01 |
| IAA production (µg/mL) | 78.45 ± 1.92 | 43.51 ± 3.91 | 5.22 ± 1.40 | 34.41 ± 1.7 |
| Salinity Ameliorating Traits | ||||
| ACCD (µM/mg/h) | 0.910 ± 1.21 | 0.782 ± 1.02 | 0.563 ± 1.01 | 0.312 ± 1.03 |
| EPS production (g/L) | 32.211 ± 1.21 | 8.12 ± 1.71 | 15.22 ± 1.91 | 23.20 ± 2.23 |
| Antioxidant Enzymes | ||||
| SOD (IU/mg protein) | 13.86 ± 1.03 | 10.15 ± 1.04 | 9.06 ± 1.21 | 7.84 ± 3.21 |
| CAT (IU/mg protein) | 0.07 ± 0.02 | 0.04 ± 0.01 | 0.02 ± 0.01 | 0.01 ± 0.06 |
| GSH (µg/mg protein) | 22.12 ± 6.54 | 17.11 ± 4.32 | 12.12 ± 3.32 | 9.13 ± 2.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kusale, S.P.; Attar, Y.C.; Sayyed, R.Z.; Malek, R.A.; Ilyas, N.; Suriani, N.L.; Khan, N.; El Enshasy, H.A. Production of Plant Beneficial and Antioxidants Metabolites by Klebsiellavariicola under Salinity Stress. Molecules 2021, 26, 1894. https://doi.org/10.3390/molecules26071894
Kusale SP, Attar YC, Sayyed RZ, Malek RA, Ilyas N, Suriani NL, Khan N, El Enshasy HA. Production of Plant Beneficial and Antioxidants Metabolites by Klebsiellavariicola under Salinity Stress. Molecules. 2021; 26(7):1894. https://doi.org/10.3390/molecules26071894
Chicago/Turabian StyleKusale, Supriya P., Yasmin C. Attar, R. Z. Sayyed, Roslinda A. Malek, Noshin Ilyas, Ni Luh Suriani, Naeem Khan, and Hesham A. El Enshasy. 2021. "Production of Plant Beneficial and Antioxidants Metabolites by Klebsiellavariicola under Salinity Stress" Molecules 26, no. 7: 1894. https://doi.org/10.3390/molecules26071894
APA StyleKusale, S. P., Attar, Y. C., Sayyed, R. Z., Malek, R. A., Ilyas, N., Suriani, N. L., Khan, N., & El Enshasy, H. A. (2021). Production of Plant Beneficial and Antioxidants Metabolites by Klebsiellavariicola under Salinity Stress. Molecules, 26(7), 1894. https://doi.org/10.3390/molecules26071894