
Effects of 5-Year Nitrogen Addition on Species Composition and Diversity of an Alpine Steppe Plant Community on Qinghai-Tibetan Plateau


 ,
,
Abstract
:1. Introduction
2. Results
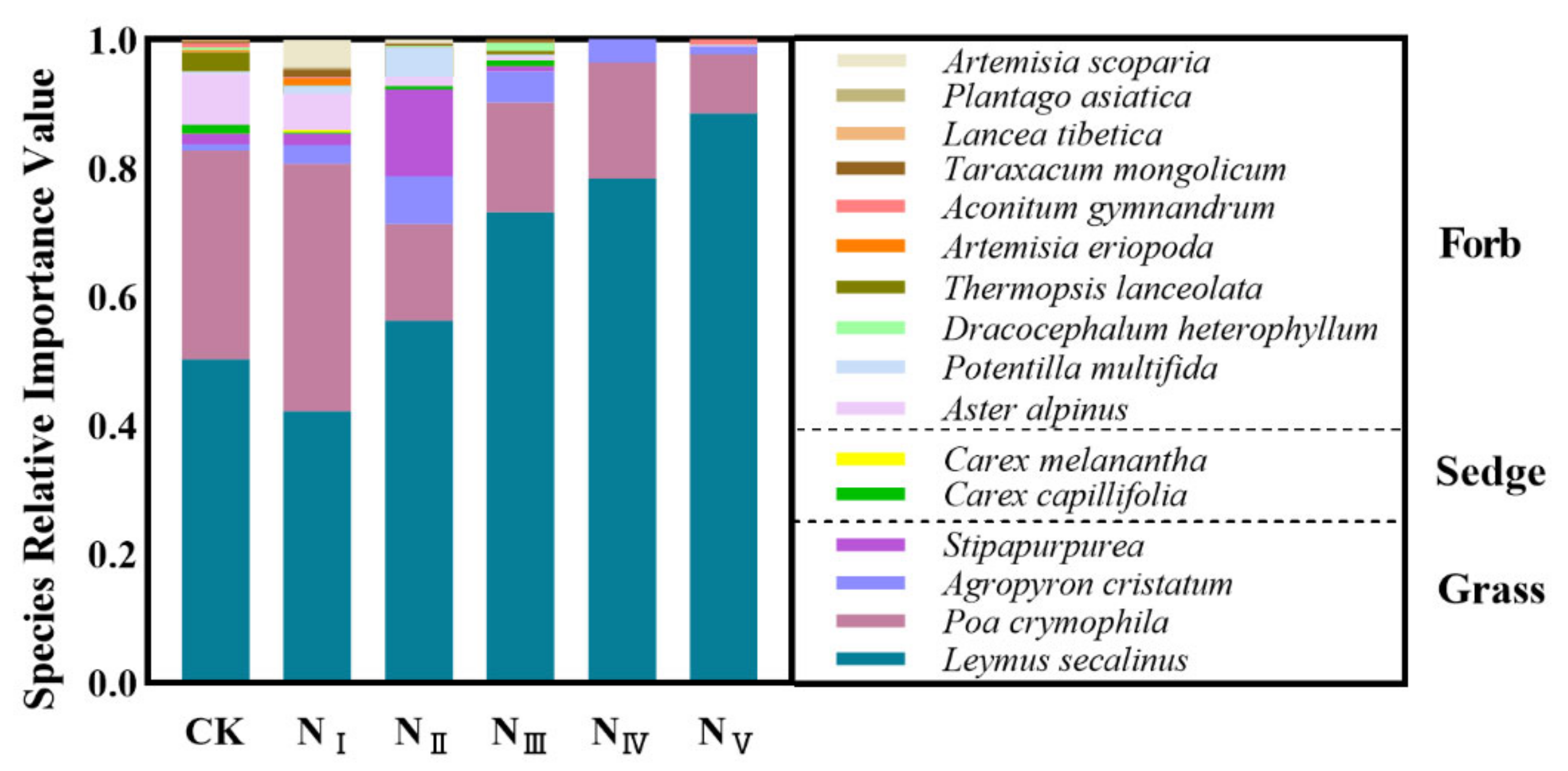
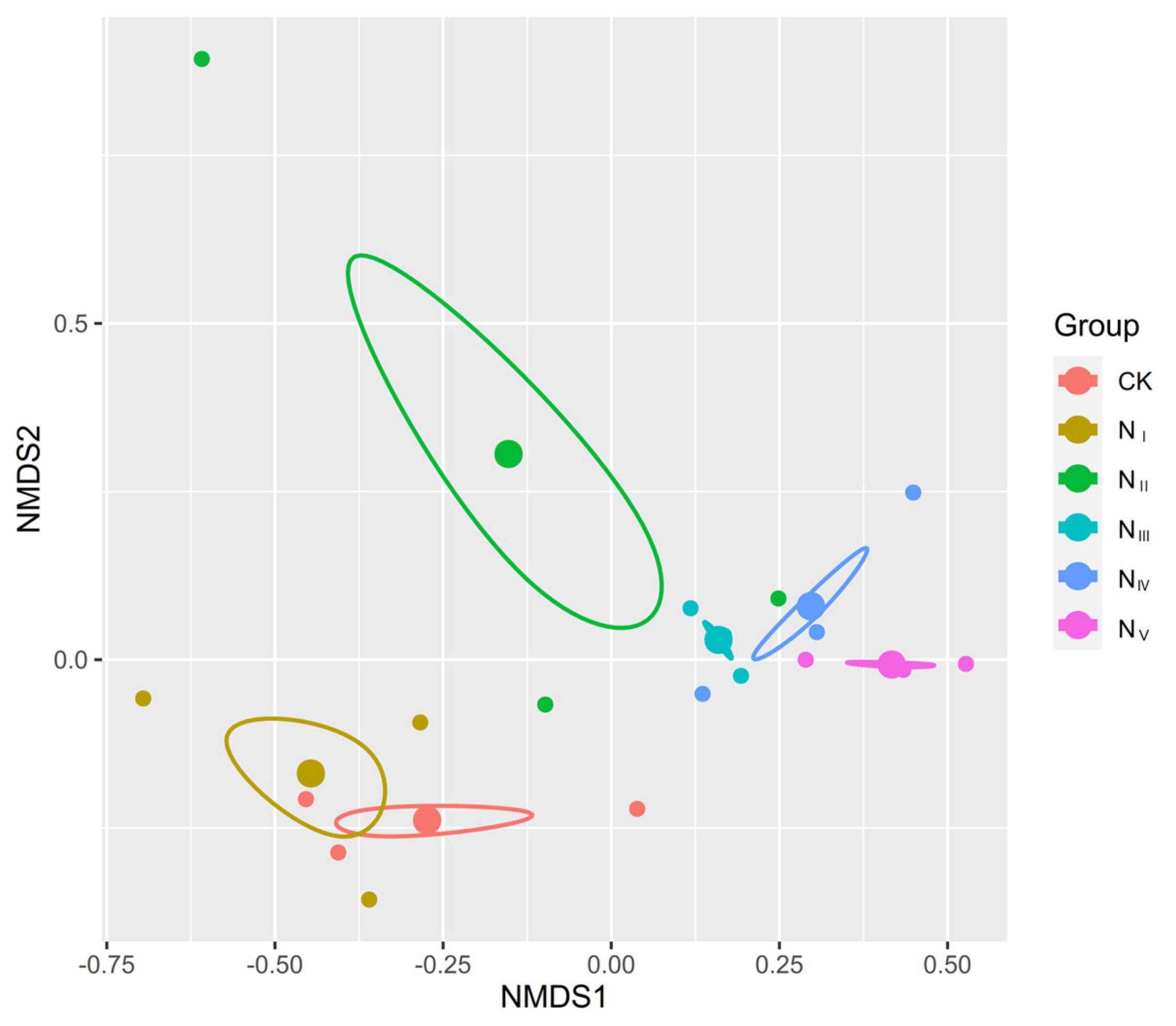
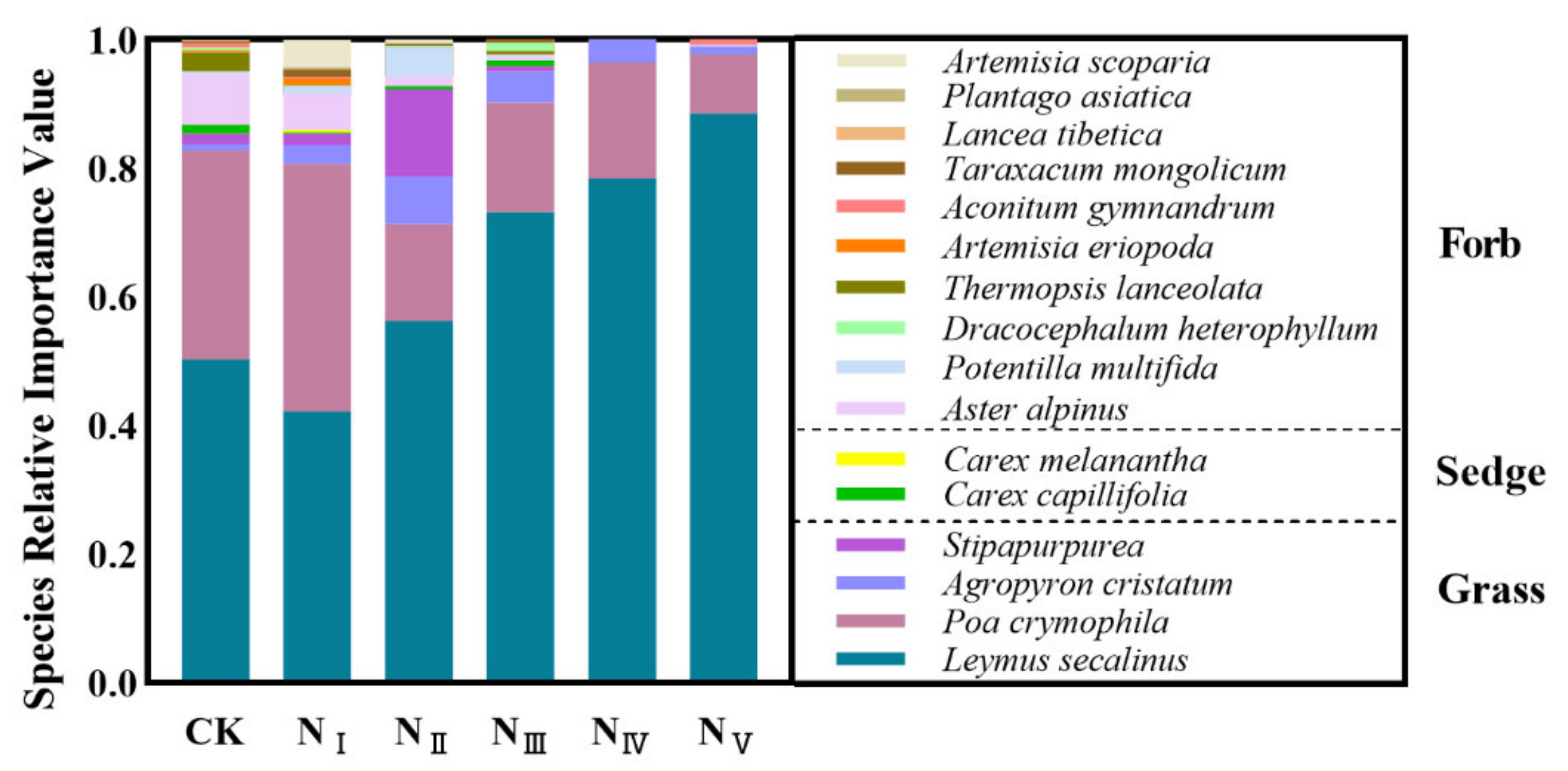
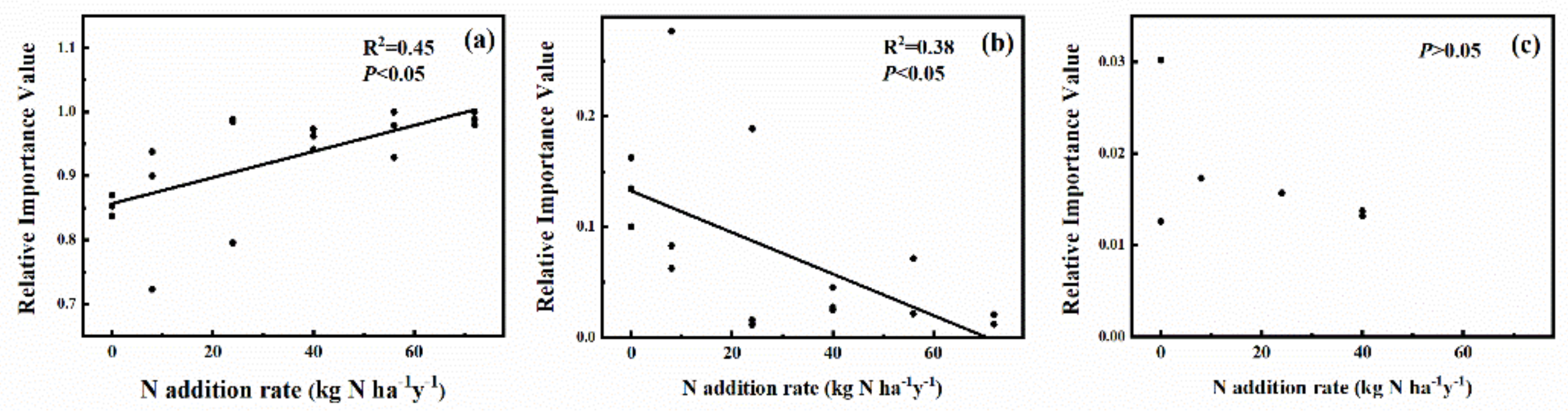
2.1. Effects of N Addition on Species Composition of the Plant Community
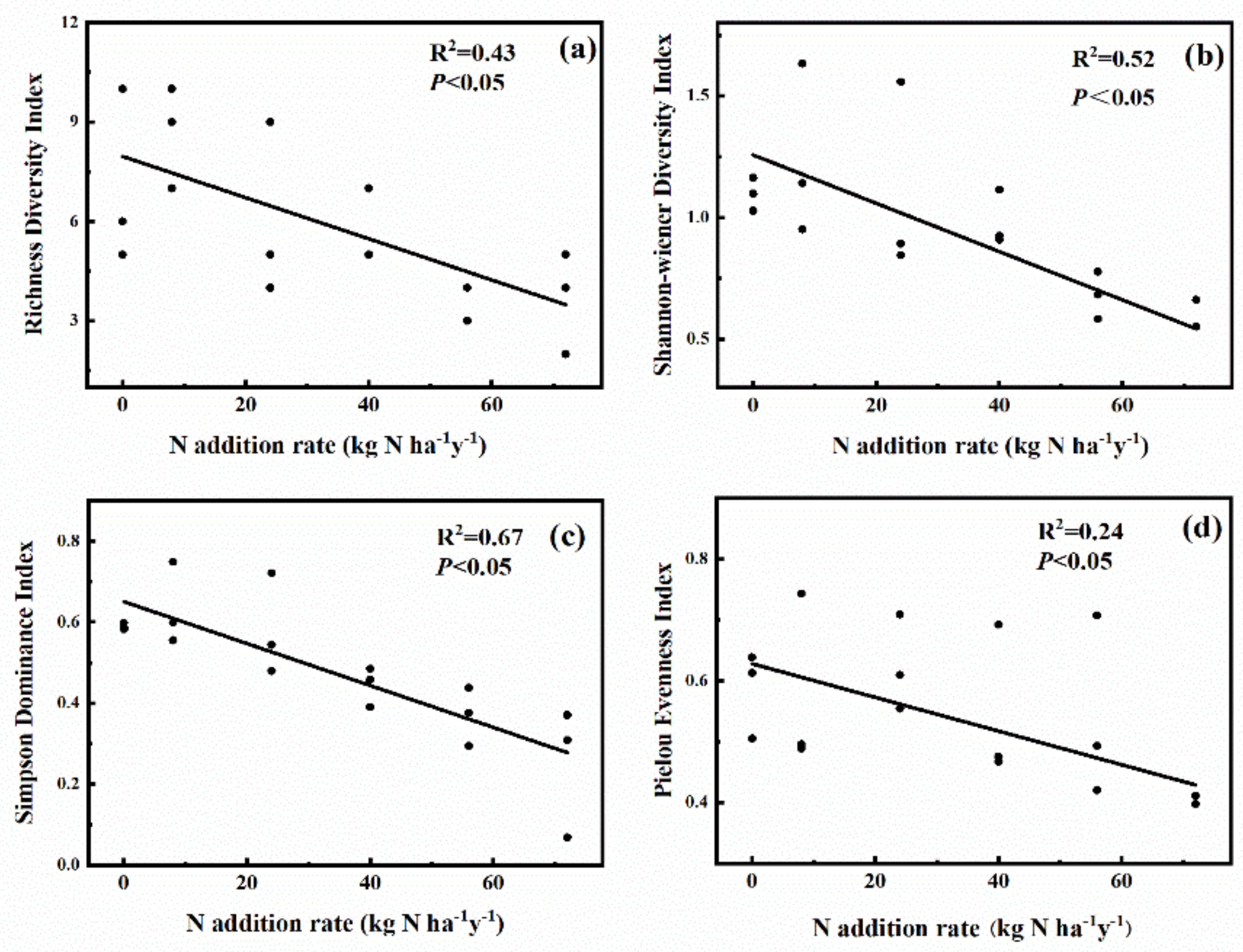
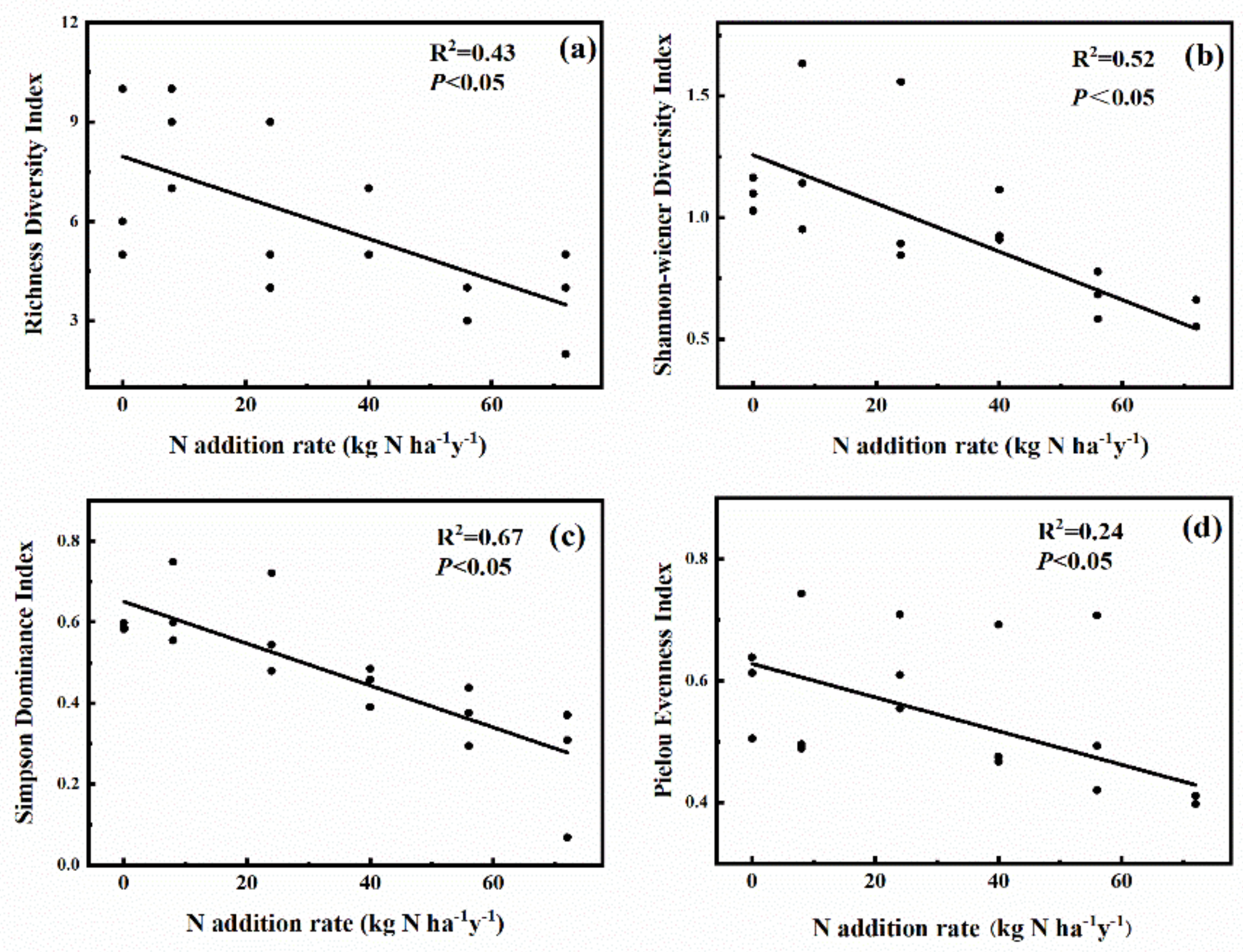
2.2. Effects of N Addition on Species Diversity of Plant Community
2.3. Relationship between Species Diversity and Soil Nutrients in the Alpine Steppe
3. Discussion
3.1. Responses of Plant Species Composition of Alpine Steppe to N Addition
3.2. Responses of Plant Species Diversity of the Alpine Steppe to N Addition
3.3. Soil Nutrients Affecting Plant Species Diversity of the Alpine Steppe under N Addition
4. Materials and Methods
4.1. Study Sites
4.2. Experimental Design
4.3. Plant and Soil Sampling
4.4. Statistical Analysis
4.4.1. Species Importance Value
4.4.2. Species Diversity Index
4.4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lü, C.Q.; Tian, H.Q. Spatial and temporal patterns of nitrogen deposition in China: Synthesis of observational data. J. Geophys. Res. 2007, 112. [Google Scholar] [CrossRef]
- Zhong, X.L.; Li, J.T.; Li, X.J.; Ye, Y.C.; Liu, S.S.; Hallett, P.D.; Ogden, M.R.; Naveed, M. Physical protection by soil aggregates stabilizes soil organic carbon under simulated N deposition in a subtropical forest of China. Geoderma 2017, 285, 323–332. [Google Scholar] [CrossRef]
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galloway, J.N.; Dentener, F.J.; Capone, D.G.; Boyer, E.W.; Howarth, R.W.; Seitzinger, S.P.; Asner, G.P.; Cleveland, C.C.; Green, P.A.; Holland, E.A.; et al. Nitrogen cycles: Past, present, and future. Biogeochemistry 2004, 70, 153–226. [Google Scholar] [CrossRef]
- Yu, G.R.; Jia, Y.L.; He, N.P.; Zhu, J.X.; Chen, Z.; Wang, Q.F.; Piao, S.L.; Liu, X.J.; He, H.L.; Guo, X.B.; et al. Stabilization of atmospheric nitrogen deposition in China over the past decade. Nat. Geosci. 2019, 12, 424–429. [Google Scholar] [CrossRef]
- Lin, J.; Wang, Y.; Sun, S.; Mu, C.; Yan, X. Effects of arbuscular mycorrhizal fungi on the growth, photosynthesis and photosynthetic pigments of Leymus chinensis seedlings under salt-alkali stress and nitrogen deposition. Sci. Total Environ. 2017, 576, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Field, C.D.; Evans, C.D.; Dise, N.B.; Hall, J.R.; Caporn, S.J.M. Long-term nitrogen deposition increases heathland carbon sequestration. Sci. Total Environ. 2017, 592, 426–435. [Google Scholar] [CrossRef]
- Shen, H.; Dong, S.; Li, S.; Xiao, J.; Han, Y.; Yang, M.; Zhang, J.; Gao, X.; Xu, Y.; Li, Y.; et al. Effects of simulated N deposition on photosynthesis and productivity of key plants from different functional groups of alpine meadow on Qinghai-Tibetan plateau. Environ. Pollut. 2019, 251, 731–737. [Google Scholar] [CrossRef]
- Bai, Y.F.; Wu, J.G.; Clark, C.M.; Naeem, S.; Pan, Q.M.; Huang, J.H.; Zhang, L.X.; Han, X.G. Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from inner Mongolia Grasslands. Glob. Change Biol. 2010, 16, 358–372. [Google Scholar] [CrossRef]
- Nakaji, T.; Fukami, M.; Dokiya, Y.; Izuta, T. Effects of high nitrogen load on growth, photosynthesis and nutrient status of Cryptomeria japonica and Pinus densiflora seedlings. Trees 2001, 15, 453–461. [Google Scholar] [CrossRef]
- Wang, J.; Gao, Y.; Zhang, Y.; Yang, J.; Smith, M.D.; Knapp, A.K.; Eissenstat, D.M.; Han, X. Asymmetry in above- and belowground productivity responses to N addition in a semi-arid temperate steppe. Glob. Change Biol. 2019, 25, 2958–2969. [Google Scholar] [CrossRef] [PubMed]
- Payne, R.J.; Dise, N.B.; Field, C.D.; Dore, A.J.; Caporn, S.J.M.; Stevens, C.J. Nitrogen deposition and plant biodiversity: Past, present, and future. Front. Ecol. Environ. 2017, 15, 431–436. [Google Scholar] [CrossRef]
- Phoenix, G.K.; Hicks, W.K.; Cinderby, S.; Kuylenstierna, J.C.I.; Stock, W.D.; Dentener, F.J.; Giller, K.E.; Austin, A.T.; Lefroy, R.D.B.; Gimeno, B.S.; et al. Atmospheric nitrogen deposition in world biodiversity hotspots: The need for a greater global perspective in assessing N deposition impacts. Glob. Change Biol. 2006, 12, 470–476. [Google Scholar] [CrossRef]
- Zhang, Y.; Loreau, M.; Lu, X.; He, N.; Zhang, G.; Han, X. Nitrogen enrichment weakens ecosystem stability through decreased species asynchrony and population stability in a temperate grassland. Glob. Chang Biol. 2016, 22, 1445–1455. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef]
- Avolio, M.L.; Koerner, S.E.; La Pierre, K.J.; Wilcox, K.R.; Wilson, G.W.T.; Smith, M.D.; Collins, S.L.; MacDougall, A. Changes in plant community composition, not diversity, during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrass prairie. J. Ecol. 2014, 102, 1649–1660. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.H.; Dong, S.K.; Zhao, Z.Z.; Sha, W.; Li, S.; Shen, H.; Xiao, J.N.; Zhang, J.; Wu, X.Y.; Jiang, X.M.; et al. Response of soil nutrients and stoichiometry to elevated nitrogen deposition in alpine grassland on the Qinghai-Tibetan Plateau. Geoderma 2019, 343, 263–268. [Google Scholar] [CrossRef]
- He, K.; Qi, Y.; Huang, Y.; Chen, H.; Sheng, Z.; Xu, X.; Duan, L. Response of aboveground biomass and diversity to nitrogen addition-a five-year experiment in semi-arid grassland of Inner Mongolia, China. Sci. Rep. 2016, 6, 31919. [Google Scholar] [CrossRef] [Green Version]
- Stevens, C.J.; Dise, N.B.; Gowing, D.J.G.; Mountford, J.O. Loss of forb diversity in relation to nitrogen deposition in the UK: Regional trends and potential controls. Glob. Change Biol. 2006, 12, 1823–1833. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Liu, H.; Song, Z.; Wang, W.; Hu, G.; Qi, Z. Response of aboveground biomass and diversity to nitrogen addition along a degradation gradient in the Inner Mongolian steppe, China. Sci. Rep. 2015, 5, 10284. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Bowker, M.A.; Tao, Y.; Wu, L.; Zhang, Y. Chronic nitrogen addition induces a cascade of plant community responses with both seasonal and progressive dynamics. Sci. Total Environ. 2018, 626, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Kwaku, E.A.; Dong, S.; Shen, H.; Li, W.; Sha, W.; Su, X.; Zhang, Y.; Li, S.; Gao, X.; Liu, S.; et al. Biomass and Species Diversity of Different Alpine Plant Communities Respond Differently to Nitrogen Deposition and Experimental Warming. Plants 2021, 10, 2719. [Google Scholar] [CrossRef] [PubMed]
- Standish, R.J.; Fontaine, J.B.; Harris, R.J.; Stock, W.D.; Hobbs, R.J. Interactive effects of altered rainfall and simulated nitrogen deposition on seedling establishment in a global biodiversity hotspot. Oikos 2012, 121, 2014–2025. [Google Scholar] [CrossRef] [Green Version]
- Stevens, C.J.; Dise, N.B.; Mountford, J.O.; Gowing, D.J. Impact of nitrogen deposition on the species richness of grasslands. Science 2004, 303, 1876–1879. [Google Scholar] [CrossRef] [Green Version]
- Klein, J.A.; Harte, J.; Zhao, X.Q. Decline in medicinal and forage species with warming is mediated by plant traits on the Tibetan Plateau. Ecosystems 2008, 11, 775–789. [Google Scholar] [CrossRef]
- Zhao, Z.; Dong, S.K.; Jiang, X.; Liu, S.; Ji, H.; Li, Y.; Han, Y.; Sha, W. Effects of warming and nitrogen deposition on CH4, CO2 and N2O emissions in alpine grassland ecosystems of the Qinghai-Tibetan Plateau. Sci. Total Environ. 2017, 592, 565–572. [Google Scholar] [CrossRef]
- Dong, S.K.; Shang, Z.H.; Gao, J.X.; Boone, R.B. Enhancing sustainability of grassland ecosystems through ecological restoration and grazing management in an era of climate change on Qinghai-Tibetan Plateau. Agric. Ecosyst. Environ. 2020, 287, 106684. [Google Scholar] [CrossRef]
- Simkin, S.M.; Allen, E.B.; Bowman, W.D.; Clark, C.M.; Belnap, J.; Brooks, M.L.; Cade, B.S.; Collins, S.L.; Geiser, L.H.; Gilliam, F.S.; et al. Conditional vulnerability of plant diversity to atmospheric nitrogen deposition across the United States. Proc. Natl. Acad. Sci. USA 2016, 113, 4086–4091. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lu, X.; Isbell, F.; Stevens, C.; Han, X.; He, N.; Zhang, G.; Yu, Q.; Huang, J.; Han, X. Rapid plant species loss at high rates and at low frequency of N addition in temperate steppe. Glob. Change Biol. 2014, 20, 3520–3529. [Google Scholar] [CrossRef]
- Li, Y.Y.; Dong, S.K.; Wen, L.; Wang, X.X.; Wu, Y. Soil carbon and nitrogen pools and their relationship to plant and soil dynamics of degraded and artificially restored grasslands of the Qinghai-Tibetan Plateau. Geoderma 2014, 213, 178–184. [Google Scholar] [CrossRef]
- Zhang, T.; Guo, R.; Gao, S.; Guo, J.X.; Sun, W. Response of plant community composition and productivity to warming and nitrogen deposition in a temperate meadow ecosystem. Biogeosciences 2015, 11, 6647–6672. [Google Scholar] [CrossRef]
- Foster, B.L.; Gross, K.L. Species Richness in a Successional Grassland: Effects of Nitrogen Enrichment and Plant Litter. Ecology 1998, 79, 2593–2602. [Google Scholar] [CrossRef]
- Clark, C.M.; Tilman, D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 2008, 451, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Lan, Z.; Bai, Y. Testing mechanisms of N-enrichment-induced species loss in a semiarid Inner Mongolia grassland: Critical thresholds and implications for long-term ecosystem responses. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3125–3134. [Google Scholar] [CrossRef] [Green Version]
- Southon, G.E.; Field, C.; Caporn, S.J.; Britton, A.J.; Power, S.A. Nitrogen deposition reduces plant diversity and alters ecosystem functioning: Field-scale evidence from a nationwide survey of UK heathlands. PLoS ONE 2013, 8, e59031. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, L.J.; Jones, L.; Sheppard, L.J.; Smart, S.M.; Bobbink, R.; Dise, N.B.; Ashmore, M.R. Evidence for differential effects of reduced and oxidised nitrogen deposition on vegetation independent of nitrogen load. Environ. Pollut. 2016, 208, 890–897. [Google Scholar] [CrossRef] [Green Version]
- Humbert, J.Y.; Dwyer, J.M.; Andrey, A.; Arlettaz, R. Impacts of nitrogen addition on plant biodiversity in mountain grasslands depend on dose, application duration and climate: A systematic review. Glob. Change Biol. 2016, 22, 110–120. [Google Scholar] [CrossRef]
- Hautier, Y.; Niklaus, P.A.; Hector, A. Competition for light causes plant biodiversity loss after eutrophication. Science 2009, 324, 636–638. [Google Scholar] [CrossRef] [Green Version]
- Suding, K.N.; Collins, S.L.; Gough, L.; Clark, C.; Cleland, E.E.; Gross, K.L.; Milchunas, D.G.; Pennings, S. Functional- and abundance-based mechanisms explain diversity loss due to N fertilization. Proc. Natl. Acad. Sci. USA 2005, 102, 4387–4392. [Google Scholar] [CrossRef] [Green Version]
- Dingaan, M.N.V.; Walker, S.; Tsubo, M.; Newby, T. Influence of Grazing on Plant Diversityproductivity Relationship in Semi-Arid Grassland of South Africa. Appl. Ecol. Environ. Res. 2016, 14, 1–13. [Google Scholar] [CrossRef]
- Huang, P.M.; Wang, S.-L.; Tzou, Y.-M.; Huang, Y.; Weng, B.; Zhuang, S.; Wang, M.K. Physicochemical and biological interfacial interactions: Impacts on soil ecosystem and biodiversity. Environ. Earth Sci. 2012, 68, 2199–2209. [Google Scholar] [CrossRef]
- Chen, S.; Wang, W.; Xu, W.; Wang, Y.; Wan, H.; Chen, D.; Tang, Z.; Tang, X.; Zhou, G.; Xie, Z.; et al. Plant diversity enhances productivity and soil carbon storage. Proc. Natl. Acad. Sci. USA 2018, 115, 4027–4032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fayiah, M.; Dong, S.K.; Li, Y.; Xu, Y.D.; Gao, X.X.; Li, S.; Shen, H.; Xiao, J.N.; Yang, Y.F.; Wessell, K. The relationships between plant diversity, plant cover, plant biomass and soil fertility vary with grassland type on Qinghai-Tibetan Plateau. Agric. Ecosyst. Environ. 2019, 286, 106659. [Google Scholar] [CrossRef]
- Li, S.; Dong, S.; Shen, H.; Han, Y.; Zhang, J.; Xu, Y.; Gao, X.; Yang, M.; Li, Y.; Zhao, Z.; et al. Different responses of multifaceted plant diversities of alpine meadow and alpine steppe to nitrogen addition gradients on Qinghai-Tibetan Plateau. Sci. Total Environ. 2019, 688, 1405–1412. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.K.; Gao, H.W.; Xu, G.C.; Hou, X.Y.; Long, R.J.; Kang, M.Y.; Lassoie, J.P. Farmer and professional attitudes to the large-scale ban on livestock grazing of grasslands in China. Environ. Conserv. 2007, 34, 246–254. [Google Scholar] [CrossRef]
- Kanianska, R. Agriculture and Its Impact on Land-Use, Environment, and Ecosystem Services. In Landscape Ecology-the Influences of Land Use and Anthropogenic Impacts of Landscape Creation; BoD–Books on Demand: Nordstedt, Germany, 2016. [Google Scholar]
- Lu, P.; Hao, T.; Li, X.; Wang, H.; Zhai, X.; Tian, Q.; Bai, W.; Stevens, C.; Zhang, W.H. Ambient nitrogen deposition drives plant-diversity decline by nitrogen accumulation in a closed grassland ecosystem. J. Appl. Ecol. 2021, 58, 1888–1898. [Google Scholar] [CrossRef]
- Hendriks, R.J.J.; Ozinga, W.A.; van den Berg, L.J.L.; Noordijk, E.; Schaminée, J.H.J.; van Groenendael, J.M. Aboveground persistence of vascular plants in relationship to the levels of airborne nutrient deposition. Plant Ecol. 2014, 215, 1277–1286. [Google Scholar] [CrossRef]
- Zong, N.; Shi, P.; Song, M.; Zhang, X.; Jiang, J.; Chai, X. Nitrogen Critical Loads for an Alpine Meadow Ecosystem on the Tibetan Plateau. Environ. Manag. 2016, 57, 531–542. [Google Scholar] [CrossRef]
- ThunjaI, T.; Boyd, C.E.; Dube, K. Pond Soil pH Measurement. J. World Aquac. Soc. 2001, 32, 141–152. [Google Scholar] [CrossRef]
- Li, Y.; Dong, S.; Liu, S.; Su, X.; Wang, X.; Zhang, Y.; Zhao, Z.; Gao, X.; Li, S.; Tang, L. Relationships between plant diversity and biomass production of alpine grasslands are dependent on the spatial scale and the dimension of biodiversity. Ecol. Eng. 2019, 127, 375–382. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Plant Diversity | ||||
|---|---|---|---|---|---|
| R | H | D | J | ||
| Soil | TN | −0.217 | −0.229 | −0.338 | −0.13 |
| TC | −0.471 * | −0.519* | −0.591 ** | −0.427 | |
| TP | −0.158 | −0.532* | −0.485 * | −0.678 ** | |
| TK | 0.066 | −0.017 | −0.017 | −0.089 | |
| NO3-N | −0.214 | −0.275 | −0.201 | −0.242 | |
| NH4-N | −0.573 * | −0.614 ** | −0.660 ** | −0.435 | |
| AP | 0.027 | −0.14 | −0.118 | −0.257 | |
| AK | 0.224 | 0.156 | 0.207 | 0.059 | |
| pH | 0.133 | −0.121 | −0.1 | −0.308 | |
| Ca | −0.425 | −0.33 | −0.462 | −0.04 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, R.; Shen, H.; Dong, S.; Li, S.; Xiao, J.; Zhi, Y.; Zhang, J.; Zuo, H.; Wu, S.; Mu, Z.; et al. Effects of 5-Year Nitrogen Addition on Species Composition and Diversity of an Alpine Steppe Plant Community on Qinghai-Tibetan Plateau. Plants 2022, 11, 966. https://doi.org/10.3390/plants11070966
Zhang R, Shen H, Dong S, Li S, Xiao J, Zhi Y, Zhang J, Zuo H, Wu S, Mu Z, et al. Effects of 5-Year Nitrogen Addition on Species Composition and Diversity of an Alpine Steppe Plant Community on Qinghai-Tibetan Plateau. Plants. 2022; 11(7):966. https://doi.org/10.3390/plants11070966
Chicago/Turabian StyleZhang, Ran, Hao Shen, Shikui Dong, Shuai Li, Jiannan Xiao, Yangliu Zhi, Jing Zhang, Hui Zuo, Shengnan Wu, Zhiyuan Mu, and et al. 2022. "Effects of 5-Year Nitrogen Addition on Species Composition and Diversity of an Alpine Steppe Plant Community on Qinghai-Tibetan Plateau" Plants 11, no. 7: 966. https://doi.org/10.3390/plants11070966
APA StyleZhang, R., Shen, H., Dong, S., Li, S., Xiao, J., Zhi, Y., Zhang, J., Zuo, H., Wu, S., Mu, Z., & Shi, H. (2022). Effects of 5-Year Nitrogen Addition on Species Composition and Diversity of an Alpine Steppe Plant Community on Qinghai-Tibetan Plateau. Plants, 11(7), 966. https://doi.org/10.3390/plants11070966