
Role for Selenium in Metabolic Homeostasis and Human Reproduction



Abstract
:1. Introduction
1.1. Elemental Selenium
1.2. Selenium and Selenoproteins
1.3. Significance of Different Forms of Selenium and Selenoproteins to Health
1.4. Sources of Selenium
1.4.1. Bread and Cereals
1.4.2. Meat, Fish and Eggs
1.4.3. Milk, Dairy Products and Beverages
1.4.4. Fruit and Vegetables
1.5. Nutritional Requirements of Selenium
2. Synthesis of Selenoproteins
2.1. Selenocysteine–tRNA [Ser]Sec Aminoacylation
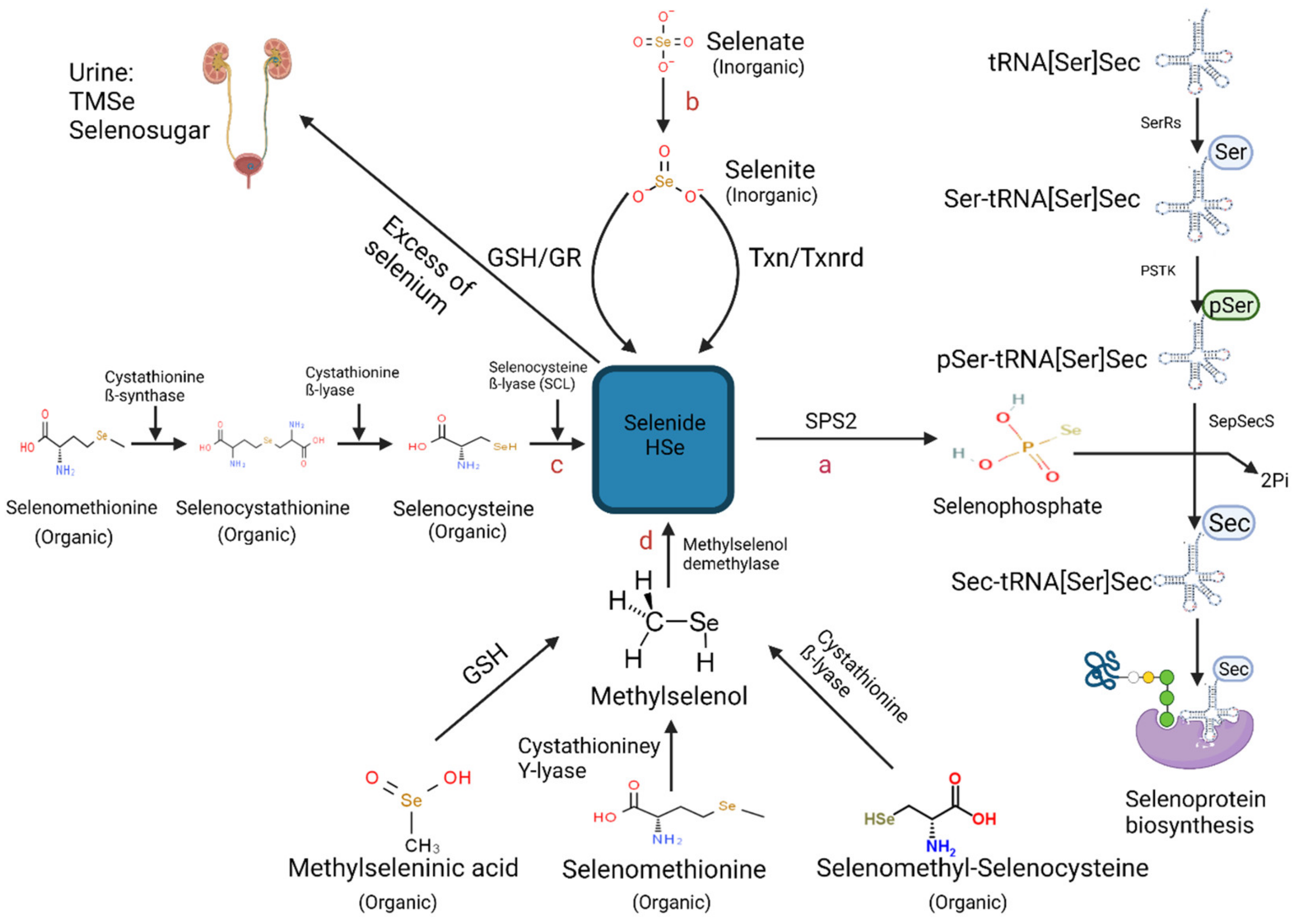
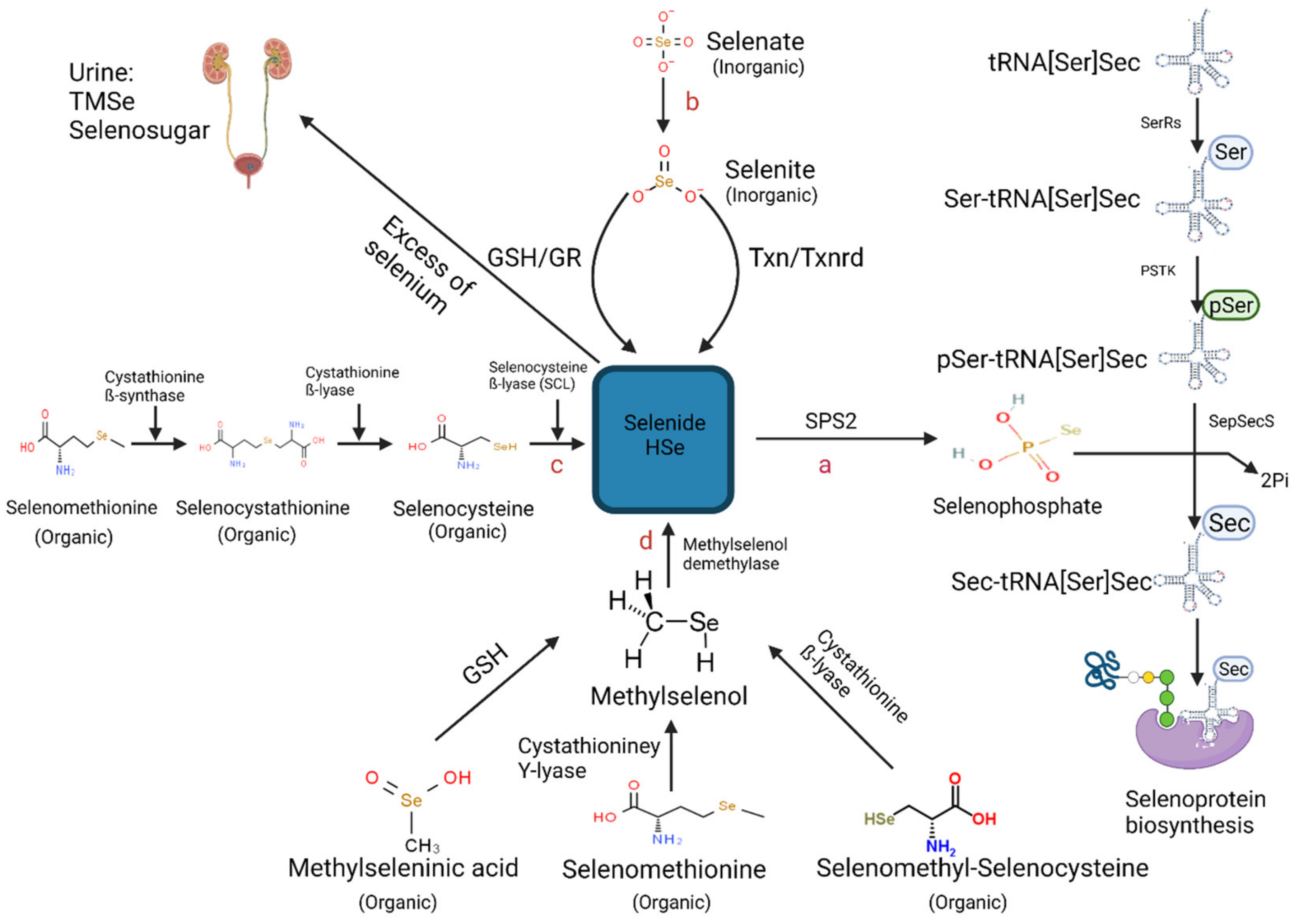
2.2. Hydrogen Selenide—The Core Precursor of Selenoprotein Synthesis
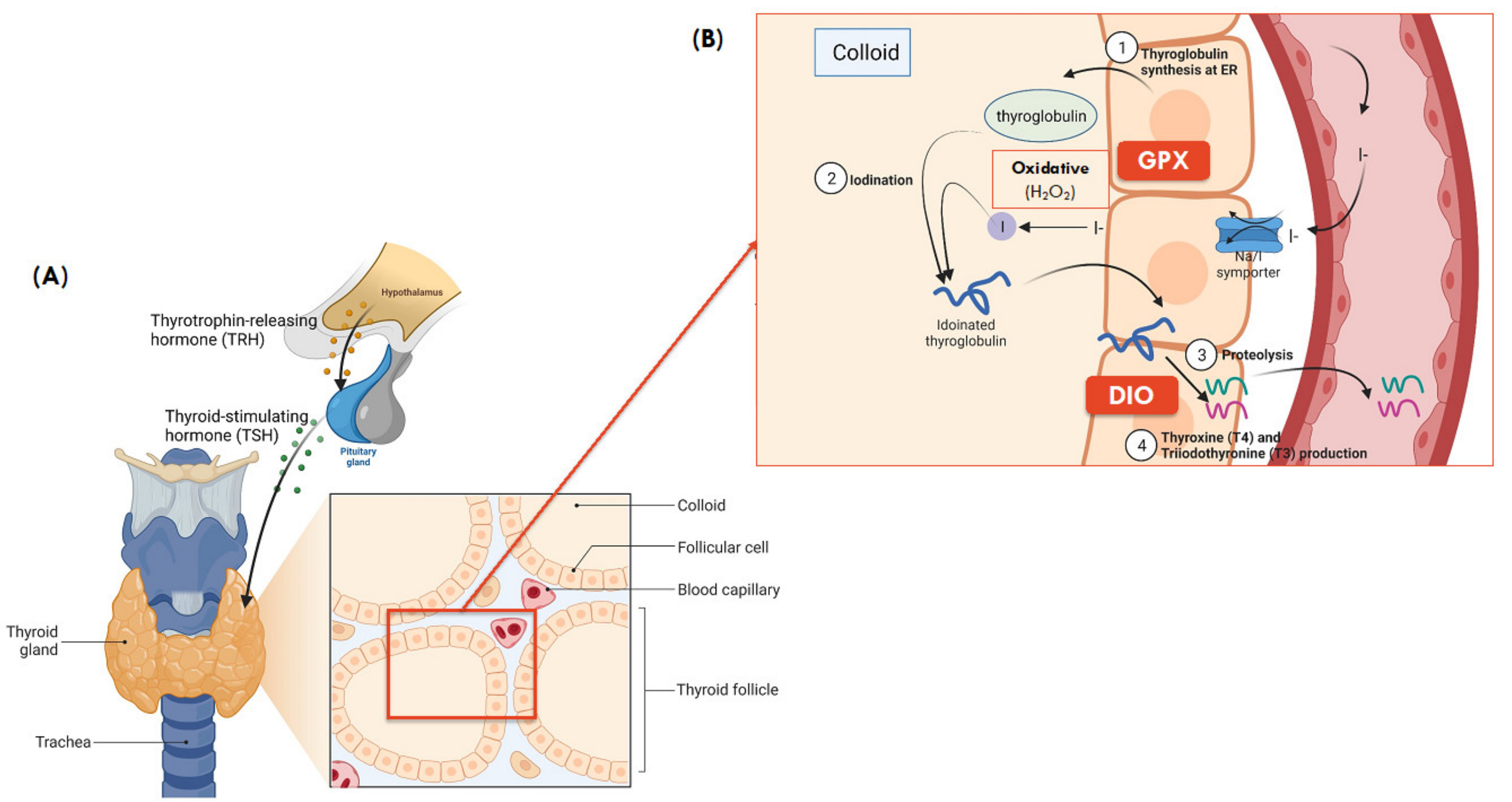
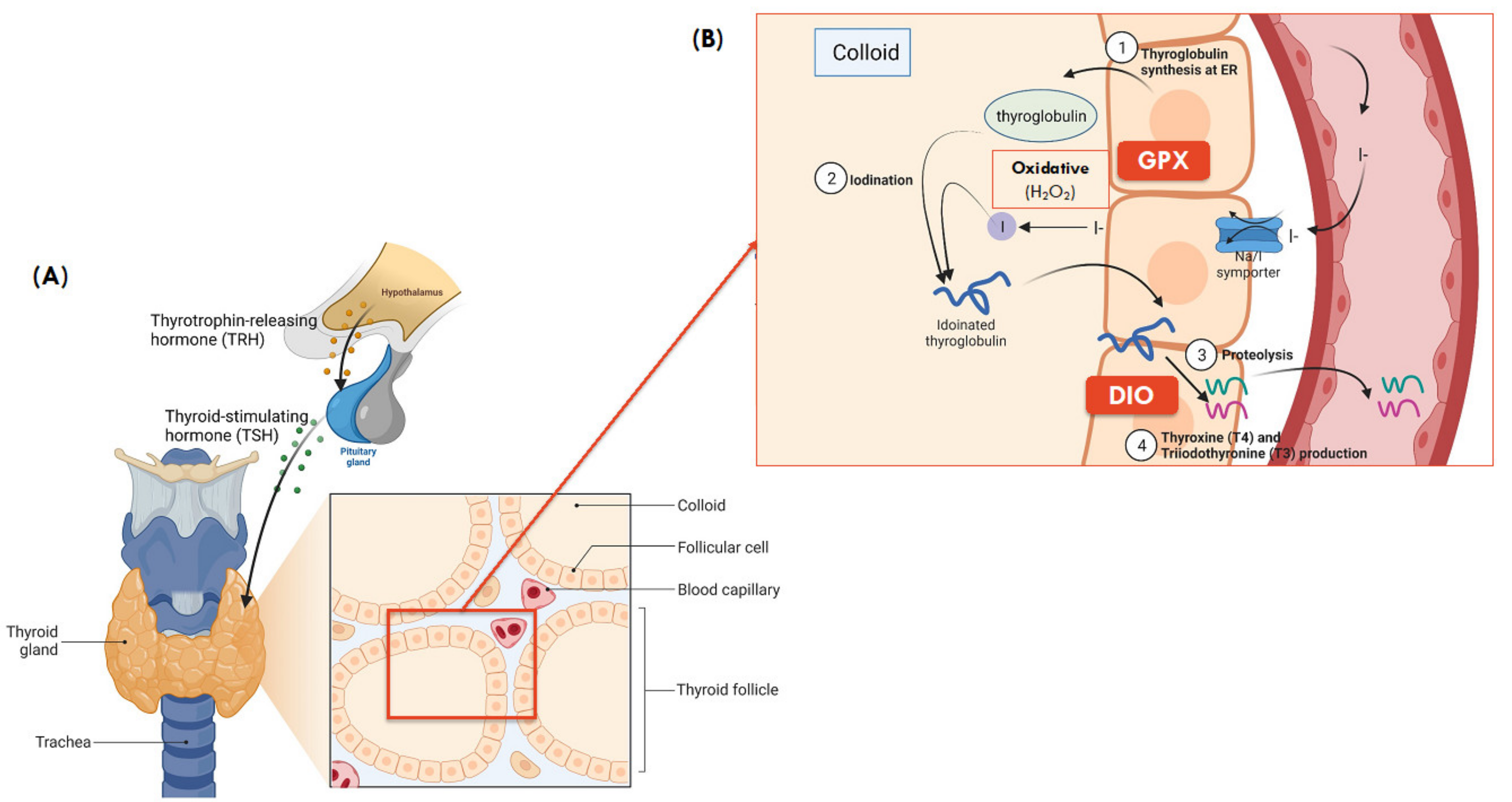
3. Selenium and Thyroid Function
3.1. Optimal Selenium Maintains Normal Thyroid Function
3.2. Selenium-Containing Selenoproteins Mediate Thyroid Function
3.3. Interplay between Selenoproteins and Thyroid Hormones Synthesis
3.4. Preferential Synthesis of Selenoprotein in the Thyroid
3.5. Thyroid and Metabolic Homeostasis
3.6. Controversies around the Application of Selenium Supplementation in Ameliorating Obesity
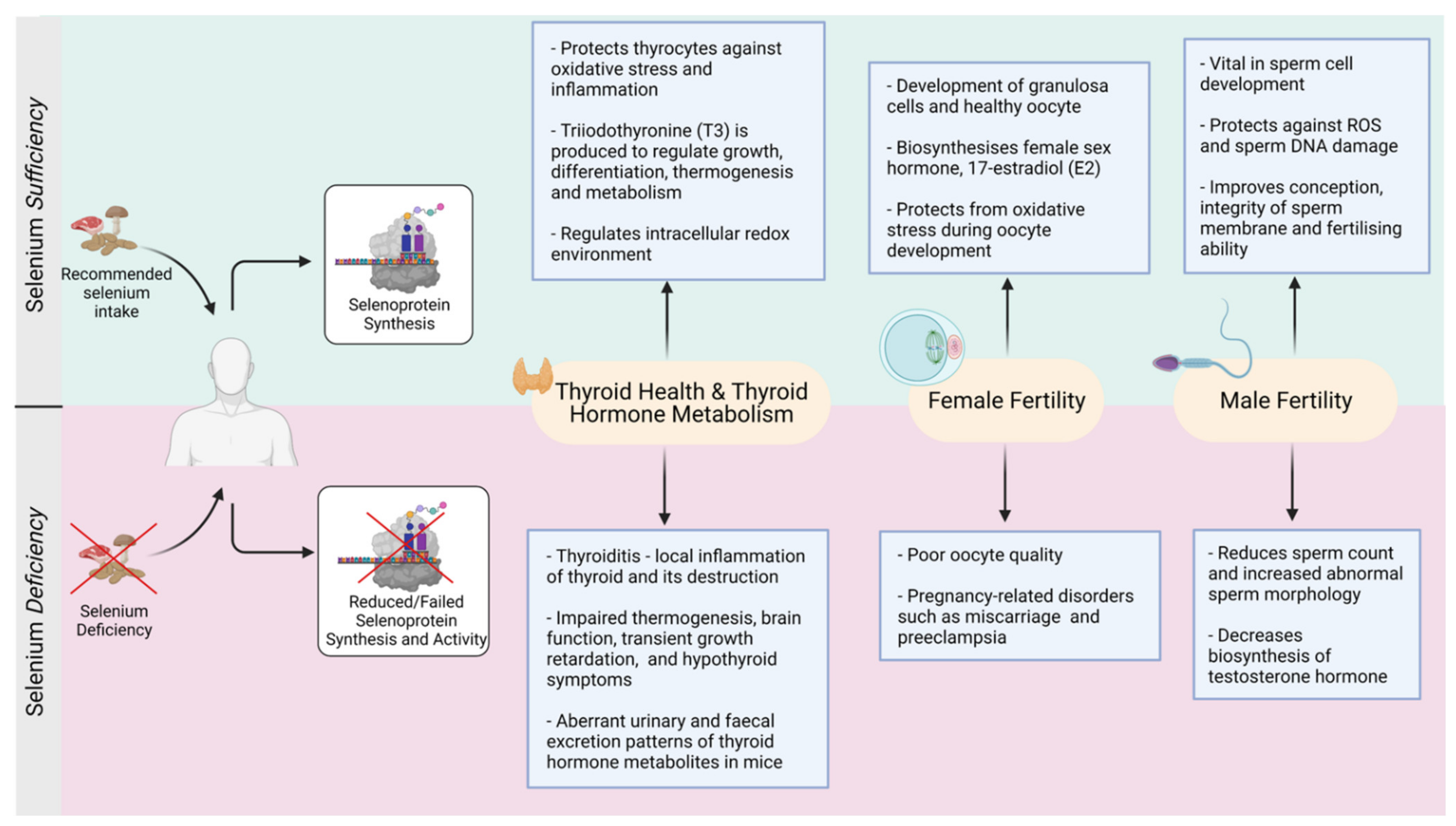
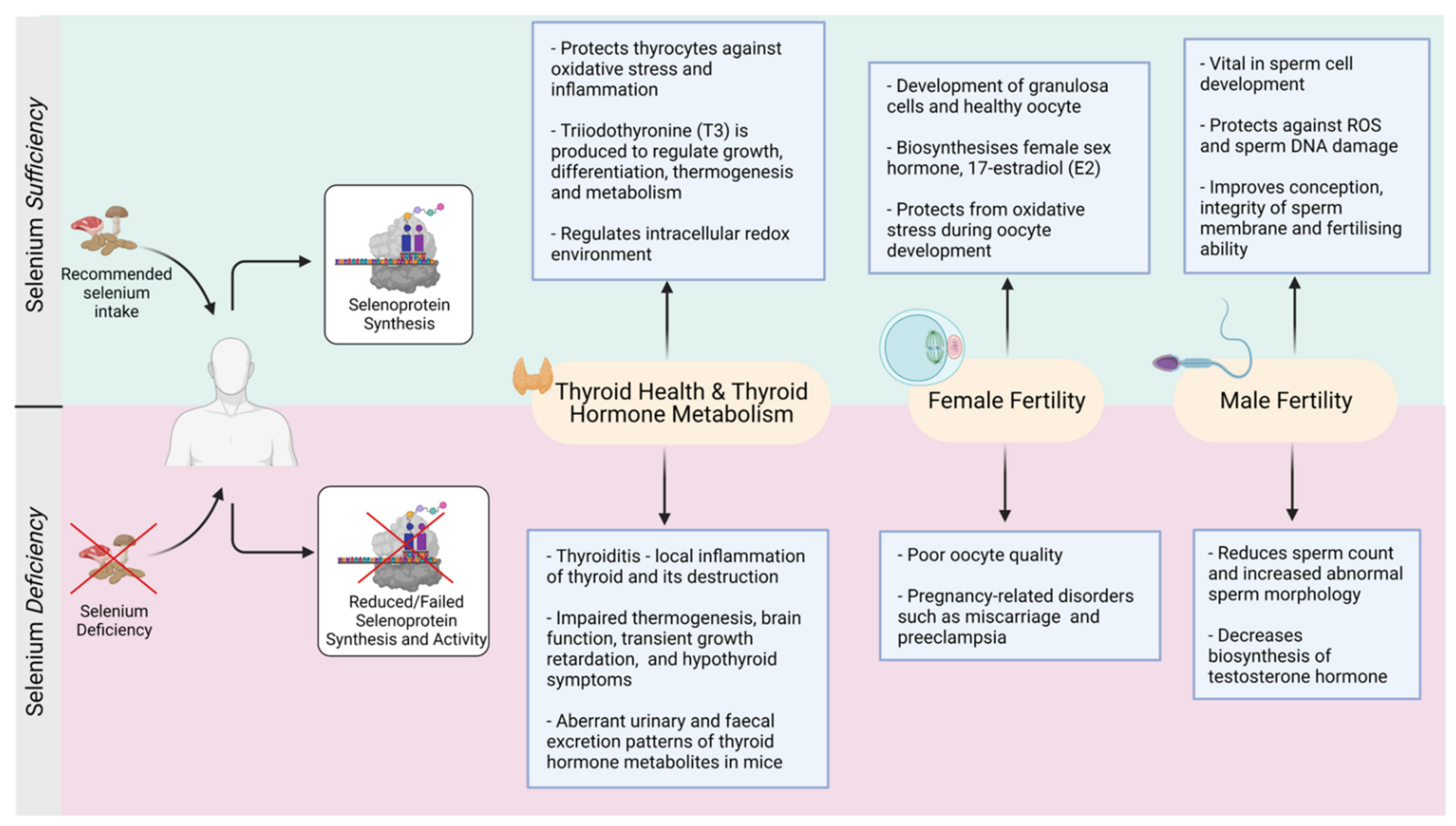
3.7. Thyroid Disorders Associated with Selenium Deficiency
3.8. Selenium Supplementation and Thyroid Disorders
4. Selenium-Thyroid and Female Reproduction
Selenium Supplementation and Female Reproduction
5. Selenium and Male Reproduction
5.1. Glutathione Peroxidase 4 (GPX4)
5.2. Selenoprotein P
5.3. GPX1 and GPX3
5.4. Selenium Supplementation and Male Fertility
6. Conclusions and Future Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bierla, K.; Dernovics, M.; Vacchina, V.; Szpunar, J.; Bertin, G.; Lobinski, R. Determination of selenocysteine and selenomethionine in edible animal tissues by 2D size-exclusion reversed-phase HPLC-ICP MS following carbamidomethylation and proteolytic extraction. Anal. Bioanal. Chem. 2008, 390, 1789–1798. [Google Scholar] [CrossRef] [PubMed]
- Boyd, R. Selenium stories. Nat. Chem. 2011, 3, 570. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, K.; Foltz, C.M. Selenium as an integral part of factor 3 against dietary necrotic liver degeneration. J. Am. Chem. Soc. 1957, 79, 3292–3293. [Google Scholar] [CrossRef]
- Doj, R.; Khanal, D.; Knight, A. Selenium: Its Role in Livestock Health and Productivity. J. Agric. Environ. 2010, 11, 101–106. [Google Scholar]
- Hoffmann, P.R.; Berry, M.J. The influence of selenium on immune responses. Mol. Nutr. Food Res. 2008, 52, 1273–1280. [Google Scholar] [CrossRef]
- Tinggi, U. Essentiality and toxicity of selenium and its status in Australia: A review. Toxicol Lett. 2003, 137, 103–110. [Google Scholar] [CrossRef]
- Bonnard, N.; Brondeau, M.-T.; Jargot, D.; Pillière, F.; Schneider, O.; Serre, P. Sélénium et Composés. 2011. Available online: http://www.inrs.fr/publications/bdd/fichetox/fiche.html?refINRS=FICHETOX_150 (accessed on 26 September 2020).
- Burk, R.F. Selenium in Biology and Human Health; Springer: New York, NY, USA, 1994. [Google Scholar]
- Simonoff, M.; Simonoff, G. Le sélénium et la vie; Masson: Paris, France, 1991. [Google Scholar]
- Ventura, M.; Melo, M.; Carrilho, F. Selenium and Thyroid Function. In Selenium; Michalke, B., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 157–173. [Google Scholar]
- Marinò, M.; Lanzolla, G.; Rotondo Dottore, G.; Marcocci, C. Chapter 32-Selenium usage and oxidative stress in Graves’ disease and Graves’ orbitopathy. In Pathology; Preedy, V.R., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 335–344. [Google Scholar]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigó, R.; Gladyshev, V.N. Characterization of Mammalian Selenoproteomes. Science 2003, 300, 1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brigelius-Flohé, R.; Flohé, L. Selenium and redox signaling. Arch. Biochem. Biophys. 2017, 617, 48–59. [Google Scholar] [CrossRef]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [Green Version]
- Letavayová, L.; Vlcková, V.; Brozmanová, J. Selenium: From cancer prevention to DNA damage. Toxicology 2006, 227, 1–14. [Google Scholar] [CrossRef]
- Pappas, A.C.; Zoidis, E.; Surai, P.F.; Zervas, G. Selenoproteins and maternal nutrition. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 151, 361–372. [Google Scholar] [CrossRef]
- Schrauzer, G.N. Selenomethionine: A review of its nutritional significance, metabolism and toxicity. J. Nutr. 2000, 130, 1653–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rayman, M.P.; Infante, H.G.; Sargent, M. Food-chain selenium and human health: Spotlight on speciation. Br. J. Nutr. 2008, 100, 238–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burk, R.F.; Hill, K.E. Regulation of Selenium Metabolism and Transport. Annu. Rev. Nutr. 2015, 35, 109–134. [Google Scholar] [CrossRef]
- Finley, J.W. Bioavailability of Selenium from Foods. Nutr. Rev. 2006, 64, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Fisinin, V.I. Selenium in pig nutrition and reproduction: Boars and semen quality-a review. Asian-Australas J. Anim. Sci. 2015, 28, 730–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surai, P.F.; Fisinin, V.I. Selenium in Livestock and Other Domestic Animals. In Selenium: Its Molecular Biology and Role in Human Health; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 595–606. [Google Scholar]
- Rayman, M.P. The use of high-selenium yeast to raise selenium status: How does it measure up? Br. J. Nutr. 2004, 92, 557–573. [Google Scholar] [CrossRef] [Green Version]
- Reich, H.J.; Hondal, R.J. Why Nature Chose Selenium. ACS Chem. Biol. 2016, 11, 821–841. [Google Scholar] [CrossRef]
- Mason, R.P.; Soerensen, A.L.; DiMento, B.P.; Balcom, P.H. The Global Marine Selenium Cycle: Insights From Measurements and Modeling. Glob. Biogeochem. Cycles 2018, 32, 1720–1737. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S. Current Knowledge on the Importance of Selenium in Food for Living Organisms: A Review. Molecules 2016, 21, 609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahid, M.; Niazi, N.K.; Khalid, S.; Murtaza, B.; Bibi, I.; Rashid, M.I. A critical review of selenium biogeochemical behavior in soil-plant system with an inference to human health. Environ. Pollut. 2018, 234, 915–934. [Google Scholar]
- Söderlund, M.; Virkanen, J.; Holgersson, S.; Lehto, J. Sorption and speciation of selenium in boreal forest soil. J. Environ. Radioact. 2016, 164, 220–231. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xiang, Y.; Zhou, Y.; Yang, Y.; Zhang, J.; Huang, H.; Shang, C.; Luo, L.; Gao, J.; Tang, L. Selenium contamination, consequences and remediation techniques in water and soils: A review. Environ. Res. 2018, 164, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Paikaray, S. Origin, Mobilization and Distribution of Selenium in a Soil/Water/Air System: A Global Perspective With Special Reference to the Indian Scenario. CLEAN–Soil Air Water 2016, 44, 474–487. [Google Scholar] [CrossRef]
- WHO. Guidelines for Drinking-Water Quality; World Health Organization: Geneva, Switzerland, 2004; Volume 1. [Google Scholar]
- Whanger, P.D. Selenocompounds in plants and animals and their biological significance. J. Am. Coll. Nutr. 2002, 21, 223–232. [Google Scholar] [CrossRef]
- Wolf, W.R.; Goldschmidt, R.J. Updated estimates of the selenomethionine content of NIST wheat reference materials by GC-IDMS. Anal. Bioanal. Chem. 2007, 387, 2449–2452. [Google Scholar] [CrossRef]
- Juniper, D.T.; Phipps, R.H.; Ramos-Morales, E.; Bertin, G. Effect of dietary supplementation with selenium-enriched yeast or sodium selenite on selenium tissue distribution and meat quality in beef cattle. J. Anim. Sci. 2008, 86, 3100–3109. [Google Scholar] [CrossRef]
- Fairweather-Tait, S.J.; Collings, R.; Hurst, R. Selenium bioavailability: Current knowledge and future research requirements. Am. J. Clin. Nutr. 2010, 91, 1484s–1491s. [Google Scholar] [CrossRef] [Green Version]
- Reyes, L.H.; Mar, J.L.; Rahman, G.M.; Seybert, B.; Fahrenholz, T.; Kingston, H.M. Simultaneous determination of arsenic and selenium species in fish tissues using microwave-assisted enzymatic extraction and ion chromatography-inductively coupled plasma mass spectrometry. Talanta 2009, 78, 983–990. [Google Scholar] [CrossRef]
- Cappon, C.J.; Smith, J.C. Chemical form and distribution of mercury and selenium in edible seafood. J. Anal. Toxicol. 1982, 6, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Lipiec, E.; Siara, G.; Bierla, K.; Ouerdane, L.; Szpunar, J. Determination of selenomethionine, selenocysteine, and inorganic selenium in eggs by HPLC-inductively coupled plasma mass spectrometry. Anal. Bioanal. Chem. 2010, 397, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Lubos, E.; Sinning, C.R.; Schnabel, R.B.; Wild, P.S.; Zeller, T.; Rupprecht, H.J.; Bickel, C.; Lackner, K.J.; Peetz, D.; Loscalzo, J.; et al. Serum selenium and prognosis in cardiovascular disease: Results from the AtheroGene study. Atherosclerosis 2010, 209, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Muñiz-Naveiro, O.; Domínguez-González, R.; Bermejo-Barrera, A.; Bermejo-Barrera, P.; Cocho, J.A.; Fraga, J.M. Selenium speciation in cow milk obtained after supplementation with different selenium forms to the cow feed using liquid chromatography coupled with hydride generation-atomic fluorescence spectrometry. Talanta 2007, 71, 1587–1593. [Google Scholar] [CrossRef]
- Kotrebai, M.; Birringer, M.; Tyson, J.F.; Block, E.; Uden, P.C. Selenium speciation in enriched and natural samples by HPLC-ICP-MS and HPLC-ESI-MS with perfluorinated carboxylic acid ion-pairing agents. Analyst 2000, 125, 71–78. [Google Scholar] [CrossRef]
- Baiomy, A.A.; Mohamed, A.E.A.; Mottelib, A.A. Effect of dietary selenium and vitamin E supplementation on productive and reproductive performance in rams. J. Vet. Med Res. 2009, 19, 39–43. [Google Scholar] [CrossRef] [Green Version]
- Ventura, M.; Melo, M.; Carrilho, F. Selenium and Thyroid Disease: From Pathophysiology to Treatment. Int. J. Endocrinol. 2017, 2017, 1297658. [Google Scholar] [CrossRef] [Green Version]
- Hosnedlová, B.; Kepinska, M.; Skalickova, S.; Fernández, C.; Ruttkay-Nedecky, B.; Malevu, T.; Sochor, J.; Baron, M.; Melčová, M.; Zidkova, J.; et al. A Summary of New Findings on the Biological Effects of Selenium in Selected Animal Species—A Critical Review. Int. J. Mol. Sci. 2017, 18, 2209. [Google Scholar] [CrossRef]
- Gereben, B.; Zavacki, A.; Ribich, S.; Kim, B.; Huang, S.; Simonides, W.; Anikó, Z.; Bianco, A. Cellular and Molecular Basis of Deiodinase-Regulated Thyroid Hormone Signaling 1. Endocr. Rev. 2008, 29, 898–938. [Google Scholar] [CrossRef] [Green Version]
- Winther, K.H.; Rayman, M.P.; Bonnema, S.J.; Hegedüs, L. Selenium in thyroid disorders—Essential knowledge for clinicians. Nat. Rev. Endocrinol. 2020, 16, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Carlson, B.A.; Lee, B.J.; Tsuji, P.A.; Tobe, R.; Park, J.M.; Schweizer, U.; Gladyshev, V.N.; Hatfield, D.L. Selenocysteine tRNA[Ser]Sec: From Nonsense Suppressor tRNA to the Quintessential Constituent in Selenoprotein Biosynthesis. In Selenium: Its Molecular Biology and Role in Human Health; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 3–12. [Google Scholar]
- Hatfield, D.L.; Gladyshev, V.N. How selenium has altered our understanding of the genetic code. Mol. Cell. Biol. 2002, 22, 3565–3576. [Google Scholar] [CrossRef] [Green Version]
- Turanov, A.A.; Everley, R.A.; Hybsier, S.; Renko, K.; Schomburg, L.; Gygi, S.P.; Hatfield, D.L.; Gladyshev, V.N. Regulation of Selenocysteine Content of Human Selenoprotein P by Dietary Selenium and Insertion of Cysteine in Place of Selenocysteine. PLoS ONE 2015, 10, e0140353. [Google Scholar] [CrossRef] [Green Version]
- Turanov, A.A.; Xu, X.M.; Carlson, B.A.; Yoo, M.H.; Gladyshev, V.N.; Hatfield, D.L. Biosynthesis of selenocysteine, the 21st amino acid in the genetic code, and a novel pathway for cysteine biosynthesis. Adv. Nutr. 2011, 2, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.M.; Turanov, A.A.; Carlson, B.A.; Yoo, M.H.; Everley, R.A.; Nandakumar, R.; Sorokina, I.; Gygi, S.P.; Gladyshev, V.N.; Hatfield, D.L. Targeted insertion of cysteine by decoding UGA codons with mammalian selenocysteine machinery. Proc. Natl. Acad. Sci. USA 2010, 107, 21430–21434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalke, B. Selenium; Springer International Publishing: Cham, Switzerland, 2018. [Google Scholar]
- Papp, L.V.; Lu, J.; Holmgren, A.; Khanna, K.K. From selenium to selenoproteins: Synthesis, identity, and their role in human health. Antioxid. Redox Signal. 2007, 9, 775–806. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Wichman, J.; Winther, K.H.; Bonnema, S.J.; Hegedüs, L. Selenium Supplementation Significantly Reduces Thyroid Autoantibody Levels in Patients with Chronic Autoimmune Thyroiditis: A Systematic Review and Meta-Analysis. Thyroid 2016, 26, 1681–1692. [Google Scholar] [CrossRef] [PubMed]
- Gärtner, R.; Gasnier, B.C.H.; Dietrich, J.W.; Krebs, B.; Angstwurm, M.W.A. Selenium Supplementation in Patients with Autoimmune Thyroiditis Decreases Thyroid Peroxidase Antibodies Concentrations. J. Clin. Endocrinol. Metab. 2002, 87, 1687–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Holmgren, A. Selenoproteins *. J. Biol. Chem. 2009, 284, 723–727. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.J.; Fiering, S.N.; Thai, B.; Wu, S.-Y.; Germain, E.S.; Parlow, A.F.; Germain, D.L.S.; Galton, V.A. Targeted Disruption of the Type 1 Selenodeiodinase Gene (Dio1) Results in Marked Changes in Thyroid Hormone Economy in Mice. Endocrinology 2006, 147, 580–589. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.L.; Wang, C.W.; Tan, S.R.; Liang, Y.; Yao, H.D.; Zhang, Z.W.; Xu, S.W. Selenium deficiency inhibits the conversion of thyroidal thyroxine (T4) to triiodothyronine (T3) in chicken thyroids. Biol. Trace Elem. Res. 2014, 161, 263–271. [Google Scholar] [CrossRef]
- Germain, D.L.S.; Hernandez, A.; Schneider, M.J.; Galton, V.A. Insights Into the Role of Deiodinases from Studies of Genetically Modified Animals. Thyroid 2005, 15, 905–916. [Google Scholar] [CrossRef]
- Rayman, M.P.; Duntas, L.H. Selenium Deficiency and Thyroid Disease. In The Thyroid and Its Diseases: A Comprehensive Guide for the Clinician; Luster, M., Duntas, L.H., Wartofsky, L., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 109–126. [Google Scholar]
- Contempre, B.; Le Moine, O.; Dumont, J.E.; Denef, J.F.; Many, M.C. Selenium deficiency and thyroid fibrosis. A key role for macrophages and transforming growth factor beta (TGF-beta). Mol. Cell. Endocrinol. 1996, 124, 7–15. [Google Scholar] [CrossRef]
- Brown, K.M.; Arthur, J.R. Selenium, selenoproteins and human health: A review. Public Health Nutr. 2001, 4, 593–599. [Google Scholar] [CrossRef] [Green Version]
- Esworthy, R.S.; Yang, L.; Frankel, P.H.; Chu, F.F. Epithelium-specific glutathione peroxidase, Gpx2, is involved in the prevention of intestinal inflammation in selenium-deficient mice. J. Nutr. 2005, 135, 740–745. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.C.; Kuo, W.R.; Chiang, F.Y.; Hsiao, P.J.; Lee, K.W.; Wu, C.W.; Juo, S.H. Glutathione peroxidase 3 gene polymorphisms and risk of differentiated thyroid cancer. Surgery 2009, 145, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Schomburg, L.; Köhrle, J. On the importance of selenium and iodine metabolism for thyroid hormone biosynthesis and human health. Mol. Nutr. Food Res. 2008, 52, 1235–1246. [Google Scholar] [CrossRef] [PubMed]
- Bermano, G.; Nicol, F.; Dyer, J.A.; Sunde, R.A.; Beckett, G.J.; Arthur, J.R.; Hesketh, J.E. Selenoprotein gene expression during selenium-repletion of selenium-deficient rats. Biol. Trace Elem. Res. 1996, 51, 211–223. [Google Scholar] [CrossRef]
- Yant, L.J.; Ran, Q.; Rao, L.; Van Remmen, H.; Shibatani, T.; Belter, J.G.; Motta, L.; Richardson, A.; Prolla, T.A. The selenoprotein GPX4 is essential for mouse development and protects from radiation and oxidative damage insults. Free Radic. Biol. Med. 2003, 34, 496–502. [Google Scholar] [CrossRef]
- Hill, K.E.; McCollum, G.W.; Boeglin, M.E.; Burk, R.F. Thioredoxin reductase activity is decreased by selenium deficiency. Biochem. Biophys. Res. Commun. 1997, 234, 293–295. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, N.; Fujii, T.; Fujii, J.; Taniguchi, N. Functional expression of rat thioredoxin reductase: Selenocysteine insertion sequence element is essential for the active enzyme. Biochem. J. 1999, 340 Pt 2, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Mullur, R.; Liu, Y.-Y.; Brent, G.A. Thyroid hormone regulation of metabolism. Physiol. Rev. 2014, 94, 355–382. [Google Scholar] [CrossRef] [Green Version]
- Miot, F.; Dupuy, C.; Dumont, J.; Rousset, B. Chapter 2 Thyroid Hormone Synthesis and Secretion. 2000. Available online: https://www.ncbi.nlm.nih.gov/books/NBK285550/ (accessed on 13 April 2021).
- Huang, Y.-H.; Tsai, M.-M.; Lin, K.-H. Thyroid hormone dependent regulation of target genes and their physiological significance. Chang. Gung Med. J. 2008, 31, 325–334. [Google Scholar]
- Arnér, E.S.J.; Holmgren, A. Physiological functions of thioredoxin and thioredoxin reductase. Eur. J. Biochem. 2000, 267, 6102–6109. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A.; Lu, J. Thioredoxin and thioredoxin reductase: Current research with special reference to human disease. Biochem. Biophys. Res. Commun. 2010, 396, 120–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.R.; Zhu, H.; Danelisen, I. Role of Peroxiredoxins in Protecting Against Cardiovascular and Related Disorders. Cardiovasc. Toxicol. 2020, 20, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Arriga, R.; Pacifici, F.; Capuani, B.; Coppola, A.; Orlandi, A.; Scioli, M.G.; Pastore, D.; Andreadi, A.; Sbraccia, P.; Tesauro, M.; et al. Peroxiredoxin 6 Is a Key Antioxidant Enzyme in Modulating the Link between Glycemic and Lipogenic Metabolism. Oxidative Med. Cell. Longev. 2019, 2019, 9685607. [Google Scholar] [CrossRef]
- Leoni, S.G.; Kimura, E.T.; Santisteban, P.; De la Vieja, A. Regulation of thyroid oxidative state by thioredoxin reductase has a crucial role in thyroid responses to iodide excess. Mol. Endocrinol. 2011, 25, 1924–1935. [Google Scholar] [CrossRef] [Green Version]
- Metere, A.; Frezzotti, F.; Graves, C.E.; Vergine, M.; De Luca, A.; Pietraforte, D.; Giacomelli, L. A possible role for selenoprotein glutathione peroxidase (GPx1) and thioredoxin reductases (TrxR1) in thyroid cancer: Our experience in thyroid surgery. Cancer Cell Int. 2018, 18, 7. [Google Scholar] [CrossRef]
- Schomburg, L.; Riese, C.; Michaelis, M.; Griebert, E.; Klein, M.O.; Sapin, R.; Schweizer, U.; Köhrle, J. Synthesis and Metabolism of Thyroid Hormones Is Preferentially Maintained in Selenium-Deficient Transgenic Mice. Endocrinology 2006, 147, 1306–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangiapane, E.; Pessione, A.; Pessione, E. Selenium and Selenoproteins: An Overview on Different Biological Systems. Curr. Protein Pept. Sci. 2014, 15, 598–607. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, W.C.; Keim, N.L. Dietary Selenium Intake Modulates Thyroid Hormone and Energy Metabolism in Men. J. Nutr. 2003, 133, 3443–3448. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, X.; Pedram, P.; Shahidi, M.; Du, J.; Yi, Y.; Gulliver, W.; Zhang, H.; Sun, G. Significant Beneficial Association of High Dietary Selenium Intake with Reduced Body Fat in the CODING Study. Nutrients 2016, 8, 24. [Google Scholar] [CrossRef]
- Hosseini, B.; Saedisomeolia, A.; Allman-Farinelli, M. Association Between Antioxidant Intake/Status and Obesity: A Systematic Review of Observational Studies. Biol. Trace Elem. Res. 2017, 175, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Banach, W.; Nitschke, K.; Krajewska, N.; Mongiałło, W.; Matuszak, O.; Muszyński, J.; Skrypnik, D. The Association between Excess Body Mass and Disturbances in Somatic Mineral Levels. Int. J. Mol. Sci. 2020, 21, 7306. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; Lin, R.; Nong, Q. Adiposity and Serum Selenium in U.S. Adults. Nutrients 2018, 10, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, C.; Xian, J.; Zeng, M.; Cai, Z.; Li, S.; Zhao, Y.; Shi, Z. Regional Difference in the Association between the Trajectory of Selenium Intake and Hypertension: A 20-Year Cohort Study. Nutrients 2021, 13, 1501. [Google Scholar] [CrossRef]
- Wang, X.-L.; Yang, T.-B.; Wei, J.; Lei, G.-H.; Zeng, C. Association between serum selenium level and type 2 diabetes mellitus: A non-linear dose–response meta-analysis of observational studies. Nutr. J. 2016, 15, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Xiao, J.; Liu, H.; Liu, H. Selenium nanoparticles alleviate hyperlipidemia and vascular injury in ApoE-deficient mice by regulating cholesterol metabolism and reducing oxidative stress†. Metallomics 2019, 12, 204–217. [Google Scholar] [CrossRef]
- Cavedon, E.; Manso, J.; Negro, I.; Censi, S.; Serra, R.; Busetto, L.; Vettor, R.; Plebani, M.; Pezzani, R.; Nacamulli, D.; et al. Selenium Supplementation, Body Mass Composition, and Leptin Levels in Patients with Obesity on a Balanced Mildly Hypocaloric Diet: A Pilot Study. Int. J. Endocrinol. 2020, 2020, 4802739. [Google Scholar] [CrossRef]
- Fontenelle, L.C.; Feitosa, M.M.; Freitas, T.E.C.; Severo, J.S.; Morais, J.B.S.; Henriques, G.S.; Oliveira, F.E.; Moita Neto, J.M.; do Nascimento Marreiro, D. Selenium status and its relationship with thyroid hormones in obese women. Clin. Nutr. ESPEN 2021, 41, 398–404. [Google Scholar] [CrossRef]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Tan, H.-Y.; Li, S.; Xu, Y.; Guo, W.; Feng, Y. Supplementation of Micronutrient Selenium in Metabolic Diseases: Its Role as an Antioxidant. Oxidative Med. Cell. Longev. 2017, 2017, 7478523. [Google Scholar] [CrossRef] [PubMed]
- Tinkov, A.A.; Ajsuvakova, O.P.; Filippini, T.; Zhou, J.-C.; Lei, X.G.; Gatiatulina, E.R.; Michalke, B.; Skalnaya, M.G.; Vinceti, M.; Aschner, M.; et al. Selenium and Selenoproteins in Adipose Tissue Physiology and Obesity. Biomolecules 2020, 10, 658. [Google Scholar] [CrossRef] [PubMed]
- Macek Jílková, Z.; Pavelka, S.; Flachs, P.; Hensler, M.; Kůs, V.; Kopecký, J. Modulation of type I iodothyronine 5’-deiodinase activity in white adipose tissue by nutrition: Possible involvement of leptin. Physiol. Res. 2010, 59, 561–569. [Google Scholar] [CrossRef]
- Brätter, P.; de Brätter, V.E.N. Influence of high dietary selenium intake on the thyroid hormone level in human serum. J. Trace Elem. Med. Biol. 1996, 10, 163–166. [Google Scholar] [CrossRef]
- Loomba, R.; Filippini, T.; Chawla, R.; Chaudhary, R.; Cilloni, S.; Datt, C.; Singh, S.; Dhillon, K.S.; Vinceti, M. Exposure to a high selenium environment in Punjab, India: Effects on blood chemistry. Sci. Total. Environ. 2020, 716, 135347. [Google Scholar] [CrossRef]
- Winther, K.H.; Bonnema, S.J.; Cold, F.; Debrabant, B.; Nybo, M.; Cold, S.; Hegedüs, L. Does selenium supplementation affect thyroid function? Results from a randomized, controlled, double-blinded trial in a Danish population. Eur. J. Endocrinol. 2015, 172, 657–667. [Google Scholar] [CrossRef]
- Combs, G.F., Jr.; Midthune, D.N.; Patterson, K.Y.; Canfield, W.K.; Hill, A.D.; Levander, O.A.; Taylor, P.R.; Moler, J.E.; Patterson, B.H. Effects of selenomethionine supplementation on selenium status and thyroid hormone concentrations in healthy adults. Am. J. Clin. Nutr. 2009, 89, 1808–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazi Tani, L.S.; Dennouni-Medjati, N.; Toubhans, B.; Charlet, L. Selenium Deficiency—From Soil to Thyroid Cancer. Appl. Sci. 2020, 10, 5368. [Google Scholar] [CrossRef]
- Szanto, I.; Pusztaszeri, M.; Mavromati, M. H2O2 Metabolism in Normal Thyroid Cells and in Thyroid Tumorigenesis: Focus on NADPH Oxidases. Antioxidants 2019, 8, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maia, A.L.; Goemann, I.M.; Meyer, E.L.; Wajner, S.M. Deiodinases: the balance of thyroid hormone: type 1 iodothyronine deiodinase in human physiology and disease. J. Endocrinol. 2011, 209, 283–297. [Google Scholar] [CrossRef] [Green Version]
- Fairweather-Tait, S.J.; Bao, Y.; Broadley, M.R.; Collings, R.; Ford, D.; Hesketh, J.E.; Hurst, R. Selenium in Human Health and Disease. Antioxid. Redox Signal. 2010, 14, 1337–1383. [Google Scholar] [CrossRef] [PubMed]
- Sturniolo, G.; Mesa, J. Selenium supplementation and autoimmune thyroid diseases. Endocrinol. Nutr. 2013, 60, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Gärtner, R.; Gasnier, B.C.H. Selenium in the treatment of autoimmune thyroiditis. BioFactors 2003, 19, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Lan, D.; Li, J.; Lin, Y.; Li, M. Selenium supplementation during in vitro maturation enhances meiosis and developmental capacity of yak oocytes. Anim. Sci. J. 2018, 89, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Hedaoo, M.K.; Khllare, K.P.; Meshram, M.D.; Sahatpure, S.K.; Patil, M.G. Study of some serum trace minerals in cyclic and non-cyclic surti buffaloes. Vet. World 2008, 1, 71–72. [Google Scholar]
- Foresta, C.; Flohé, L.; Garolla, A.; Roveri, A.; Ursini, F.; Maiorino, M. Male Fertility Is Linked to the Selenoprotein Phospholipid Hydroperoxide Glutathione Peroxidase1. Biol. Reprod. 2002, 67, 967–971. [Google Scholar] [CrossRef] [Green Version]
- Kommisrud, E.; Østerås, O.; Vatn, T. Blood Selenium Associated with Health and Fertility in Norwegian Dairy Herds. Acta Vet. Scand. 2005, 46, 229. [Google Scholar] [CrossRef] [Green Version]
- Basini, G.; Tamanini, C. Selenium stimulates estradiol production in bovine granulosa cells: Possible involvement of nitric oxide☆. Domest. Anim. Endocrinol. 2000, 18, 1–17. [Google Scholar] [CrossRef]
- Ceko, M.; Hummitzsch, K.; Hatzirodos, N.; Bonner, W.; Aitken, J.; Russell, D.; Lane, M.; Rodgers, R.; Harris, H. X-Ray fluorescence imaging and other analyses identify selenium and GPX1 as important in female reproductive function. Metallomics 2015, 7, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Grazul-Bilska, A.T.; Caton, J.S.; Arndt, W.; Burchill, K.; Thorson, C.; Borowczyk, E.; Bilski, J.J.; Redmer, D.A.; Reynolds, L.P.; Vonnahme, K.A. Cellular proliferation and vascularization in ovine fetal ovaries: Effects of undernutrition and selenium in maternal diet. Reproduction 2009, 137, 699. [Google Scholar] [CrossRef] [Green Version]
- Galton, V.A.; Martinez, E.; Hernandez, A.; Germain, E.A.S.; Bates, J.M.; Germain, D.L.S. The type 2 iodothyronine deiodinase is expressed in the rat uterus and induced during pregnancy. Endocrinology 2001, 142, 2123–2128. [Google Scholar] [CrossRef]
- Galton, V.A.; Martinez, E.; Hernandez, A.; Germain, E.A.S.; Bates, J.M.; Germain, D.L.S. Pregnant rat uterus expresses high levels of the type 3 iodothyronine deiodinase. J. Clin. Investig. 1999, 103, 979–987. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.A.; Dorfman, D.M.; Genest, D.R.; Salvatore, D.; Larsen, P.R. Type 3 iodothyronine deiodinase is highly expressed in the human uteroplacental unit and in fetal epithelium. J. Clin. Endocrinol. Metab. 2003, 88, 1384–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ejima, K.; Koji, T.; Nanri, H.; Kashimura, M.; Ikeda, M. Expression of thioredoxin and thioredoxin reductase in placentae of pregnant mice exposed to lipopolysaccharide. Placenta 1999, 20, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Ejima, K.; Nanri, H.; Toki, N.; Kashimura, M.; Ikeda, M. Localization of thioredoxin reductase and thioredoxin in normal human placenta and their protective effect against oxidative stress. Placenta 1999, 20, 95–101. [Google Scholar] [CrossRef]
- Duntas, L.H. Selenium and at-risk pregnancy: Challenges and controversies. Thyroid Res. 2020, 13, 16. [Google Scholar] [CrossRef] [PubMed]
- Hofstee, P.; Bartho, L.A.; McKeating, D.R.; Radenkovic, F.; McEnroe, G.; Fisher, J.J.; Holland, O.J.; Vanderlelie, J.J.; Perkins, A.V.; Cuffe, J.S.M. Maternal selenium deficiency during pregnancy in mice increases thyroid hormone concentrations, alters placental function and reduces fetal growth. J. Physiol. 2019, 597, 5597–5617. [Google Scholar] [CrossRef] [PubMed]
- Mesdaghinia, E.; Rahavi, A.; Bahmani, F.; Sharifi, N.; Asemi, Z. Clinical and Metabolic Response to Selenium Supplementation in Pregnant Women at Risk for Intrauterine Growth Restriction: Randomized, Double-Blind, Placebo-Controlled Trial. Biol. Trace Elem. Res. 2017, 178, 14–21. [Google Scholar] [CrossRef]
- Uhm, S.J.; Gupta, M.K.; Yang, J.H.; Lee, S.H.; Lee, H.T. Selenium improves the developmental ability and reduces the apoptosis in porcine parthenotes. Mol. Reprod. Dev. Inc. Gamete Res. 2007, 74, 1386–1394. [Google Scholar] [CrossRef]
- Shi, L.; Ren, Y.; Zhang, C.; Yue, W.; Lei, F. Effects of organic selenium (Se-enriched yeast) supplementation in gestation diet on antioxidant status, hormone profile and haemato-biochemical parameters in Taihang Black Goats. Anim. Feed. Sci. Technol. 2018, 238, 57–65. [Google Scholar] [CrossRef]
- Yang, H.; Qazi, I.H.; Pan, B.; Angel, C.; Guo, S.; Yang, J.; Zhang, Y.; Ming, Z.; Zeng, C.; Meng, Q.; et al. Dietary Selenium Supplementation Ameliorates Female Reproductive Efficiency in Aging Mice. Antioxidants 2019, 8, 634. [Google Scholar] [CrossRef] [Green Version]
- Riese, C.; Michaelis, M.; Mentrup, B.; Götz, F.; Köhrle, J.; Schweizer, U.; Schomburg, L. Selenium-dependent pre- and posttranscriptional mechanisms are responsible for sexual dimorphic expression of selenoproteins in murine tissues. Endocrinology 2006, 147, 5883–5892. [Google Scholar] [CrossRef] [Green Version]
- Beckett, G.J.; Arthur, J.R. Selenium and endocrine systems. J. Endocrinol. 2005, 184, 455–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knapen, M.F.C.M.; Zusterzeel, P.L.M.; Peters, W.H.M.; Steegers, E.A.P. Glutathione and glutathione-related enzymes in reproduction: A review. Eur. J. Obstet. Gynecol. Reprod. Biol. 1999, 82, 171–184. [Google Scholar] [CrossRef]
- Safarinejad, M.R.; Safarinejad, S. Efficacy of Selenium and/or N-Acetyl-Cysteine for Improving Semen Parameters in Infertile Men: A Double-Blind, Placebo Controlled, Randomized Study. J. Urol. 2009, 181, 741–751. [Google Scholar] [CrossRef]
- Ursini, F.; Heim, S.; Kiess, M.; Maiorino, M.; Roveri, A.; Wissing, J.; Flohé, L. Dual function of the selenoprotein PHGPx during sperm maturation. Science 1999, 285, 1393–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, H.; Hirao, F.; Sakamoto, T.; Sekine, K.; Mizukura, Y.; Saito, M.; Kitamoto, T.; Hayasaka, M.; Hanaoka, K.; Nakagawa, Y. Early embryonic lethality caused by targeted disruption of the mouse PHGPx gene. Biochem. Biophys. Res. Commun. 2003, 305, 278–286. [Google Scholar] [CrossRef]
- Parillo, F.; Sylla, L.; Palombi, C.; Monaci, M.; Stradaioli, G. Immunocytochemical Localisation of Phospholipid Hydroperoxide Glutathione Peroxidase in Bull’s Spermatogenic Cells. Ital. J. Anim. Sci. 2014, 13, 3483. [Google Scholar] [CrossRef]
- Olson, G.E.; Winfrey, V.P.; NagDas, S.K.; Hill, K.E.; Burk, R.F. Apolipoprotein E Receptor-2 (ApoER2) Mediates Selenium Uptake from Selenoprotein P by the Mouse Testis*. J. Biol. Chem. 2007, 282, 12290–12297. [Google Scholar] [CrossRef] [Green Version]
- Griswold, M.D. 50 years of spermatogenesis: Sertoli cells and their interactions with germ cells. Biol. Reprod. 2018, 99, 87–100. [Google Scholar] [CrossRef]
- Hill, K.E.; Zhou, J.; McMahan, W.J.; Motley, A.K.; Atkins, J.F.; Gesteland, R.F.; Burk, R.F. Deletion of Selenoprotein P Alters Distribution of Selenium in the Mouse*. J. Biol. Chem. 2003, 278, 13640–13646. [Google Scholar] [CrossRef] [Green Version]
- Noblanc, A.; Kocer, A.; Chabory, E.; Vernet, P.; Saez, F.; Cadet, R.; Conrad, M.; Drevet, J.R. Glutathione Peroxidases at Work on Epididymal Spermatozoa: An Example of the Dual Effect of Reactive Oxygen Species on Mammalian Male Fertilizing Ability. J. Androl. 2011, 32, 641–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahsan, U.; Kamran, Z.; Raza, I.; Ahmad, S.; Babar, W.; Riaz, M.H.; Iqbal, Z. Role of selenium in male reproduction—A review. Anim. Reprod. Sci. 2014, 146, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Irvine, D.S. Glutathione as a treatment for male infertility. Rev. Reprod. 1996, 1, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Asri-Rezaei, S.; Nourian, A.; Shalizar-Jalali, A.; Najafi, G.; Nazarizadeh, A.; Koohestani, M.; Karimi, A. Selenium supplementation in the form of selenium nanoparticles and selenite sodium improves mature male mice reproductive performances. Iran. J. Basic Med Sci. 2018, 21, 577–585. [Google Scholar]
- Mirone, M.; Giannetta, E.; Isidori, A. Selenium and reproductive function. A systematic review. J. Endocrinol. Invest. 2013, 36, 28–36. [Google Scholar] [PubMed]
- Agarwal, A.; Ahmad, G.; Sharma, R. Reference values of reactive oxygen species in seminal ejaculates using chemiluminescence assay. J. Assist. Reprod. Genet. 2015, 32, 1721–1729. [Google Scholar] [CrossRef] [Green Version]
- Kaushal, N.; Bansal, M.P. Diminished reproductive potential of male mice in response to selenium-induced oxidative stress: Involvement of HSP70, HSP70-2, and MSJ-1. J. Biochem. Mol. Toxicol. 2009, 23, 125–136. [Google Scholar] [CrossRef]
- Shalini, S.; Bansal, M.P. Role of selenium in spermatogenesis: Differential expression of cjun and cfos in tubular cells of mice testis. Mol. Cell. Biochem. 2006, 292, 27–38. [Google Scholar] [CrossRef]
- Shalini, S.; Bansal, M.P. Alterations in selenium status influences reproductive potential of male mice by modulation of transcription factor NFκB. BioMetals 2007, 20, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Bano, I.; Malhi, M.; Khatri, P.; Soomro, S.A.; Sajjad, H.; Leghari, A.; Awais, M.; Kandhro, S.; Lakho, S.A.; Soomro, M. Effect of dietary selenium yeast supplementation on morphology and antioxidant status in testes of young goat. Pak. J. Zool. 2019, 51, 979–988. [Google Scholar] [CrossRef]
- Stefanov, R.; Chervenkov, M.; Anev, G.; Maksimovic, N.; Andreeva, M.; Ivanova, T.; Milovanovic, A. Effect of supplementation with inorganic and organic selenium on sperm quality and quantity in north-east Bulgarian merino rams. Biotechnol. Anim. Husb. 2018, 34, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Martins, S.M.M.K.; Afonso, E.R.; Parazzi, L.J.; de Andrade, A.F.C.; Leal, D.F.; Gameiro, A.H.; de Sant’Anna Moretti, A.; de Arruda, R.P. Organic selenium supplementation is cost-effective for increasing the number of seminal doses produced by sexually mature boars. Rev. Bras. Zootec. 2018, 47. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Selenoproteins Relevant to Thyroid Health | Cellular Distribution Within Thyroid | Functions Under Optimal Conditions | Thyroid and Other Pathologies Associated with the Respective Selenoprotein Deficiency/Abnormality | |
|---|---|---|---|---|
| Iodothyronine deiodinase (DIO) | DIO1 | Plasma membrane | Converts inactive thyroxine (T4) to the biologically active triiodothyronine (T3) thyroid form, and control circulating T3 levels | DIO1 knockout mice had normal growth, development, and fertility, however they exhibited aberrant circulating TH and excretory iodothyronines [53,58]. Se deficiency in chickens inhibited the levels of DIO1, DIO2 and DIO3, which indirectly suppressed the conversion of T4 to T3 [59]. |
| DIO2 | ER membrane | Intracellular conversion of T4 to T3 in thyrocytes, contributing to the major source of T3 in circulation in euthyroidism | DIO2 knockout mice exhibited impaired thermogenesis, brain function, transient growth retardation, in addition to aberrant circulating TH [53,60]. | |
| DIO3 | Plasma membrane | Inactivates thyroid hormones from T4 to reverse T3 (rT3), and T3 to T2 | Fetal DIO3 knockout in mice is significant among all DIOs. It was characterised by reduced viability, growth retardation, impaired fertility, and hypothyroid symptoms, along with reduced T3 and increased T4 levels in the circulation [53] | |
| Glutathione peroxidase (GPX) | GPX1 | Cytosol | Protection against oxidative stress through reduction of H2O2 and lipid hydroperoxides | Reduced Se-dependent GPX activity, leading to diffusion of H2O2 into the thyroid parenchyma, followed by local inflammation and ultimately its destruction [61,62] |
| GPX2 | Not expressed in thyroid | Expression is specific to epithelial cells including colonic crypts and that in gastrointestinal tract and lungs, which offers protection against ingested lipid hydroperoxides [63] | N/A to thyroid health. However, GPX2 was suggested to be the most important Se-dependent antioxidant in colon and early defense against colon cancer [63]. Although no negative impacts of Se deficiency have been illustrated in humans, GPX2 knockout mice when deprived in Se showed enhanced ileocolitis (inflammation) in the intestinal mucosa than that of Se sufficiency [64]. | |
| GPX3 | Thyroid colloid (apical side of thyrocyte membrane) | Antioxidant in extracellular fluid; thyroid protection from excessive H2O2 in thyrocytes and follicular lumen, which is not used by thyroid peroxidase (TPO) for the iodination of thyroglobulin during the process of thyroid hormone synthesis | Se deficiencies reduced Se-dependent GPX activity, leading to diffusion of H2O2 into the thyroid parenchyma, followed by local inflammation and ultimately its destruction [61,62] In a Taiwanese population, SNPs in GPX3 were significantly associated with the risk of differentiated thyroid cancer [65] | |
| GPX4 | Mitochondria | Reduction of phospholipid hydroperoxides in mitochondria, offering protection of mitochondrial membrane in thyrocytes against peroxidation [61,66] Modulate apoptosis | Limited information on the effects of GPX abnormalities on thyroid function. In Se-deficient mice, GPX4 activity was not significantly reduced when compared to that of GPX1 that had a 50% reduction [67]. Interestingly, embryonic mice with a full GPX4-gene deletion was fatal [68]. | |
| Thioredoxin reductase (TXNRD) | TXNRD1 | Cytosol | Antioxidant by catalysing NADPH-dependent reduction of thioredoxin, regulating the intracellular redox environment | Limited data on the biological effects of TXNRD abnormalities on thyroid function. However, Se deficiency in mice caused a 4.5% and 11% reduction in TXNRD activity in liver and kidney respectively, while TXNRD level was unchanged [69]. It was postulated that the lack of Sec interferes with the biosynthesis of TXNRD and incorporation into the polypeptide chain at termination, which ultimately produces a non-functional protein [70]. |
| TXNRD2 | Mitochondria | Regulate cell proliferation | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mojadadi, A.; Au, A.; Salah, W.; Witting, P.; Ahmad, G. Role for Selenium in Metabolic Homeostasis and Human Reproduction. Nutrients 2021, 13, 3256. https://doi.org/10.3390/nu13093256
Mojadadi A, Au A, Salah W, Witting P, Ahmad G. Role for Selenium in Metabolic Homeostasis and Human Reproduction. Nutrients. 2021; 13(9):3256. https://doi.org/10.3390/nu13093256
Chicago/Turabian StyleMojadadi, Albaraa, Alice Au, Wed Salah, Paul Witting, and Gulfam Ahmad. 2021. "Role for Selenium in Metabolic Homeostasis and Human Reproduction" Nutrients 13, no. 9: 3256. https://doi.org/10.3390/nu13093256
APA StyleMojadadi, A., Au, A., Salah, W., Witting, P., & Ahmad, G. (2021). Role for Selenium in Metabolic Homeostasis and Human Reproduction. Nutrients, 13(9), 3256. https://doi.org/10.3390/nu13093256