
New Laboratory Protocol to Determine the Oxidative Stress Profile of Human Nasal Epithelial Cells Using Flow Cytometry

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Experimental Section
2.1. Biological Samples
2.2. Reagents
2.3. Determination of Dead and Live Cells
2.4. Reactive Oxygen Species and Reactive Nitrogen Species Assessment
2.5. Plasmatic Membrane Potential Assessment
2.6. Mitochondrial Assessment
2.7. Intracellular Calcium Assessment
2.8. Oxidative Damage to Biomolecules
2.9. Apoptosis Assay
2.10. Positive Control Incubations
2.11. Cytometer Settings and Data Analysis
3. Results
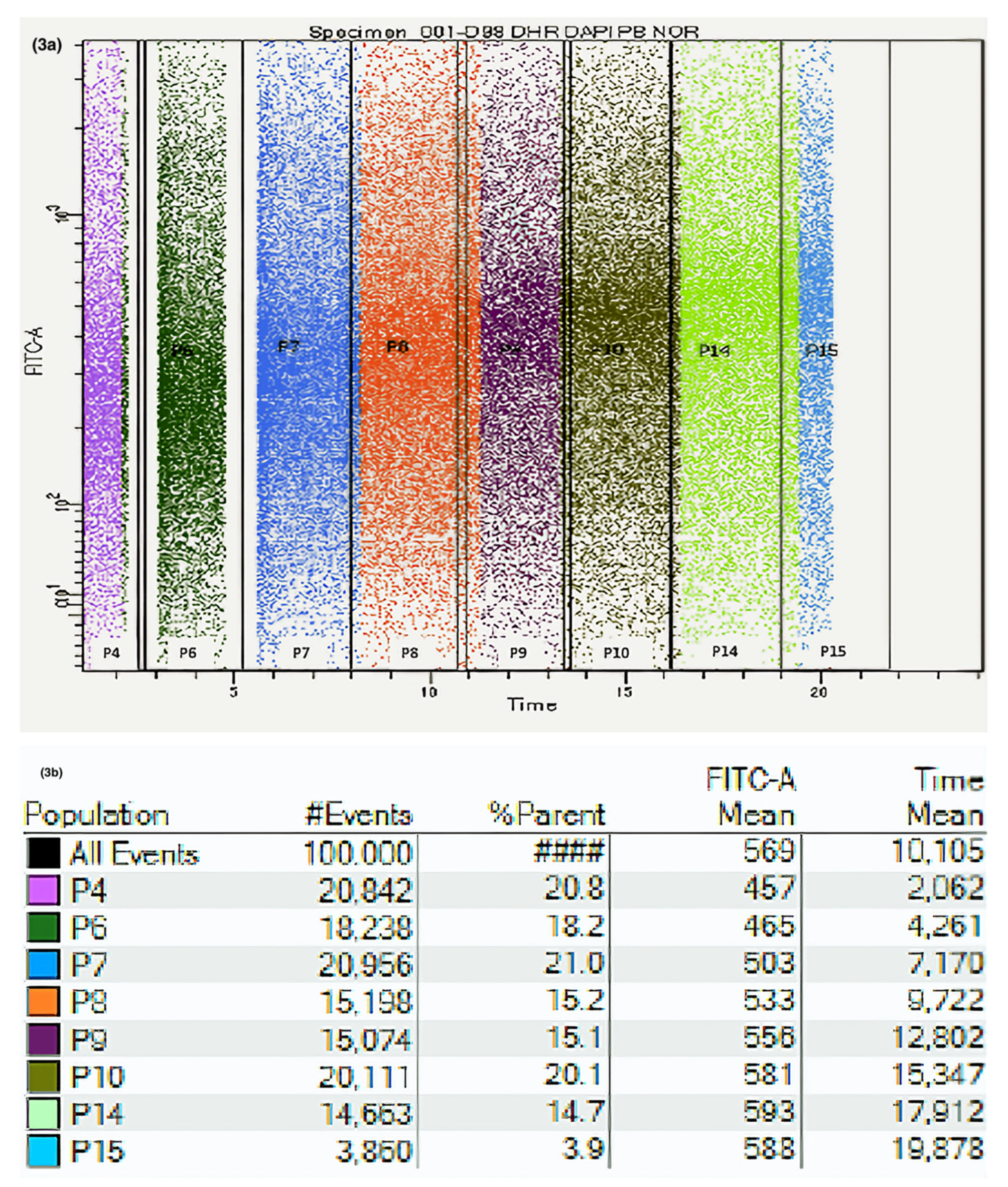
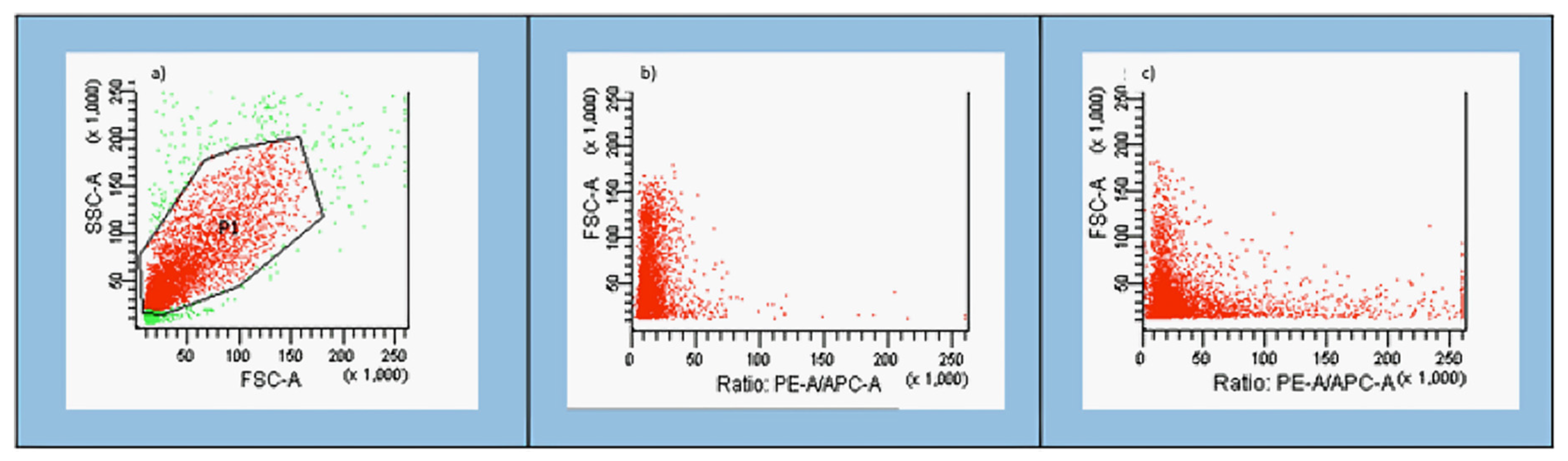
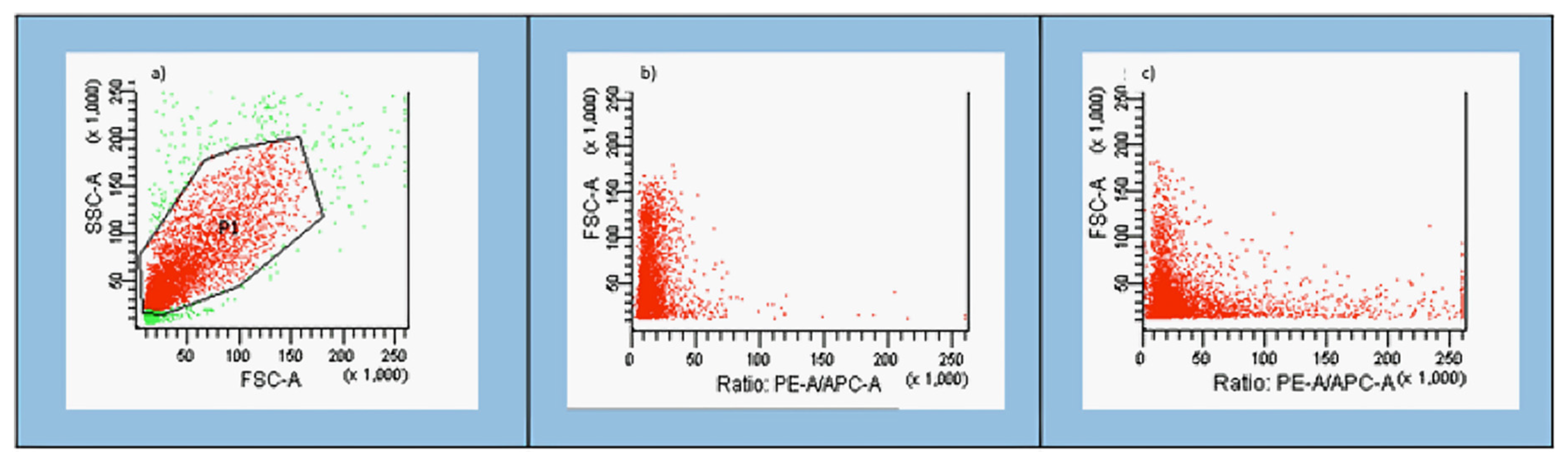
3.1. Gating Strategy
3.2. ROS and RNS Generation
3.3. GSH Detection
3.4. Intracellular Ca2+ and Plasma Membrane Potential Detection
3.5. Mitochondrial Function
3.6. Oxidative Damage in Lipids and Proteins Analysis
3.7. Apoptosis and Cell Death Detection
4. Discussion
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Le Bras, M.; Clément, M.-V.; Pervaiz, S.; Brenner, C. Reactive oxygen species and the mitochondrial signaling pathway of cell death. Histol. Histopathol. 2005, 20, 205–219. [Google Scholar]
- Weidinger, A.; Kozlov, A.V. Biological activities of reactive oxygen and nitrogen species: Oxidative stress versus signal transduction. Biomolecules 2015, 5, 472–484. [Google Scholar] [CrossRef] [Green Version]
- Figueira, T.R.; Barros, M.H.; Camargo, A.A.; Castilho, R.F.; Ferreira, J.C.; Kowaltowski, A.J.; Sluse, F.E.; Souza-Pinto, N.C.; Vercesi, A.E. Mitochondria as a source of reactive oxygen and nitrogen species: From molecular mechanisms to human health. Antioxid. Redox Signal. 2013, 18, 2029–2074. [Google Scholar] [CrossRef] [PubMed]
- Schroder, P.; Krutmann, J. Environmental oxidative stress—Environmental sources of ROS. Handb. Environ. Chem. 2005, 2, 19–31. [Google Scholar] [CrossRef]
- Gupta, R.K.; Patel, A.K.; Shah, N.; Choudhary, A.K.; Jha, U.K.; Yadav, U.C.; Gupta, P.K.; Pakuwal, U. Oxidative stress and antioxidants in disease and cancer: A review. Asian Pac. J. Cancer Prev. 2014, 15, 4405–4409. [Google Scholar] [CrossRef] [Green Version]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- Pamplona, R.; Costantini, D. Molecular and structural antioxidant defenses against oxidative stress in animals. Am. J. Physiol. Integr. Comp. Physiol. 2011, 301, R843–R863. [Google Scholar] [CrossRef] [Green Version]
- Alfadda, A.A.; Sallam, R.M. Reactive oxygen species in health and disease. J. Biomed. Biotechnol. 2012, 2012, 936486. [Google Scholar] [CrossRef]
- Evans, M.D.; Cooke, M.S. Factors contributing to the outcome of oxidative damage to nucleic acids. BioEssays 2004, 26, 533–542. [Google Scholar] [CrossRef]
- Rahman, T.; Hosen, I.; Islam, M.M.T.; Shekhar, H.U. Oxidative stress and human health. Adv. Biosci. Biotechnol. 2012, 3, 997–1019. [Google Scholar] [CrossRef] [Green Version]
- Guerra, J.I.E. Oxidative stress, diseases and antioxidant treatment. An. Med. Interna 2001, 18, 326–335. [Google Scholar]
- Günther, J.; Seyfert, H.-M. The first line of defence: Insights into mechanisms and relevance of phagocytosis in epithelial cells. Semin. Immunopathol. 2018, 40, 555–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, L.; Brighton, L.E.; Carson, J.L.; Ii, W.A.F.; Jaspers, I. Culturing of human nasal epithelial cells at the air liquid interface. J. Vis. Exp. 2013, e50646. [Google Scholar] [CrossRef]
- Thavagnanam, S.; Parker, J.C.; McBrien, M.E.; Skibinski, G.; Shields, M.D.; Heaney, L.G. Nasal epithelial cells can act as a physiological surrogate for paediatric asthma studies. PLoS ONE 2014, 9, e85802. [Google Scholar] [CrossRef] [Green Version]
- Domej, W.; Oetll, K.; Renner, W. Oxidative stress and free radicals in COPD—Implications and relevance for treatment. Int. J. Chronic Obs. Pulm. Dis. 2014, 9, 1207–1224. [Google Scholar] [CrossRef] [Green Version]
- Mishra, V.; Banga, J.; Silveyra, P. Oxidative stress and cellular pathways of asthma and inflammation: Therapeutic strategies and pharmacological targets. Pharm. Ther. 2017, 181, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Escribano, A.; Pastor, S.; Reula, A.; Castillo, S.; Vicente, S.; Sanz, F.; Casas, F.; Torres, M.; Fernández-Fabrellas, E.; Codoñer-Franch, P.; et al. Accelerated telomere attrition in children and teenagers with α1-antitrypsin deficiency. Eur. Respir. J. 2016, 48, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Torres-Durán, M.; Lopez-Campos, J.L.; Barrecheguren, M.; Miravitlles, M.; Martinez-Delgado, B.; Castillo, S.; Escribano, A.; Baloira, A.; Navarro-Garcia, M.M.; Pellicer, D.; et al. Alpha-1 antitrypsin deficiency: Outstanding questions and future directions. Orphanet J. Rare Dis. 2018, 13, 114. [Google Scholar] [CrossRef] [Green Version]
- Escribano, A.; Amor, M.; Pastor, S.; Castillo, S.; Sanz, F.; Codoñer-Franch, P.; Dasí, F. Decreased glutathione and low catalase activity contribute to oxidative stress in children with α-1 antitrypsin deficiency: Table 1. Thorax 2014, 70, 82–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magallón, M.; Navarro-García, M.M.; Dasí, F. Oxidative Stress in COPD. J. Clin. Med. 2019, 8, 1953. [Google Scholar] [CrossRef] [Green Version]
- Zihlif, N.; Paraskakis, E.; Tripoli, C.; Lex, C.; Bush, A. Markers of airway inflammation in primary ciliary dyskinesia studied using exhaled breath condensate. Pediatr. Pulmonol. 2006, 41, 509–514. [Google Scholar] [CrossRef]
- Rickham, P.P. Human experimentation: Code of ethics of W.M.A. BMJ 1964, 2, 177. [Google Scholar] [CrossRef] [Green Version]
- Wallberg, F.; Tenev, T.; Meier, P. Analysis of apoptosis and necroptosis by fluorescence-activated cell sorting. Cold Spring Harb. Protoc. 2016, 2016, 087387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Gurtu, V.; Kain, S.R.; Yan, G. Early detection of apoptosis using a fluorescent conjugate of annexin V. Biotechniques 1997, 23, 525–531. [Google Scholar] [CrossRef]
- Kalyanaraman, B. Oxidative chemistry of fluorescent dyes: Implications in the detection of reactive oxygen and nitrogen species. Biochem. Soc. Trans. 2011, 39, 1221–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyanaraman, B.; Hardy, M.; Podsiadly, R.; Cheng, G.; Zielonka, J. Recent developments in detection of superoxide radical anion and hydrogen peroxide: Opportunities, challenges, and implications in redox signaling. Arch. Biochem. Biophys. 2017, 617, 38–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eruslanov, E.; Kusmartsev, S. Identification of ROS using oxidized DCFDA and flow-cytometry. Methods Mol. Biol. 2010, 594, 57–72. [Google Scholar] [CrossRef]
- Crow, J.P. Dichlorodihydrofluorescein and Dihydrorhodamine 123 are sensitive indicators of Peroxynitritein Vitro: Implications for intracellular measurement of reactive nitrogen and oxygen species. Nitric Oxide 1997, 1, 145–157. [Google Scholar] [CrossRef]
- Dikalov, S.I.; Harrison, D.G. Methods for detection of mitochondrial and cellular reactive oxygen species. Antioxid. Redox Signal. 2014, 20, 372–382. [Google Scholar] [CrossRef] [Green Version]
- Díez, I.; Calatayud, S.; Hernandez, C.; Quintana, E.; O’Connor, J.; Esplugues, J.; Barrachina, M.D. Nitric oxide, derived from inducible nitric oxide synthase, decreases hypoxia inducible factor-1α in macrophages during aspirin-induced mesenteric inflammation. Br. J. Pharm. 2010, 159, 1636–1645. [Google Scholar] [CrossRef] [Green Version]
- Namin, S.M.; Nofallah, S.; Joshi, M.S.; Kavallieratos, K.; Tsoukias, N.M. Kinetic analysis of DAF-FM activation by NO: Toward calibration of a NO-sensitive fluorescent dye. Nitric Oxide 2013, 28, 39–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedley, D.W.; Chow, S. Evaluation of methods for measuring cellular glutathione content using flow cytometry. Cytometry 1994, 15, 349–358. [Google Scholar] [CrossRef] [Green Version]
- Sebastià, J.; Cristòfol, R.; Martin, M.; Rodríguez-Farré, E.; Sanfeliu, C. Evaluation of fluorescent dyes for measuring intracellular glutathione content in primary cultures of human neurons and neuroblastoma SH-SY5Y. Cytom. Part A 2002, 51, 16–25. [Google Scholar] [CrossRef]
- Evans, J.A.; Darlington, D.N.; Gann, D.S. A circulating factor(s) mediates cell depolarization in hemorrhagic shock. Ann. Surg. 1991, 213, 549–557. [Google Scholar] [CrossRef]
- Perry, S.W.; Norman, J.P.; Barbieri, J.; Brown, E.B.; Gelbard, H.A. Mitochondrial membrane potential probes and the proton gradient: A practical usage guide. Biotechnique 2011, 50, 98–115. [Google Scholar] [CrossRef]
- Petrat, F.; Pindiur, S.; Kirsch, M.; De Groot, H. “Mitochondrial” photochemical drugs do not release toxic amounts of 1O2 within the mitochondrial matrix space. Arch. Biochem. Biophys. 2003, 412, 207–215. [Google Scholar] [CrossRef]
- Scaduto, R.C.; Grotyohann, L.W. Measurement of mitochondrial membrane potential using fluorescent rhodamine derivatives. Biophys. J. 1999, 76, 469–477. [Google Scholar] [CrossRef] [Green Version]
- Presley, A.D.; Fuller, K.M.; Arriaga, E.A. MitoTracker Green labeling of mitochondrial proteins and their subsequent analysis by capillary electrophoresis with laser-induced fluorescence detection. J. Chromatogr. B 2003, 793, 141–150. [Google Scholar] [CrossRef]
- Doherty, E.; Perl, A. Measurement of mitochondrial mass by flow cytometry during oxidative stress. React. Oxyg. Species 2017, 4, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, P.; Rajesh, M.; Yoshihiro, K.; Haskó, G.; Pacher, P. Simple quantitative detection of mitochondrial superoxide production in live cells. Biochem. Biophys. Res. Commun. 2007, 358, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, B.C.; Lin, V.S.; Chang, C.J. Preparation and use of MitoPY1 for imaging hydrogen peroxide in mitochondria of live cells. Nat. Protoc. 2013, 8, 1249–1259. [Google Scholar] [CrossRef] [Green Version]
- Gee, K.; Brown, K.; Chen, W.-N.; Bishop-Stewart, J.; Gray, D.; Johnson, I. Chemical and physiological characterization of fluo-4 Ca2+-indicator dyes. Cell Calcium 2000, 27, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Wesseling, M.C.; Wagner-Britz, L.; Boukhdoud, F.; Asanidze, S.; Nguyen, D.B.; Kaestner, L.; Bernhardt, I. Measurements of intracellular Ca2+ content and phosphatidylserine exposure in human red blood cells: Methodological issues. Cell. Physiol. Biochem. 2016, 38, 2414–2425. [Google Scholar] [CrossRef] [PubMed]
- Naguib, Y.M.A. Antioxidant Activities of Astaxanthin and Related Carotenoids. J. Agric. Food Chem. 2000, 48, 1150–1154. [Google Scholar] [CrossRef] [PubMed]
- Raudsepp, P.; Brüggemann, D.A.; Andersen, M.L. Detection of radicals in single droplets of oil-in-water emulsions with the lipophilic fluorescent probe BODIPY665/676 and confocal laser scanning microscopy. Free Radic. Biol. Med. 2014, 70, 233–240. [Google Scholar] [CrossRef]
- Chaudhuri, A.R.; De Waal, E.M.; Pierce, A.; Van Remmen, H.; Ward, W.F.; Richardson, A. Detection of protein carbonyls in aging liver tissue: A fluorescence-based proteomic approach. Mech. Ageing Dev. 2006, 127, 849–861. [Google Scholar] [CrossRef] [PubMed]
- Perumalsamy, H.; Sankarapandian, K.; Kandaswamy, N.; Balusamy, S.R.; Periyathambi, D.; Raveendiran, N. Cellular effect of styrene substituted biscoumarin caused cellular apoptosis and cell cycle arrest in human breast cancer cells. Int. J. Biochem. Cell Biol. 2017, 92, 104–114. [Google Scholar] [CrossRef]
- Akasaki, Y.; Alvarez-Garcia, O.; Saito, M.; Caramés, B.; Iwamoto, Y.; Lotz, M.K. FoxO transcription factors support oxidative stress resistance in human chondrocytes. Arthritis Rheumatol. 2014, 66, 3349–3358. [Google Scholar] [CrossRef] [Green Version]
- Beltrán, B.; Nos, P.; Dasí, F.; Iborra, M.; Bastida, G.; Martínez, M.; Oʼconnor, J.-E.; Sáez, G.; Moret, I.; Ponce, J. Mitochondrial dysfunction, persistent oxidative damage, and catalase inhibition in immune cells of naïve and treated Crohn’s disease. Inflamm. Bowel Dis. 2010, 16, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Herrera, G.; Martínez, A.; O’Cornor, J.; Blanco, M. Functional assays of oxidative stress using genetically engineered Escherichia coli Strains. Curr. Protoc. Cytom. 2003, 24, 11–16. [Google Scholar] [CrossRef]
- Thor, H.; Smith, M.T.; Hartzell, P.; Bellomo, G.; Jewell, S.A.; Orrenius, S. The metabolism of menadione (2-methyl-1,4-naphthoquinone) by isolated hepatocytes. A study of the implications of oxidative stress in intact cells. J. Biol. Chem. 1982, 257, 12419–12425. [Google Scholar] [CrossRef]
- Khailova, L.S.; Rokitskaya, T.I.; Kotova, E.A.; Antonenko, Y.N. Effect of cyanide on mitochondrial membrane depolarization induced by uncouplers. Biochemistry 2017, 82, 1140–1146. [Google Scholar] [CrossRef]
- Katerji, M.; Filippova, M.; Duerksen-Hughes, P. Approaches and methods to measure oxidative stress in clinical samples: Research applications in the cancer field. Oxidative Med. Cell. Longev. 2019, 2019, 1279250. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, B.; Sblendorio, V. Oxidative stress tests: Overview on reliability and use. Part II. Eur. Rev. Med Pharm. Sci. 2008, 11, 383–399. [Google Scholar]
- Palmieri, B.; Sblendorio, V. Oxidative stress tests: Overview on reliability and use. Part I. Eur. Rev. Med. Pharm. Sci. 2007, 11, 309–342. [Google Scholar]
- Palmieri, B.; Sblendorio, V. Current status of measuring oxidative stress. Methods Mol. Biol. 2009, 594, 3–17. [Google Scholar] [CrossRef]
- Rahman, I.; Biswas, S.K. Non-invasive biomarkers of oxidative stress: Reproducibility and methodological issues. Redox Rep. 2004, 9, 125–143. [Google Scholar] [CrossRef]
- Bhatti, J.S.; Bhatti, G.K.; Reddy, P.H. Mitochondrial dysfunction and oxidative stress in metabolic disorders—A step towards mitochondria based therapeutic strategies. Biochim. Biophys. Acta Basis Dis. 2017, 1863, 1066–1077. [Google Scholar] [CrossRef]
- Tinning, P.W.; Franssen, A.J.P.M.; Hridi, S.U.; Bushell, T.J.; McConnell, G. A 340/380 Nm Light Emitting Diode Illuminator for Fura-2 AM Ratiometric Ca2+ imaging of live cells with better than 5 NM Precision. J Microsc. 2018, 269, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reula, A.; Lucas, J.S.; Moreno-Galdó, A.; Romero, T.; Milara, X.; Carda, C.; Mata-Roig, M.; Escribano, A.; Dasi, F.; Armengot-Carceller, M. New insights in primary ciliary dyskinesia. Expert Opin. Orphan Drugs 2017, 5, 537–548. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reactive Species | Measurement | Stock Concentration | Storage Temperature | Final Concentration | Flow Cytometer | Excitation Laser | Detector | Cell Number | Total Volume | Incubation Time | Incubation Temperature |
|---|---|---|---|---|---|---|---|---|---|---|---|
| DIBAC | Plasma membrane potential | 100 µM | −20 °C | 1.2 µM | FACS Verse | Blue (488 nm) | 527/32507 LP | 8.000 | 250 µL | 20 min | 37 °C |
| FLUO-4 | Intracellular Ca2+ | 50 µM | −20 °C | 0.5 µM | LSR Fortessa X-20 | Blue (488 nm) | 530/30505LP | 8.000 | 250 µL | 20 min | 37 °C |
| CMFDA | Reduced thiols (GSH) | 10 µM | −20 °C | 25 nM | FACS Verse | Blue (488 nm) | 527/32507 LP | 8.000 | 250 µL | 20 min | 37 °C |
| DAF-FMDA | NO | 1.25 mM | −20 °C | 1 µM | LSR Fortessa X-20 | Blue (488 nm) | 530/30505LP | 8.000 | 250 µL | 20 min | 37 °C |
| DCF | Intracellular peroxides | 1 mg/mL | −20 °C | 2.5 µg/mL | FACS Verse | Blue (488 nm) | 527/32507 LP | 8.000 | 250 µL | 20 min | 37 °C |
| MitoSOX | Motochondrial O2− | 0.5 mM | −20 °C | 640 nM | FACS Verse | Blue (488 nm) | 700/54665LP | 8.000 | 250 µL | 20 min | 37 °C |
| Mitotracker Green | Mitochondrial mass | 10 µM | −20 °C | 78 nM | FACS Verse | Blue (488 nm) | 527/32507 LP | 8.000 | 250 µL | 20 min | 37 °C |
| BODIPY 665/676 | Oxidized/reduced lipid ratio | 1 mM | −20 °C | 800 nM | FACS Aria III | Blue (488 nm) Red (635 nm) | 586/42556LP and 780/60735LP | 12.000 | 250 µL | 30 min | 37 °C |
| TMRM | Mitochondrial Ψm | 240 µM | −20 °C | 600 nM | FACS Verse | Blue (488 nm) | 586/42560LP | 8.000 | 250 µL | 20 min | 37 °C |
| DHR 123 | ONOO− | 5 mM | −20 °C | 100 µM | LSR Fortessa X-20 | Blue (488 nm) | 530/30505LP | 12.000 | 250 µL | 20 min | 37 °C |
| HE | O2− | 1 mg/mL | −20 °C | 2.5 µg/mL | FACS Verse | Blue (488 nm) | 700/54665LP | 8.000 | 250 µL | 20 min | 37 °C |
| FTC | Protein carbonylation | 1 mM | −20 °C | 800 nM | FACS Verse | Blue (488 nm) | 527/32507 LP | 8.000 | 250 µL | 20 min | 37 °C |
| MitoPY | Mitochondrial H2O2 | 1 mM | −20 °C | 4 µM | FACS Verse | Blue (488 nm) | 527/32507 LP | 8.000 | 250 µL | 20 min | 37 °C |
| DAPI | Cell death | 1 mg/mL | −20 °C | 800 ng/mL | FACS Verse | Violet (405 nm) | 448/45 | 8.000 | 250 µL | 20 min | 37 °C |
| PI | Cell death | 1 mg/mL | 4 °C | 8 µg/mL | FACS Verse | Blue (488 nm) | 586/42560LP | 12.000 | 100 µL | 15 min | RT |
| Annexin V | Apoptosis | - | 4 °C | - | FACS Verse | Blue (488 nm) | 527/32507 LP | 12.000 | 100 µL | 15 min | RT |
| Reactive Species | Measurement | Inductor | Stock Concentration | Final Concentration | Storage Temperature | Inductor Incubation Time | Inductor Incubation Temperature | Reactive Incubation Time | Reactive Incubation Temperature |
|---|---|---|---|---|---|---|---|---|---|
| DIBAC | Plasmatic membrane potential | t-BHP | 7.7 mM | 100 μM | 4 °C | 15 min | 37 °C | 30 min | 37 °C |
| FLUO-4 | Intracellular Ca2+ | Ionomycin | 1.338 mM | 50 μM | −20 °C | Kynetics | Kynetics | ||
| CMFDA | Reduced thiols | DEM | - | 20 mM | −20 °C | 90 min | 37 °C | 30 min | 37 °C |
| DAF-FM DA | NO | NOR-1 | 1 mg/mL | 16 μg/mL | −20 °C | Kynetics | Kynetics | ||
| DCF | Intracellular peroxides | t-BHP | 7.7 mM | 100 μM | 4 °C | 15 min | 37 °C | 30 min | 37 °C |
| MitoSOX | Mitochondrial O2− | PB | 2.8 mg/mL | 2.24 μg/mL | −20 °C | 15 min | 37 °C | 30 min | 37 °C |
| BODIPY 665/676 | Oxidized/reduced lipids ratio | t-BHP | 7.7 mM | 100 μM | 4 °C | 15 min | 37 °C | 30 min | 37 °C |
| TMRM | Mitochondrial Ψm | FCCP | 10 mM | 52 μM | −20 °C | 15 min | 37 °C | 30 min | 37 °C |
| DHR 123 | ONOO- | NOR-1 and PB | 1 mg/mL and 2.8 mg/mL | 16 μg/mL and 2.24 μg/mL | −20 °C | Kynetics | Kynetics | ||
| HE | O2− | PB | 2.8 mg/mL | 2.24 μg/mL | −20 °C | 15 min | 37 °C | 30 min | 37 °C |
| FTC | Protein carbonylation | Menadione | 10 mg/mL | 1 mM | −20 °C | 60 min | 37 °C | 30 min | 37 °C |
| MitoPY | Mitochondrial H2O2 | t-BHP | 7.7 mM | 100 μM | 4 °C | 15 min | 37 °C | 30 min | 37 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reula, A.; Pellicer, D.; Castillo, S.; Magallón, M.; Armengot, M.; Herrera, G.; O’Connor, J.-E.; Bañuls, L.; Navarro-García, M.M.; Escribano, A.; et al. New Laboratory Protocol to Determine the Oxidative Stress Profile of Human Nasal Epithelial Cells Using Flow Cytometry. J. Clin. Med. 2021, 10, 1172. https://doi.org/10.3390/jcm10061172
Reula A, Pellicer D, Castillo S, Magallón M, Armengot M, Herrera G, O’Connor J-E, Bañuls L, Navarro-García MM, Escribano A, et al. New Laboratory Protocol to Determine the Oxidative Stress Profile of Human Nasal Epithelial Cells Using Flow Cytometry. Journal of Clinical Medicine. 2021; 10(6):1172. https://doi.org/10.3390/jcm10061172
Chicago/Turabian StyleReula, Ana, Daniel Pellicer, Silvia Castillo, María Magallón, Miguel Armengot, Guadalupe Herrera, José-Enrique O’Connor, Lucía Bañuls, María Mercedes Navarro-García, Amparo Escribano, and et al. 2021. "New Laboratory Protocol to Determine the Oxidative Stress Profile of Human Nasal Epithelial Cells Using Flow Cytometry" Journal of Clinical Medicine 10, no. 6: 1172. https://doi.org/10.3390/jcm10061172
APA StyleReula, A., Pellicer, D., Castillo, S., Magallón, M., Armengot, M., Herrera, G., O’Connor, J.-E., Bañuls, L., Navarro-García, M. M., Escribano, A., & Dasí, F. (2021). New Laboratory Protocol to Determine the Oxidative Stress Profile of Human Nasal Epithelial Cells Using Flow Cytometry. Journal of Clinical Medicine, 10(6), 1172. https://doi.org/10.3390/jcm10061172